Abstract
Gout, recognized as the most common form of inflammatory arthritis, arises from the accumulation of uric acid crystals, leading to intense pain, particularly in the big toe. This condition has traditionally been associated with the overproduction or reduced clearance of uric acid. Recent studies, however, have underscored the significant role of the gut microbiota in uric acid metabolism, impacting both its production and elimination. This emerging understanding suggests that maintaining gut health could offer innovative approaches to treating gout, complementing traditional dietary and pharmacological interventions. It highlights the potential of probiotics or microbiome-based therapies, indicating a future where treatments are tailored to an individual’s microbiome. This offers a fresh perspective on gout management and underscores the broader influence of the microbiota on health and disease.
1. Introduction
Gout, the most common form of inflammatory arthritis, has been known since ancient times. Recent findings on the occurrence and new cases of gout show significant variation, depending on the population examined and the research techniques used. These studies indicate a prevalence rate from less than 1% to 6.8%, and an incidence rate ranging from 0.58 to 2.89 per 1000 person-years [1]. Gout predominantly strikes males (20 times more often than females). Gout is a form of arthritis characterized by sudden and severe joint pain, often affecting the big toe. It is caused by the accumulation of uric acid crystals in the joints [2].
This condition comes from either an overproduction of uric acid or its inadequate elimination, causing sudden, severe pain, redness, warmth, and swelling in joints, often resolving in 5–10 days (Figure 1). The scientific name for uric acid (UA) is 2,6,8-trioxypurine, and UA is found in nearly all organisms and is excreted as a crystal slurry. In mammals, UA is exclusively produced from purine metabolism, while urea results from protein breakdown [3].
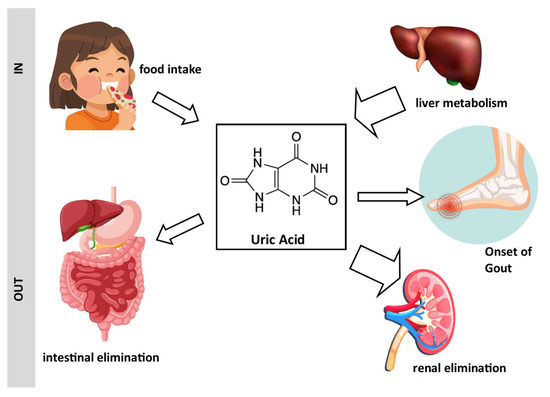
Figure 1.
Schematic representation of the origin of uric acid and its association with gout: Uric acid originates from dietary sources and hepatic biosynthesis. It is eliminated from the body through renal excretion and the gastrointestinal tract. Excessive levels of uric acid can lead to the development of gout. Drawing created with the assistance of Macrovector Image and brgfx on Freepik.
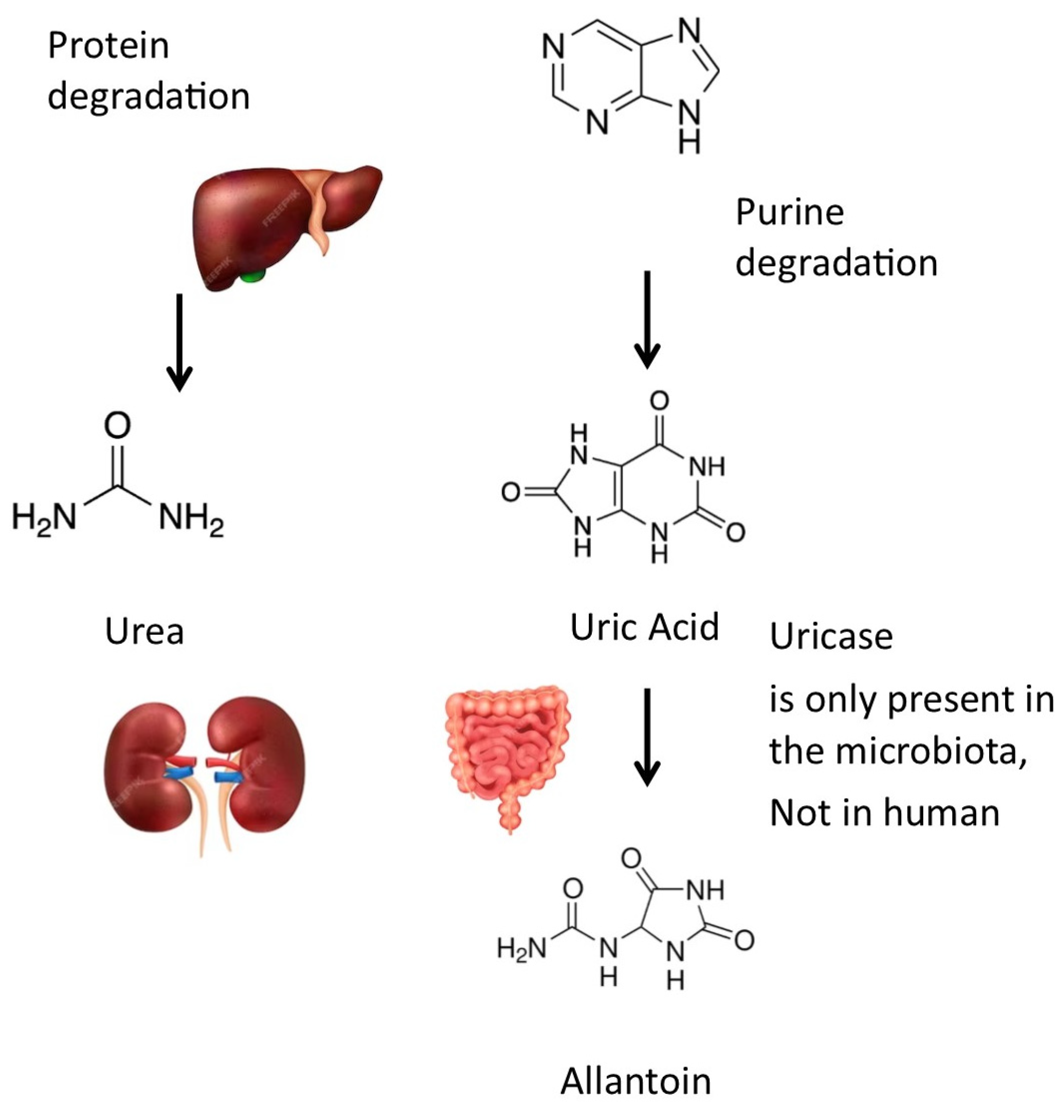
Uric acid can be converted into allantoin by uricase (or urate oxidase (EC 1.7.3.3)). Uricase, an enzyme present in certain bacteria within the gut microbiota, plays a significant role in the metabolism of uric acid. Unlike humans, who lack uricase due to evolutionary gene loss, these bacteria possess the ability to break down uric acid into allantoin [4] (Figure 2). Allantoin is more soluble and less likely to crystallize in joints. The activity of microbial uricase in the gut could potentially lower systemic uric acid levels and reduce the risk of gout, a condition characterized by painful joint inflammation due to uric acid crystal deposition. This suggests that modulating the gut microbiota to enhance the abundance of uricase-producing bacteria might be a promising strategy to manage hyperuricemia and gout. Such findings emphasize the potential therapeutic benefits of targeting the gut microbiome in gout prevention and management.
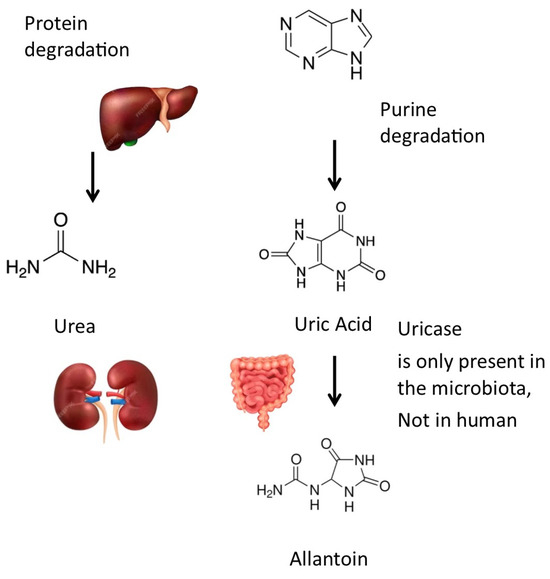
Figure 2.
Metabolic pathways for uric acid: Uric acid can be transformed into allantoin by uricase, an enzyme that is not expressed by human cells but by many bacteria of the gut microbiota. Drawing created with the assistance of Macrovector Image and brgfx on Freepik.
The presence of uric acid in the human body emanates from a dynamic interplay of its biosynthesis and excretory processes. The liver is the primary site of uric acid production. About two-thirds of the body’s urate pool is generated endogenously from the breakdown of purines, with the remainder originating from dietary sources (Figure 1). Each day, a majority (approximately 70%) of the uric acid produced is expelled through the kidneys, while the remaining part is cleared into the biliary tract and subsequently transformed into allantoin by colonic bacterial uricase.
Diet is not the primary source of purine in the organism but can act as an adjustment factor in the case of excessive uric acid production. It is easier to limit the consumption of foods rich in uric acid or purine than to reduce hepatic metabolism. Foodstuffs rich in purine are listed in Table 1. Data were selected from [5,6]. Beers and wine are also considered to be purine-rich drinks. High-purine diets typically include red meats, certain seafood (like anchovies and sardines), and alcoholic beverages, particularly beer. Despite its long history, gout continues to pose significant socio-economic challenges and adversely affects patients’ quality of life [7] due to its increasing prevalence [8].

Table 1.
Purine content of selected foods and its correspondence to uric acid.
The basis of gout’s pathogenesis lies in the connections among genetic predisposition, environmental factors, and aberrations in purine metabolism [9], and most clinical instances of hyperuricemia are attributed to impaired renal elimination [4], but ablation of the microbiota may also cause severe hyperuricemia [10] and the subsequent precipitation of monosodium urate crystals in joints, tendons, and other tissues.
Although the kidneys are primarily responsible for uric acid elimination, a significant amount of uric acid is also eliminated from the human body through a process that involves the intestin and gut microbiota. Increasing the uric acid removal capabilities of the gut microbiota may provide an alternative and potentially preventive approach to decrease the incidence of gout episodes [11]. Additionally, the gut microbiota has been implicated in modulating immune responses that could exacerbate or alleviate inflammatory processes associated with gout [12].
Despite this progress, the detailed mechanisms through which the gut microbiota interacts with metabolic and immune pathways in gout remain underexplored. This review seeks to consolidate existing knowledge on the microbiota–gout interaction, focusing on how microbiota diversity and function can be leveraged to manage or potentially prevent gout through dietary and probiotic interventions.
Alterations in gut microbiota composition may influence uric acid metabolism and contribute to the pathogenesis of gout. Certain gut bacteria may modulate uric acid levels by affecting its production, excretion, or reabsorption in the intestines and kidneys [13]. Understanding the interplay between the microbiota and gout could pave the way for novel therapeutic approaches, such as probiotics or targeted microbiome interventions, to manage this disorder more effectively.
The human gut microbiota is a complex and dynamic ecosystem of microorganisms. This combination of bacteria, viruses, fungi, and archaea plays a pivotal role in maintaining homeostasis and influencing various aspects of host physiology [14].
Beyond contributing to digestion and nutrient metabolism, the microbiota actively participates in the regulation of immune homeostasis, maintenance of barrier functions, and synthesis of bioactive compounds. Perturbations in the composition and functionality of the microbiota, termed dysbiosis, have been associated with a spectrum of diseases, ranging from gastrointestinal disorders to autoimmune conditions, metabolic syndromes, neurological disorders [15], and arthritis.
The relationship between arthritis and the gut microbiota has attracted significant research interest, particularly in the context of autoimmune diseases such as rheumatoid arthritis (RA). Studies have shown that the gut microbiota composition differs notably between individuals with RA and healthy controls, indicating a potential role in the disease’s pathogenesis [16]. Dysbiosis, or an imbalance in the gut microbiome, has been linked to increased inflammation and immune system dysregulation, contributing to the development and progression of RA [17]. Specific bacteria, such as Prevotella copri, have been found in higher abundance in RA patients and are associated with the activation of inflammatory pathways [18]. Dysbiosis affects the gut’s barrier function, leading to systemic immune responses that exacerbate arthritis symptoms [19].
The composition of the gut microbiota can vary significantly among individuals, influenced by factors such as age, diet, genetics, environment, and health status. Despite this variability, certain microbial phyla are consistently found in the human gut. The major phyla include the following [20]:
- Firmicutes: Firmicutes are among the dominant bacterial phyla in the human gut. They include various genera, such as Clostridium, Lactobacillus, and Ruminococcus.
- Bacteroidetes: Bacteroidetes are another major bacterial phylum in the gut microbiota. Bacteroides is a prominent genus within this phylum.
- Actinobacteria: This phylum includes genera like Bifidobacterium, which are known for their beneficial roles in the gut, such as the fermentation of dietary fibers.
- Proteobacteria: This phylum consists of a diverse group of bacteria, including Escherichia coli (E. coli) and Helicobacter pylori.
- Verrucomicrobia: Although less abundant than Firmicutes and Bacteroidetes, Verrucomicrobia includes the genus Akkermansia, which has been associated with a healthy gut environment.
- Fusobacteria: This phylum is present in lower abundance and includes various species like Fusobacterium.
The gut microbiota is not limited to bacteria; it also includes viruses, fungi, archaea, and other microorganisms [21].
This review aims to consolidate current knowledge on the reciprocal interactions between the gut microbiota and gout. The microbiota, with its diversity, may possess the capacity to metabolize uric acid through both anabolic and catabolic pathways, potentially influencing uricemia regulation. Additionally, there is a potential for gout and its related inflammation to influence the composition of the microbiota. Through an examination of recent studies, clinical observations, and experimental findings, we aim to present a comprehensive overview of how changes in microbiota composition and function may impact gout in humans. Additionally, we will explore potential therapeutic approaches targeting the microbiota in the context of gout and/or hyperuricemia.
1.1. Database Search
This review utilized a comprehensive and systematic approach, employing the PRISMA (Preferred Reporting Items for Systematic Reviews and Meta-Analyses) strategy [22].
This literature search was conducted to address the review question “Does the gut microbiota interact with the onset and/or the progression of gout and conversely?” The review question was limited to human health. The bibliographic search was performed on both Web of Science and PubMed. We looked for “gout”, “microbiota”, “dysbiosis”, and “uric acid” in the abstracts. The search was limited to human studies. The search period was limited to 2014–2024. The bibliographic search specifically focused on articles written in English. The last entry dates for the databases were 1 February 2024 (PubMed) and 2 February 2024 (WoS). The process of selecting relevant publications involved two stages: (i) rapid assessment of relevance based on information in the titles and abstracts of the publications, allowing for the exclusion of obviously irrelevant publications, (ii) followed by a detailed assessment of the full-text document if required.
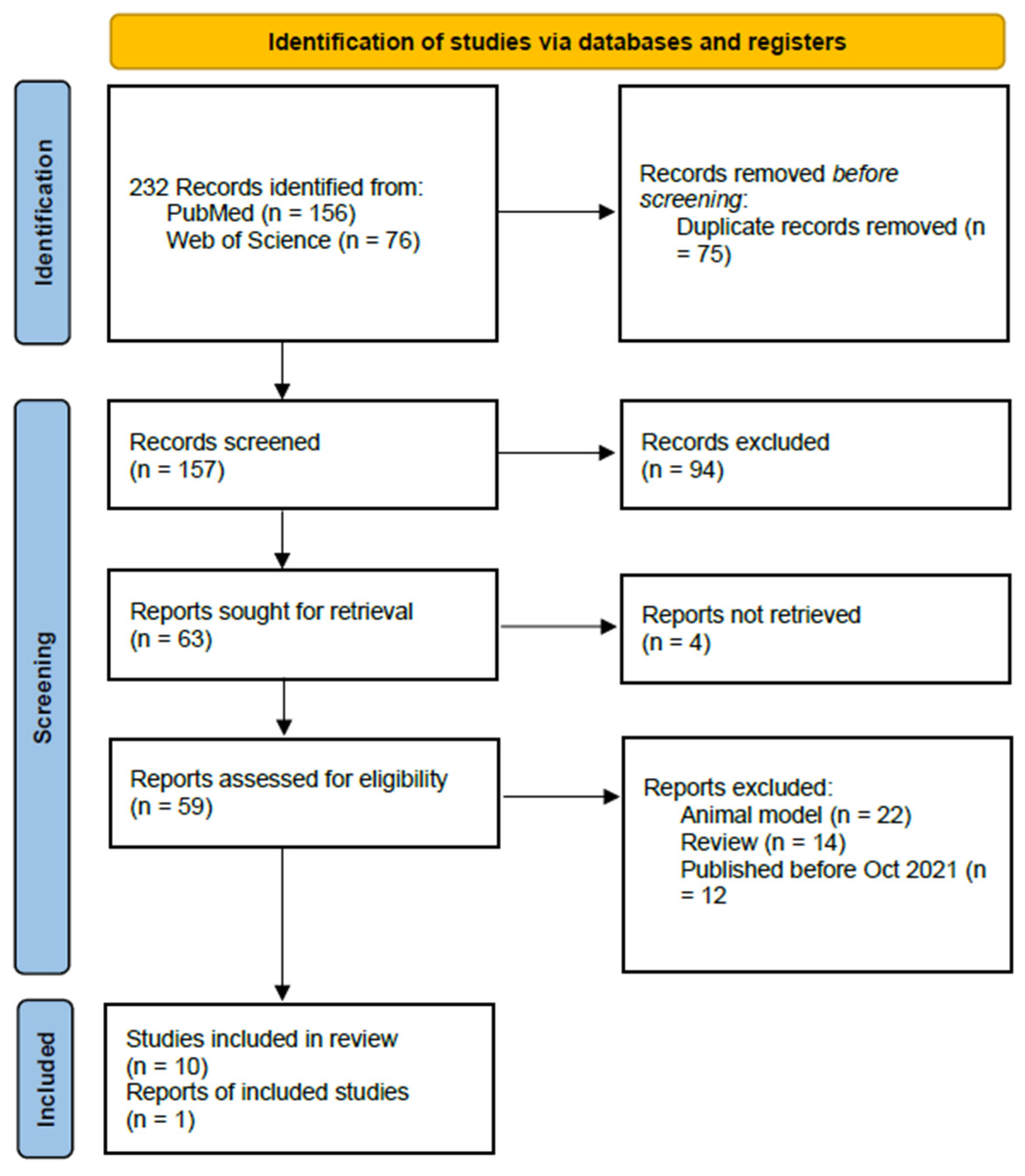
The selection process led to the identification of 232 publications. Among these, a systematic review was published in 2022 with a bibliographic review conducted from 2013 to October 2021. The current study analyzes the data from this review and provides an update with new information published since.
Employing this strategy, 156 references were retrieved from PubMed and 76 from Web of Science. Duplicate records were removed, resulting in a compilation of 157 articles. These publications underwent an initial screening process aimed at identifying relevant studies. As a result of this process, 59 publications were retained for further consideration. A subsequent round of screening excluded 22 articles that focused on animal models, particularly rodents, along with 14 that were narrative reviews and 12 that were published prior to 2021 and had been previously analyzed in the review by Shirmani-Rad [23] (during the period covered by this author, we did not identify any additional references). Our bibliographic research culminated in the selection and analysis of 10 articles presenting original data, published between October 2021 and the present (Figure 3).
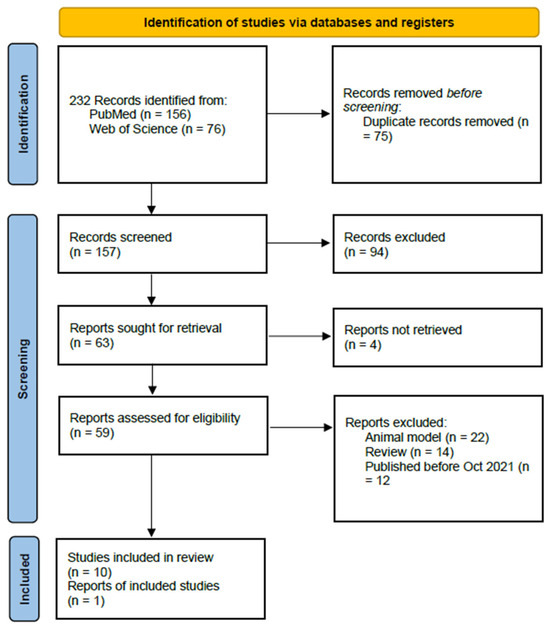
Figure 3.
PRISMA flow [22].
Although the Cochrane database was consulted for this research, it did not yield any new information.
To identify all published papers and limit any possible omissions that could introduce bias, we utilized a snowballing approach, which involves examining the references cited in relevant articles to discover additional sources.
1.2. Snowballing
Snowballing was used to identify additional articles. As a general rule, after assessing the eligibility of the returned articles, eligible ones can be uploaded [24]. We analyzed all the bibliographic references from these selected publications. This method did not yield any further publications.
2. Identification of Links between Gout and the Gut Microbiota
In this article, we compile data from both this systematic review and all papers published over the past three years that explore the interactions between gout and the gut microbiota. Given the proposal of fecal transplantation as a prospective treatment for the condition, our analysis also concentrates on data found in the literature within the same timeframe.
2.1. Analysis of Information Available until 2021
In the systematic review published by Shirvani-Rad in 2022 [23], the authors compiled the information coming from 15 studies (10 in humans).
This systematic review summarizes intestinal dysbiosis in gout to illustrate possible correlations. Among all of the studies, only five of them reported changes in alpha diversity and richness indices. Three studies showed lower richness in gout patients, which may be linked to intestinal dysbiosis and inflammation in gout. In contrast, two studies reported higher richness. These discrepancies could be related to differences between humans and animals, or to the disease state (asymptomatic hyperuricemia).
At the phylum level, the abundance of Bacteroidetes was found to be increased in the microbiota of gout patients in three studies and decreased in one study. The abundance of Firmicutes appeared to be increased in the gut microbiota of gout patients in three studies and decreased in two studies. A single study reported an Actinobacteria abundance decrease, while an increase in the abundance of these bacteria was reported in three studies. Additionally, Actinobacteria abundance increased in healthy controls in one study. Two studies reported increased Proteobacteria abundance in gout patients, and the opposite was reported in two studies. At the phylum level, the results of all of these studies exhibit contradictions and heterogeneities. Differences in population characteristics, gout severity, and methodologies could account for these contradictions.
At the genus level, an increase in Oscillibacter, Butyricicoccus, and Dialister was observed in the microbiota of gout patients in two studies. Increased abundance of Bacteroides in patients was reported in five studies, while the abundance of Roseburia and Alistipes was reported as increased in gout patients in three studies. Studies were contradictory regarding Faecalibacterium. Furthermore, two studies revealed that, at the genus level, the number of Bifidobacterium was reduced in gout patients.
In conclusion, this very complete systematic review outlines the intricate landscape of intestinal dysbiosis in gout, emphasizing variations in alpha diversity and richness indices across studies. The conflicting results regarding richness in gout patients may stem from differences in study populations or disease states. The authors concluded by suggesting that “further studies are needed to generate basic knowledge for clarifying possible underlying mechanisms and any probable causal relationship” [23].
2.2. Update since 2021
Since the completion of this review, 10 additional original papers have been published, offering new insights into the relationship between the gut microbiota and gout. In the forthcoming discussion, we will thoroughly examine the findings presented in these recent publications, aiming to broaden our understanding of the impact of intestinal dysbiosis on gout. We divide this update into three distinct parts: papers dealing with dysbiosis, clinical approaches, and fecal transplantation papers. These papers are presented in a chronological way in each sub-chapter. All of this information is summarized and compiled in Table 2.
Table 2.
Summary table of the original articles used for writing this review.
2.2.1. Dysbiosis and Gout
In the work published by Wei et al. in 2022 [25], the authors explored the link between the intestinal microbiota and hyperuricemia in humans. Utilizing data from two distinct population studies, the research began by assessing the relationship among intestinal microbiota imbalance, hyperuricemia, and serum urate levels in a rural Chinese population (termed the discovery cohort). This initial investigation was then corroborated through a subsequent study involving urban Chinese residents (referred to as the validation cohort). The entire study involved a total of over 1000 individuals.
Patients with hyperuricemia exhibited a moderately decreased richness and diversity of the microbiome. The authors of this study reported that a lower relative abundance of the genus Coprococcus was also strongly associated with higher SUA levels.
In both cohorts, the Kyoto Encyclopedia of Genes and Genomes (KEGG) metabolic pathways, including amino acid metabolisms, nucleotide metabolisms, and microbial functions related to replication, differed significantly between subjects with hyperuricemia and controls.
The major conclusion of this article is that microbiota alteration occurs in patients with gout. Furthermore, this study describes the beneficial effects that Coprococcus may have—an effect that seems to be mediated by an increased production of short-chain fatty acids (propionate and butyrate). The authors do not explicitly propose the use of this bacterial strain as a probiotic [25]; however, they should.
The study by Cao et al. (2022) explored the association between the intestinal microbiota and the biochemical characteristics of patients with uric-acid renal stones (UAS) in a Chinese population [26]. The overall goal of this study was to identify bacteria likely to affect the pathogenesis of uric-acid renal stones. This study was carried out on 117 patients with UAS, 123 patients with gout, and 135 healthy controls.
The characteristics of the intestinal microbiota were analyzed using 16S ribosomal RNA (rRNA) gene sequencing. The authors reported that the richness, diversity, and relative abundance of dominant bacteria at the phylum and genus levels in the intestinal microbiota of patients with UAS were significantly different from those of other subjects.
The abundance of Bacteroides and Fusobacterium was significantly positively correlated with the serum uric acid levels of UAS patients. In individuals forming uric-acid renal stones, the richness and diversity of the intestinal microbiota were different from those of gout patients and the control population: Bacteroides and Fusobacterium were positively correlated with the serum UA levels of UAS patients. These findings could represent a non-invasive target for the prevention and treatment of UAS, requiring further large-scale research.
The objective of the study by Wang et al. (2023) was to explore the causal relationship between the composition of the intestinal microbiota and gout [27]. To investigate the cause-and-effect connection between the makeup of the intestinal microbiota and the occurrence of gout, a two-sample Mendelian randomization (MR) approach was used. This study compiled over 500,000 patients from over 70 studies.
Mendelian randomization utilizes genetic variations as instrumental variables, allowing for the evaluation of causal connections between exposures and outcomes. From this study, several conclusions can be drawn.
The Actinobacteria phylum exhibits a negative correlation with serum uric acid levels, and intriguingly, elevated SUA levels may reciprocally increase the abundance of the Actinobacteria phylum. This suggests a potential negative feedback loop and regulatory mechanism between the Actinobacteria phylum and SUA levels. Additionally, both SUA levels and gout exert simultaneous influences on the abundance of the Faecalibacterium and Prevotella genera. Furthermore, the presence of Anaerotignum in the intestinal microbiota appears to confer a protective effect against gout, likely attributable to the production of butyric acid. Conversely, the presence in the gut microbiota of the Porphyromonadaceae family, the Melainabacteria, and the Ruminococcaceae UCG011 genus were identified as risk factors for gout.
This is a very well-conducted study, even if a remark can be made on Mendelian randomization in relation to pleiotropy. Pleiotropy occurs when a genetic variant used as an instrumental variable in MR influences multiple traits or pathways, not just the exposure of interest. This can lead to biased results in several ways, but this remark can essentially be made for any Mendelian randomization. This is just a remark, not a criticism.
2.2.2. Clinical Studies
Rodriguez and collaborators, in 2022, published the results of a study aiming to explore the ability of various Ligilactobacillus salivarius strains to metabolize purine-related metabolites, targeting the identification of a potential probiotic strain beneficial for individuals with a history of hyperuricemia [28]. The research was conducted in two phases: firstly, assessing the bacterial uptake of purine and selecting efficient strains, followed by a clinical trial using the chosen strain.
Among the 13 Ligilactobacillus salivarius strains initially analyzed, most showed a reduced ability to transport uric acid compared to inosine and guanosine, except for the MPac90 strain, which equally transported all three. Significantly, L. salivarius CECT 30632 outperformed others by completely converting inosine and guanosine (100%) and uric acid (50%), making it the candidate for clinical testing.
In the clinical trial, 30 hyperuricemic patients with recurrent gout were randomized to receive either L. salivarius CECT 30632 (109 CFU/day; 15 patients) or allopurinol (100 to 300 mg/day; 15 patients) over six months. The probiotic group, consuming L. salivarius CECT 30632, exhibited a notable reduction in gout episodes and better tolerance compared to the allopurinol group. Particularly, the probiotic intervention significantly lowered serum urate levels from an average of 9.04 mg/dL (range 8.72 to 9.36) to 7.90 mg/dL (range 7.58 to 8.22).
These research findings suggest that consumption of L. salivarius CECT 30632 is effective in lowering serum urate levels and reducing the incidence of gout episodes. This could lead to a decreased reliance on drug treatments for managing hyperuricemia and gout. Although the results are encouraging, the small number of participants in the study might limit the generalizability of these outcomes. Nevertheless, the study is significant because it highlights the potential role of probiotics in the management of gout, opening new opportunities for non-pharmacological interventions in treating this condition.
The work carried out by Chen et al. in 2022 [29] aimed at profiling the intestinal bacteriome, mycobiome, and virome in the context of gout. This study was carried out on 26 patients with gout and 28 healthy controls. Metagenomic sequencing of their stool samples was performed to characterize their microbiota. Profound alterations were observed in the intestinal bacteriome, mycobiome, and virome of gout patients. This analysis led to the identification of 1117 differentially abundant bacterial species, 23 fungal species, and 4115 viral operational taxonomic units. The co-abundance network revealed complex interactions among these multi-kingdom signatures, signifying their putative collective influence on the disease. Additionally, these microbial signatures demonstrated effective discrimination between patients and controls. The authors concluded that these differences might be considered as having diagnostic utility.
Kim’s article [30] aims to understand the differences in microbiota composition that may exist between patients with gout and those with asymptomatic hyperuricemia. The idea is ultimately to look for a therapeutic means to lower uric acid levels by interacting with the intestinal microbiota. In this study, fecal matter from 8 patients with asymptomatic hyperuricemia and over 30 patients with gout was analyzed. The gout patients were separated into two groups based on the severity of their disease. As with all such studies, some disparity exists even within a single group. Nonetheless, patients with gout showed a lower diversity than asymptomatic patients. The microbiota of gout patients showed a decreased Firmicutes/Bacteroidetes ratio and, conversely, an increased Prevotella/Bacteroides ratio. The authors of this publication concluded on different compositions between the microbiota of gout patients or asymptomatic patients and proposed a possible treatment for gout through modification of the intestinal microbiota. The authors also presented a summary table of bacteria promoting gout or, conversely, more present in asymptomatic patients. However, the absence of a real non-gout, non-hyperuricemic control group is detrimental. This article presents the results of a study that is relatively modest in quantity and presents results that confirm those already obtained in other studies. It is also noteworthy that this study was conducted only on men.
The goal of the paper published by Hou et al. in 2023 [31] was to investigate the interactive causal effects between the intestinal microbiota and host urate metabolism, and to explore the underlying mechanism.
They conducted large-scale bidirectional Mendelian randomization (MR) and bivariate linkage disequilibrium score regression to explore genetic causality and correlation between microbiota phenotypes and urate phenotypes.
In this paper, the authors reported that the intestinal microbiota and host urate metabolism had a bidirectional causal association, highlighting the critical role of crosstalk between host and microbiota, most notably in hyperuricemic patients. They also indicated that mediation analysis showed that the Bifidobacteriales order and Bifidobacteriaceae family had protective effects on urate levels. This effect seemed to be mediated by an increased level of docosahexaenoic acid, an omega-3 fatty acid.
The intestinal microbiota plays an essential role in the host’s production, catabolism, metabolism, and excretion of uric acid. It can either convert purines into uric acid by secreting active enzymes or accelerate uric acid degradation by synthesizing urate-metabolizing enzymes. One conclusion of this work could be that Bifidobacteria should be considered as a possible probiotic for treating hyperuricemia.
The paper by Martinez-Nava [32] evaluated the differences between taxonomic profiles by sequencing of the V3-V4 region of the 16S rRNA gene in the intestinal microbiota, and it predicted functional profiles of the intestinal microbiota in individuals with asymptomatic hyperuricemia and 162 people, including patients with gout.
The authors reported that no significant difference was observed in the α-diversity metrics among the study groups but, after adjusting the models with fixed effects, notable findings emerged. They found a significantly lower richness in individuals with asymptomatic hyperuricemia compared to both controls and gout patients. Furthermore, a significant difference in richness was observed between gout patients and controls.
The intestinal microbiota of individuals with asymptomatic hyperuricemia exhibited higher abundance of bacteria from the Proteobacteria phylum and bacteria from three Clostridia classes within the Firmicutes phylum.
The functional prediction of the gut microbiome indicated notable differences in pathway enrichment among the studied groups. Individuals with asymptomatic hyperuricemia, when compared to controls, showed an increase in bacterial proteins related to environmental information processing but a decrease in those associated with organismal systems and metabolism.
These results suggest that the intestinal microbiota of individuals with asymptomatic hyperuricemia produces more purines due to the absence of transcriptional repression of the pur operon, potentially leading to higher purine production than in healthy individuals. Comparing healthy subjects to gout patients did not reveal significant differences in the abundance of these proteins, indicating that anti-urate treatment could be involved in regulating purine production in the intestinal microbiota.
These results demonstrate that although some bacterial genera are common to both gout patients and individuals with arthritis, the latter exhibit less richness and more unique features in gout patients compared to controls.
The article from Ul-Haq et al. in 2022 [33] aimed to characterize differences in microbiota composition in relation to uric acid levels. In this study, 17 fecal samples from healthy individuals were compared with those of 48 gout patients treated with febuxostat. These 48 patients included 28 subjects with a decrease in uric acid levels and 20 subjects for whom uric acid levels were not controlled. The authors reported that the microbiota composition of the patient group with controlled uricemia was relatively close to that of healthy individuals. The authors also concluded that febuxostat allows for the restoration of microbiota composition in gout patients. This result indicates that, by modifying uricemia, it is possible to alter the microbiota composition. Among all observed differences, the presence of P. copri seems to be a marker for gout.
The authors suggested using seven bacteria as diagnostic markers for gout. It was also reported in this publication that there was a decrease in the levels of gamma Bifidobacteria in gout patients, a result that had already been published previously [34].
2.2.3. FMT and Gout
Fecal microbiota transplantation (FMT) involves the transfer of stool from a healthy donor to the gastrointestinal tract of a patient, with the aim of restoring a healthy microbial balance [35]. By altering the gut microbiota composition, FMT could help in reducing serum uric acid levels, thereby managing gout symptoms, such as by reducing inflammation, altering gut permeability, and limiting the impact on comorbidities. Currently, FMT is primarily used for treating Clostridium difficile infections and is being studied for other conditions [36].
Everyone’s gut microbiome is unique, and FMT may offer a personalized treatment approach. FMT has been recognized as a secure and effective approach to modulate gut dysbiosis. In 2022, Xie et al. conducted a study using WMT (washed microbiota transplantation) that induced a decrease in serum uric acid levels in gout patients, in accordance with findings on FMT’s effects in hyperuricemic mice [37]. WMT is an approach derived from FMT that, according to these authors, offers advantages in terms of safety and quality control compared to crude FMT. In this initial study, conducted with 11 male patients with gout, the objective was to assess the impacts of WMT on serum uric acid levels, symptoms, and the intestinal barrier function in individuals experiencing acute and recurrent gout.
In this preliminary investigation, they observed a reduction in serum uric acid levels and improved gout symptoms in individuals with acute and recurrent gout following washed microbiota transplantation (WMT) treatment. Additionally, there was an enhancement in the compromised intestinal barrier function after WMT.
However, it is important to note that the application of FMT (and WMT) in gout is still under investigation, and more research is needed to understand its efficacy and safety. The safety of FMT is still a real issue [38].
3. Conclusions
Through the exploration of the intricate relationship between gut microbiota composition and gout pathogenesis, it becomes evident that targeting the microbiota holds significant interest in ameliorating the burden of this debilitating disorder. By intervening at the microbial level, it might be possible to unlock novel strategies to modulate uric acid metabolism, mitigate inflammation and, ultimately, alleviate the symptoms and progression of gout.
Data across various studies, while not entirely consistent, generally support a link between the microbiota composition and the development or progression of gout. This consensus underscores the importance of microbiome modulation in managing gout’s symptoms and progression through dietary and bacterial interventions [28,29,30].
One can, however, note that there is a large heterogeneity in these studies: some use Mendelian randomization, others explore microbiota diversity, and others are clinical studies.
Prebiotics, mainly dietary fibers that nurture beneficial gut bacteria, and probiotics, comprising live beneficial bacteria, are central to these interventions [39]. They have shown potential in reducing serum urate levels and the frequency of gout episodes by modulating the gut microbiome.
The utility of probiotics, such as specific strains of Ligilactobacillus salivarius, has been demonstrated to effectively lower urate levels, a key factor in gout’s development [28]. This reduction not only alleviates the symptoms of gout but also potentially decreases the dependency on conventional urate-lowering drugs. By altering the gut microbiota composition, prebiotics can indirectly influence urate metabolism and excretion.
Fecal microbiota transplantation (FMT) for the treatment of gout is an innovative and emerging area of research that stems from the growing understanding of the gut microbiome’s role in health and disease. Given the intricate relationship between the gut microbiome and metabolic health, exploring FMT (or WMT [40]) as a treatment for gout is both fascinating and promising. However, it is crucial to approach this potential therapy with caution [41]. While altering the gut microbiome presents a novel avenue for managing gout, the scientific community must back it up with robust evidence from well-designed studies. The safety concerns and ethical implications of FMT also warrant careful consideration.
However, it is important to note that, while these findings are encouraging, they are still in the early stages. Most studies have limited participant numbers and are often focused on specific bacterial strains. Therefore, more extensive and diverse clinical trials are necessary to fully understand the efficacy and safety of prebiotics and probiotics in gout treatment.
Despite the promising potential of targeting the microbiota in gout treatment, there are several limitations to the current research that must be acknowledged. Firstly, much of the evidence linking gut microbiota modulation to reduced uric acid levels and improved gout outcomes comes from small-scale studies or those with a narrow focus on specific microbial strains. This limits the generalizability of the findings. Furthermore, the mechanisms by which the microbiota influences uric acid levels and gout are not fully understood, and the causal relationships remain to be clearly defined. Inconsistencies in data across studies highlight the need for a more standardized approach in microbiome research.
Future research in this area should prioritize larger, multicentric clinical trials that not only validate these initial findings but also assess the long-term effects and safety of interventions targeting the microbiome. Additionally, studies should broaden to include a wider array of prebiotics and probiotics, alongside other novel interventions such as fecal microbiota transplantation (FMT). FMT, in particular, presents a compelling method for gout management due to its potential for profound alterations in the microbiome. Nevertheless, the implementation of such treatments requires thorough consideration of safety and ethical issues, particularly because of the intimate nature and inherent risks associated with FMT [42].
The exploration of these strategies holds promise for opening new therapeutic pathways for gout, traditionally managed with pharmacological agents that many patients find insufficient or problematic. By advancing our understanding of the gut microbiome’s role in disease mechanisms, we can pave the way for more personalized and effective treatments for gout and other related metabolic disorders.
Funding
This research received no external funding.
Data Availability Statement
Both authors performed the literature search and the initial writing. Final revision was done by JD.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Dehlin, M.; Jacobsson, L.; Roddy, E. Global epidemiology of gout: Prevalence, incidence, treatment patterns and risk factors. Nat. Rev. Rheumatol. 2020, 16, 380–390. [Google Scholar] [CrossRef] [PubMed]
- Skinner, K.A.; Tan, S.; Parks, D.A. Uric Acid Metabolism. In Encyclopedia of Life Sciences; John Wiley & Sons: Hoboken, NJ, USA, 2001. [Google Scholar] [CrossRef]
- Demarquoy, J.; Fairand, A.; Gautier, C.; Vaillant, R. Regulation of argininosuccinate synthetase level by corticosteroid and pancreatic hormones during perinatal period. Mol. Cell Biochem. 1995, 143, 47–51. [Google Scholar] [CrossRef] [PubMed]
- Doherty, M. New insights into the epidemiology of gout. Rheumatology 2009, 48 (Suppl. S2), ii2–ii8. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, K.; Aoyagi, Y.; Fukuuchi, T.; Inazawa, K.; Yamaoka, N. Total purine and purine base content of common foodstuffs for facilitating nutritional therapy for gout and hyperuricemia. Biol. Pharm. Bull. 2014, 37, 709–721. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, K.; Takayanagi, F.; Fukuuchi, T.; Yamaoka, N.; Yasuda, M.; Mawatari, K.I.; Fujimori, S. Determination of total purine and purine base content of 80 food products to aid nutritional therapy for gout and hyperuricemia. Nucleosides Nucleotides Nucleic Acids 2020, 39, 1449–1457. [Google Scholar] [CrossRef]
- Flores, N.M.; Nuevo, J.; Klein, A.B.; Baumgartner, S.; Morlock, R. The economic burden of uncontrolled gout: How controlling gout reduces cost. J. Med. Econ. 2019, 22, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Helget, L.N.; Mikuls, T.R. Health disparities in gout. Curr. Opin. Rheumatol. 2024, 36, 120–125. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; You, C. The biomarkers discovery of hyperuricemia and gout: Proteomics and metabolomics. PeerJ 2023, 11, e14554. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Jarman, J.B.; Low, Y.S.; Augustijn, H.E.; Huang, S.; Chen, H.; DeFeo, M.E.; Sekiba, K.; Hou, B.H.; Meng, X.; et al. A widely distributed gene cluster compensates for uricase loss in hominids. Cell 2023, 186, 4472–4473. [Google Scholar] [CrossRef]
- Tong, S.; Zhang, P.; Cheng, Q.; Chen, M.; Chen, X.; Wang, Z.; Lu, X.; Wu, H. The role of gut microbiota in gout: Is gut microbiota a potential target for gout treatment. Front. Cell Infect. Microbiol. 2022, 12, 1051682. [Google Scholar] [CrossRef]
- Rosenbaum, J.T.; Asquith, M.J. The Microbiome: A Revolution in Treatment for Rheumatic Diseases? Curr. Rheumatol. Rep. 2016, 18, 62. [Google Scholar] [CrossRef] [PubMed]
- Chu, Y.; Sun, S.; Huang, Y.; Gao, Q.; Xie, X.; Wang, P.; Li, J.; Liang, L.; He, X.; Jiang, Y.; et al. Metagenomic analysis revealed the potential role of gut microbiome in gout. NPJ Biofilms Microbiomes 2021, 7, 66. [Google Scholar] [CrossRef] [PubMed]
- Gonzales-Luna, A.J.; Carlson, T.J.; Garey, K.W. Gut microbiota changes associated with. Gut Microbes 2023, 15, 2223345. [Google Scholar] [CrossRef] [PubMed]
- Demarquoy, J.; Othman, H.; Demarquoy, C. Modify gut microbiome in autism: A promising strategy? Explor. Neurosci. 2023; in press. [Google Scholar] [CrossRef] [PubMed]
- Horta-Baas, G.; Romero-Figueroa, M.D.S.; Montiel-Jarquín, A.J.; Pizano-Zárate, M.L.; García-Mena, J.; Ramírez-Durán, N. Intestinal Dysbiosis and Rheumatoid Arthritis: A Link between Gut Microbiota and the Pathogenesis of Rheumatoid Arthritis. J. Immunol. Res. 2017, 2017, 4835189. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Wei, Y.; Zhu, Y.; Xie, Z.; Hai, Q.; Li, Z.; Qin, D. Gut microbiota and rheumatoid arthritis: From pathogenesis to novel therapeutic opportunities. Front. Immunol. 2022, 13, 1007165. [Google Scholar] [CrossRef] [PubMed]
- Pianta, A.; Arvikar, S.; Strle, K.; Drouin, E.E.; Wang, Q.; Costello, C.E.; Steere, A.C. Evidence of the Immune Relevance of Prevotella copri, a Gut Microbe, in Patients With Rheumatoid Arthritis. Arthritis Rheumatol. 2017, 69, 964–975. [Google Scholar] [CrossRef] [PubMed]
- Teng, F.; Klinger, C.N.; Felix, K.M.; Bradley, C.P.; Wu, E.; Tran, N.L.; Umesaki, Y.; Wu, H.J. Gut Microbiota Drive Autoimmune Arthritis by Promoting Differentiation and Migration of Peyer’s Patch T Follicular Helper Cells. Immunity 2016, 44, 875–888. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Yan, Y.; Webb, R.J.; Li, Y.; Mehrabani, S.; Xin, B.; Sun, X.; Wang, Y.; Mazidi, M. Psychological Stress and Gut Microbiota Composition: A Systematic Review of Human Studies. Neuropsychobiology 2023, 82, 247–262. [Google Scholar] [CrossRef]
- Perler, B.K.; Friedman, E.S.; Wu, G.D. The Role of the Gut Microbiota in the Relationship Between Diet and Human Health. Annu. Rev. Physiol. 2023, 85, 449–468. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. Syst. Rev. 2021, 10, 89. [Google Scholar] [CrossRef]
- Shirvani-Rad, S.; Khatibzade-Nasari, N.; Ejtahed, H.S.; Larijani, B. Exploring the role of gut microbiota dysbiosis in gout pathogenesis: A systematic review. Front. Med. 2023, 10, 1163778. [Google Scholar] [CrossRef]
- Wohlin, C. Guidelines for snowballing in systematic literature studies and a replication in software engineering. In Proceedings of the 18th International Conference on Evaluation and Assessment in Software Engineering (EASE’14), London, UK, 12–14 May 2014; Association for Computing Machinery: New York, NY, USA, 2014; pp. 1–10. [Google Scholar]
- Wei, J.; Zhang, Y.; Dalbeth, N.; Terkeltaub, R.; Yang, T.; Wang, Y.; Yang, Z.; Li, J.; Wu, Z.; Zeng, C.; et al. Association Between Gut Microbiota and Elevated Serum Urate in Two Independent Cohorts. Arthritis Rheumatol. 2022, 74, 682–691. [Google Scholar] [CrossRef]
- Cao, C.; Jin, X.; Ding, Q.; Zhu, J.; Yang, D.; Fan, B. The altered composition of gut microbiota and biochemical features as well as dietary patterns in a southern Chinese population with recurrent renal calcium oxalate stones. Urolithiasis 2023, 51, 95. [Google Scholar] [CrossRef]
- Wang, M.; Fan, J.; Huang, Z.; Zhou, D.; Wang, X. Causal Relationship between Gut Microbiota and Gout: A Two-Sample Mendelian Randomization Study. Nutrients 2023, 15, 4260. [Google Scholar] [CrossRef]
- Rodríguez, J.M.; Garranzo, M.; Segura, J.; Orgaz, B.; Arroyo, R.; Alba, C.; Beltrán, D.; Fernández, L. A randomized pilot trial assessing the reduction of gout episodes in hyperuricemic patients by oral administration of. Front. Microbiol. 2023, 14, 1111652. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Zhang, Y.; Yao, X.; Yan, Q.; Li, S.; Zhong, Q.; Liu, Z.; Tang, F.; Liu, C.; Li, H.; et al. Characterizations of the multi-kingdom gut microbiota in Chinese patients with gouty arthritis. BMC Microbiol. 2023, 23, 363. [Google Scholar] [CrossRef]
- Kim, H.W.; Yoon, E.J.; Jeong, S.H.; Park, M.C. Distinct Gut Microbiota in Patients with Asymptomatic Hyperuricemia: A Potential Protector against Gout Development. Yonsei Med. J. 2022, 63, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Hou, T.; Dai, H.; Wang, Q.; Hou, Y.; Zhang, X.; Lin, H.; Wang, S.; Li, M.; Zhao, Z.; Lu, J.; et al. Dissecting the causal effect between gut microbiota, DHA, and urate metabolism: A large-scale bidirectional Mendelian randomization. Front. Immunol. 2023, 14, 1148591. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Nava, G.A.; Méndez-Salazar, E.O.; Vázquez-Mellado, J.; Zamudio-Cuevas, Y.; Francisco-Balderas, A.; Martínez-Flores, K.; Fernández-Torres, J.; Lozada-Pérez, C.; Guido-Gómora, D.L.; Martínez-Gómez, L.E.; et al. The impact of short-chain fatty acid-producing bacteria of the gut microbiota in hyperuricemia and gout diagnosis. Clin. Rheumatol. 2023, 42, 203–214. [Google Scholar] [CrossRef]
- Ul-Haq, A.; Lee, K.A.; Seo, H.; Kim, S.; Jo, S.; Ko, K.M.; Moon, S.J.; Kim, Y.S.; Choi, J.R.; Song, H.Y.; et al. Characteristic alterations of gut microbiota in uncontrolled gout. J. Microbiol. 2022, 60, 1178–1190. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Zhang, J.; Wang, Z.; Ang, K.Y.; Huang, S.; Hou, Q.; Su, X.; Qiao, J.; Zheng, Y.; Wang, L.; et al. Intestinal Microbiota Distinguish Gout Patients from Healthy Humans. Sci. Rep. 2016, 6, 20602. [Google Scholar] [CrossRef] [PubMed]
- Airola, C.; Severino, A.; Porcari, S.; Fusco, W.; Mullish, B.H.; Gasbarrini, A.; Cammarota, G.; Ponziani, F.R.; Ianiro, G. Future Modulation of Gut Microbiota: From Eubiotics to FMT, Engineered Bacteria, and Phage Therapy. Antibiotics 2023, 12, 868. [Google Scholar] [CrossRef] [PubMed]
- Servetas, S.L.; Daschner, P.J.; Guyard, C.; Thomas, V.; Affagard, H.; Sergaki, C.; Sokol, H.; Wargo, J.A.; Wu, G.D.; Sabot, P. Evolution of FMT—From early clinical to standardized treatments. Biologicals 2022, 76, 31–35. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Tang, S.; Han, J.; Fan, S.; Huang, Y.; Zhang, Z.; Zhou, J.; Ming, T.; Li, Y.; Su, X. Apostichopus japonicus Oligopeptide Induced Heterogeneity in the Gastrointestinal Tract Microbiota and Alleviated Hyperuricemia in a Microbiota-Dependent Manner. Mol. Nutr. Food Res. 2021, 65, e2100147. [Google Scholar] [CrossRef]
- Vendrik, K.E.; Chernova, V.O.; Kuijper, E.J.; Terveer, E.M.; van Hilten, J.J.; Contarino, M.F.; FMT4PD study group. Safety and feasibility of faecal microbiota transplantation for patients with Parkinson’s disease: A protocol for a self-controlled interventional donor-FMT pilot study. BMJ Open 2023, 13, e071766. [Google Scholar] [CrossRef] [PubMed]
- Sanlier, N.; Kocabas, Ş. The effect of probiotic, prebiotic and gut microbiota on ASD: A review and future perspectives. Crit. Rev. Food Sci. Nutr. 2023, 63, 2319–2330. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.R.; Yang, X.Y.; Deng, Z.H.; Zheng, Y.M.; Zhang, R.; Wu, L.H.; Cai, J.Y.; Kong, L.P.; Xia, H.H.; He, X.X. Effects of Washed Microbiota Transplantation on Serum Uric Acid Levels, Symptoms, and Intestinal Barrier Function in Patients with Acute and Recurrent Gout: A Pilot Study. Dig. Dis. 2022, 40, 684–690. [Google Scholar] [CrossRef] [PubMed]
- Bhutiani, N.; Schucht, J.E.; Miller, K.R.; McClave, S.A. Technical Aspects of Fecal Microbial Transplantation (FMT). Curr. Gastroenterol. Rep. 2018, 20, 30. [Google Scholar] [CrossRef]
- Qu, Z.; Tian, P.; Yang, B.; Zhao, J.; Wang, G.; Chen, W. Fecal microbiota transplantation for diseases: Therapeutic potential, methodology, risk management in clinical practice. Life Sci. 2022, 304, 120719. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).