
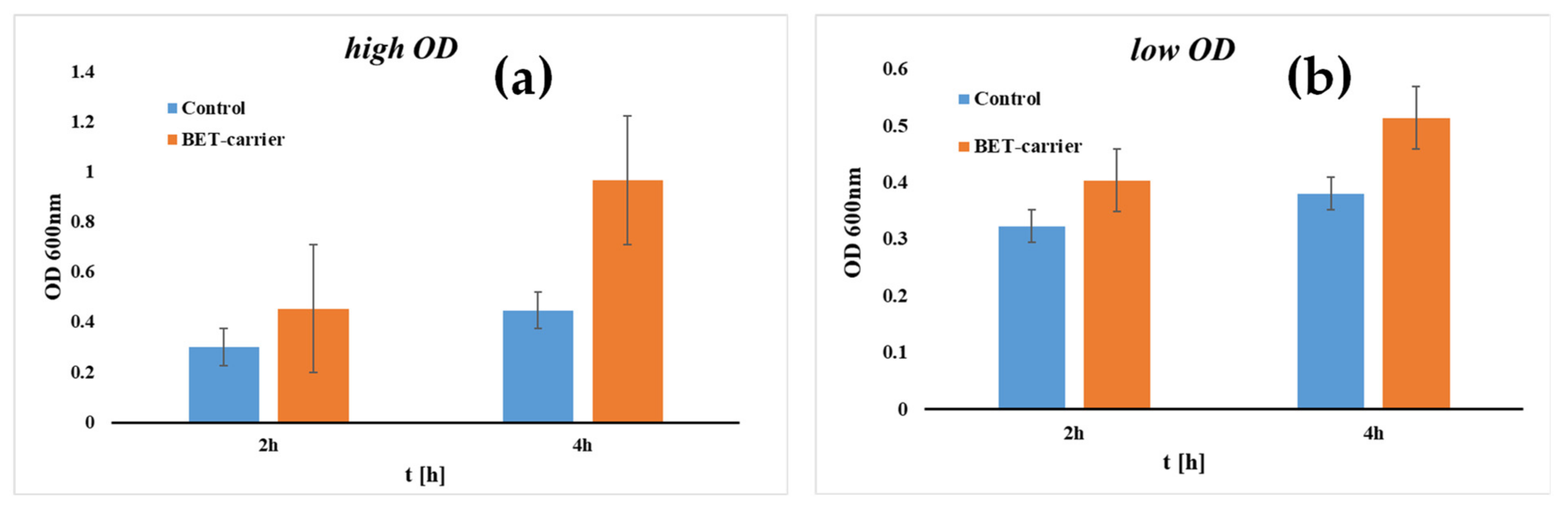
Biospeckle Analysis and Biofilm Electrostatic Tests, Two Useful Methods in Microbiology


 ,
,  ,
,  and
and 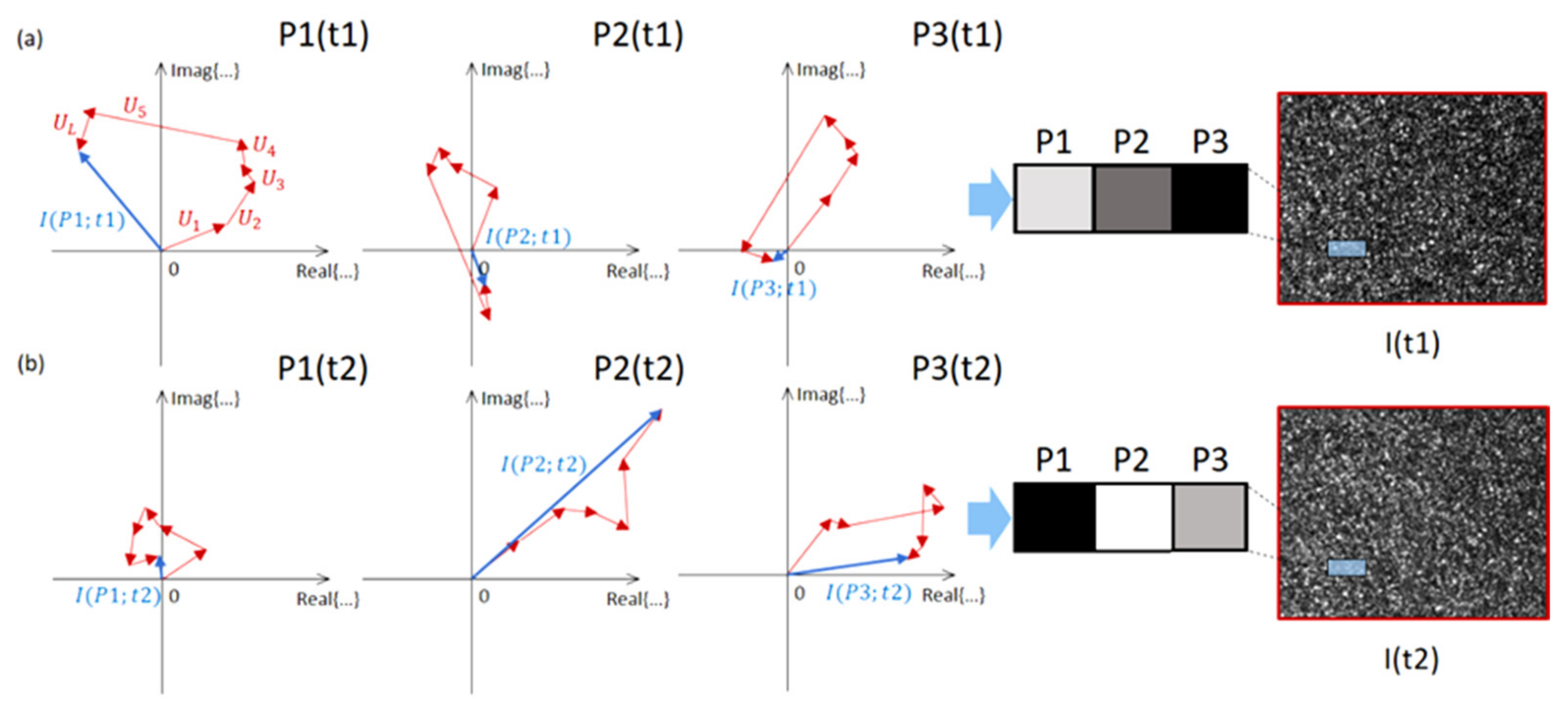{kind=link}
{kind=link}
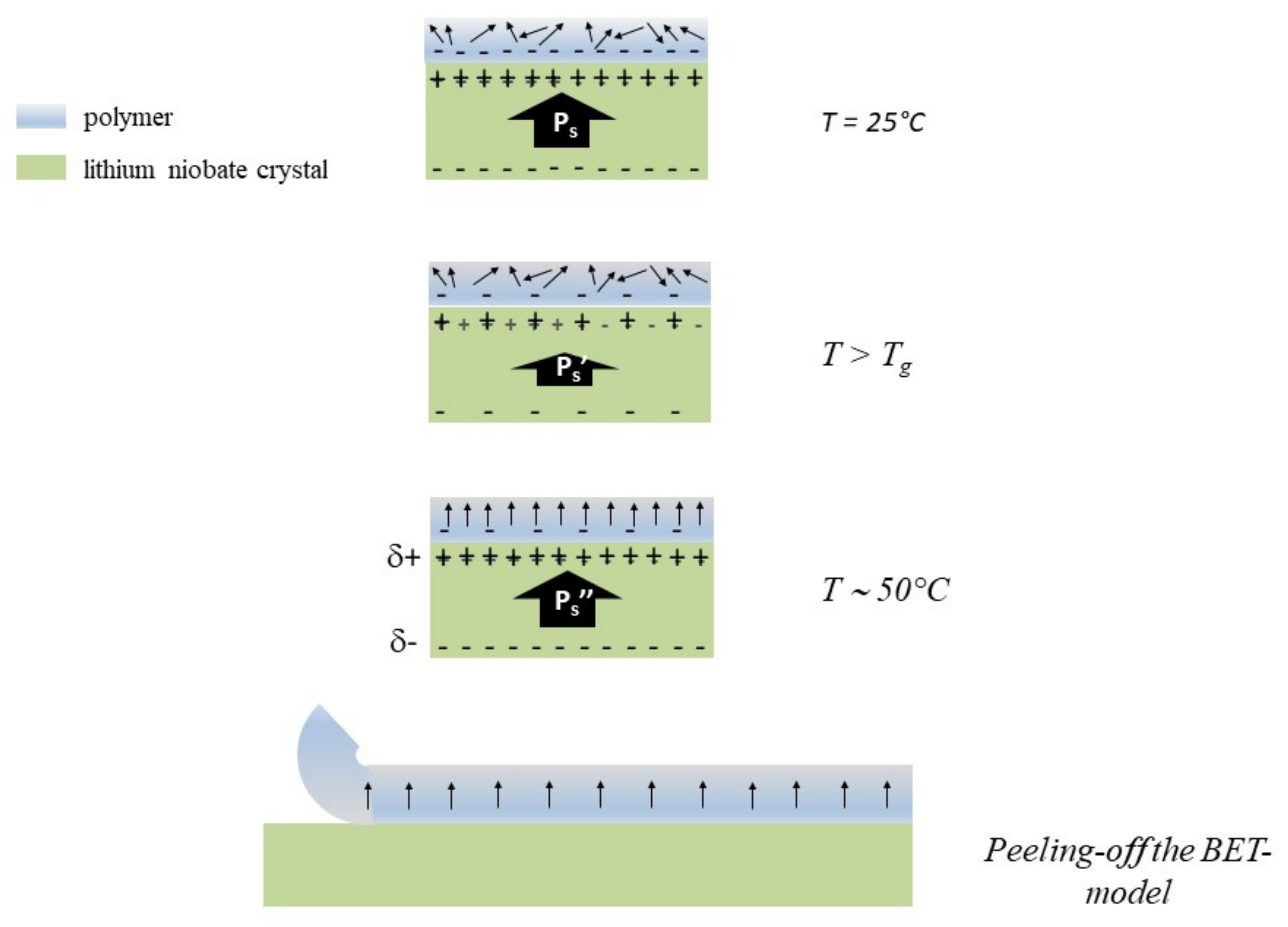
{kind=link}
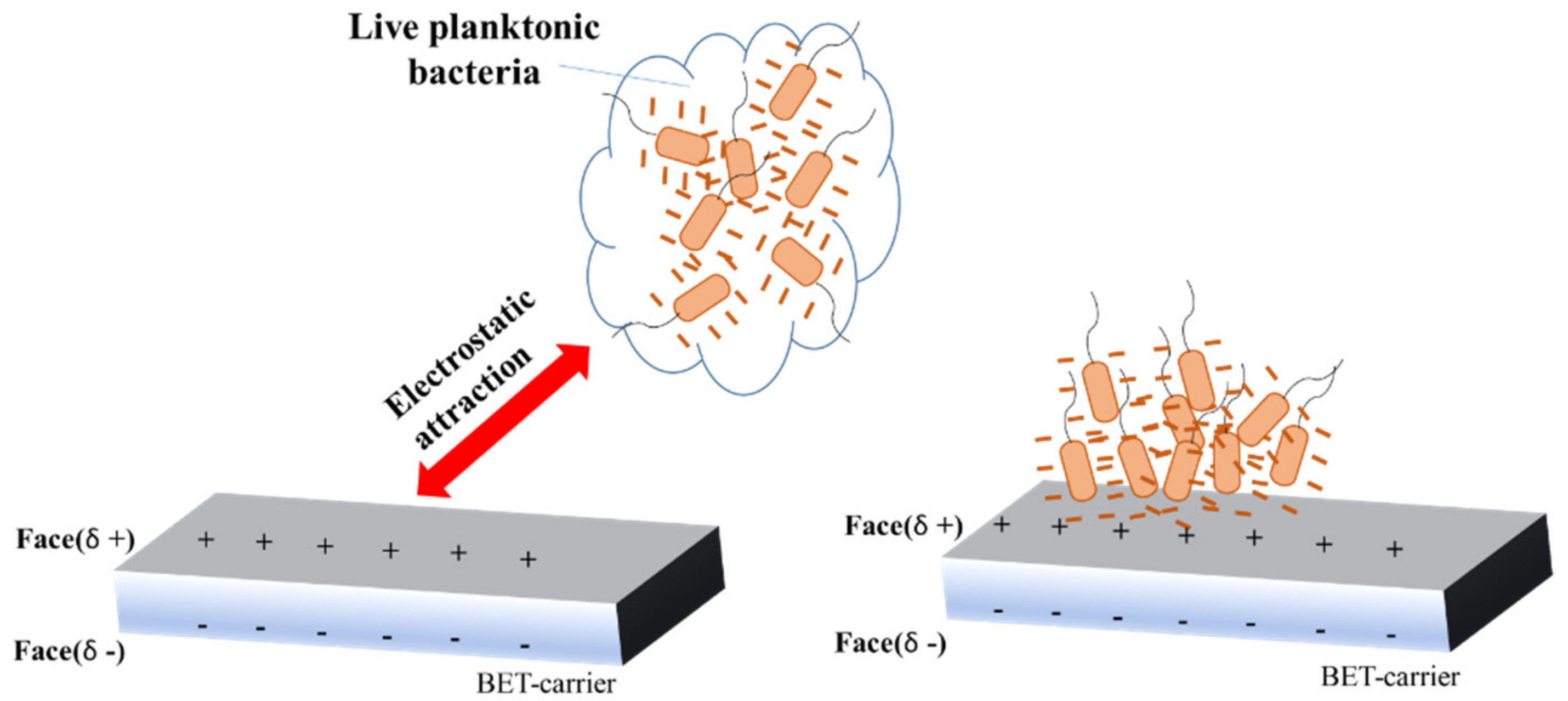
{kind=link}
{kind=link}
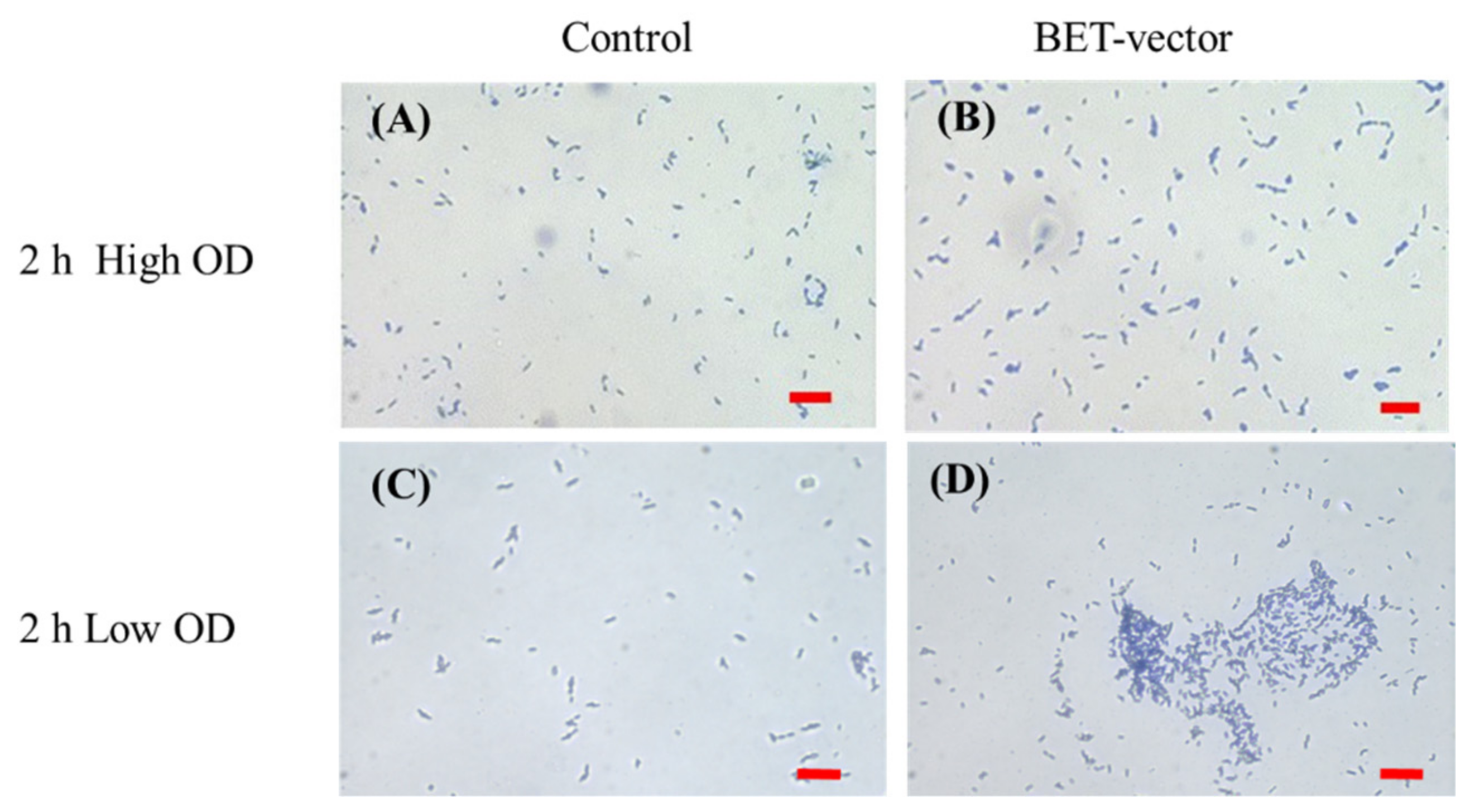{kind=link}
{kind=link}
Abstract
Share and Cite
Oleandro, E.; Grilli, S.; Rega, R.; Mugnano, M.; Bianco, V.; Valentino, M.; Mandracchia, B.; Nazzaro, F.; Coppola, R.; Ferraro, P. Biospeckle Analysis and Biofilm Electrostatic Tests, Two Useful Methods in Microbiology. Appl. Microbiol. 2021, 1, 557-572. https://doi.org/10.3390/applmicrobiol1030036
Oleandro E, Grilli S, Rega R, Mugnano M, Bianco V, Valentino M, Mandracchia B, Nazzaro F, Coppola R, Ferraro P. Biospeckle Analysis and Biofilm Electrostatic Tests, Two Useful Methods in Microbiology. Applied Microbiology. 2021; 1(3):557-572. https://doi.org/10.3390/applmicrobiol1030036
Chicago/Turabian StyleOleandro, Emilia, Simonetta Grilli, Romina Rega, Martina Mugnano, Vittorio Bianco, Marika Valentino, Biagio Mandracchia, Filomena Nazzaro, Raffaele Coppola, and Pietro Ferraro. 2021. "Biospeckle Analysis and Biofilm Electrostatic Tests, Two Useful Methods in Microbiology" Applied Microbiology 1, no. 3: 557-572. https://doi.org/10.3390/applmicrobiol1030036
APA StyleOleandro, E., Grilli, S., Rega, R., Mugnano, M., Bianco, V., Valentino, M., Mandracchia, B., Nazzaro, F., Coppola, R., & Ferraro, P. (2021). Biospeckle Analysis and Biofilm Electrostatic Tests, Two Useful Methods in Microbiology. Applied Microbiology, 1(3), 557-572. https://doi.org/10.3390/applmicrobiol1030036