
TRI14 Is Critical for Fusarium graminearum Infection and Spread in Wheat


 , , ,
, , ,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Strains and Plant Cultivation
2.2. Identification of TRI Gene Orthologs in Fungi
2.3. FHB Virulence Assays
2.4. Gene Expression Analyses by Quantitative PCR (qPCR)
2.5. Fungal Biomass Quantification
2.6. Trichothecene Production in Liquid Culture and in Planta
2.7. ROS Assays
2.8. Effect of Plant Defense Compounds on GZ3639 and ∆tri14 Growth
2.9. Tri14 Three-Dimensional (3D) Structure Prediction
2.10. Statistical Analysis
3. Results
3.1. TRI14 Is Conserved among Trichothecene-Producing Fungi
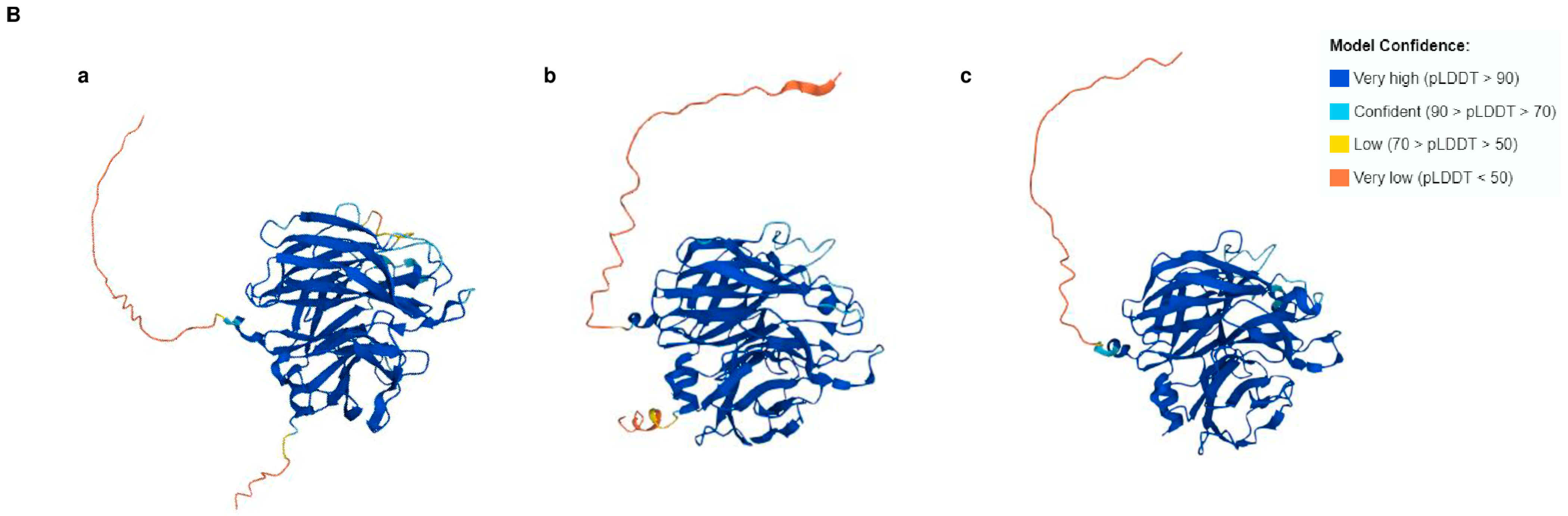
3.2. Tri14 Orthologs Share Conserved Three-Dimensional Structures
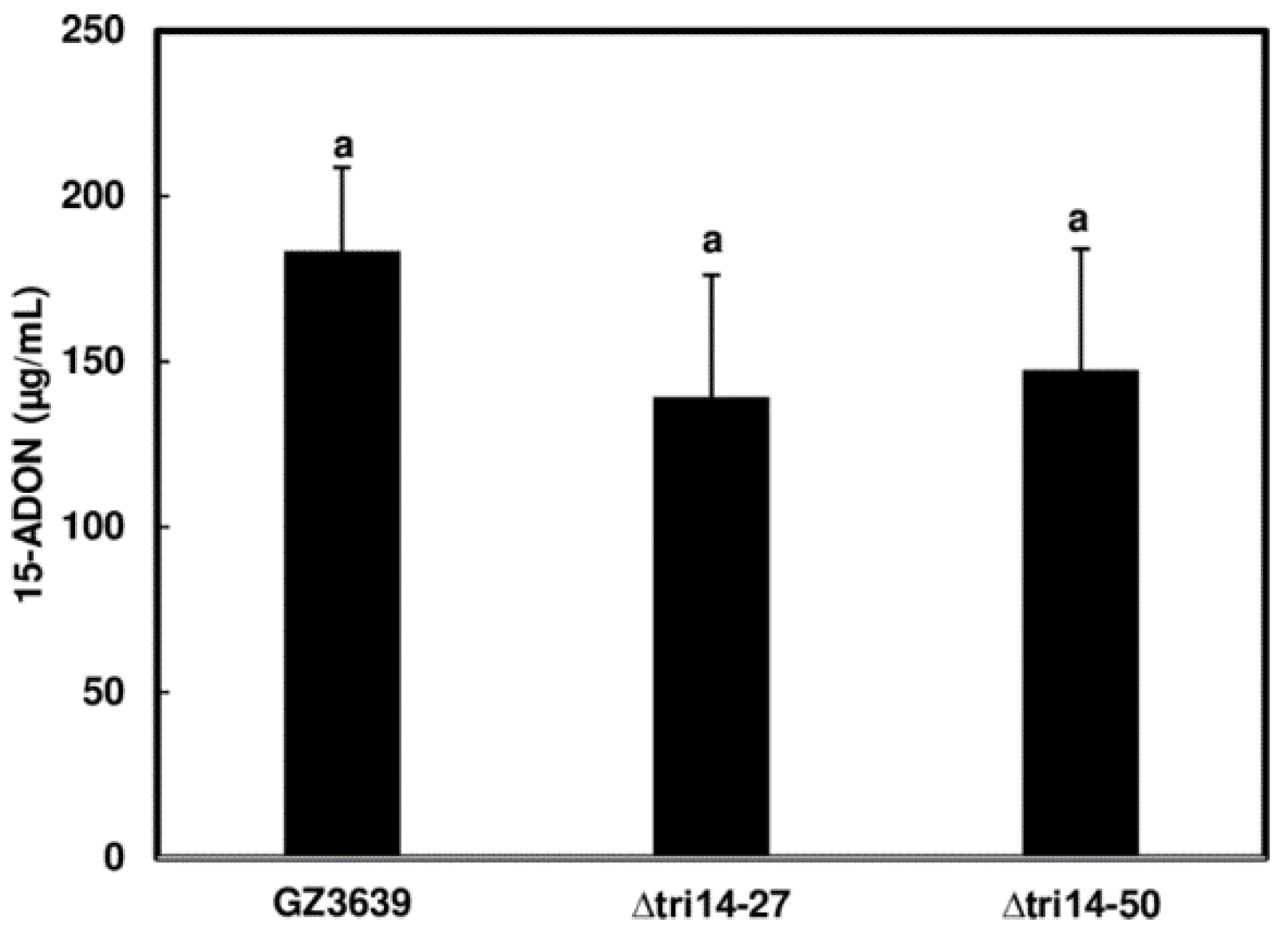
3.3. TRI14 Deletion Does Not Affect DON Production of F. graminearum in Liquid Cultures
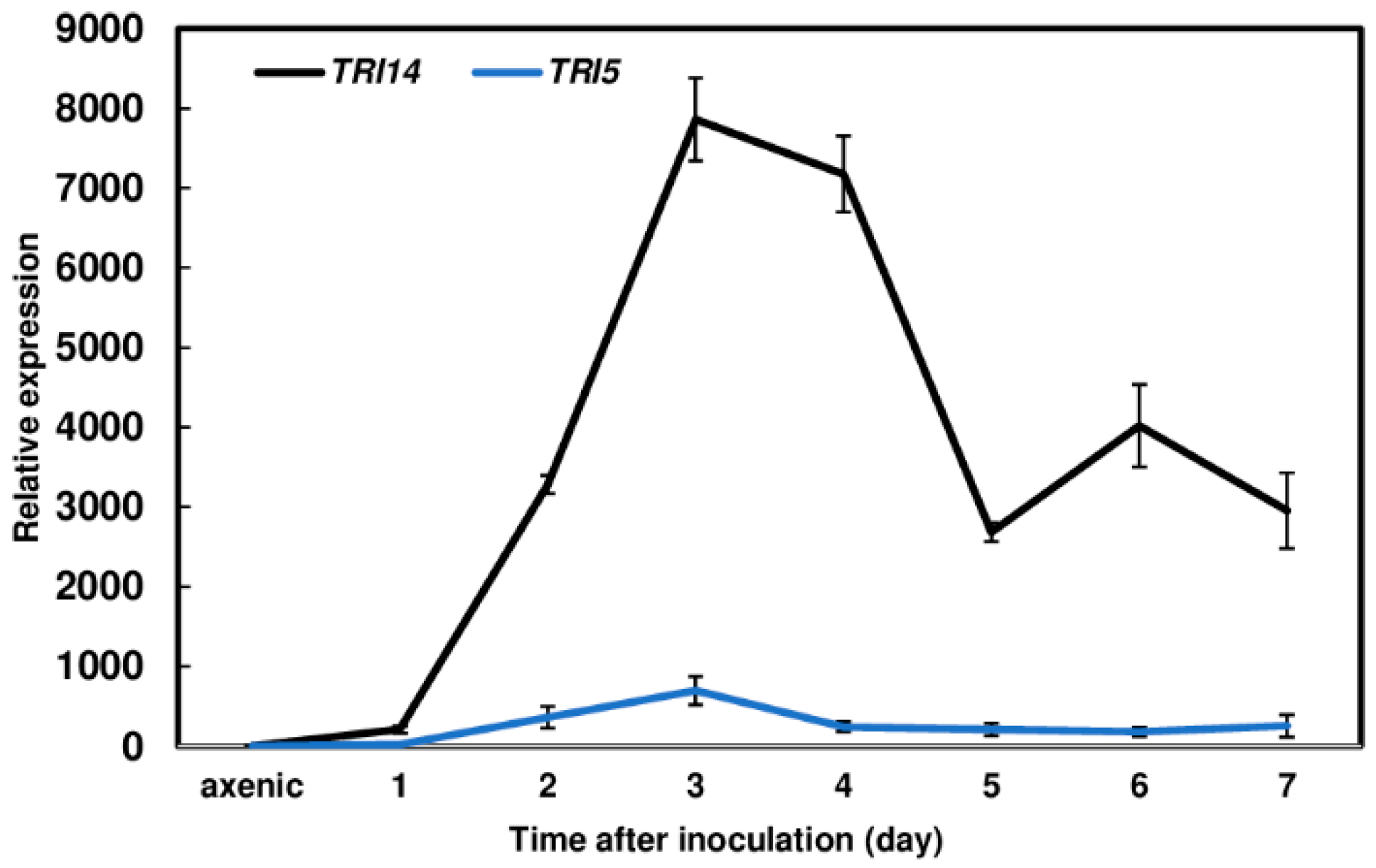
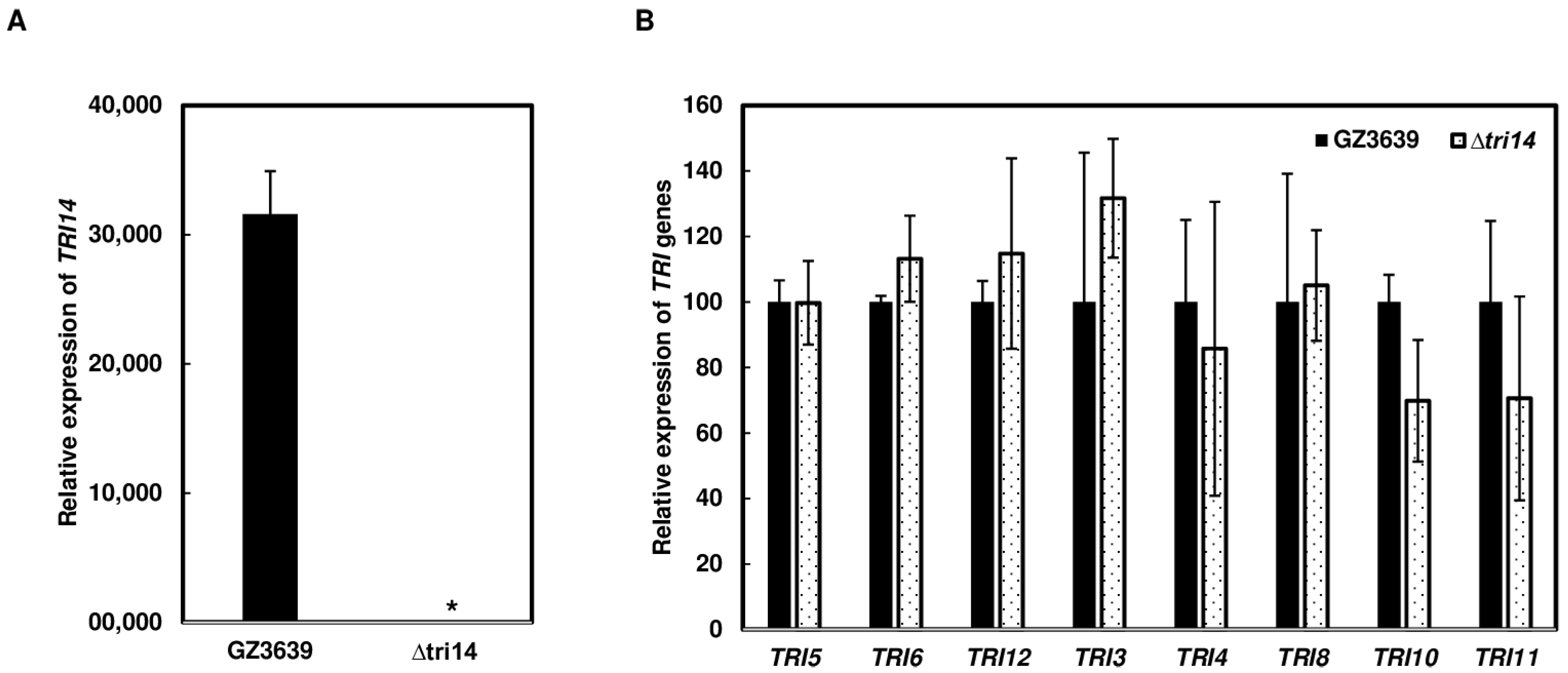
3.4. TRI14 Expression Is Highly Induced during Wheat Head Infection
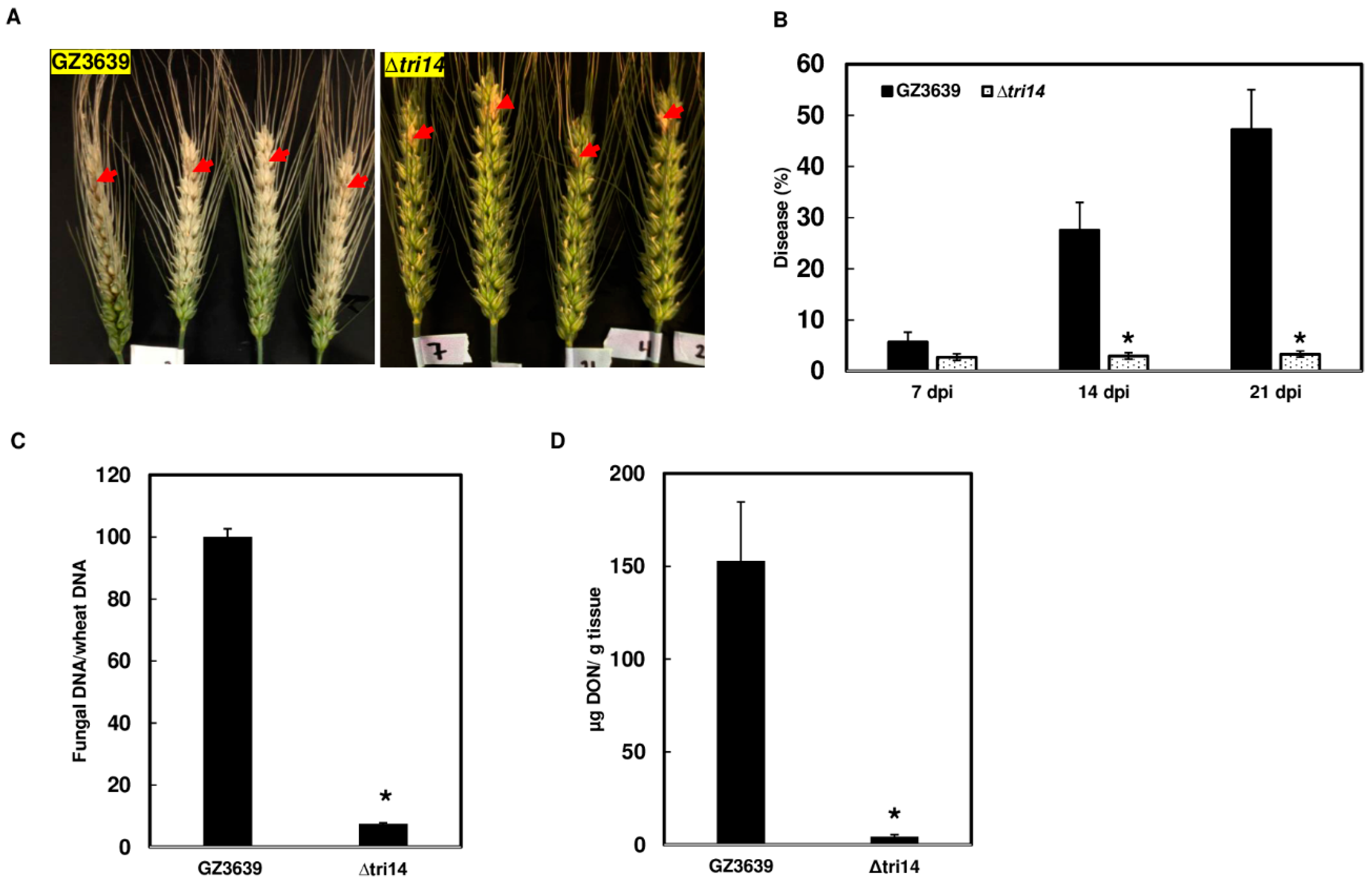
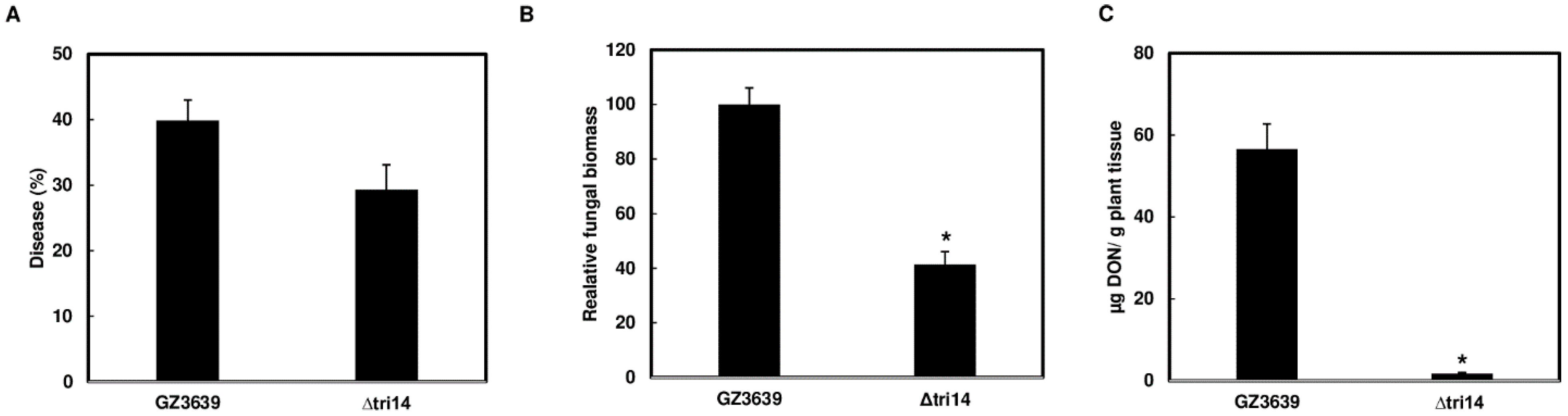
3.5. ∆tri14 Has Restricted FHB Symptoms, Less Growth and DON Production in the Inoculated Spikelets
3.6. ∆tri14 Has Reduced Growth and DON Production in Dip-Inoculated Wheat Heads
3.7. Deletion of TRI14 Does Not Affect TRI Cluster Gene Expression
3.8. Deletion of TRI14 Reduces Fungal Growth and Lesions in Wheat Coleoptiles
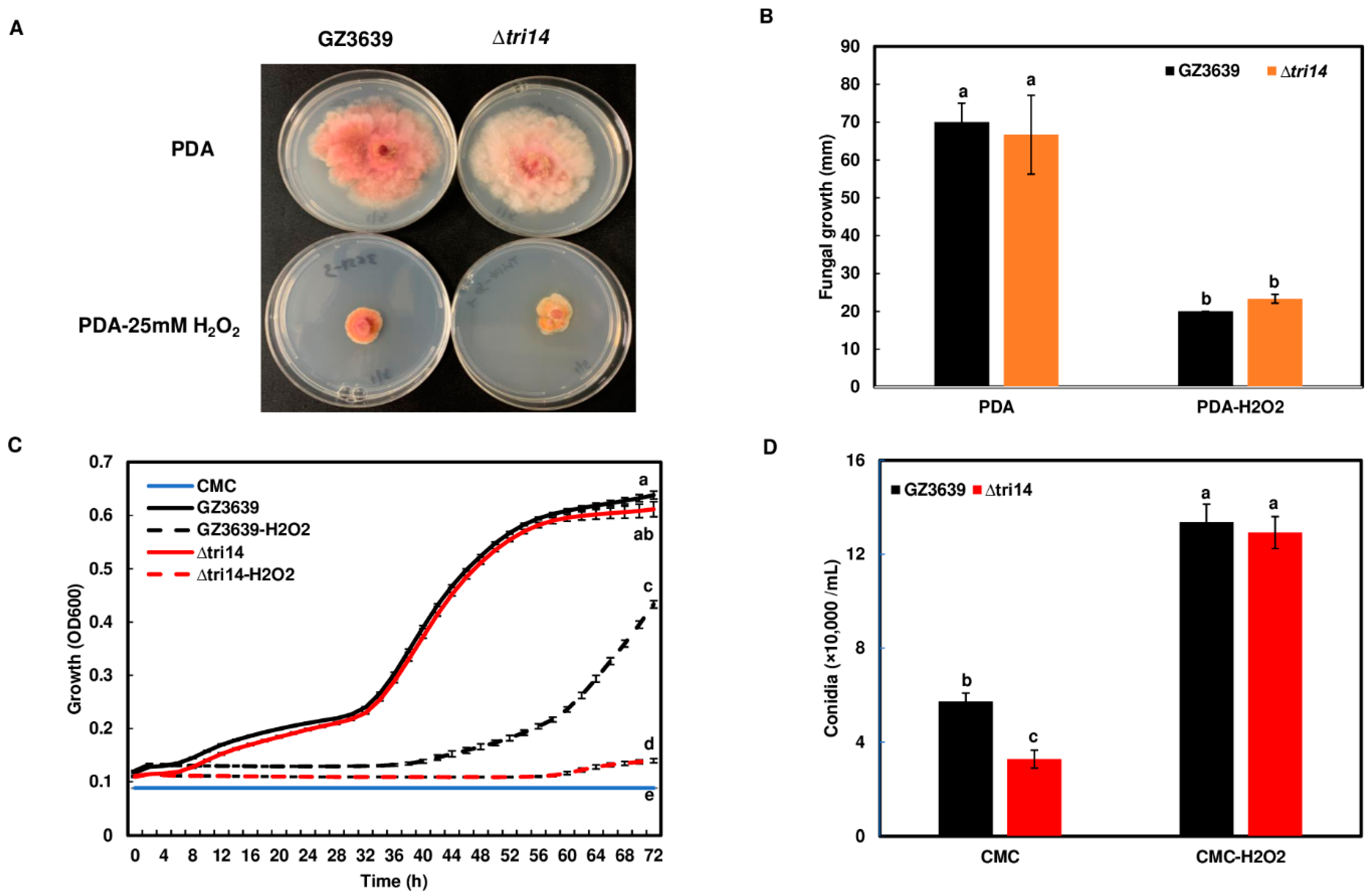
3.9. TRI14 Mutant Is More Sensitive to H2O2 under Certain Growth Conditions
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Proctor, R.H.; McCormick, S.P.; Gutierrez, S. Genetic bases for variation in structure and biological activity of trichothecene toxins produced by diverse fungi. Appl. Microbiol. Biot. 2020, 104, 5185–5199. [Google Scholar] [CrossRef]
- Desjardins, A.E.; McCormick, S.P.; Appell, M. Structure-activity relationships of trichothecene toxins in an Arabidopsis thaliana leaf assay. J. Agric. Food Chem. 2007, 55, 6487–6492. [Google Scholar] [CrossRef]
- Wilson, W.; Dahl, B.; Nganje, W. Economic costs of Fusarium head blight, scab and deoxynivalenol. World Mycotoxin J. 2018, 11, 291–302. [Google Scholar] [CrossRef]
- Bai, G.H.; Desjardins, A.E.; Plattner, R.D. Deoxynivalenol-nonproducing fusarium graminearum causes initial infection, but does not cause disease spread in wheat spikes. Mycopathologia 2002, 153, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Proctor, R.H.; Hohn, T.M.; McCormick, S.P. Reduced virulence of Gibberella zeae caused by disruption of a trichothecene toxin biosynthetic gene. Mol. Plant Microbe Interact. 1995, 8, 593–601. [Google Scholar] [CrossRef]
- Proctor, R.H.; McCormick, S.P.; Kim, H.S.; Cardoza, R.E.; Stanley, A.M.; Lindo, L.; Kelly, A.; Brown, D.W.; Lee, T.; Vaughan, M.M.; et al. Evolution of structural diversity of trichothecenes, a family of toxins produced by plant pathogenic and entomopathogenic fungi. PLoS Pathog. 2018, 14, e1006946. [Google Scholar] [CrossRef]
- Proctor, R.H.; Hao, G.; Kim, H.S.; Whitaker, B.K.; Laraba, I.; Vaughan, M.M.; McCormick, S.P. A Novel Trichothecene Toxin Phenotype Associated with Horizontal Gene Transfer and a Change in Gene Function in Fusarium. Toxins 2022, 15, 12. [Google Scholar] [CrossRef] [PubMed]
- Villani, A.; Proctor, R.H.; Kim, H.S.; Brown, D.W.; Logrieco, A.F.; Amatulli, M.T.; Moretti, A.; Susca, A. Variation in secondary metabolite production potential in the Fusarium incarnatum-equiseti species complex revealed by comparative analysis of 13 genomes. BMC Genom. 2019, 20, 314. [Google Scholar] [CrossRef] [PubMed]
- McCormick, S.P.; Hohn, T.M.; Desjardins, A.E. Isolation and characterization of Tri3, a gene encoding 15-O- acetyltransferase from Fusarium sporotrichioides. Appl. Environ. Microbiol. 1996, 62, 353–359. [Google Scholar] [CrossRef]
- Alexander, N.J.; Proctor, R.H.; McCormick, S.P. Genes, gene clusters, and biosynthesis of trichothecenes and fumonisins in Fusarium. Toxin Rev. 2009, 24, 198–215. [Google Scholar] [CrossRef]
- Dyer, R.B.; Plattner, R.D.; Kendra, D.F.; Brown, D.W. Fusarium graminearum TRI14 is required for high virulence and DON production on wheat but not for DON synthesis in vitro. J. Agric. Food Chem. 2005, 53, 9281–9287. [Google Scholar] [CrossRef] [PubMed]
- Hao, G.; McCormick, S.; Usgaard, T.; Tiley, H.; Vaughan, M.M. Characterization of three Fusarium graminearum effectors and their roles during Fusarium Head Blight. Front. Plant Sci. 2020, 11, 579553. [Google Scholar] [CrossRef]
- Jansen, C.; von Wettstein, D.; Schafer, W.; Kogel, K.H.; Felk, A.; Maier, F.J. Infection patterns in barley and wheat spikes inoculated with wild-type and trichodiene synthase gene disrupted Fusarium graminearum. Proc. Natl. Acad. Sci. USA 2005, 102, 16892–16897. [Google Scholar] [CrossRef]
- Hao, G.; McCormick, S.; Tiley, H.; Gutierrez, S.; Yulfo-Soto, G.; Vaughan, M.M.; Ward, T.J. NX Trichothecenes Are Required for Fusarium graminearum Infection of Wheat. Mol. Plant Microbe Interact. 2023, 36, 294–304. [Google Scholar] [CrossRef] [PubMed]
- Bowden, R.L.; Leslie, J.F. Nitrate non-utilizing mutants of Gibberella zeae and their use in determining vegetative compatibility. Exp. Mycol. 1992, 16, 308–315. [Google Scholar] [CrossRef]
- Tuite, J. Plant Pathological Methods: Fungi and Bacteria; Burgess Publishing Company: Minneapolis, MN, USA, 1969; 239p. [Google Scholar]
- Bai, G.-H.; Shaner, G. Variation in Fusarium graminearum and cultivar resistance to wheat scab. Plant Dis. 1996, 80, 975–979. [Google Scholar] [CrossRef]
- Gutierrez, S.; McCormick, S.P.; Cardoza, R.E.; Kim, H.S.; Yugueros, L.L.; Vaughan, M.M.; Carro-Huerga, G.; Busman, M.; Saenz de Miera, L.E.; Jaklitsch, W.M.; et al. Distribution, function, and evolution of a gene essential for trichothecene toxin biosynthesis in Trichoderma. Front. Microbiol. 2021, 12, 791641. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.W.; Jia, L.J.; Zhang, Y.; Jiang, G.; Li, X.; Zhang, D.; Tang, W.H. In planta stage-specific fungal gene profiling elucidates the molecular strategies of Fusarium graminearum growing inside wheat coleoptiles. Plant Cell 2012, 24, 5159–5176. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Gardiner, D.M.; Kazan, K.; Manners, J.M. Novel genes of Fusarium graminearum that negatively regulate deoxynivalenol production and virulence. Mol. Plant-Microbe Interact. 2009, 22, 1588–1600. [Google Scholar] [CrossRef]
- Hao, G.; McCormick, S.; Vaughan, M.M.; Naumann, T.A.; Kim, H.S.; Proctor, R.; Kelly, A.; Ward, T.J. Fusarium graminearum arabinanase (Arb93B) enhances wheat head blight susceptibility by suppressing plant immunity. Mol. Plant Microbe Interact. 2019, 32, 888–898. [Google Scholar] [CrossRef] [PubMed]
- Hao, G.; Tiley, H.; McCormick, S. Chitin triggers tissue-specific immunity in wheat associated with Fusarium Head Blight. Front. Plant Sci. 2022, 13, 832502. [Google Scholar] [CrossRef] [PubMed]
- Hao, G.; Naumann, T.A.; Chen, H.; Bai, G.; McCormick, S.; Kim, H.; Tian, B.; Trick, H.N.; Naldrett, M.J.; Proctor, R. Fusarium graminearum effector FgNls1 targets plant nuclei to induce wheat head blight. Mol. Plant Microbe Interact. 2023, 36, 478–488. [Google Scholar] [CrossRef] [PubMed]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Zidek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Gibrat, J.F.; Madej, T.; Bryant, S.H. Surprising similarities in structure comparison. Curr. Opin. Struct. Biol. 1996, 6, 377–385. [Google Scholar] [CrossRef] [PubMed]
- Telford, J.C.; Yeung, J.H.; Xu, G.; Kiefel, M.J.; Watts, A.G.; Hader, S.; Chan, J.; Bennet, A.J.; Moore, M.M.; Taylor, G.L. The Aspergillus fumigatus sialidase is a 3-deoxy-D-glycero-D-galacto-2-nonulosonic acid hydrolase (KDNase): Structural and mechanistic insights. J. Biol. Chem. 2011, 286, 10783–10792. [Google Scholar] [CrossRef] [PubMed]
- Carapito, R.; Imberty, A.; Jeltsch, J.M.; Byrns, S.C.; Tam, P.H.; Lowary, T.L.; Varrot, A.; Phalip, V. Molecular basis of arabinobio-hydrolase activity in phytopathogenic fungi: Crystal structure and catalytic mechanism of Fusarium graminearum GH93 exo-alpha-L-arabinanase. J. Biol. Chem. 2009, 284, 12285–12296. [Google Scholar] [CrossRef] [PubMed]
- Corfield, T. Bacterial sialidases--roles in pathogenicity and nutrition. Glycobiology 1992, 2, 509–521. [Google Scholar] [CrossRef] [PubMed]
- Vilhelmsson, M.; Zargari, A.; Crameri, R.; Rasool, O.; Achour, A.; Scheynius, A.; Hallberg, B.M. Crystal structure of the major Malassezia sympodialis allergen Mala s 1 reveals a beta-propeller fold: A novel fold among allergens. J. Mol. Biol. 2007, 369, 1079–1086. [Google Scholar] [CrossRef]
- Amarasinghe, C.C.; Fernando, W.G. Comparative Analysis of Deoxynivalenol Biosynthesis Related Gene Expression among Different Chemotypes of Fusarium graminearum in Spring Wheat. Front. Microbiol. 2016, 7, 1229. [Google Scholar] [CrossRef]
- Hao, G.; Rhoades, N.A.; McCormick, S. Chitin and laminarin additively trigger wheat reactive oxygen species but not resistance to Fusarium head blight. Plant Direct 2023, 7, e538. [Google Scholar] [CrossRef]
- Walter, S.; Nicholson, P.; Doohan, F.M. Action and reaction of host and pathogen during Fusarium head blight disease. New Phytol. 2010, 185, 54–66. [Google Scholar] [CrossRef]
- Glenn, A.E.; Hinton, D.M.; Yates, I.E.; Bacon, C.W. Detoxification of corn antimicrobial compounds as the basis for isolating Fusarium verticillioides and some other Fusarium species from corn. Appl. Environ. Microbiol. 2001, 67, 2973–2981. [Google Scholar] [CrossRef]
- Wouters, F.C.; Blanchette, B.; Gershenzon, J.; Vassao, D.G. Plant defense and herbivore counter-defense: Benzoxazinoids and insect herbivores. Phytochem. Rev. 2016, 15, 1127–1151. [Google Scholar] [CrossRef]
- Boenisch, M.J.; Schafer, W. Fusarium graminearum forms mycotoxin producing infection structures on wheat. BMC Plant Biol. 2011, 11, 110. [Google Scholar] [CrossRef]
- Pritsch, C.; Muehlbauer, G.J.; Bushnell, W.R.; Somers, D.A.; Vance, C.P. Fungal development and induction of defense response genes during early infection of wheat spikes by Fusarium graminearum. Mol. Plant Microbe Interact. 2000, 13, 159–169. [Google Scholar] [CrossRef]
- Desmond, O.J.; Manners, J.M.; Stephens, A.E.; Maclean, D.J.; Schenk, P.M.; Gardiner, D.M.; Munn, A.L.; Kazan, K. The Fusarium mycotoxin deoxynivalenol elicits hydrogen peroxide production, programmed cell death and defence responses in wheat. Mol. Plant Pathol. 2008, 9, 435–445. [Google Scholar] [CrossRef]
- Seong, K.Y.; Pasquali, M.; Zhou, X.; Song, J.; Hilburn, K.; McCormick, S.; Dong, Y.; Xu, J.R.; Kistler, H.C. Global gene regulation by Fusarium transcription factors Tri6 and Tri10 reveals adaptations for toxin biosynthesis. Mol. Microbiol. 2009, 72, 354–367. [Google Scholar] [CrossRef]
- Boedi, S.; Berger, H.; Sieber, C.; Munsterkotter, M.; Maloku, I.; Warth, B.; Sulyok, M.; Lemmens, M.; Schuhmacher, R.; Guldener, U.; et al. Comparison of Fusarium graminearum Transcriptomes on Living or Dead Wheat Differentiates Substrate-Responsive and Defense-Responsive Genes. Front. Microbiol. 2016, 7, 1113. [Google Scholar] [CrossRef]
- Grant, J.J.; Loake, G.J. Role of reactive oxygen intermediates and cognate redox signaling in disease resistance. Plant Physiol. 2000, 124, 21–29. [Google Scholar] [CrossRef]
- Li, T.; Kim, D.; Lee, J. NADPH oxidase gene, FgNoxD, plays a critical role in development and virulence in Fusarium graminearum. Front. Microbiol. 2022, 13, 822682. [Google Scholar] [CrossRef]
- Lee, Y.; Son, H.; Shin, J.Y.; Choi, G.J.; Lee, Y.W. Genome-wide functional characterization of putative peroxidases in the head blight fungus Fusarium graminearum. Mol. Plant Pathol. 2018, 19, 715–730. [Google Scholar] [CrossRef]
- Heller, J.; Tudzynski, P. Reactive oxygen species in phytopathogenic fungi: Signaling, development, and disease. Annu. Rev. Phytopathol. 2011, 49, 369–390. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hao, G.; Proctor, R.H.; Brown, D.W.; Rhoades, N.A.; Naumann, T.A.; Kim, H.; Gutiėrrez, S.; McCormick, S.P. TRI14 Is Critical for Fusarium graminearum Infection and Spread in Wheat. Appl. Microbiol. 2024, 4, 839-855. https://doi.org/10.3390/applmicrobiol4020058
Hao G, Proctor RH, Brown DW, Rhoades NA, Naumann TA, Kim H, Gutiėrrez S, McCormick SP. TRI14 Is Critical for Fusarium graminearum Infection and Spread in Wheat. Applied Microbiology. 2024; 4(2):839-855. https://doi.org/10.3390/applmicrobiol4020058
Chicago/Turabian StyleHao, Guixia, Robert H. Proctor, Daren W. Brown, Nicholas A. Rhoades, Todd A. Naumann, HyeSeon Kim, Santiago Gutiėrrez, and Susan P. McCormick. 2024. "TRI14 Is Critical for Fusarium graminearum Infection and Spread in Wheat" Applied Microbiology 4, no. 2: 839-855. https://doi.org/10.3390/applmicrobiol4020058
APA StyleHao, G., Proctor, R. H., Brown, D. W., Rhoades, N. A., Naumann, T. A., Kim, H., Gutiėrrez, S., & McCormick, S. P. (2024). TRI14 Is Critical for Fusarium graminearum Infection and Spread in Wheat. Applied Microbiology, 4(2), 839-855. https://doi.org/10.3390/applmicrobiol4020058