Abstract
With the anticipated population growth by 2050, the demand for high-quality protein for human consumption is set to rise. To enhance the sustainability of U.S. cattle production, producers and researchers have traditionally concentrated on improving the feed efficiency of steers through advancements in genetics, nutrition, and microbiome tools, resulting in a more marketable beef product. However, without successful pregnancies, there would be no marketable animals to feed. Despite extensive research on hormonal impacts on cattle physiology, including nutrition and reproduction, there is limited knowledge about how the rumen microbial environment is impacted by pregnancy and feed efficiency in female beef cattle. Understanding the rumen microbiome’s role in feed efficiency and its response to hormonal changes during pregnancy is crucial for advancing sustainable beef cattle production. Therefore, this review highlights the importance of understanding the microbial dynamics in the rumen of pregnant beef females. Although progress has been made, gaps remain in understanding how varying nutritional requirements throughout pregnancy affect the rumen microbiome, highlighting the need for continued research. Addressing these areas will lead to more efficient and sustainable cattle production practices, benefiting beef production and contributing to global food security.
1. Introduction
As the world’s population is expected to reach 9.5 billion by the year 2050, the demand for high-quality protein sources is expected to increase [1]. Subsequently, this comes at the cost of diminishing essential resources such as land, feed ingredients, grain commodities, and labor [2]. A shift toward efficient production practices is warranted, including techniques to increase crop and carcass yields, implement environmentally conscious management practices, and maintain feed-efficient beef cattle. These key strategies will contribute to improving the sustainability of food production, ultimately benefiting U.S. agricultural practices.
Feed efficiency in ruminant animals considers their ability to convert inputs into profitable outputs. In beef cattle, these inputs consist of low-quality feed options for humans, which are then converted into high-quality end products like milk and meat. Importantly, this conversion process relies on the rumen microbiome, consisting of bacteria, protozoa, fungi, and archaea, and its fermentative capacity, supplying approximately 70% of the ruminant’s energy requirement [3,4]. Research has suggested a variety of relationships between host feed efficiency status and the rumen microbiome populations [5,6]. In addition to feed conversion and processing, the rumen microbiome has also been linked to host health, disease state [7], and methane production [8]. The composition and structure of the rumen microbiome are influenced by a variety of different factors, including age [9], diet [10], environment [11], and host physiology [12,13].
Different production sectors within the beef cattle industry have a unique definition of what is considered a quality end product, as different management practices are implemented to ensure the operation meets production goals. Feedlots are primarily focused on increasing red muscle tissue and fat deposition. Highly concentrated diets are commonly offered as an efficient way to meet the animal’s nutritional requirements at a certain point in time [14]. In cow–calf operations, producers are more focused on female health and adequate body condition throughout the year, which encourages proper fetal growth and health during pregnancy, positively impacting the female’s capacity to raise a healthy calf [15]. In this setting, producers often rely on forage-based diets that are fed on an inclining plane of nutrition with the intent to develop future generations of efficient cattle.
An animal that is feed efficient in one production system may not necessarily be feed efficient in another. Shi et al. [16] found that shifting growing heifer diets to include a greater percentage of grain allowed heifers to achieve a greater average daily gain and feed conversion ratio. When cattle diets contain a greater proportion of grain compared to forage, they typically are expected to more efficiently convert the energy from the feed into muscle mass [17]. Grains are typically highly digestible and spend less time undergoing ruminal processing and fermentation, whereas forages take longer to undergo the fermentation process once in the rumen [18]. However, roughages, or physically effective fiber, should be included in cattle diets for at least 5–10% of the total dry matter intake to encourage the rumination process and promote ruminal health [19,20]. Ultimately, understanding feed efficiency in the context of production type should aid in the progression of economics and outputs.
Historically, there has been a heavy focus on studying steer rumen microbial environment and feed efficiency through stages in production [5,6,21]. These animals are managed to promote growth and muscle development. Previous studies have utilized novel microbial technologies and varying management strategies, including a variety of feed rations, ionophores, and growth-promoting implants, to determine their impact on rumen microbiome health, rumen microbiome structure, and animal performance [21,22]. However, few studies have aimed to determine the interplay of the heifer and cow rumen microbiome and feed efficiency status, especially during pregnancy [23].
A productive female is the most important asset to a profitable cow–calf operation [24]. Pregnancy in beef females is one of the most important periods to producers, where it is optimal to have females that can produce healthy and efficient calves and maintain a stable rumen microbial environment throughout all stages of production. Although there is a higher expectation of profit and value in a market animal, without a successful pregnancy, there would be no animal to market. Ideally, females are a long-term constituent of the herd and are a consistent contributor of genetic attributes for the next generation of cattle, and it is important to consider the rumen microbial environment to improve the cow–calf enterprise. During pregnancy, the female undergoes a demanding physiological stage, and the rumen microbiome during this stage is essential to produce and provide energy for herself and the fetus [25]. Additionally, studies suggested that the rumen microbiome is a moderately heritable trait, and when the goal of production is to maintain feed-efficient animals, understanding feed efficiency status and the rumen microbial environment of females within the herd is necessary to preserve beneficial genetics [26,27]. Therefore, this review underscores the critical importance of understanding the physiological changes and microbial dynamics in the rumen during pregnancy in beef females. By examining the rumen environment and its influence on feed efficiency during this demanding stage, researchers and producers can develop strategies to optimize the health and productivity of the herd, ensuring the long-term sustainability and profitability of cow–calf enterprises. These insights are critical for advancing knowledge and improving practices in the beef industry.
2. Feed Efficiency, Diet, and the Rumen Microbiome of Fed Cattle Systems
The selection of feed-efficient cattle has been a production focus for the past several decades [28]. Roughly 70–80% of costs associated with beef cattle production are nutrition-related [29]. With the rising costs in feed commodities in current years [30], it is imperative for cattle to become more efficient in feed utilization to ensure profitability and the sustainability of production practices.
Feed efficiency can be calculated in multiple ways. Feed conversion ratios have been utilized previously to calculate feed efficiency and rely on weight gain and growth to determine efficiency status. A larger number represents better efficiency and conversion rates when using a Gain:Feed intake ratio (G:F) [28]. These methods are geared toward younger, growing animals and do not consider energy requirements for other later stages of production. The residual feed intake (RFI) is a common feed efficiency measure utilized by producers and researchers. RFI is defined as the difference between the actual dry matter intake (DMI) of an animal and the expected DMI required for maintenance and growth, estimated through a regression equation involving metabolic BW and average daily gain (ADG) [28]. RFI considers the energy requirements for the current stage of production to determine nutrient requirements. An animal with a low RFI requires less feed than expected to meet production threshold standards, resulting in a lower input cost. An animal with a high RFI consumes more feed than expected to meet production thresholds, resulting in a higher input cost or loss in profit from poor performance [31]. Feed efficiency research has recently increased focus on the rumen microbiome due to its fermentative capacity and impact on the nutritional status of the animal [32,33]. Several studies have noted differences in rumen microbiome structure in animals differing in feed efficiency status across multiple species of ruminants [34]. In steers, differences in the relative abundances of higher orders of taxa, low-abundance bacteria, alpha diversity, and beta diversity of the microbiome have been identified in greater feed-efficient animals. Zhang et al. [35] demonstrated that low-RFI steers tended to have greater eukaryotic richness in the rumen when compared to high-RFI steers. Additionally, in the same study, a PCoA failed to demonstrate significant beta diversity differences in the rumen microbial communities of the two groups of steers [35]. Zhang et al. [35] also concluded that the relative abundance of two eukaryotic taxa (kingdom Fungi and genus Entodinium) were affected by steers divergent in feed efficiency status. These studies are valuable resources for identifying key microbial factors of feed efficiency in feedlot production systems.
The relationship between rumen microbes and various ruminant species is complex, and understanding this association may be crucial for optimizing feed efficiency and animal health. However, the relationship between rumen microbes and RFI across various ruminant species is a dynamic and species-specific interaction influenced by microbial composition, host genetics, and diet [27]. Indeed, the microbial community composition of the rumen and camelid foregut has been demonstrated to be highly variable among diet, host species, and geography when analyzed from 742 samples from 32 animal species across 35 countries [10]. However, the authors noted that 67.1% of all bacterial reads detected in all samples were assigned to Prevotella, Butyrivibrio, Ruminococcus, and unclassified Lachnospiraceae, Ruminococcaceae, Bacteroidales, and Clostridiales, suggesting that there is a core rumen microbiome among ruminants. Yet, they were not equally abundant across species, indicating although these communities may be important for ruminal function, host species often host microbial populations that are better adapted to maximizing species-specific nutrient use efficiency.
Feed efficiency is a moderately heritable trait [36]. Importantly, Sasson et al. [37] determined that there is a core group of heritable rumen microbes. Furthermore, there were other production traits that were able to be independently associated with heritable microbes, such as DMI, energy-harvesting efficiency, and milk protein. In sheep, Ellison et al. [38] identified a group of microbes that were found to be heritable, including Methanobrevibacter smithii and Mitsuokella jalaludini, that differed in abundance in high- and low-feed-efficiency groups. In dairy cattle, Shabat et al. [39] identified microbes that differed in high- and low-feed-efficiency groups, as well as differences in abundance and richness within those phenotypic groups. Defining these heritable microbes in the context of cattle production is important because they can enhance feed efficiency and overall herd productivity, offering a pathway to improve genetic selection and sustainable farming practices.
Diets vary among producers, geographical locations, and production operations [40]. Diet is also one of the greater factors impacting the rumen microbiome and its environment (Figure 1) [41]. The rumen incubation period of different forages, as described by Elliot and others [42], impacts the temporal stability of the rumen, as well as the timeline of rumen colonization events. Fernando et al. [43] demonstrated, when examining eight ruminally cannulated beef steers, a significant shift of rumen microbial populations in favor of amylolytic and other starch-digesting bacteria during a transition from a forage-based diet to a concentrate diet. Appropriately, this is in response to the increase in highly fermentable substrates in the high-concentrate diet. At the phylum level, an increase in Bacteroidetes populations and a decrease in Firmicutes and Proteobacteria populations are a common effect of transitioning cattle from forage-based diets to high-concentrate diets. Typically, these rapid shifts in bacterial populations also occur when cattle are transitioned into a finishing stage (e.g., feedlot operation); therefore, these operations typically implement a meticulous process to adapt cattle to high-concentrate diets [44,45]. The rapid transition of cattle diets can also induce subacute ruminal acidosis (SARA), where the rumen pH decreases and impacts the ability of the rumen to function normally. This can include damaging the lining of the gastrointestinal tract, limiting the absorption of volatile fatty acids, the formation of liver abscesses, and ultimately inhibiting optimal animal performance [46]. When more readily fermentable carbohydrates, as found in concentrate diets, are rapidly increased, the rumen enters this acidotic state, which hinders bacterial fibrolytic activity and promotes an ideal environment for amylolytic and lactic acid-utilizing bacteria [47,48,49,50]. Ogunade et al. [22] demonstrated that several commensal bacterial species, such as Fibrobacter succinogenes and two Ruminococcus species, were reduced with the onset of SARA in feedlot steers, indicating a reduction in diversity with the onset of acidotic changes.
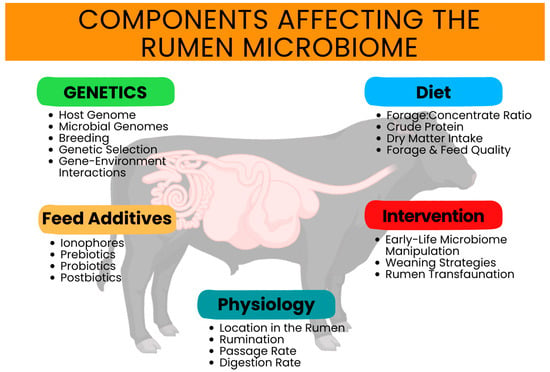
Figure 1.
Components affecting the rumen microbiome.
Microbiome and rumen chemistry changes due to SARA can also impact ruminal tissue, further exacerbating the deleterious effects of a rapid dietary transition. For example, when cattle were transitioned from forage diets (90.6% dry matter, 114 g of crude protein/kg dry matter, 600 g of neutral detergent fiber/kg dry matter, 173 g of non-fiber carbohydrate/kg dry matter, 69 g of starch/kg dry matter; 1.91 mCal/kg of dry matter) to grain diets (35% chopped hay and 65% mixed grain; 88.9% dry matter, 117 g of crude protein/kg dry matter, 307 g of neutral detergent fiber/kg dry matter, 504 g of non-fiber carbohydrate/kg dry matter, 409 g of starch/kg dry matter; 2.60 mCal/kg of dry matter), the rumen epithelium was compromised due to the reduction in cell adhesion in stratified squamous epithelial layers and sloughing of the stratum corneum [51]. These findings are common effects as a result of dietary impacts on the rumen microbiome commonly associated with the fed cattle system. While cow–calf enterprises differ from cattle-feeding operations in terms of diet and management practices, this research is valuable for highlighting these differences and improving the management of calves produced in productive cow–calf operations.
3. Pregnancy Effects on Gut Microbes
Within the past decade, the majority of beef cattle nutritional research has focused on feed efficiency, methane mitigation, and their rumen microbiome associations in steers [5,6]. Few studies have focused on the rumen microbiome of breeding females that consume roughly 70% of all feed supply across sectors of the beef industry [52]. The few studies that utilize female cattle tend to occur within the dairy industry [53,54]. In beef production, cow–calf operations are more commonly based around pasture or forage-based nutritional programs [55]. This sector of the beef industry focuses on meeting the nutritional needs of the animal to enhance reproductive performance and, in turn, profitability [56].
Successful reproductive performance in the cow–calf sector results in a healthy pregnancy and live, marketable calf. Bovine prenatal development was first outlined by Winters et al. [57] and Swett et al. [58] and has more recently been studied by Lyne et al. [59], Ferrell et al. [60], and Prior and Laster [61], with a particular point of interest focusing on the physiological changes throughout pregnancy, especially during the third trimester. This period is the most critical, as the fetus undergoes the largest growth transformation and final organ development [62]. With the increase in fetal growth and development, the female’s nutritional requirements are expected to also increase. On average, it has been reported that pregnant cows should receive 20% or more dietary energy and protein to aid in fetal growth, maintain proper body condition going into calving, and prepare for lactation, which is the most demanding stage of production [63]. Ensuring maternal nutritional requirements are met will offset potential negative effects, such as malformation and other deficiencies in the offspring [64,65,66]. Weller et al. [65] suggested that maternal overnutrition may be detrimental to offspring’s future reproductive capabilities. Their research examined 62 multiparous crossbred cows and observed a reduction in the number of follicles in heifers and a disruption of testicular development in young bulls. Wu et al. [66] demonstrated that a restriction of feed in the first trimester of gestation inhibited muscle and adipose tissue development in the offspring. Campos et al. [64] further acknowledged the advantages of meeting maternal nutritional needs for skeletal muscle tissue, adipose fat, and connective tissue development during the fetal stage when examining zebu and crossbred cattle, further defining the importance of nutrition during this physiologically demanding stage of production.
The gut microbiome is expected to acclimate in response to the imposed change in nutrient demands and dietary nutrient profile provided during pregnancy [67]. Differences in rumen microbiome during the prepartum and post-partum interval in dairy cattle have been identified, specifically as an indicator of production traits such as milk production [68]. Lima et al. [68] demonstrated differences in the relative abundance of amylolytic and cellulolytic bacteria, as well as a shift of fungi and protozoal abundances between the prepartum and postpartum period in dairy cattle, which were highly influential in milk production outputs. However, these changes were also likely influenced by dietary differences between the two groups, as prepartum cows were provided a diet high in fiber content (neutral detergent fiber [NDF] = 38.2%; amylase-treated NDF [aNDF] = 43.3%) and low in energy density (1.39 Mcal/kg), and postpartum cows received a diet lower in fiber (NDF = 24.1%; aNDF = 30.1%) but higher in energy density (1.69 Mcal/kg), achieved through increased starch and fat supplementation [68]. Martin et al. [23], however, demonstrated diet-independent differences in ruminal bacterial communities prior to and throughout pregnancy in Angus heifers [23]. The abundance of several metabolites and bacterial communities in Angus heifers fluctuated throughout pregnancy, likely in response to nutrient demands and physiological changes. Additionally, these metabolic and microbial shifts were linked to the heifers’ previously established feed efficiency status [23]. These changes, when correlated with feed efficiency, could help identify key microbial and metabolic markers that influence cattle productivity and efficiency. This knowledge could lead to more tailored nutritional strategies to optimize health, growth, and feed utilization in pregnant cattle, ultimately improving overall herd management and sustainability in beef production.
Beyond the limited work in dairy cattle, the effect of pregnancy on the gut microbiome of women and other vertebrate species has been reported [69,70,71]. Koren et al. [70] discovered that the gut microbiota of 256 pregnant women during the first trimester of pregnancy was similar to a healthy, non-lactating, non-pregnant woman; however, there was a shift of phylogenetic composition and microbial function similar to disease-associated dysbiosis over the course of pregnancy. However, the mechanism underlying this response failed to be identified. In the same study, it was found that by the third trimester, there was a noticeable increase in Proteobacteria and Actinobacteria. Women in this study often had symptoms of gut inflammation, which was further supported by Mukhopadhya et al. [72], who noted similar abundances tied to gut inflammation when populations exceeded a defined threshold of commensal proteobacterial colonization. While the composition and functions of the human gut microbiome in response to physiological changes during pregnancy have been well described, there is still a gap in understanding the established and anticipated changes in the rumen microbiome of beef cattle during pregnancy.
4. Pregnancy Effects on Rumen Microbiome and Metabolic Status in Cattle
Metabolic health and feed conversion remain a consistent goal, especially regarding reproduction in the cow–calf sector. Studies from nearly the last fifty years have highly supported the relevance of hormones and metabolic products in cattle productivity and performance [73,74,75,76,77,78,79,80,81,82]. In terms of reproduction, metabolic products are a driving factor stimulating ovulation, embryonic development, ovarian structures, steroid production, and estrus length in beef cattle [83].
The cow’s demand for glucose and other nutrients is expected to increase to support fetal growth and lactation after calving [84]. Unfortunately, there is often a decrease in appetite, impacting feed intake, potentially in relation to fetal size or estrogen secretion [85]. Cattle become more reliant upon the rumen microbes to effectively metabolize feedstuffs into usable nutrients to support maternal and reproductive function [86]. Beever [87] noted that increasing dietary protein and energy during the late term to counteract the anticipated reduction in intake is beneficial up to calving. After calving, the altered diet can lead to negative effects such as increased body condition score, reduced fertility, and diminished production outputs.
Additional research highly supports the importance of meeting nutritional requirements during gestation. When carbohydrate intake is not sufficient to meet the demands of production, metabolic diseases such as ketosis, tetany, and milk fever are likely to be detrimental to the animal’s production [74]. During periods of nutrient restriction, cows experience metabolic changes characterized by decreased blood glucose and insulin secretion. As a result, adipose tissues are mobilized, leading to increased ketogenesis and the production of ketone bodies, commonly referred to as ketosis [88,89]. When female cattle do not receive adequate nutrition to support a new pregnancy, it has been shown that internal signaling involving the somatotrophic axis (GH, IGF1, insulin, and IGFBP2) and leptin can impair fertility. This occurs because the cow’s metabolic state is insufficient to sustain pregnancy [90]. Nevertheless, metabolic outputs are crucial at every stage of production, particularly for successful reproductive activity and healthy pregnancy.
The onset of pregnancy triggers a range of hormonal fluctuations. The precise impact of pregnancy-related hormones on the composition of beef cattle rumen microbiomes remains to be clarified. Beyond ruminants, Mulak et al. [91] suggested that pregnancy hormones, such as progesterone and estrogen, impact bacterial metabolism, growth, and virulence of pathogens in the gastrointestinal tract. Another study demonstrated that host sex hormone levels and diet impact the gut microbiome in 341 female and 348 male mice [92]. Additionally, host gut microbiota could assist in regulating and modulating steroid sex hormone levels [93].
Several studies to date have explored the association between hormone levels and the host microbiome, including both the rumen and reproductive tract, providing insight into potential strategies to impact rumen and reproductive microbiome health (Figure 2). Henniger et al. [94] utilized 50 crossbred steers on either a moderate or aggressive growth hormone implant strategy containing estradiol and trenbolone acetate. They failed to find many significant differences in the rumen bacterial composition of steers differing in hormonal supplementation [94]. This was believed to be due to the absence of sex hormone receptors in the rumen epithelium. Further, when examining the effects of protein supplementation on the rumen and uterine microbiome of 60 commercial Angus pre- and post-pubertal heifers, the researchers failed to find significant variation between the two maturity groups [95]. This further supports the lack of a relationship between hormone fluctuations and rumen microbiome response. Unlike the rumen, the uterus has hormone receptors and is therefore more sensitive to hormonal fluctuations; however, the variation in the microbiome was minimal. Interestingly, another study indicated that eight of the most abundant OTUs present in the vaginal microbiome were also found in greater abundance in the ruminant gastrointestinal tract and fecal matter [96]. Ruminococcaceae, a highly abundant bacterial family in the rumen, has also been found to be the most abundant in the female reproductive tract [97]. It is believed that the presence of microbes in both physiological tracts is partly due to the location of the vagina, which may allow microbes to enter from the digestive tract and other areas of the body [96]. Laguardia-Nascimento et al. [96] concluded that there was no evidence of hormonal differences associated with pregnant or non-pregnant cattle throughout puberty or pregnancy influencing the microbiome of the vaginal tract. However, it was noted that more research should be performed to clarify and validate their findings on other physiological changes and processes. Undoubtedly, pregnancy induces a range of physiological changes, and the host response—particularly concerning nutrition, metabolism, and reproduction—in cattle has been extensively documented. However, the limited understanding of how the rumen environment responds to pregnancy highlights the need for further research to determine its impact on beef cattle.
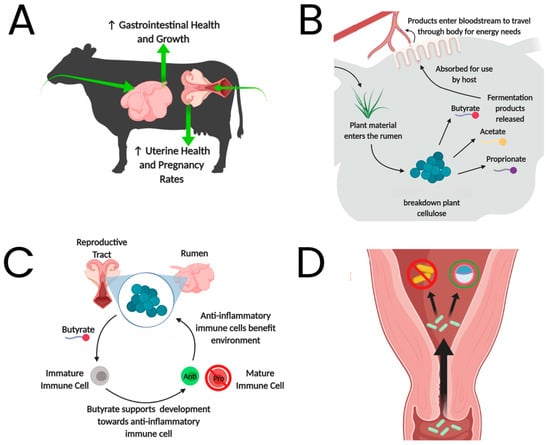
Figure 2.
Potential routes for applied microbiology to impact ruminal and reproductive microbiome health and stability. (A) Microbial and microbiome interventions to improve gut and reproductive health; (B) Example of ruminal microbial activity to provide energy to the host; (C) Example of how ruminal energy precursors, such as butyrate, may impact the host systemically; and (D) Microbial species or probiotics administered into the vagina to eliminate pathogens and benefit potential pregnancies by improving the uterine environment. Adapted from Myer et al., 2020 [98,99].
5. Next Generation and Future Work
As demand for sustainable practices in agriculture increases, there is growing emphasis on the development of microbial tools to maintain feed efficiency in beef cattle operations. The rumen microbiome composition of a beef female is critical for host health [100], productivity, and feed efficiency [5,6]. Identifying the role of the rumen microbiome and its connection with feed efficiency in beef females across various life stages could enhance opportunities for producing the next generation of feed-efficient cattle. Research examining the relationship between the rumen microbiome during pregnancy and feed efficiency in beef heifers could offer valuable insights to enhance the economic and environmental sustainability of beef cattle production in the United States.
Author Contributions
Conceptualization, M.G.M., P.R.M., K.J.M. and B.H.V.; investigation, M.G.M.; resources, P.R.M.; writing—original draft preparation, M.G.M. and J.F.C.-L.; writing—review and editing, M.G.M., P.R.M., J.F.C.-L., B.H.V. and K.J.M.; supervision, P.R.M.; funding acquisition, M.G.M. and P.R.M. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the Tennessee Beef Promotion Board and the USDA-NIFA Hatch/Multistate Project W-3010-TEN00578: Integrated Approach to Enhance Efficiency of Feed Utilization in Beef Production Systems; Accession Number: 1025826. The funding bodies had no role in the design of the study, collection, analyses, interpretation of data, or writing the manuscript.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- World Population Prospects-Population Division-United Nations. Available online: https://population.un.org/wpp/ (accessed on 30 August 2024).
- Henchion, M.; Hayes, M.; Mullen, A.M.; Fenelon, M.; Tiwari, B. Future Protein Supply and Demand: Strategies and Factors Influencing a Sustainable Equilibrium. Foods 2017, 6, 53. [Google Scholar] [CrossRef] [PubMed]
- Flint, H.J.; Bayer, E.A. Plant Cell Wall Breakdown by Anaerobic Microorganisms from the Mammalian Digestive Tract. Ann. N. Y. Acad. Sci. 2008, 1125, 280–288. [Google Scholar] [CrossRef] [PubMed]
- Seymour, W.M.; Campbell, D.R.; Johnson, Z.B. Relationships between rumen volatile fatty acid concentrations and milk production in dairy cows: A literature study. Anim. Feed. Sci. Technol. 2005, 119, 155–169. [Google Scholar] [CrossRef]
- Li, F.; Hitch, T.C.A.; Chen, Y.; Creevey, C.J.; Guan, L.L. Comparative metagenomic and metatranscriptomic analyses reveal the breed effect on the rumen microbiome and its associations with feed efficiency in beef cattle. Microbiome 2019, 7, 6. [Google Scholar] [CrossRef]
- Myer, P.R.; Smith, T.P.L.; Wells, J.E.; Kuehn, L.A.; Freetly, H.C. Rumen Microbiome from Steers Differing in Feed Efficiency. PLoS ONE 2015, 10, e0129174. [Google Scholar] [CrossRef]
- McCann, J.C.; Luan, S.; Cardoso, F.C.; Derakhshani, H.; Khafipour, E.; Loor, J.J. Induction of subacute ruminal acidosis affects the ruminal microbiome and epithelium. Front. Microbiol. 2016, 7, 701. [Google Scholar] [CrossRef] [PubMed]
- Wallace, R.J.; Rooke, J.A.; McKain, N.; Duthie, C.-A.; Hyslop, J.J.; Ross, D.W.; Waterhouse, A.; Watson, M.; Roehe, R. The rumen microbial metagenome associated with high methane production in cattle. BMC Genom. 2015, 16, 839. [Google Scholar] [CrossRef]
- Yáñez-Ruiz, D.R.; Macías, B.; Pinloche, E.; Newbold, C.J. The persistence of bacterial and methanogenic archaeal communities residing in the rumen of young lambs. FEMS Microbiol. Ecol. 2010, 72, 272–278. [Google Scholar] [CrossRef]
- Henderson, G.; Cox, F.; Ganesh, S.; Jonker, A.; Young, W.; Global Rumen Census Collaborators; Abecia, L.; Angarita, E.; Aravena, P.; Nora Arenas, G.; et al. Rumen microbial community composition varies with diet and host, but a core microbiome is found across a wide geographical range. Sci. Rep. 2015, 5, 14567. [Google Scholar] [CrossRef]
- Mizrahi, I.; Wallace, R.J.; Moraïs, S. The rumen microbiome: Balancing food security and environmental impacts. Nat. Rev. Microbiol. 2021, 19, 553–566. [Google Scholar] [CrossRef]
- Weimer, P.J. Redundancy, resilience, and host specificity of the ruminal microbiota: Implications for engineering improved ruminal fermentations. Front. Microbiol. 2015, 6, 296. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Peng, Y.-J.; Chen, Y.; Klinger, C.M.; Oba, M.; Liu, J.-X.; Guan, L.L. Assessment of microbiome changes after rumen transfaunation: Implications on improving feed efficiency in beef cattle. Microbiome 2018, 6, 62. [Google Scholar] [CrossRef] [PubMed]
- Loerch, S.C. Effects of feeding growing cattle high-concentrate diets at a restricted intake on feedlot performance. J. Anim. Sci. 1990, 68, 3086–3095. [Google Scholar] [CrossRef] [PubMed]
- Bertelsen, M.; Vaarst, M. Shaping cow-calf contact systems: Farmers’ motivations and considerations behind a range of different cow-calf contact systems. J. Dairy Sci. 2023, 106, 7769–7785. [Google Scholar] [CrossRef]
- Shi, H.; Zhang, J.; Li, S.; Ji, S.; Cao, Z.; Zhang, H.; Wang, Y. Effects of a wide range of dietary forage-to-concentrate ratios on nutrient utilization and hepatic transcriptional profiles in limit-fed Holstein heifers. BMC Genom. 2018, 19, 148. [Google Scholar] [CrossRef]
- Dixon, R.M.; Stockdale, C.R. Associative effects between forages and grains: Consequences for feed utilisation. Aust. J. Agric. Res. 1999, 50, 757–774. [Google Scholar] [CrossRef]
- Owens, F.N.; Basalan, M. Ruminal Fermentation. In Rumenology; Millen, D.D., De Beni Arrigoni, M., Lauritano Pacheco, R.D., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 63–102. ISBN 978-3-319-30533-2. [Google Scholar]
- Allen, M.S. Relationship Between Fermentation Acid Production in the Rumen and the Requirement for Physically Effective Fiber. J. Dairy Sci. 1997, 80, 1447–1462. [Google Scholar] [CrossRef]
- National Academies of Sciences, Engineering, and Medicine; Division on Earth and Life Studies; Board on Agriculture and Natural Resources; Committee on Nutrient Requirements of Beef Cattle. Nutrient Requirements of Beef Cattle: Eighth Revised Edition; National Academies Press: Washington, DC, USA, 2016; ISBN 978-0-309-31702-3. [Google Scholar]
- Auffret, M.D.; Stewart, R.D.; Dewhurst, R.J.; Duthie, C.-A.; Watson, M.; Roehe, R. Identification of Microbial Genetic Capacities and Potential Mechanisms within the Rumen Microbiome Explaining Differences in Beef Cattle Feed Efficiency. Front. Microbiol. 2020, 11, 1229. [Google Scholar] [CrossRef]
- Ogunade, I.; Pech-Cervantes, A.; Schweickart, H. Metatranscriptomic Analysis of Sub-Acute Ruminal Acidosis in Beef Cattle. Animals 2019, 9, 232. [Google Scholar] [CrossRef]
- Martin, M.G.; McLean, K.J.; Voy, B.H.; Myer, P.R. Pregnancy Influences on The Rumen Environment of Angus Heifers Differing in Feed Efficiency. Master’s Thesis, University of Tennessee, Knoxville, TN, USA, 2023. Available online: https://trace.tennessee.edu/utk_gradthes/9961 (accessed on 23 September 2024).
- Cabrera, V.E. A simple formulation and solution to the replacement problem: A practical tool to assess the economic cow value, the value of a new pregnancy, and the cost of a pregnancy loss. J. Dairy Sci. 2012, 95, 4683–4698. [Google Scholar] [CrossRef]
- Pohler, K.G.; Franco, G.A.; Reese, S.T.; Smith, M.F. Chapter 3—Physiology and pregnancy of beef cattle. In Animal Agriculture; Bazer, F.W., Lamb, G.C., Wu, G., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 37–55. ISBN 978-0-12-817052-6. [Google Scholar]
- Jin, S.; Zhang, Z.; Zhang, G.; He, B.; Qin, Y.; Yang, B.; Yu, Z.; Wang, J. Maternal Rumen Bacteriota Shapes the Offspring Rumen Bacteriota, Affecting the Development of Young Ruminants. Microbiol. Spectr. 2023, 11, e03590-22. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Li, C.; Chen, Y.; Liu, J.; Zhang, C.; Irving, B.; Fitzsimmons, C.; Plastow, G.; Guan, L.L. Host genetics influence the rumen microbiota and heritable rumen microbial features associate with feed efficiency in cattle. Microbiome 2019, 7, 92. [Google Scholar] [CrossRef]
- Koch, R.M.; Swiger, L.A.; Chambers, D.; Gregory, K.E. Efficiency of Feed Use in Beef Cattle. J. Anim. Sci. 1963, 22, 486–494. [Google Scholar] [CrossRef]
- Stewart, R.L. Cutting Costs, Not Corners: Managing Cattle in Tough Times; UGA Cooperative Extension: Athens, GA, USA, 2010. [Google Scholar]
- Becker, G.S. Livestock Feed Costs: Concerns and Options; Congressional Research Service, Library of Congress: Washington, DC, USA, 2008. [Google Scholar]
- Herd, R.M.; Archer, J.A.; Arthur, P.F. Reducing the cost of beef production through genetic improvement in residual feed intake: Opportunity and challenges to application1. J. Anim. Sci. 2003, 81, E9–E17. [Google Scholar] [CrossRef]
- Guan, L.L.; Nkrumah, J.D.; Basarab, J.A.; Moore, S.S. Linkage of microbial ecology to phenotype: Correlation of rumen microbial ecology to cattle’s feed efficiency. FEMS Microbiol. Lett. 2008, 288, 85–91. [Google Scholar] [CrossRef]
- Hernandez-Sanabria, E.; Goonewardene, L.A.; Wang, Z.; Durunna, O.N.; Moore, S.S.; Guan, L.L. Impact of Feed Efficiency and Diet on Adaptive Variations in the Bacterial Community in the Rumen Fluid of Cattle. Appl. Environ. Microbiol. 2012, 78, 1203–1214. [Google Scholar] [CrossRef]
- McLoughlin, S.; Spillane, C.; Claffey, N.; Smith, P.E.; O’Rourke, T.; Diskin, M.G.; Waters, S.M. Rumen Microbiome Composition Is Altered in Sheep Divergent in Feed Efficiency. Front. Microbiol. 2020, 11, 1981. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, F.; Chen, Y.; Wu, H.; Meng, Q.; Guan, L.L. Metatranscriptomic Profiling Reveals the Effect of Breed on Active Rumen Eukaryotic Composition in Beef Cattle with Varied Feed Efficiency. Front. Microbiol. 2020, 11, 367. [Google Scholar] [CrossRef] [PubMed]
- Arthur, P.F.; Archer, J.A.; Johnston, D.J.; Herd, R.M.; Richardson, E.C.; Parnell, P.F. Genetic and phenotypic variance and covariance components for feed intake, feed efficiency, and other postweaning traits in Angus cattle. J. Anim. Sci. 2001, 79, 2805–2811. [Google Scholar] [CrossRef]
- Sasson, G.; Kruger Ben-Shabat, S.; Seroussi, E.; Doron-Faigenboim, A.; Shterzer, N.; Yaacoby, S.; Berg Miller, M.E.; White, B.A.; Halperin, E.; Mizrahi, I. Heritable Bovine Rumen Bacteria Are Phylogenetically Related and Correlated with the Cow’s Capacity to Harvest Energy from Its Feed. mBio 2017, 8, e00703-17. [Google Scholar] [CrossRef]
- Ellison, M.J.; Conant, G.C.; Lamberson, W.R.; Cockrum, R.R.; Austin, K.J.; Rule, D.C.; Cammack, K.M. Diet and feed efficiency status affect rumen microbial profiles of sheep. Small Rumin. Res. 2017, 156, 12–19. [Google Scholar] [CrossRef]
- Shabat, S.K.B.; Sasson, G.; Doron-Faigenboim, A.; Durman, T.; Yaacoby, S.; Berg Miller, M.E.; White, B.A.; Shterzer, N.; Mizrahi, I. Specific microbiome-dependent mechanisms underlie the energy harvest efficiency of ruminants. ISME J. 2016, 10, 2958–2972. [Google Scholar] [CrossRef] [PubMed]
- Owens, F.N.; Secrist, D.S.; Hill, W.J.; Gill, D.R. The effect of grain source and grain processing on performance of feedlot cattle: A review. J. Anim. Sci. 1997, 75, 868–879. [Google Scholar] [CrossRef] [PubMed]
- Newbold, C.J.; Ramos-Morales, E. Review: Ruminal microbiome and microbial metabolome: Effects of diet and ruminant host. Animal 2020, 14, s78–s86. [Google Scholar] [CrossRef]
- Elliott, C.L.; Edwards, J.E.; Wilkinson, T.J.; Allison, G.G.; McCaffrey, K.; Scott, M.B.; Rees-Stevens, P.; Kingston-Smith, A.H.; Huws, S.A. Using ‘Omic Approaches to Compare Temporal Bacterial Colonization of Lolium perenne, Lotus corniculatus, and Trifolium pratense in the Rumen. Front. Microbiol. 2018, 9, 2184. [Google Scholar] [CrossRef] [PubMed]
- Fernando, S.C.; Purvis, H.T.; Najar, F.Z.; Sukharnikov, L.O.; Krehbiel, C.R.; Nagaraja, T.G.; Roe, B.A.; DeSilva, U. Rumen Microbial Population Dynamics during Adaptation to a High-Grain Diet. Appl. Environ. Microbiol. 2010, 76, 7482–7490. [Google Scholar] [CrossRef]
- Kittelmann, S.; Kirk, M.R.; Jonker, A.; McCulloch, A.; Janssen, P.H. Buccal Swabbing as a Noninvasive Method to Determine Bacterial, Archaeal, and Eukaryotic Microbial Community Structures in the Rumen. Appl. Environ. Microbiol. 2015, 81, 7470–7483. [Google Scholar] [CrossRef]
- Petri, R.M.; Schwaiger, T.; Penner, G.B.; Beauchemin, K.A.; Forster, R.J.; McKinnon, J.J.; McAllister, T.A. Characterization of the Core Rumen Microbiome in Cattle during Transition from Forage to Concentrate as Well as during and after an Acidotic Challenge. PLoS ONE 2013, 8, e83424. [Google Scholar] [CrossRef] [PubMed]
- Elam, C.J. Acidosis in Feedlot Cattle: Practical Observations. J. Anim. Sci. 1976, 43, 898–901. [Google Scholar] [CrossRef]
- Klieve, A.V.; Hennessy, D.; Ouwerkerk, D.; Forster, R.J.; Mackie, R.I.; Attwood, G.T. Establishing populations of Megasphaera elsdenii YE 34 and Butyrivibrio fibrisolvens YE 44 in the rumen of cattle fed high grain diets. J. Appl. Microbiol. 2003, 95, 621–630. [Google Scholar] [CrossRef]
- Nagaraja, T.G.; Titgemeyer, E.C. Ruminal Acidosis in Beef Cattle: The Current Microbiological and Nutritional Outlook. J. Dairy Sci. 2007, 90, E17–E38. [Google Scholar] [CrossRef] [PubMed]
- Petri, R.M.; Forster, R.J.; Yang, W.; McKinnon, J.J.; McAllister, T.A. Characterization of rumen bacterial diversity and fermentation parameters in concentrate fed cattle with and without forage. J. Appl. Microbiol. 2012, 112, 1152–1162. [Google Scholar] [CrossRef] [PubMed]
- Russell, J.B. Rumen Microbiology and Its Role in Ruminant Nutrition; Department of Microbiology, Cornell University: Ithaca, NY, USA, 2002. [Google Scholar]
- Steele, M.A.; Croom, J.; Kahler, M.; AlZahal, O.; Hook, S.E.; Plaizier, K.; McBride, B.W. Bovine rumen epithelium undergoes rapid structural adaptations during grain-induced subacute ruminal acidosis. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2011, 300, R1515–R1523. [Google Scholar] [CrossRef]
- Lamb, G.C.; Black, T.E.; Bischoff, K.M.; Mercadante, V.R.G. The Importance of Feed Efficiency in the Cow Herd; University of Florida: Marianna, FL, USA, 2013. [Google Scholar]
- Difford, G.F.; Plichta, D.R.; Løvendahl, P.; Lassen, J.; Noel, S.J.; Højberg, O.; Wright, A.-D.G.; Zhu, Z.; Kristensen, L.; Nielsen, H.B.; et al. Host genetics and the rumen microbiome jointly associate with methane emissions in dairy cows. PLoS Genet. 2018, 14, e1007580. [Google Scholar] [CrossRef]
- Plaizier, J.C.; Mesgaran, M.D.; Derakhshani, H.; Golder, H.; Khafipour, E.; Kleen, J.L.; Lean, I.; Loor, J.; Penner, G.; Zebeli, Q. Review: Enhancing gastrointestinal health in dairy cows. Animal 2018, 12, s399–s418. [Google Scholar] [CrossRef] [PubMed]
- Greenwood, P.L. Review: An overview of beef production from pasture and feedlot globally, as demand for beef and the need for sustainable practices increase. Animal 2021, 15, 100295. [Google Scholar] [CrossRef]
- Richards, M.W.; Spitzer, J.C.; Warner, M.B. Effect of Varying Levels of Postpartum Nutrition and Body Condition at Calving on Subsequent Reproductive Performance in Beef Cattle23. J. Anim. Sci. 1986, 62, 300–306. [Google Scholar] [CrossRef]
- Winters, L.M.; Green, W.W.; Comstock, R.E. Prenatal Development of the Bovine; University of Minnesota: Minneapolis, MN, USA, 1942; Available online: https://conservancy.umn.edu/bitstream/handle/11299/204085/mn1000_agexpstn_tb_151.pdf?sequence=1 (accessed on 23 September 2024).
- Swett, W.W.; Matthews, C.A.; Fohrman, M.H. Development of the Fetus in the Dairy Cow; University of Minnesota: Minneapolis, MN, USA, 1948; Available online: https://ageconsearch.umn.edu/record/170289/files/tb964.pdf (accessed on 23 September 2024).
- Lyne, A.G. Pre-natal growth of cattle. Proc. Aust. Soc. Anim. Prod. 1960, 3, 153–161. [Google Scholar]
- Ferrell, C.L.; Garrett, W.N.; Hinman, N. Growth, Development and Composition of the Udder and Gravid Uterus of Beef Heifers during Pregnancy. J. Anim. Sci. 1976, 42, 1477–1489. [Google Scholar] [CrossRef]
- Prior, R.L.; Laster, D.B. Development of the Bovine Fetus1. J. Anim. Sci. 1979, 48, 1546–1553. [Google Scholar] [CrossRef]
- Mao, W.H.; Albrecht, E.; Teuscher, F.; Yang, Q.; Zhao, R.Q.; Wegner, J. Growth- and Breed-related Changes of Fetal Development in Cattle. Asian-Australas. J. Anim. Sci. 2008, 21, 640–647. [Google Scholar] [CrossRef]
- Hall, J.B.; Seay, W.W.; Baker, S.M. Nutrition and Feeding of the Cow-Calf Herd: Production Cycle Nutrition and Nutrient Requirements of Cows, Pregnant Heifers and Bulls; Virginia Cooperative Extension: Blacksburg, VA, USA, 2001; Available online: https://vtechworks.lib.vt.edu/bitstreams/cc702ee1-e2de-47ae-ae40-04f70de23a14/download (accessed on 23 September 2024).
- Campos, V.; Teixeira, S.; Bianco, B.; Silva, F.A.; Chizzotti, M. Nutrient Requirements of Zebu and Crossbred Cattle-BR-CORTE; Universidade Federal de Viçosa: Viçosa, Brazil, 2023; ISBN 978-85-8179-194-4. [Google Scholar]
- Weller, M.M.D.C.A.; Fortes, M.R.S.; Marcondes, M.I.; Rotta, P.P.; Gionbeli, T.R.S.; Valadares Filho, S.C.; Campos, M.M.; Silva, F.F.; Silva, W.; Moore, S.; et al. Effect of maternal nutrition and days of gestation on pituitary gland and gonadal gene expression in cattle. J. Dairy Sci. 2016, 99, 3056–3071. [Google Scholar] [CrossRef]
- Wu, G.; Bazer, F.W.; Wallace, J.M.; Spencer, T.E. BOARD-INVITED REVIEW: Intrauterine growth retardation: Implications for the animal sciences1. J. Anim. Sci. 2006, 84, 2316–2337. [Google Scholar] [CrossRef] [PubMed]
- Derakhshani, H.; Tun, H.M.; Cardoso, F.C.; Plaizier, J.C.; Khafipour, E.; Loor, J.J. Linking Peripartal Dynamics of Ruminal Microbiota to Dietary Changes and Production Parameters. Front. Microbiol. 2017, 7, 2143. [Google Scholar] [CrossRef]
- Lima, F.S.; Oikonomou, G.; Lima, S.F.; Bicalho, M.L.S.; Ganda, E.K.; de Oliveira Filho, J.C.; Lorenzo, G.; Trojacanec, P.; Bicalho, R.C. Prepartum and Postpartum Rumen Fluid Microbiomes: Characterization and Correlation with Production Traits in Dairy Cows. Appl. Environ. Microbiol. 2015, 81, 1327–1337. [Google Scholar] [CrossRef]
- Elderman, M.; Hugenholtz, F.; Belzer, C.; Boekschoten, M.; de Haan, B.; de Vos, P.; Faas, M. Changes in intestinal gene expression and microbiota composition during late pregnancy are mouse strain dependent. Sci. Rep. 2018, 8, 10001. [Google Scholar] [CrossRef]
- Koren, O.; Goodrich, J.K.; Cullender, T.C.; Spor, A.; Laitinen, K.; Bäckhed, H.K.; Gonzalez, A.; Werner, J.J.; Angenent, L.T.; Knight, R.; et al. Host Remodeling of the Gut Microbiome and Metabolic Changes during Pregnancy. Cell 2012, 150, 470–480. [Google Scholar] [CrossRef] [PubMed]
- Phillips, C.D.; Phelan, G.; Dowd, S.E.; McDONOUGH, M.M.; Ferguson, A.W.; Delton Hanson, J.; Siles, L.; Ordóñez-Garza, N.; San Francisco, M.; Baker, R.J. Microbiome analysis among bats describes influences of host phylogeny, life history, physiology and geography. Mol. Ecol. 2012, 21, 2617–2627. [Google Scholar] [CrossRef]
- Mukhopadhya, I.; Hansen, R.; El-Omar, E.M.; Hold, G.L. IBD—What role do Proteobacteria play? Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 219–230. [Google Scholar] [CrossRef]
- Castro, N.; Kawashima, C.; van Dorland, H.A.; Morel, I.; Miyamoto, A.; Bruckmaier, R.M. Metabolic and energy status during the dry period is crucial for the resumption of ovarian activity postpartum in dairy cows. J. Dairy Sci. 2012, 95, 5804–5812. [Google Scholar] [CrossRef]
- D’Occhio, M.J.; Baruselli, P.S.; Campanile, G. Influence of nutrition, body condition, and metabolic status on reproduction in female beef cattle: A review. Theriogenology 2019, 125, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Lucy, M. Functional Differences in the Growth Hormone and Insulin-like Growth Factor Axis in Cattle and Pigs: Implications for Post-partum Nutrition and Reproduction. Reprod. Domest. Anim. 2008, 43, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Lucy, M.C. Regulation of Ovarian Follicular Growth by Somatotropin and Insulin-Like Growth Factors in Cattle1. J. Dairy Sci. 2000, 83, 1635–1647. [Google Scholar] [CrossRef] [PubMed]
- Lucy, M.C.; Butler, S.T.; Garverick, H.A. Endocrine and metabolic mechanisms linking postpartum glucose with early embryonic and foetal development in dairy cows. Animal 2014, 8, 82–90. [Google Scholar] [CrossRef]
- Roche, J.R.; Burke, C.R.; Meier, S.; Walker, C.G. Nutrition × reproduction interaction in pasture-based systems: Is nutrition a factor in reproductive failure? Anim. Prod. Sci. 2011, 51, 1045–1066. [Google Scholar] [CrossRef]
- Samadi, F.; Phillips, N.J.; Blache, D.; Martin, G.B.; D’Occhio, M.J. Interrelationships of nutrition, metabolic hormones and resumption of ovulation in multiparous suckled beef cows on subtropical pastures. Anim. Reprod. Sci. 2013, 137, 137–144. [Google Scholar] [CrossRef]
- Sartori, R.; Gimenes, L.U.; Monteiro, P.L.J.; Melo, L.F.; Baruselli, P.S.; Bastos, M.R. Metabolic and endocrine differences between Bos taurus and Bos indicus females that impact the interaction of nutrition with reproduction. Theriogenology 2016, 86, 32–40. [Google Scholar] [CrossRef]
- Silva, J.R.V.; Figueiredo, J.R.; van den Hurk, R. Involvement of growth hormone (GH) and insulin-like growth factor (IGF) system in ovarian folliculogenesis. Theriogenology 2009, 71, 1193–1208. [Google Scholar] [CrossRef]
- Velazquez, M.A.; Spicer, L.J.; Wathes, D.C. The role of endocrine insulin-like growth factor-I (IGF-I) in female bovine reproduction. Domest. Anim. Endocrinol. 2008, 35, 325–342. [Google Scholar] [CrossRef]
- Sartori, R.; Guardieiro, M.M.; Surjus, R.S.; Melo, L.F.; Prata, A.B.; Ishiguro, M.; Bastos, M.R.; Nascimento, A.B. Metabolic hormones and reproductive function in cattle. Anim. Reprod. 2018, 10, 199–205. [Google Scholar]
- LeBlanc, S. Monitoring programs for transition dairy cows. In Proceedings of the 26th World Biuatrics Congress, Nice, France, 15 October 2006. [Google Scholar]
- Smith, V.G.; Edgerton, L.A.; Hafs, H.D.; Convey, E.M. Bovine Serum Estrogens, Progestins and Glucocorticoids during Late Pregnancy, Parturition and Early Lactation. J. Anim. Sci. 1973, 36, 391–396. [Google Scholar] [CrossRef] [PubMed]
- Journet, M.; Remond, B. Physiological factors affecting the voluntary intake of feed by cows: A review. Livest. Prod. Sci. 1976, 3, 129–146. [Google Scholar] [CrossRef]
- Beever, D.E. The impact of controlled nutrition during the dry period on dairy cow health, fertility and performance. Anim. Reprod. Sci. 2006, 96, 212–226. [Google Scholar] [CrossRef] [PubMed]
- Grummer, R.R. Etiology of Lipid-Related Metabolic Disorders in Periparturient Dairy Cows. J. Dairy Sci. 1993, 76, 3882–3896. [Google Scholar] [CrossRef]
- Spain, J.N.; Scheer, W.A. The 100-day contract with the dairy cow: 30 days prepartum to 70 days postpartum. In Proceedings of the Tri-State Dairy Nutrition Conference, Fort Wayne, IN, USA, 17–18 April 2001; Volume 414, p. 13. Available online: https://citeseerx.ist.psu.edu/document?repid=rep1&type=pdf&doi=3d2aa04b7a561887ac2197f8730d7f4de3b92654#page=20 (accessed on 23 September 2024).
- Wathes, D. Mechanisms Linking Metabolic Status and Disease with Reproductive Outcome in the Dairy Cow. Reprod. Domest. Anim. 2012, 47, 304–312. [Google Scholar] [CrossRef]
- Mulak, A.; Taché, Y.; Larauche, M. Sex hormones in the modulation of irritable bowel syndrome. World J. Gastroenterol. WJG 2014, 20, 2433–2448. [Google Scholar] [CrossRef] [PubMed]
- Org, E.; Mehrabian, M.; Parks, B.W.; Shipkova, P.; Liu, X.; Drake, T.A.; Lusis, A.J. Sex differences and hormonal effects on gut microbiota composition in mice. Gut Microbes 2016, 7, 313–322. [Google Scholar] [CrossRef]
- Adlercreutz, H.; Martin, F.; Järvenpää, P.; Fotsis, T. Steroid absorption and enterohepatic recycling. Contraception 1979, 20, 201–223. [Google Scholar] [CrossRef]
- Henniger, M.T.; Wells, J.E.; Hales, K.E.; Lindholm-Perry, A.K.; Freetly, H.C.; Kuehn, L.A.; Schneider, L.G.; McLean, K.J.; Campagna, S.R.; Christopher, C.J.; et al. Effects of a Moderate or Aggressive Implant Strategy on the Rumen Microbiome and Metabolome in Steers. Front. Anim. Sci. 2022, 3, 889817. [Google Scholar] [CrossRef]
- Ault-Seay, T.B.; Brandt, K.J.; Henniger, M.T.; Payton, R.R.; Mathew, D.J.; Moorey, S.E.; Schrick, F.N.; Pohler, K.G.; Smith, T.P.L.; Rhinehart, J.D.; et al. Bacterial Communities of the Uterus and Rumen during Heifer Development with Protein Supplementation. Front. Anim. Sci. 2022, 3, 903909. [Google Scholar] [CrossRef]
- Laguardia-Nascimento, M.; Branco, K.M.G.R.; Gasparini, M.R.; Giannattasio-Ferraz, S.; Leite, L.R.; Araujo, F.M.G.; de Matos Salim, A.C.; Nicoli, J.R.; de Oliveira, G.C.; Barbosa-Stancioli, E.F. Vaginal Microbiome Characterization of Nellore Cattle Using Metagenomic Analysis. PLoS ONE 2015, 10, e0143294. [Google Scholar] [CrossRef] [PubMed]
- Ault, T.B.; Clemmons, B.A.; Reese, S.T.; Dantas, F.G.; Franco, G.A.; Smith, T.P.L.; Edwards, J.L.; Myer, P.R.; Pohler, K.G. Bacterial taxonomic composition of the postpartum cow uterus and vagina prior to artificial insemination1. J. Anim. Sci. 2019, 97, 4305–4313. [Google Scholar] [CrossRef] [PubMed]
- Myer, P.R.; Seay, T.B.; Rhinehart, J. Cattle Gut Microbe Series: Lactobacillus Species; W905. University of Tennessee: Knoxville, TN, USA, 2020. [Google Scholar]
- Myer, P.R.; Seay, T.B.; Rhinehart, J. Cattle Gut Microbe Series: Ruminococcus Species; W398. University of Tennessee: Knoxville, TN, USA, 2020. [Google Scholar]
- Zeineldin, M.; Barakat, R.; Elolimy, A.; Salem, A.Z.M.; Elghandour, M.M.Y.; Monroy, J.C. Synergetic action between the rumen microbiota and bovine health. Microb. Pathog. 2018, 124, 106–115. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).