
Genetic Engineering of Filamentous Fungi: Prospects for Obtaining Fourth-Generation Biological Products

 ,
,  , , and
, , and
Abstract
1. Introduction
2. The Generations of Bio-Input Technology
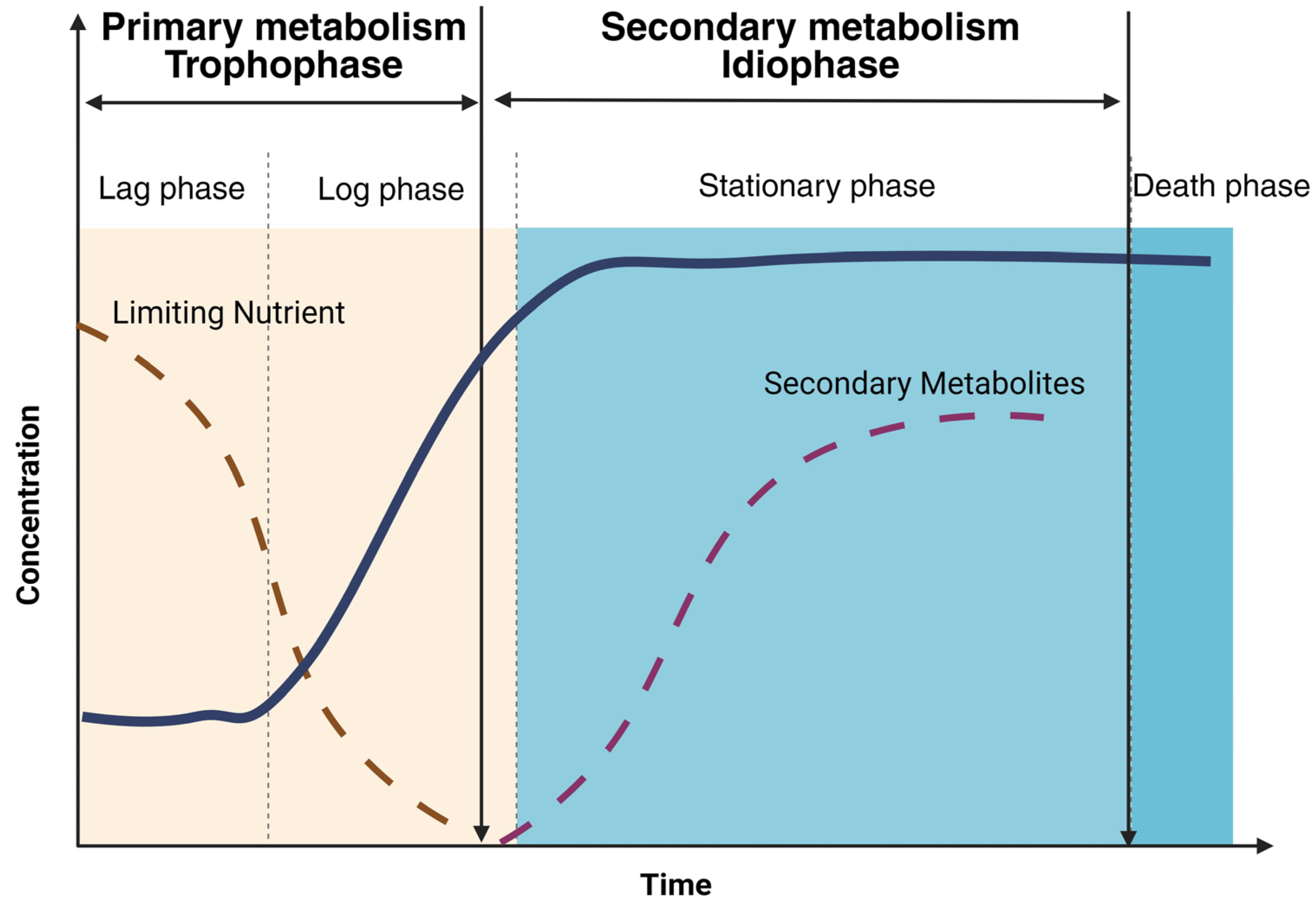
3. Secondary Metabolites from Filamentous fungi
4. Tools for the Genetic Transformation of Filamentous fungi

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Method | Principle | Advantages | Disadvantages | Reference |
|---|---|---|---|---|
| TMP | Use of cell wall degrading enzymes to get protoplasts. DNA transfer occurs by adding PEG and CaCl2. | Spores, spores in the process of germination, and hyphae are usable. | The transformation rate depends on the efficiency of the lytic enzyme used to get the protoplasts. Requires cell regeneration procedure. High number of DNA copies is inserted | [66,67] |
| TMA | A. tumefaciens carries two vectors (the binary vector with the DNA of interest between the left and right repeats and the T vector containing the virulence region important for DNA transfer). During the co-cultivation of A. tumefaciens with the fungus, they achieve DNA transfer. | Spores, spores in the process of germination, and hyphae are usable. Low copy number of inserted DNA improves targeted integration. | Several parameters during the co-cultivation of A. tumefaciens and the filamentous fungus to be transformed affect the transformation rate. Time-consuming technique | [68] |
| EP | DNA transfer is mediated by reversible permeabilization of the membrane induced by local application of electrical pulses. | Spores, spores in the process of germination, and hyphae are usable. A simple and inexpensive technique | The formation of protoplasts is often necessary to perform the technique | [69] |
| TB | The DNA is coated with tungsten or gold and inserted into the cell to be transformed using a microparticle accelerator. | No pre-treatment of recipient cells | Special equipment is necessary. | [70] |
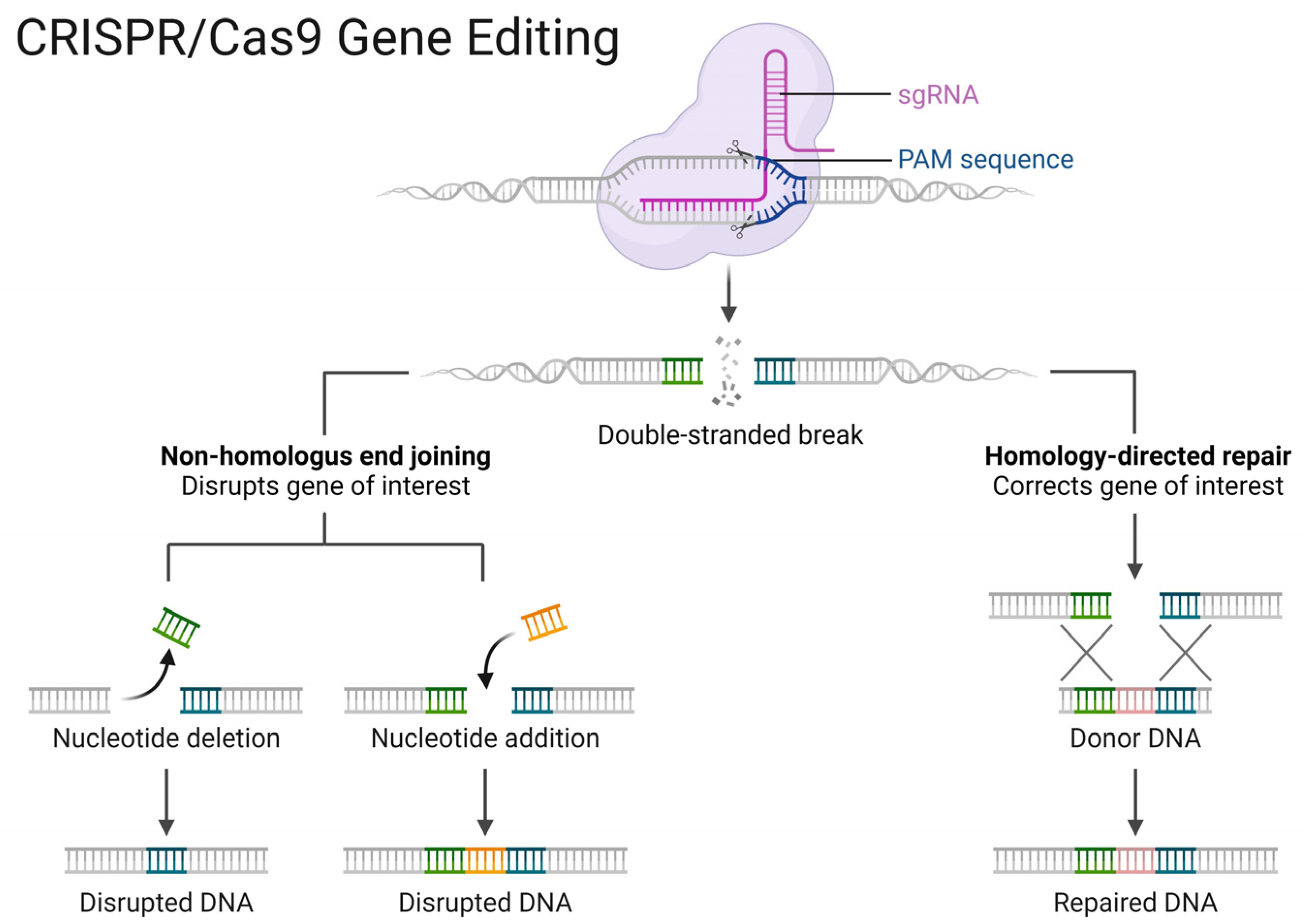
| CRISPR (RNPs) | Cas9 and guide RNAs can be delivered in form of DNA, RNA/mRNA, or ribonucleoprotein (RNP). The delivery methods are usually divided into physical (electroporation and microinjection), viral (lentiviral, adenoviral, and AAV vectors), and non-viral (plasmids, lipid and polymeric nanoparticles, and extracellular vesicles) ones. | The delivery of CRISPR ribonucleoproteins (RNPs) for genome editing in vitro and in vivo has important advantages over other delivery methods, including reduced off-target and immunogenic effects | Effective delivery of RNPs remains challenging in certain cell types due to low efficiency and cell toxicity | [71,72,73] |
5. Protein Secretion Pathways in Filamentous fungi
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Kumar, A.; Chadha, S.; Rath, D. Chapter 17—CRISPR-Cas9 system for functional genomics of filamentous fungi: Applications and challenges. In Fungi Bio-Prospects in Sustainable Agriculture, Environment and Nano-Technology; Sharma, V.K., Shah, M.P., Parmar, S., Kumar, A., Eds.; Academic Press: Cambridge, MA, USA, 2021; pp. 541–576. [Google Scholar] [CrossRef]
- Nevalainen, H.; Peterson, R. Making recombinant proteins in filamentous fungi-are we expecting too much? Front. Microbiol. 2014, 5, 75. [Google Scholar] [CrossRef] [PubMed]
- Demain, A.L. Small bugs, big business: The economic power of the microbe. Biotechnol. Adv. 2000, 18, 499–514. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Zhong, C.; Xiao, H. Genetic Engineering of Filamentous Fungi for Efficient Protein Expression and Secretion. Front. Bioeng. Biotechnol. 2020, 8, 293. [Google Scholar] [CrossRef] [PubMed]
- Brasil. Instrução Normativa Conjunta SDA/SDC nº 2, de 12 de Julho de 2013. Estabelece as Especificações de Referência de Produtos Fitossanitários com uso Aprovado para a Agricultura Orgânica; União, D.O.D., Ed.; 2013; pp. 6–8. Available online: https://www.gov.br/agricultura/pt-br/assuntos/insumos-agropecuarios/insumos-agricolas/agrotoxicos/produtos-fitossanitarios/arquivos-especificacao-de-referencia/in-conjunta-sda-sdc-no-2-de-12-de-julho-de-2013.pdf/view (accessed on 16 January 2024).
- Quintela, E.D. Lalguard Java: Produto Biológico a Base Do Fungo Cordyceps Javanica Para o Controle de Pragas—Parceria Embrapa e Lallemand Plant Care; Embrapa Rice and Beans: Santo Antônio de Goiás, Brasil, 2022. Available online: https://www.embrapa.br/busca-de-solucoes-tecnologicas/-/produto-servico/11043/bioinseticida-a-base-de-cordyceps-javanica-para-controle-biologico-de-mosca-branca (accessed on 1 May 2024).
- Corrêa, B.; da Silveira Duarte, V.; Silva, D.M.; Mascarin, G.M.; Júnior, I.D. Comparative analysis of blastospore production and virulence of Beauveria bassiana and Cordyceps fumosorosea against soybean pests. BioControl 2020, 65, 323–337. [Google Scholar] [CrossRef]
- Woo, S.L.; Hermosa, R.; Lorito, M.; Monte, E. Trichoderma: A multipurpose, plant-beneficial microorganism for eco-sustainable agriculture. Nat. Rev. Microbiol. 2023, 21, 312–326. [Google Scholar] [CrossRef] [PubMed]
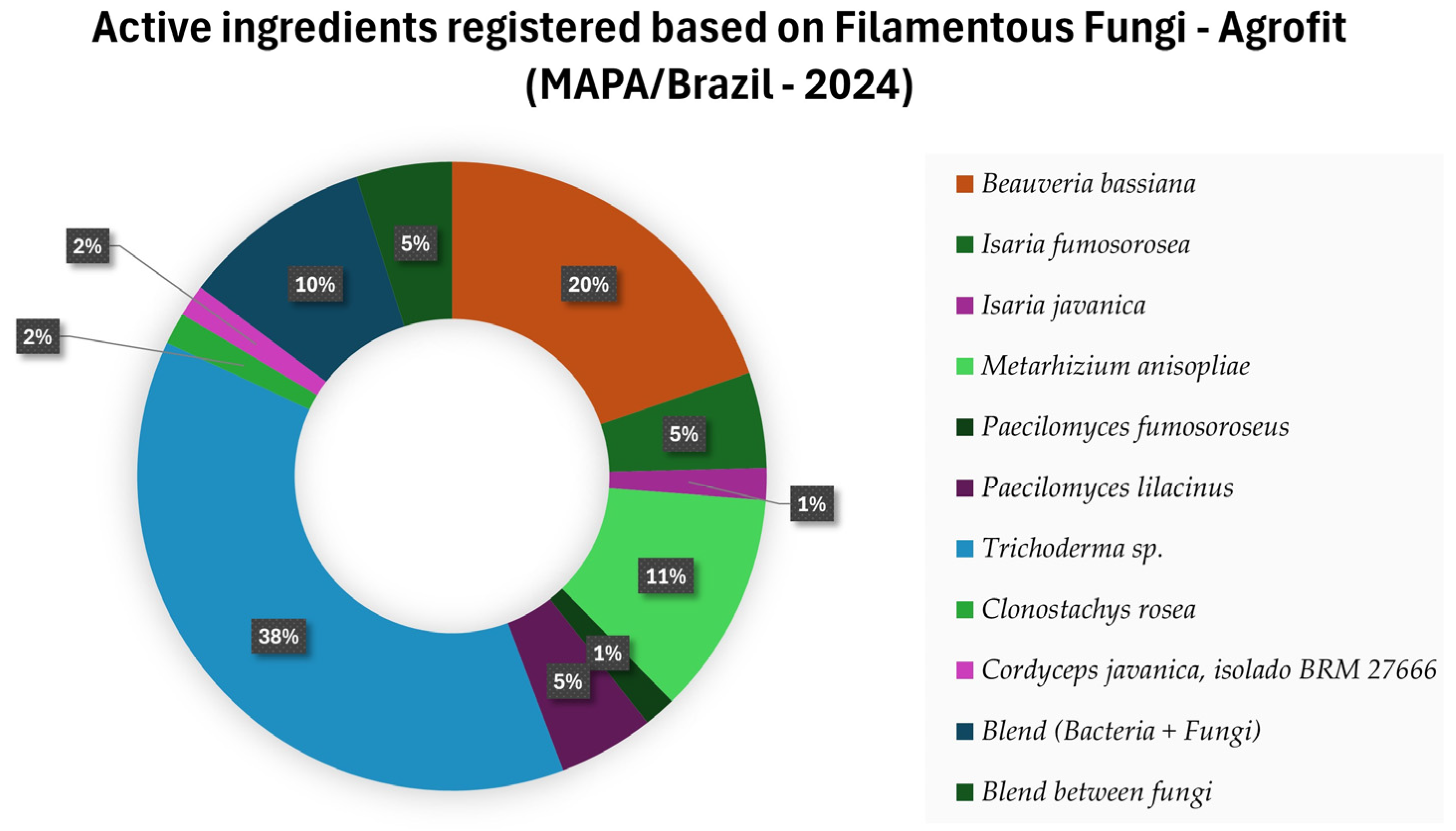
- MAPA. Consulta de Produtos Formulados. 2024. Available online: https://agrofit.agricultura.gov.br/agrofit_cons/principal_agrofit_cons (accessed on 3 February 2024).
- Joop, G.; Vilcinskas, A. Coevolution of parasitic fungi and insect hosts. Zoology 2016, 119, 350–358. [Google Scholar] [CrossRef] [PubMed]
- Molnár, I.; Gibson, D.M.; Krasnoff, S.B. Secondary metabolites from entomopathogenic Hypocrealean fungi. Nat. Prod. Rep. 2010, 27, 1241–1275. [Google Scholar] [CrossRef]
- Rohlfs, M.; Churchill, A.C. Fungal secondary metabolites as modulators of interactions with insects and other arthropods. Fungal Genet. Biol. 2011, 48, 23–34. [Google Scholar] [CrossRef]
- Wang, H.; Peng, H.; Li, W.; Cheng, P.; Gong, M. The Toxins of Beauveria bassiana and the Strategies to Improve Their Virulence to Insects. Front. Microbiol. 2021, 12, 705343. [Google Scholar] [CrossRef]
- Rukachaisirikul, V.; Chantaruk, S.; Tansakul, C.; Saithong, S.; Chaicharernwimonkoon, L.; Pakawatchai, C.; Isaka, M.; Intereya, K. A cyclopeptide from the Insect pathogenic fungus Cordyceps sp. BCC 1788. J. Nat. Prod. 2006, 69, 305–307. [Google Scholar] [CrossRef]
- Kumar, S.; Dahiya, R.; Khokra, S.L.; Mourya, R.; Chennupati, S.V.; Maharaj, S. Total Synthesis and Pharmacological Investigation of Cordyheptapeptide A. Molecules 2017, 22, 682. [Google Scholar] [CrossRef]
- Meyer, V. Genetic engineering of filamentous fungi—Progress, obstacles and future trends. Biotechnol. Adv. 2008, 26, 177–185. [Google Scholar] [CrossRef]
- Vanegas, K.G.; Rendsvig, J.K.H.; Jarczynska, Z.D.; Cortes, M.V.D.C.B.; van Esch, A.P.; Morera-Gómez, M.; Contesini, F.J.; Mortensen, U.H. A Mad7 System for Genetic Engineering of Filamentous Fungi. J. Fungi 2023, 9, 16. [Google Scholar] [CrossRef]
- Unkles, S.E.; Valiante, V.; Mattern, D.J.; Brakhage, A.A. Synthetic biology tools for bioprospecting of natural products in eukaryotes. Chem. Biol. 2014, 21, 502–508. [Google Scholar] [CrossRef]
- Schuetze, T.; Meyer, V. Polycistronic gene expression in Aspergillus niger. Microb. Cell Factories 2017, 16, 162. [Google Scholar] [CrossRef]
- Geib, E.; Brock, M. ATNT: An enhanced system for expression of polycistronic secondary metabolite gene clusters in Aspergillus niger. Fungal Biol. Biotechnol. 2017, 4, 13. [Google Scholar] [CrossRef]
- Mattern, D.J.; Valiante, V.; Horn, F.; Petzke, L.; Brakhage, A.A. Rewiring of the Austinoid Biosynthetic Pathway in Filamentous Fungi. ACS Chem. Biol. 2017, 12, 2927–2933. [Google Scholar] [CrossRef]
- MAPA. Bio-Inputs Enhance Sustainability of Agricultural Production in Brazil. 2024. Available online: https://www.gov.br/agricultura/pt-br/assuntos/inovacao/bioinsumos/material-para-imprensa/en/bio-inputs-enhance-sustainability-of-agricultural-production-in-brazil.pdf (accessed on 18 February 2024).
- Ziolkowska, J.R. Chapter 1—Biofuels Technologies. In Biofuels for a More Sustainable Future; Ren, J., Scipioni, A., Manzardo, A., Liang, H., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 1–19. [Google Scholar] [CrossRef]
- Brasil. Projeto de Lei nº 3668, 2021. Dispõe Sobre a Produção, o Registro, Comercialização, uso, Destino dos Resíduos e Embalagens, o Registro, Inspeção e Fiscalização, a Pesquisa e Experimentação, e os Incentivos à Produção de Bioinsumos para Agricultura e dá Outras Providências. Deputados, C.D., Ed.; Brasília, 2021. Available online: https://www25.senado.leg.br/web/atividade/materias/-/materia/150351 (accessed on 16 January 2024).
- Brasil. Projeto de Lei nº 658: Dispõe Sobre a Classificação, Tratamento e Produção de Bioinsumos por meio do Manejo Biológico on Farm; Ratifica o Programa Nacional de Bioinsumos e dá Outras Providências. Brasilia, 2021. Available online: https://www.camara.leg.br/propostas-legislativas/2271161 (accessed on 16 January 2024).
- Brasil. Portaria Conjunta SDA/MAPA—IBAMA—ANVISA n.º 1. Estabelece Procedimentos a Serem Adotados para o Registro de Produtos Microbiológicos Empregados no Controle de Pragas ou como Desfolhantes, Dessecantes, Estimuladores, Inibidores de Crescimento, Além de Revogar os atos Normativos Vigentes, Pertinentes à esta Matéria: Instrução Normativa Conjunta Ministério da Agricultura Pecuária e Abastecimento/Anvisa/Ibama nº 03 de 10 de Março de 2006 e o Ato CGAA/DSV/SDA nº 06. Brasil, D.O.d.R.F.d., Ed.; Brasília-DF, 2023; Vol. n.º 84. Available online: http://pesquisa.in.gov.br/imprensa/jsp/visualiza/index.jsp?data=04/05/2023&jornal=515&pagina=7 (accessed on 17 February 2024).
- Ajanovic, A. Biofuels versus food production: Does biofuels production increase food prices? Energy 2021, 36, 2070–2076. [Google Scholar] [CrossRef]
- Harvey, M.; Pilgrim, S. The new competition for land: Food, energy, and climate change. Food Policy 2011, 36, S40–S51. [Google Scholar] [CrossRef]
- Méndez-González, F.; Loera-Corral, O.; Saucedo-Castañeda, G.; Favela-Torres, E. Chapter 7—Bioreactors for the Production of Biological Control Agents Produced by Solid-State Fermentation. In Current Developments in Biotechnology and Bioengineering; Pandey, A., Larroche, C., Soccol, C.R., Eds.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 109–121. [Google Scholar] [CrossRef]
- Faria, M.R.D.; Wraight, S.P. Mycoinsecticides and Mycoacaricides: A comprehensive list with worldwide coverage and international classification of formulation types. Biol. Control 2007, 43, 237–256. [Google Scholar] [CrossRef]
- Li, Z.; Alves, S.B.; Roberts, D.W.; Fan, M.; Delalibera, I., Jr.; Tang, J.; Lopes, R.B.; Faria, M.; Rangel, D.E.N. Biological control of insects in Brazil and China: History, current programs and reasons for their successes using entomopathogenic fungi. Biocontrol Sci. Technol. 2010, 20, 117–136. [Google Scholar] [CrossRef]
- Kirst, H.A. The spinosyn family of insecticides: Realizing the potential of natural products research. J. Antibiot. 2010, 63, 101–111. [Google Scholar] [CrossRef]
- Cortes, M.V.D.C.B.; Barbosa, E.T.; Oliveira, M.I.D.S.; Maciel, L.H.R.; Lobo Junior, M.; Contesini, F.J.; Filippi, M.C.C.D.; Silva-Lobo, V.L.D. Trichoderma harzianum marker-free strain construction based on efficient CRISPR/Cas9 recyclable system: A helpful tool for the study of biological control agents. Biol. Control 2023, 184, 105281. [Google Scholar] [CrossRef]
- Schueffler, A.; Anke, T. Fungal natural products in research and development. Nat. Prod. Rep. 2014, 31, 1425–1448. [Google Scholar] [CrossRef]
- Bérdy, J. Bioactive Microbial Metabolites. J. Antibiot. 2005, 58, 1–26. [Google Scholar] [CrossRef]
- Liaud, N.; Giniés, C.; Navarro, D.; Fabre, N.; Crapart, S.; Gimbert, I.H.; Levasseur, A.; Raouche, S.; Sigoillot, J.-C. Exploring fungal biodiversity: Organic acid production by 66 strains of filamentous fungi. Fungal Biol. Biotechnol. 2014, 1, 1. [Google Scholar] [CrossRef]
- Wösten, H.A.B. Filamentous fungi for the production of enzymes, chemicals and materials. Curr. Opin. Biotechnol. 2019, 59, 65–70. [Google Scholar] [CrossRef]
- Awad, H.M.; EL-Shahed, K.Y.I.; Aziz, R.; Sarmidi, M.R.; El-Enshasy, H.A. Antibiotics as Microbial Secondary Metabolites: Production and Application. J. Teknol. 2012, 59, 101–111. [Google Scholar] [CrossRef]
- Trail, F.; Mahanti, N.; Rarick, M.; Mehigh, R.; Liang, S.H.; Zhou, R.; Linz, J.E. Physical and transcriptional map of an aflatoxin gene cluster in Aspergillus parasiticus and functional disruption of a gene involved early in the aflatoxin pathway. Appl. Environ. Microbiol. 1995, 61, 2665–2673. [Google Scholar] [CrossRef]
- Lysøe, E.; Seong, K.Y.; Kistler, H.C. The transcriptome of Fusarium graminearum during the infection of wheat. Mol. Plant Microbe Interact. 2011, 24, 995–1000. [Google Scholar] [CrossRef]
- Lind, A.L.; Lim, F.Y.; Soukup, A.A.; Keller, N.P.; Rokas, A. An LaeA- and BrlA-Dependent Cellular Network Governs Tissue-Specific Secondary Metabolism in the Human Pathogen Aspergillus fumigatus. mSphere 2018, 3, 10–1128. [Google Scholar] [CrossRef]
- Spraker, J.E.; Wiemann, P.; Baccile, J.A.; Venkatesh, N.; Schumacher, J.; Schroeder, F.C.; Sanchez, L.M.; Keller, N.P. Conserved Responses in a War of Small Molecules between a Plant-Pathogenic Bacterium and Fungi. mBio 2018, 9, 1–14. [Google Scholar] [CrossRef]
- Tkacz, J.S.; Lange, L. Advances in Fungal Biotechnology for Industry, Agriculture, and Medicine; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2004. [Google Scholar]
- Keller, N.P. Fungal secondary metabolism: Regulation, function and drug discovery. Nat. Rev. Microbiol. 2019, 17, 167–180. [Google Scholar] [CrossRef]
- Nielsen, J.C.; Prigent, S.; Grijseels, S.; Workman, M.; Ji, B.; Nielsen, J. Comparative Transcriptome Analysis Shows Conserved Metabolic Regulation during Production of Secondary Metabolites in Filamentous Fungi. mSystems 2019, 4, 1–14. [Google Scholar] [CrossRef]
- Mosunova, O.V.; Navarro-Muñoz, J.C.; Collemare, J. The Biosynthesis of Fungal Secondary Metabolites: From Fundamentals to Biotechnological Applications. In Encyclopedia of Mycology; Elsevier: Amsterdam, The Netherlands, 2021. [Google Scholar]
- Niu, X.; Thaochan, N.; Hu, Q. Diversity of Linear Non-Ribosomal Peptide in Biocontrol Fungi. J. Fungi 2020, 6, 61. [Google Scholar] [CrossRef]
- Collemare, J.; Billard, A.; Böhnert, H.U.; Lebrun, M.-H. Biosynthesis of secondary metabolites in the rice blast fungus Magnaporthe grisea: The role of hybrid PKS-NRPS in pathogenicity. Mycol. Res. 2008, 112, 207–215. [Google Scholar] [CrossRef]
- Herbst, D.A.; Townsend, C.A.; Maier, T. The architectures of iterative type I PKS and FAS. Nat. Prod. Rep. 2018, 35, 1046–1069. [Google Scholar] [CrossRef]
- Yu, D.; Xu, F.; Zeng, J.; Zhan, J. Type III polyketide synthases in natural product biosynthesis. IUBMB Life 2012, 64, 285–295. [Google Scholar] [CrossRef]
- Hashimoto, M.; Seshime, Y.; Kitamoto, K.; Uchiyama, N.; Goda, Y.; Fujii, I. Identification of csypyrone B2 and B3 as the minor products of Aspergillus oryzae type III polyketide synthase CsyB. Bioorg. Med. Chem. Lett. 2013, 23, 650–653. [Google Scholar] [CrossRef]
- Sun, L.; Wang, S.; Zhang, S.; Yu, D.; Qin, Y.; Huang, H.; Wang, W.; Zhan, J. Identification of a type III polyketide synthase involved in the biosynthesis of spirolaxine. Appl. Microbiol. Biotechnol. 2016, 100, 7103–7113. [Google Scholar] [CrossRef]
- Ramakrishnan, D.; Tiwari, M.K.; Manoharan, G.; Sairam, T.; Thangamani, R.; Lee, J.K.; Marimuthu, J. Molecular characterization of two alkylresorcylic acid synthases from Sordariomycetes fungi. Enzym. Microb. Technol. 2018, 115, 16–22. [Google Scholar] [CrossRef]
- Yan, H.; Sun, L.; Huang, J.; Qiu, Y.; Xu, F.; Yan, R.; Zhu, D.; Wang, W.; Zhan, J. Identification and heterologous reconstitution of a 5-alk(en)ylresorcinol synthase from endophytic fungus Shiraia sp. Slf14. J. Microbiol. 2018, 56, 805–812. [Google Scholar] [CrossRef]
- Kaneko, A.; Morishita, Y.; Tsukada, K.; Taniguchi, T.; Asai, T. Post-genomic approach based discovery of alkylresorcinols from a cricket-associated fungus, Penicillium soppi. Org. Biomol. Chem. 2019, 17, 5239–5243. [Google Scholar] [CrossRef]
- Manoharan, G.; Sairam, T.; Thangamani, R.; Ramakrishnan, D.; Tiwari, M.K.; Lee, J.K.; Marimuthu, J. Identification and characterization of type III polyketide synthase genes from culturable endophytes of ethnomedicinal plants. Enzym. Microb. Technol. 2019, 131, 109396. [Google Scholar] [CrossRef]
- Minami, A.; Ugai, T.; Ozaki, T.; Oikawa, H. Predicting the chemical space of fungal polyketides by phylogeny-based bioinformatics analysis of polyketide synthase-nonribosomal peptide synthetase and its modification enzymes. Sci. Rep. 2020, 10, 13556. [Google Scholar] [CrossRef]
- Rusu, A.V.; Trif, M.; Rocha, J.M. Microbial Secondary Metabolites via Fermentation Approaches for Dietary Supplementation Formulations. Molecules 2023, 28, 6020. [Google Scholar] [CrossRef]
- Wang, S.; Chen, H.; Tang, X.; Zhang, H.; Chen, W.; Chen, Y.Q. Molecular tools for gene manipulation in filamentous fungi. Appl. Microbiol. Biotechnol. 2017, 101, 8063–8075. [Google Scholar] [CrossRef]
- Wang, Q.; Coleman, J.J. Progress and Challenges: Development and Implementation of CRISPR/Cas9 Technology in Filamentous Fungi. Comput. Struct. Biotechnol. J. 2019, 17, 761–769. [Google Scholar] [CrossRef]
- Mojica, F.J.M.; Díez-Villaseñor, C.; García-Martínez, J.; Almendros, C. Short motif sequences determine the targets of the prokaryotic CRISPR defence system. Microbiology 2009, 155, 733–740. [Google Scholar] [CrossRef]
- Sternberg, S.H.; Redding, S.; Jinek, M.; Greene, E.C.; Doudna, J.A. DNA interrogation by the CRISPR RNA-guided endonuclease Cas9. Nature 2014, 507, 62–67. [Google Scholar] [CrossRef]
- Kim, H.; Kim, J.-S. A guide to genome engineering with programmable nucleases. Nat. Rev. Genet. 2014, 15, 321–334. [Google Scholar] [CrossRef] [PubMed]
- Boettcher, M.; McManus, M.T. Choosing the Right Tool for the Job: RNAi, TALEN, or CRISPR. Mol. Cell 2015, 58, 575–585. [Google Scholar] [CrossRef] [PubMed]
- Mei, Y.Z.; Zhu, Y.L.; Huang, P.W.; Yang, Q.; Dai, C.C. Strategies for gene disruption and expression in filamentous fungi. Appl. Microbiol. Biotechnol. 2019, 103, 6041–6059. [Google Scholar] [CrossRef] [PubMed]
- Case, M.E.; Schweizer, M.; Kushner, S.R.; Giles, N.H. Efficient transformation of Neurospora crassa by utilizing hybrid plasmid DNA. Proc. Natl. Acad. Sci. USA 1979, 76, 5259–5263. [Google Scholar] [CrossRef] [PubMed]
- Turgeon, B.G.; Condon, B.; Liu, J.; Zhang, N. Protoplast transformation of filamentous fungi. Methods Mol. Biol. 2010, 638, 3–19. [Google Scholar] [CrossRef] [PubMed]
- de Groot, M.J.A.; Bundock, P.; Hooykaas, P.J.J.; Beijersbergen, A.G.M. Agrobacterium tumefaciens-mediated transformation of filamentous fungi. Nat. Biotechnol. 1998, 16, 839–842. [Google Scholar] [CrossRef] [PubMed]
- Richey, M.G.; Marek, E.T.; Schardl, C.L.; Smith, D.A. Transformation of filamentous fungi with plasmid DNA by electroporation. Phytopathology 1989, 79, 844–847. [Google Scholar] [CrossRef]
- Bhairi, S.M.; Staples, R.C. Transient expression of the β-glucuronidase gene introduced into Uromyces appendiculatus uredospores by particle bombardment. Phytopathology 1992, 82, 986–989. [Google Scholar] [CrossRef]
- Chen, K.; Stahl, E.C.; Kang, M.H.; Xu, B.; Allen, R.; Trinidad, M.; Doudna, J.A. Engineering self-deliverable ribonucleoproteins for genome editing in the brain. Nat. Commun. 2024, 15, 1727. [Google Scholar] [CrossRef]
- Zhang, Y.; Iaffaldano, B.; Qi, Y. CRISPR ribonucleoprotein-mediated genetic engineering in plants. Plant Commun. 2021, 2, 100168. [Google Scholar] [CrossRef]
- Horodecka, K.; Düchler, M. CRISPR/Cas9: Principle, Applications, and Delivery through Extracellular Vesicles. Int. J. Mol. Sci. 2021, 22, 6072. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Lv, G.; Tu, Y.; Cheng, X.; Duan, Y.; Zeng, B.; He, B. Applications of CRISPR/Cas9 in the Synthesis of Secondary Metabolites in Filamentous Fungi. Front. Microbiol. 2021, 12, 638096. [Google Scholar] [CrossRef] [PubMed]
- Fuller, K.K.; Chen, S.; Loros, J.J.; Dunlap, J.C. Development of the CRISPR/Cas9 System for Targeted Gene Disruption in Aspergillus fumigatus. Eukaryot. Cell 2015, 14, 1073–1080. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Chen, L.; Jiang, Y.; Zhou, Z.; Zou, G. Efficient genome editing in filamentous fungus Trichoderma reesei using the CRISPR/Cas9 system. Cell Discov. 2015, 1, 15007. [Google Scholar] [CrossRef] [PubMed]
- Matsu-Ura, T.; Baek, M.; Kwon, J.; Hong, C. Efficient gene editing in Neurospora crassa with CRISPR technology. Fungal Biol. Biotechnol. 2015, 2, 4. [Google Scholar] [CrossRef] [PubMed]
- Nødvig, C.S.; Nielsen, J.B.; Kogle, M.E.; Mortensen, U.H. A CRISPR-Cas9 System for Genetic Engineering of Filamentous Fungi. PLoS ONE 2015, 10, e0133085. [Google Scholar] [CrossRef] [PubMed]
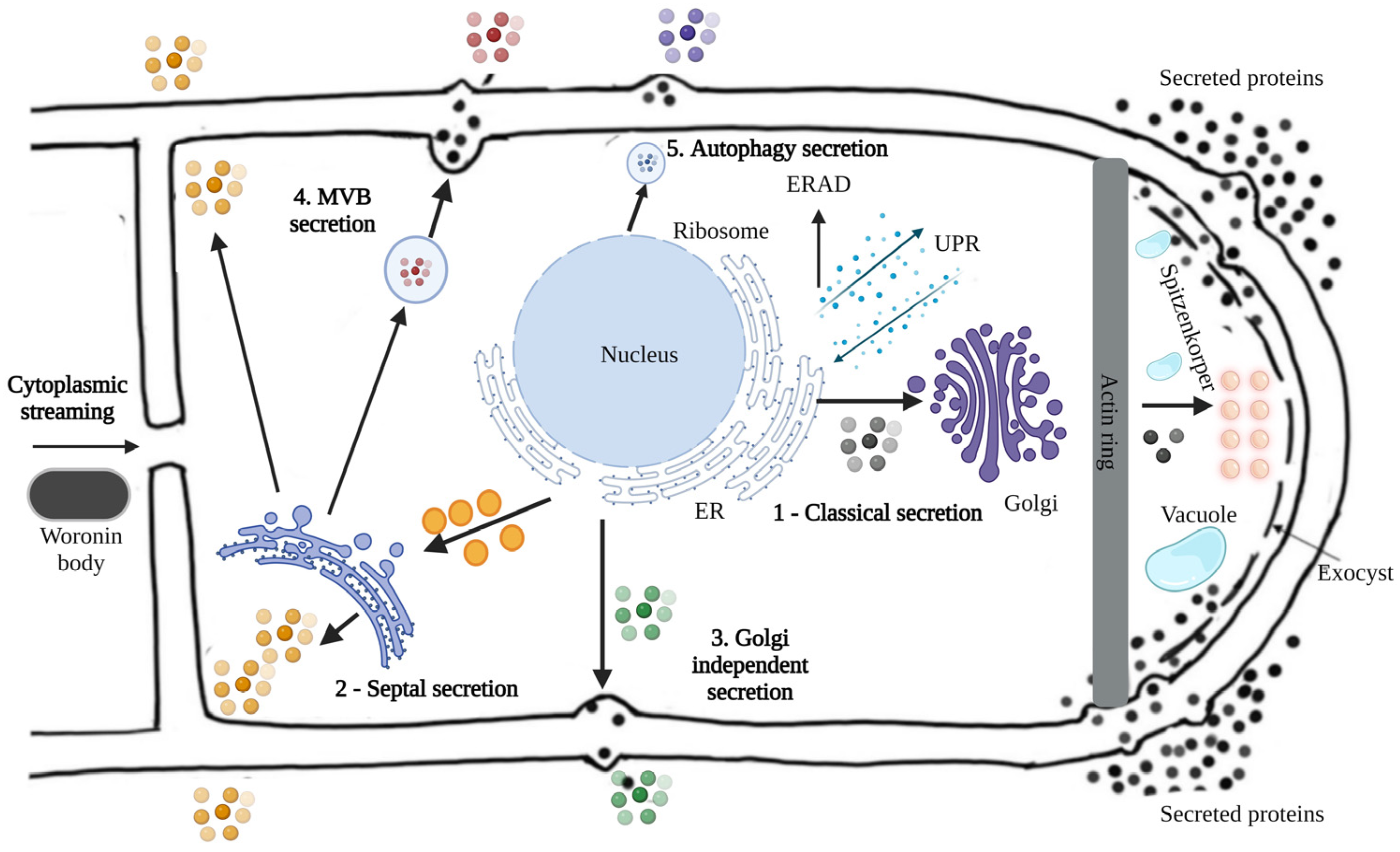
- Conesa, A.; Punt, P.J.; van Luijk, N.; van den Hondel, C.A. The secretion pathway in filamentous fungi: A biotechnological view. Fungal Genet. Biol. 2001, 33, 155–171. [Google Scholar] [CrossRef] [PubMed]
- Hayakawa, Y.; Ishikawa, E.; Shoji, J.Y.; Nakano, H.; Kitamoto, K. Septum-directed secretion in the filamentous fungus Aspergillus oryzae. Mol. Microbiol. 2011, 81, 40–55. [Google Scholar] [CrossRef]
- Fiedler, M.R.M.; Cairns, T.C.; Koch, O.; Kubisch, C.; Meyer, V. Conditional Expression of the Small GTPase ArfA Impacts Secretion, Morphology, Growth, and Actin Ring Position in Aspergillus niger. Front. Microbiol. 2018, 9, 878. [Google Scholar] [CrossRef]
- Shoji, J.Y.; Kikuma, T.; Kitamoto, K. Vesicle trafficking, organelle functions, and unconventional secretion in fungal physiology and pathogenicity. Curr. Opin. Microbiol. 2014, 20, 1–9. [Google Scholar] [CrossRef]
- Miura, N.; Ueda, M. Evaluation of Unconventional Protein Secretion by Saccharomyces cerevisiae and other Fungi. Cells 2018, 7, 128. [Google Scholar] [CrossRef] [PubMed]
- Sakekar, A.A.; Gaikwad, S.R.; Punekar, N.S. Protein expression and secretion by filamentous fungi. J. Biosci. 2021, 46, 5. [Google Scholar] [CrossRef]
- Virag, A.; Harris, S.D. The Spitzenkörper: A molecular perspective. Mycol. Res. 2006, 110, 4–13. [Google Scholar] [CrossRef]
- Haßdenteufel, S.; Johnson, N.; Paton, A.W.; Paton, J.C.; High, S.; Zimmermann, R. Chaperone-Mediated Sec61 Channel Gating during ER Import of Small Precursor Proteins Overcomes Sec61 Inhibitor-Reinforced Energy Barrier. Cell Rep. 2018, 23, 1373–1386. [Google Scholar] [CrossRef]
- Saloheimo, M.; Pakula, T.M. The cargo and the transport system: Secreted proteins and protein secretion in Trichoderma reesei (Hypocrea jecorina). Microbiology 2012, 158, 46–57. [Google Scholar] [CrossRef] [PubMed]
- Mitra, N.; Sinha, S.; Ramya, T.N.; Surolia, A. N-linked oligosaccharides as outfitters for glycoprotein folding, form and function. Trends Biochem. Sci. 2006, 31, 156–163. [Google Scholar] [CrossRef] [PubMed]
- Bernasconi, R.; Molinari, M. ERAD and ERAD tuning: Disposal of cargo and of ERAD regulators from the mammalian ER. Curr. Opin. Cell Biol. 2011, 23, 176–183. [Google Scholar] [CrossRef]
- Wang, G.; Zhang, D.; Chen, S. Effect of earlier unfolded protein response and efficient protein disposal system on cellulase production in Rut C30. World J. Microbiol. Biotechnol. 2014, 30, 2587–2595. [Google Scholar] [CrossRef]
- Spang, A. Membrane traffic in the secretory pathway. Cell. Mol. Life Sci. 2008, 65, 2781–2789. [Google Scholar] [CrossRef]
- Hutagalung, A.H.; Novick, P.J. Role of Rab GTPases in membrane traffic and cell physiology. Physiol. Rev. 2011, 91, 119–149. [Google Scholar] [CrossRef]
- Söllner, T.; Whiteheart, S.W.; Brunner, M.; Erdjument-Bromage, H.; Geromanos, S.; Tempst, P.; Rothman, J.E. SNAP receptors implicated in vesicle targeting and fusion. Nature 1993, 362, 318–324. [Google Scholar] [CrossRef] [PubMed]
- Valkonen, M.; Kalkman, E.R.; Saloheimo, M.; Penttilaö, M.; Read, N.D.; Duncan, R.R. Spatially Segregated SNARE Protein Interactions in Living Fungal Cells. J. Biol. Chem. 2007, 282, 22775–22785. [Google Scholar] [CrossRef] [PubMed]
- Saunders, G.; Picknett, T.M.; Tuite, M.F.; Ward, M. Heterologous gene expression in filamentous fungi. Trends Biotechnol. 1989, 7, 283–287. [Google Scholar] [CrossRef]
- Jeenes, D.J.; Mackenzie, D.A.; Roberts, I.N.; Archer, D.B. Heterologous protein production by filamentous fungi. Biotechnol. Genet. Eng. Rev. 1991, 9, 327–367. [Google Scholar]
- Nevalainen, K.M.; Te’o, V.S.; Bergquist, P.L. Heterologous protein expression in filamentous fungi. Trends Biotechnol. 2005, 23, 468–474. [Google Scholar] [CrossRef] [PubMed]
- Su, X.; Schmitz, G.; Zhang, M.; Mackie, R.I.; Cann, I.K. Heterologous gene expression in filamentous fungi. Adv. Appl. Microbiol. 2012, 81, 1–61. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Wang, Y.H.; Liu, T.Q.; Zhang, H.; Zhang, H.; Li, J. The GlaA signal peptide substantially increases the expression and secretion of α-galactosidase in Aspergillus niger. Biotechnol. Lett. 2018, 40, 949–955. [Google Scholar] [CrossRef] [PubMed]
- Ohno, A.; Maruyama, J.; Nemoto, T.; Arioka, M.; Kitamoto, K. A carrier fusion significantly induces unfolded protein response in heterologous protein production by Aspergillus oryzae. Appl. Microbiol. Biotechnol. 2011, 92, 1197–1206. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, D.I.; Olsthoorn, M.M.; Maillet, I.; Akeroyd, M.; Breestraat, S.; Donkers, S.; van der Hoeven, R.A.; van den Hondel, C.A.; Kooistra, R.; Lapointe, T.; et al. Effective lead selection for improved protein production in Aspergillus niger based on integrated genomics. Fungal Genet. Biol. 2009, 46 (Suppl. 1), S141–S152. [Google Scholar] [CrossRef]
- Wu, Y.; Sun, X.; Xue, X.; Luo, H.; Yao, B.; Xie, X.; Su, X. Overexpressing key component genes of the secretion pathway for enhanced secretion of an Aspergillus niger glucose oxidase in Trichoderma reesei. Enzym. Microb. Technol. 2017, 106, 83–87. [Google Scholar] [CrossRef]
- Hoang, H.D.; Maruyama, J.; Kitamoto, K. Modulating endoplasmic reticulum-Golgi cargo receptors for improving secretion of carrier-fused heterologous proteins in the filamentous fungus Aspergillus oryzae. Appl. Environ. Microbiol. 2015, 81, 533–543. [Google Scholar] [CrossRef] [PubMed]
- Garst, A.D.; Bassalo, M.C.; Pines, G.; Lynch, S.A.; Halweg-Edwards, A.L.; Liu, R.; Liang, L.; Wang, Z.; Zeitoun, R.; Alexander, W.G.; et al. Genome-wide mapping of mutations at single-nucleotide resolution for protein, metabolic and genome engineering. Nat. Biotechnol. 2017, 35, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xue, W.; Sims, A.H.; Zhao, C.; Wang, A.; Tang, G.; Qin, J.; Wang, H. Isolation of four pepsin-like protease genes from Aspergillus niger and analysis of the effect of disruptions on heterologous laccase expression. Fungal Genet. Biol. 2008, 45, 17–27. [Google Scholar] [CrossRef] [PubMed]


| Generations | Biofuel Technology [23] | Bio-Input Technology | |
|---|---|---|---|
| Traditional Technology | The First Generation | First-generation biofuels are derived from agricultural crops intended for human consumption. For instance, (a) Biodiesel is obtained through the chemical processes of transesterification and esterification from oilseed plants or vegetable materials. (b) Ethanol is derived from plant biomass containing fermentable sugars, which undergo fermentation to produce fuel. | First generation bio-inputs of microbial origin refer to the utilization of cells or spores of microorganisms, such as bacteria, fungi, or viruses, for the management of pests and/or diseases in agricultural crops. These bio-inputs can be produced through various methods: (a) Liquid (submerged) fermentation is employed when the objective is to harvest cells, particularly for the production of bacterial cells and spores [29]. (b) Solid fermentation is utilized when the goal is to acquire spores, particularly for the cultivation of fungal spores [29]. (c) Two-phase fermentation involves an initial stage of liquid fermentation, followed by solid-state fermentation [30,31]. (d) In vivo cultivation of insects is employed to obtain viruses for bio-input production [29]. |
| Advanced Technologies | The Second Generation | Second-generation biofuels originate from non-food crops, such as crop residues, wood, and dedicated energy crops cultivated explicitly for biofuel production. | Second-generation bio-inputs entail the amalgamation of diverse microorganisms within a formulation aimed at broadening the spectrum of action exhibited by biological agents. Some examples of application of the 2nd generation of bioinputs: (a) Use of isolates Bacillus amyloliquefaciens, CCT 7901 + Trichoderma asperellum URM 8120 + Trichoderma harzianum, URM 8119 in the control of belly rot (Rhizoctonia solani), cottony soft rot (Sclerotinia sclerotiorum) and anthracnose of bean (Colletotrichum lindemuthianum); (b) Bacillus licheniformis, ATCC 12713 + Bacillus subtilis, ATCC 6051 + Paecilomyces lilacinus (Purpureocilium lilacinum), CPQBA 040-11 DRM 10 in the control of Root-knot nematode (Meloidogyne incognita) and Root-lesion nematode (Pratylenchus brachyurus); (c) Bacillus thuringiensis, S 234 + Metarhizium rileyi, CG 1153 in the control of Soybean caterpillar (Anticarsia gemmatalis), Soybean looper (Chrysodeixis includens), Cotton bollworm (Helicoverpa armigera), The black army worm (Spodoptera cosmioides), Southern armyworm (Spodoptera eridania) and the fall armyworm (Spodoptera frugiperda); (d) Beauveria bassiana, IBCB 66 + Metarhizium anisopliae, IBCB 425 in the control of Grasshopper leafhopper (Deois flavopicta) and The neotropical brown stink bug (Euschistus heros) (Supplementary Table S1) [9]. |
| The Third Generation | Third-generation biofuels are predicated on advancements in biomass production, with algae serving as the principal raw material within this category. | Third-generation bio-inputs refer to those designed to acquire and refine the primary and secondary metabolites generated by microorganisms through fermentation processes. Within this context, the active component comprises organic molecules or proteins that exert influence over pest and/or disease management in agricultural crops. An example of a third generation bioinput is the fermentation of S. spinosa produces a natural mixture containing spinosyn A as the major component and spinosyn D as the minor component, a mixture that has been named spinosad. Several commercial products containing spinosad as their active ingredient for insect control on organic crops, and fruitfly and fire ant bait traps. The products are also useful in integrated pest management and insecticide resistance management programs [32]. | |
| The Fourth Generation | Fourth-generation biofuels strive to offer enhanced sustainability in production methodologies, integrating biofuel manufacturing with carbon dioxide (CO2) capture and storage techniques such as oxy-combustion processes or employing genetic engineering and nanotechnology approaches. | Fourth-generation bio-inputs, the focus of this review article, seek to present heightened sustainability in production strategies through the utilization of genetic engineering or nanotechnology methodologies. This encompasses the application of the CRISPR Cas system for genetic editing and augmenting the secretion of proteins and metabolites by microorganisms. A pioneering study demonstrated the modification of Trichoderma harzianum using the CRISPR/Cas9 marker-free system, targeting the albA (pks4) and ku70 genes. This achievement marks the first successful application of this recyclable system for constructing fungal strains with agricultural applications [33]. |
| Protein of Interest | Species | Strategy | Improvements | Method | Reference |
|---|---|---|---|---|---|
| α-Galactosidase | A. niger | Replacement of the original signal peptide by a glucoamylase signal peptide (GlaA) in A. niger | Approximately 9-fold increase | TMA | [99] |
| Chymosin | A. oryzae | Fusion of a target protein with a naturally secreted α-amylase protein | 2-fold increase | TMP | [100] |
| β-Glucuronidase | A. niger | The regulation of UPR and ERAD is achieved through the overexpression of sttC and the deletion of dorA. | Not measured | HR | [101] |
| Glucose oxidase | T. reesei | Regulation of UPR and ERAD via overexpression of bip1 or hac1 | 1.5 to 1.8 times increase | PEG-mediated TMP | [102] |
| Glucose oxidase | T. reesei | Optimization of the intracellular transport process by overexpression of snc1 | 2.2-fold increase | PEG-mediated TMP | [102] |
| Prochymosin | A. niger | Optimizing the intracellular transport process can be achieved by deleting Aovip36 or Aoemp47, as well as fusing the target protein with α-amylase. | Approximately 2-fold increase | TMP | [103] |
| Cellulase | T. reesei | Construction of a strain deficient in protease production because of the deletion of res-1, cre-1, gh-1 and alp-1 | 5-fold increase | CREATE | [104] |
| Laccase | A. niger | Construction of a strain deficient in protease production by deletion of pepAa, pepAb or pepAd | 1.21-to-1.42-fold increase | PEG-mediated TMP | [105] |
| Glucoamylase | A. niger | Regulation of mycelium morphology by racA deletion | 4-fold increase | TMP | [81] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oliveira, L.R.; Gonçalves, A.R.; Quintela, E.D.; Colognese, L.; Cortes, M.V.d.C.B.; de Filippi, M.C.C. Genetic Engineering of Filamentous Fungi: Prospects for Obtaining Fourth-Generation Biological Products. Appl. Microbiol. 2024, 4, 794-810. https://doi.org/10.3390/applmicrobiol4020055
Oliveira LR, Gonçalves AR, Quintela ED, Colognese L, Cortes MVdCB, de Filippi MCC. Genetic Engineering of Filamentous Fungi: Prospects for Obtaining Fourth-Generation Biological Products. Applied Microbiology. 2024; 4(2):794-810. https://doi.org/10.3390/applmicrobiol4020055
Chicago/Turabian StyleOliveira, Lorena Resende, Ariany Rosa Gonçalves, Eliane Dias Quintela, Leandro Colognese, Marcio Vinicius de C. Barros Cortes, and Marta Cristina Corsi de Filippi. 2024. "Genetic Engineering of Filamentous Fungi: Prospects for Obtaining Fourth-Generation Biological Products" Applied Microbiology 4, no. 2: 794-810. https://doi.org/10.3390/applmicrobiol4020055
APA StyleOliveira, L. R., Gonçalves, A. R., Quintela, E. D., Colognese, L., Cortes, M. V. d. C. B., & de Filippi, M. C. C. (2024). Genetic Engineering of Filamentous Fungi: Prospects for Obtaining Fourth-Generation Biological Products. Applied Microbiology, 4(2), 794-810. https://doi.org/10.3390/applmicrobiol4020055