Abstract
Kazakhstan cultivates over 12 million hectares of wheat, primarily spring wheat in the northern region. Spring wheat yields are low, ranging from 1.2 to 1.7 t/ha depending on weather conditions. Northern Kazakhstan is served by five spring wheat breeding programs: A.I. Barayev Research and Production Centre for Grain Farming and Agricultural Experimental Stations located in the Aktobe, Karagandy, Kostanay, and North Kazakhstan regions. In 2022, a germplasm set was assembled, including cultivars and breeding lines from the five breeding programs, totaling 84 genotypes. This set was evaluated in field trials during 2022 and 2023 at the breeding programs that contributed to the germplasm (except Aktobe). The material was also screened for molecular markers associated with genes for agronomic traits. The study objective was to compare the diversity and performance of germplasm originating from different breeding programs and identify potential underlying drivers. Breeding sites grouped based on variations in air temperature, precipitation, and grain yield demonstrated both similarities and differences among sites. However, these similarities were not reflected in the agronomic performance of materials originating from different locations. The expectation that germplasm would perform best for grain yield at its “home” location was not always confirmed. Grouping of germplasm based on genetic diversity of 20 molecular markers was not related to similarities in environmental conditions at the places of origin. The performance and diversity of germplasm from each of the five breeding programs is apparently driven by factors beyond environment, including breeding strategy and methodology, parental pool, and, in the absence of modern tools, breeders’ intuition and selection robustness. Kazakh spring wheat breeding programs require improvement to remain competitive in the face of increasing pressure from introduced foreign cultivars.
1. Introduction
Kazakhstan is one of the world’s major wheat producers, with approximately 12 million hectares under cultivation. Most of the crop is produced in the northern part of the country above 50° N latitude. This region has a short growing season for rainfed spring wheat, which is sown in May and harvested in September. The cropping system includes spring barley, oil crops (sunflower, flax, safflower, and rapeseed), pulses (dry peas, lentils, and chickpeas), and forage crops (maize, sorghum, Sudan grass, and cereal–legume mixtures). Spring wheat yields are low, rarely exceeding 1.2–1.5 t/ha. Climate change has resulted in warmer temperatures, an expanded growing season, and a marginal increase in precipitation [1]. The main abiotic stress is moisture limitation, which reduces productivity on average for 1–2 years out of every five. Kazakhstan’s wheat grain exports contribute significantly to regional and global food security.
Despite increased input use, the introduction of new machinery and technologies, and the adoption of foreign cultivars, wheat yield dynamics in the country have shown a stagnating trend over the last decade [2]. The lifespan of spring wheat cultivars in Kazakhstan exceeds 10–12 years. Currently grown cultivars may not satisfy producer needs or withstand the abiotic and biotic stresses associated with climate change. The Register of officially released spring wheat cultivars for 2023 comprises 131 genotypes, including 62% bred locally, 29% from Russia, and 9% from abroad, primarily from Europe [3]. Despite increasing pressure from foreign cultivars, Kazakh spring wheat cultivars remain the backbone of production.
Spring wheat breeding in Northern Kazakhstan involves public breeding programs at five agricultural research establishments: the A.I. Barayev Research and Production Centre for Grain Farming (Kaz. Grain SPC) near Astana and agricultural experimental stations (AESs) in the main spring wheat-producing regions of Karagandy (Krg), North Kazakhstan (N-Kaz), Kostanay (Karabalyk, Kar), and Aktobe (Akt). These research institutions serve over 12 million hectares of arable land across a territory spanning 1600 km from east to west and 800 km from south to north. All five breeding programs are well-established, with continuous cultivar development spanning 50–60 years. Each program contributes cultivars to the registration list that are adapted to local conditions and grown by producers. However, the increasing interest of the farming community in cultivars and the successful introduction of foreign varieties raise questions about the relative adaptation and diversity of contemporary Kazakh spring wheat germplasm. The primary study hypothesis was that spring wheat germplasm developed at a specific breeding program in Kazakhstan would be better adapted to the conditions in which it was created, while germplasm diversity would reflect the diversity of agroecology as well as the breeding approaches and methods employed.
Several studies on this subject were conducted within the framework of the Kazakhstan-Siberian Network on Spring Wheat Improvement (KASIB). Morgounov et al. [4] studied a core set of 142 spring wheat genotypes from 15 breeding programs at three sites in Kazakhstan and one site in Russia over three years for agronomic traits and the presence of effective molecular markers. Among the top 30 highest-yielding genotypes across all sites and years, 21 genotypes originated from Russia. There were also clear differences between programs in the frequency of molecular markers controlling adaptation and agronomic traits. Tajibayev et al. [5] evaluated a large set of KASIB spring durum wheat at contrasting sites in southern Kazakhstan near Almaty and in the north near Astana for two years. Unexpectedly, germplasm bred in the south was competitive for grain yield in the north and vice versa, while the most stable yield performance was demonstrated by cultivars bred in Omsk, Russia, at both sites. These two studies demonstrated that germplasm adaptation and performance were not necessarily connected to the place of germplasm origin but may perform well outside of their immediate target region.
In the current study, a set of 84 spring bread wheat cultivars and lines from five breeding programs in Kazakhstan was tested for agronomic traits at four origin sites over two years. The set was also genotyped for molecular markers associated with genes contributing to the expression of agronomic traits in the region. Weather and grain yield data from the sites of five breeding programs were extracted for 2014–2023 to evaluate similarities between locations. The study objective was to compare the adaptation, agronomic performance, and genetic diversity of germplasm originating from five breeding programs and relate these characteristics to the environmental conditions in which the breeding took place.
2. Materials and Methods
2.1. Spring Wheat Germplasm
A set of 84 cultivars and breeding lines from five breeding programs was used in this study (Table S1). The material was selected and submitted by each participating breeding program, representing common cultivars and advanced breeding lines from the final stages of testing. The number of genotypes varied from 12 (Aktobe AES) to 20 (Karabalyk AES). Advanced breeding lines represented the majority of the germplasm tested (81%).
2.2. Experimental Sites and Trial Methodology
The locations of the five breeding programs that improve spring wheat for the main production regions of Kazakhstan are presented in Figure 1. All programs are located above 50° N with variable precipitation and air temperature and are representative of their respective spring wheat production regions.
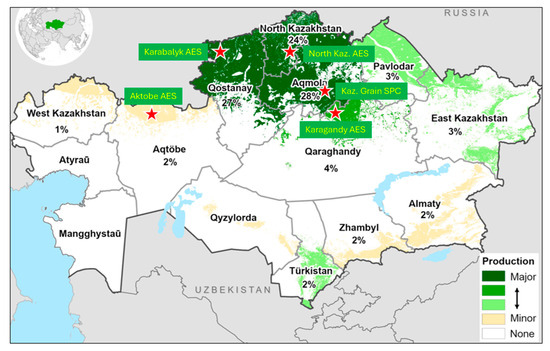
Figure 1.
Location of the breeding programs and experimental sites and wheat production areas in Kazakhstan (Karabalyk AES: 53.850809 N, 62.126478 E; KazGrain SPC: 51.643077 N, 71.028923 E; North Kaz. AES: 54.168868 N, 69.514140 E; Karagandy AES: 50.164469 N, 72.697252 E; Aktobe AES: 50.290666 N, 57.558675 E). Percentage below the region name indicates the share of total wheat area. (Map source: https://ipad.fas.usda.gov/rssiws/al/kz_cropprod.aspx, accessed 15 May 2025).
The soil at the experimental sites was represented by two types of chernozem: ordinary (Karabalyk and North Kazakhstan AES) and southern (Kaz Grain SPC, Karagandy and Aktobe AES). These soils are characterized by relatively high organic matter content (3–5%), potassium availability, and variable nitrogen and phosphorus requirements depending on the cropping system and preceding crop.
Trial agronomy followed common practices: (a) preceding crop was black fallow; (b) soil preparation for planting using harrowing and/or cultivation; (c) planting at optimal dates between 10 and 20 May; (d) seeding rate of 200–300 seeds per m2; (e) herbicide control for common weeds. No fertilizer or disease control was applied in the trials. Normally in the production environment, spring wheat following the black fallow is not fertilized.
The trials were conducted following uniform methodology, with 2–3 m2 plots and two replications in a Randomized Complete Block Design (RCBD). Agronomic traits were evaluated following the CIMMYT methodology described by Pask et al. [6]. The main adaptation traits evaluated were days from emergence to heading and plant height. The main yield components evaluated in the trials were spike length, number of spikelets per spike, number of grains per spike and spikelet, and 1000-kernel weight (TKW). These traits were evaluated in only one replication. Grain yield was evaluated in two replications by harvesting the entire plot. The reaction of the germplasm to diseases (leaf and stem rust, tan spot) was evaluated at the Kazakh Research Institute of Biosafety in Zhambyl region under artificial inoculation using a mixture of common races.
2.3. Genotyping and KASP Assay
Genomic DNA from 84 genotypes was extracted from individual seedlings using the Delaporta protocol with modifications [7]. The DNA concentration of each sample was adjusted to 50 ng/μL. Twenty KASP assays were newly designed using SNPs associated with plant adaptation and agronomic traits (Table 1). All of these markers were previously documented by Turuspekov et al. [8] and Amalova et al. [9,10,11] through genome-wide association studies (GWASs) and biparental mapping. Specific KASP markers in Table 1 were selected based on previously reported statistically significant associations (p < 0.05) with studied traits and reproducibility of associations across multiple environments. The KASP assays were developed by LGC Genomics Company using twenty-one representative SNPs selected for this purpose [12]. The KASP reaction was carried out in a 10 μL volume containing 5 μL of DNA and 5 μL of the prepared genotyping mix (2 × KASP master mix and primer mix). KASP amplification utilized the following thermal cycling profile: initial denaturation at 94 °C for 15 min, followed by 40 cycles of 94 °C for 20 s and 60 °C for 1 min.

Table 1.
List of KASP markers used in the study.
2.4. Statistics
Biplot analysis was employed to investigate the relationships between breeding sites, weather conditions, adaptation, and yield traits. ANOVA was conducted for adaptation and yield traits, and LSD calculations were used to evaluate the significance of differences between genotypes. R Studio version 3.4 [13] 1 December 2024 was utilized for these analyses. The significance of differences between germplasm originating from different breeding programs was assessed using standard error calculated in MS Excel. Genetic diversity was assessed using Shannon’s information index, unbiased diversity, and percentage of polymorphic loci in GenAlEx ver.6.5 [14]. When discussing yield and other agronomic traits in the text, the mean value of 84 genotypes under investigation is referred to at a certain breeding station.
3. Results
3.1. Variation of Breeding Sites for Weather Conditions and Grain Yield
The first stage of the analysis in this study was to determine the differences between breeding sites for two key weather variables, air temperature and precipitation, as well as grain yield. The last ten years of observations (2014–2023) were taken from meteorological stations and breeding trials. Average annual rainfall varied from around 340 mm (Kaz. Grain SPC, Karagandy and Aktobe AES) to 380 mm at Karabalyk AES and over 400 mm at North Kazakhstan AES. The sites are similarly ranked for precipitation from January to August (Figure 2). The availability of moisture for crops is low and remains a major challenge for achieving stable and high yields.
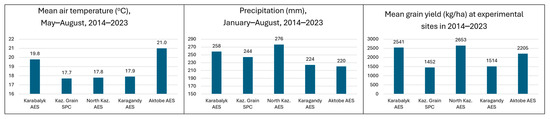
Figure 2.
Mean air temperature in May–August (°C), precipitation in January–August (mm), and spring wheat grain yield (kg/ha) for 2014–2023 across five spring breeding sites.
For the spring wheat growing season from May to August, three sites in the central region (Karagandy, Astana, and North Kazakhstan) were, on average, 2 °C cooler compared to the western sites of Aktobe and Karabalyk (Figure 2). The highest yield during the ten-year period was achieved at North Kazakhstan AES (2653 kg/ha), followed by Karabalyk AES (2541 kg/ha) and Aktobe AES (2205 kg/ha). The yield at Astana (Kaz. Grain SPC) and Karagandy was only in the range of 1500 kg/ha. Based on average values, the five breeding sites can be classified as follows: (a) warm and wet with high yield (Karabalyk AES); (b) warm and dry with average yield (Aktobe AES); (c) cool and wet with high yield (North Kazakhstan AES); (d) cool and dry with low yield (Kaz. Grain SPC, Karagandy AES). There are substantial differences between the breeding sites for both weather conditions and resulting grain yield.
The biplot analysis, using these three parameters (air temperature, precipitation, and yield) for ten individual years, provides additional information on the similarities and differences between the breeding sites (Figure 3). Clusters 1 and 2 are almost entirely composed of Astana and Karagandy breeding cites, confirming their similarity. Cluster 5 comprises five Aktobe years and one Karabalyk year. Karabalyk is grouped with North Kazakhstan AES in Cluster 3 and with Aktobe in Cluster 4. North Kazakhstan AES is the most variable for the three parameters, being present in four out of five clusters. This biplot confirmed the similarity between Kaz. Grain SPC and Karagandy AES. Aktobe AES was unique in some years, while in others it was similar to Karabalyk AES, which in turn was similar to North Kazakhstan AES, the most variable site among all.
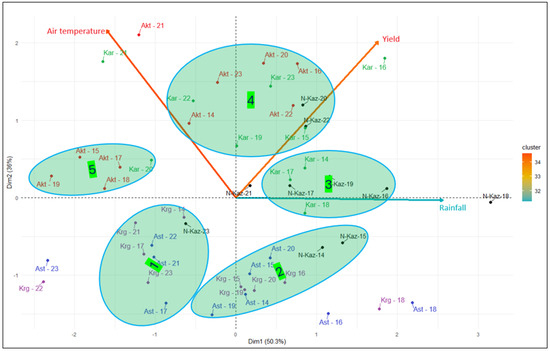
Figure 3.
Biplot analysis of five breeding programs for variation in average air temperature in May–August, precipitation in January–August, and spring wheat grain yield in 2014–2023 (Ast—Kaz. Grain SPC, Kar—Karabalyk AES, Krg—Karagandy AES, N-Kaz—North Kazakhstan AES; number refers to the year).
3.2. Variation for Agronomic Traits at Experimental Sites in 2022–2023
The weather conditions at experimental sites in 2022 were warmer and drier compared to the decade of 2014–2023. The amount of precipitation at Kaz. Grain SPC in January–August was only 55% of the decade average and 82% at Karabalyk AES. A similar lack of moisture occurred in 2023 at Kaz. Grain SPC and North Kazakhstan AES. The resulting average grain yield for 2022–2023 was 3118 kg/ha at Karabalyk AES, followed by 2652 kg/ha at North Kazakhstan AES and a yield of only around 1500 kg/ha at Astana and Karagandy (Table 2, Table S2).
Table 2.
Average agronomic traits for the breeding sites for 2022–2023 (mean site values designated with different letters are significantly different at p < 0.05).
The number of days from emergence to heading varied from 41.0 to 44.1 days, with a 2–3-day-shorter period at Karagandy AES (Table 2). The lowest plant height of 47.1 cm was at the driest site in Astana. Spring wheat at Karabalyk AES formed the second longest spike (8.64 cm) with a high number of grains per spike (26.6), lower spikelet fertility (1.78), and relatively high 1000-kernel weight (37.1 g). Spring wheat at North Kazakhstan AES was characterized by a shorter spike (6.87 cm), the highest spikelet fertility (2.44), and the largest grain (38.8 g). Kaz. Grain SPC and Karagandy AES had similar yields; however, the contribution of the yield components was significantly different. The former had the lowest spike fertility and grain size, as reflected by TKW, while at Karagandy AES, these two traits were the highest among all sites.
The rate of development is crucial for short-season spring wheat in Northern Kazakhstan. The differences in the average number of days from emergence to heading among genotypes varied within one week. All the material was divided into three groups based on the average number of days to heading across all sites and years (Table 3). Interestingly, the later-heading germplasm had significantly longer spikes, more spikelets, and more grains per spike. There was no difference in plant height and little difference between the three groups for TKW. Intermediate and late heading groups were significantly higher yielding compared to the early group. This relationship has important implications for the selection of high-yielding genotypes, which is best performed within the same heading/maturity group.
Table 3.
Agronomic traits of spring wheat germplasm across all sites and years varying in number of days from emergence to heading, 2022–2023 (mean values designated with different letters are significantly different at p < 0.05).
The results of ANOVA for grain yield demonstrated the high significance of the effects of the main factors (genotypes, sites, and years) and all interactions (Table S3). The similarity of germplasm performance at eight trials was evaluated by pairwise correlation coefficients of grain yield of 84 genotypes. In most cases, the correlation was below 0.3, indicating a lack of consistency in germplasm performance across sites and years, as well as high genotype × environment interaction. Results of only two trial combinations (out of 28) showed relatively high and significant correlation coefficients: Karabalyk AES-2022—North Kazakhstan AES-2023 (0.464) and Karabalyk AES-2023—Kaz Grain SPC-2022 (0.471). This once again signifies that germplasm performance for grain yield at four breeding programs is highly specific and variable between seasons.
3.3. Relative Performance of Spring Wheat Germplasm Originating from Different Breeding Programs
The relative performance of spring wheat material from five breeding programs for the key adaptation and yield traits when tested at four breeding sites is presented in Figure 4 and Figure 5 and Table S4. The germplasm originating from Karabalyk and North Kazakhstan AES consistently headed 1.5–2.0 days later compared to other breeding programs. There were no significant differences in plant height between the germplasm groups from different breeding programs. In fact, almost all material was tall, representing conventional extensive types of cultivars. For the number of grains per spike, material from Karabalyk and North Kazakhstan AES had an advantage for the overall performance. Spring wheat developed at Aktobe AES had the highest TKW, expressed at three out of four sites.
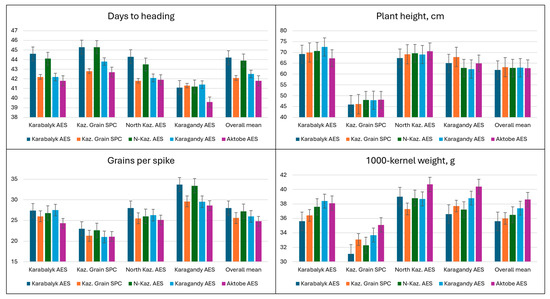
Figure 4.
Average values of adaptation and agronomic traits (2022–2023) of the germplasm originated from five breeding programs (columns with different colors) when tested at four breeding programs and across all sites (stacks of columns with the site written below).
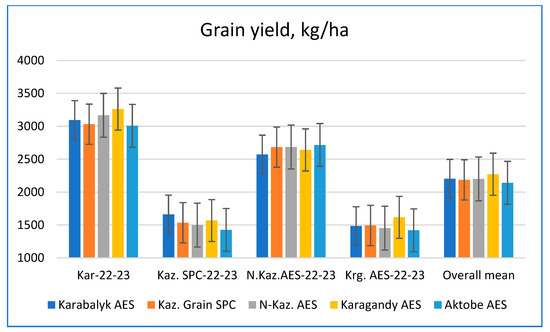
Figure 5.
The average grain yield (kg/ha) of the germplasm originated from five breeding programs (columns with different colors) when tested at four breeding programs and across all sites (stacks of columns with the site written below).
The severity of stem and leaf rust evaluated by the Kazakh Biosafety Research Institute showed minor differences between the germplasms originating from five breeding programs. The lowest average stem rust severity was demonstrated by the material from Kaz. Grain SPC (26.1%) and the highest by Aktobe AES (34.2%). For leaf rust, the lowest reading was for Karabalyk AES germplasm (19.1%) and the highest again was for Aktobe AES (25.4%). The frequency of rust-resistant genotypes was low in all breeding programs, indicating potential avenues for enhancing these traits.
The expectation for grain yield performance was that locally bred material would have an advantage over material developed at other breeding programs. Comparing mean value of the germplasm originating from different programs at four breeding sites did not show significant differences (Figure 5). However, the following tendencies were detected. The germplasm from Karagandy AES performed best when tested at Karabalyk AES over two years of study. The trial at Kaz. Grain SPC demonstrated a slight insignificant advantage of germplasm from Karabalyk AES. At North Kazakhstan AES, germplasm from Aktobe AES performed slightly better than other groups. At Karagandy AES, locally developed germplasm was higher yielding in both years. It also had a slight advantage for yield performance across all eight trials.
3.4. Genetic Diversity of Spring Wheat Germplasm Originating from Different Breeding Programs
Twenty KASP markers were polymorphic for the 84 studied genotypes. The average effective number of alleles was 1.509, and the percentage of polymorphic loci was 90.0% (Table 4). This is comparable to the genetic diversity of wheat germplasm from other regions and countries. Unbiased Nei’s genetic diversity index ranged from the lowest, 0.301 in Kaz Grain SPC, to the highest, 0.374 in the North Kazakhstan AES material. Genetic diversity of the germplasm from Karagandy and Aktobe AES was also lower compared to North Kazakhstan and Karabalyk AES.
Table 4.
Genetic diversity of 84 spring wheat accessions using 20 polymorphic KASP assays.
The principal coordinates analysis plot illustrated the relationships among breeding material with different origins (Figure 6). The two principal coordinates, PC1 and PC2, accounted for 60.1% and 24.8% of the total variation, respectively. Despite the geographic and environmental similarities of Kaz. Grain SPC and Karagandy AES, the germplasm from these two breeding programs was the most distant based on PCA1. In contrast, the germplasm from Aktobe and North Kaz AES, which have different climatic conditions, had a closer genetic relationship. Karabalyk AES material was placed apart from all other groups. The relationship between breeding sites varied significantly depending on the criteria used for classification: weather parameters and yield for the period 2014–2023 or molecular markers.
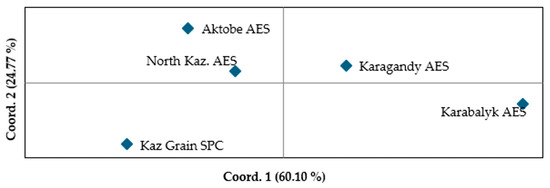
Figure 6.
Principal coordinate analysis based on Nei’s unbiased genetic distance for five groups of spring wheat germplasm originating from different breeding programs using 20 polymorphic KASP markers.
3.5. Molecular Marker Effects and Frequency of Alleles in Spring Wheat Germplasm Originating from Different Breeding Programs
The molecular markers of the genes used in the study were selected as having association with genes affecting specific traits. However, the analysis was made using all markers for all traits with expectation of multiple effects. Out of 20 KASP markers used, only seven were associated with the genes which demonstrated significant effects on the number of days to heading and grain yield (Table 5). However, the effects of the markers varied in value and direction depending on the site. Allele A of marker ipbb_ta_SH_162 had a significant negative effect on the heading date at Kaz. Grain SPC (−2.59%), a relatively large negative effect on grain yield at North Kazakhstan AES (−5.79%), and a large positive effect at Karagandy AES (+7.46%). Allele A of marker ipbb_SH_223, initially selected as linked to genes having effect on wheat development rate, had a positive significant effect on number of days to heading at three sites and a large negative effect on grain yield at Kaz. Grain SPC (−9.47%). For grain yield, the presence of the marker ipbb_PAxP_291 provided positive effects at all four sites, varying from 0.08% at North Kazakhstan AES to 8.07% at Kaz. Grain SPC. Initially this marker was selected for association with the genes affecting grain yield (Table 1).
Table 5.
Estimated effects of molecular marker SNP alleles associated with plant adaptation and yield-related components on number of days to heading and grain yield at four testing sites in 2022–2023.
The frequency of favorable alleles was calculated for each group of germplasm originating from five breeding programs (Table S5). There were large differences in the presence of specific alleles in different groups of material. Allele A of the marker ipbb_SH_223, which had a large negative effect on yield at Kaz Grain SPC (−9.47%), was absent in its germplasm. At the same time, allele A of the marker ipbb_PAxP_291, which has a large positive effect on yield at Kaz. Grain SPC (+8.07%), had a high frequency exceeding 93%.
The analysis was made to find out if the allele frequency was related to their effect on grain yield in the trials conducted at four experimental sites in 2022–2023. For each of the seven KASP markers listed above, the difference between the average allele frequency for all 84 genotypes and the frequency in the group of germplasm from a specific breeding program was calculated. This difference was plotted against the allele effect in a specific location. The assumption was that the allele with a negative effect on grain yield in a breeding program would have a lower frequency in the germplasm originating from this program. Conversely, alleles with positive effects on yield will be selected for and have a higher frequency in the respective germplasm. The results presented in Figure 7 demonstrate the higher frequency of alleles with a positive effect on grain yield in the germplasm originating from the site where this effect was observed. This important tendency demonstrates the reliability of the molecular markers used in the study and contributes to explaining the differences in performance among germplasm originating from different breeding programs.
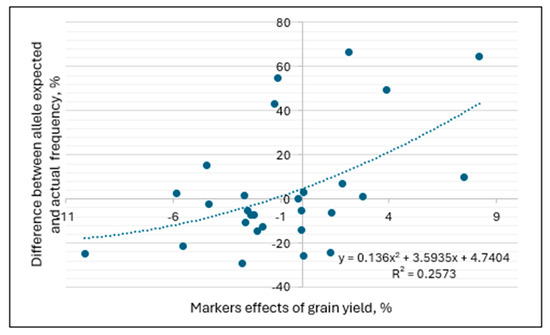
Figure 7.
Relationship between the effects of molecular markers and the allele frequencies in the germplasm tested and selected in four breeding programs.
3.6. High-Yielding Spring Wheat Genotypes Identified During the Study
The top 20% of highest-yielding genotypes based on performance across all sites and years were selected from each heading date group (early, intermediate, late) (Table 6). The absolute winner was cultivar Karagandinskaya 31 from the late group with 2617 kg/ha grain yield, or 6.6% higher than the second-best genotype 342-08 from the intermediate group developed at Kaz. Grain SPC (2453 kg/ha). All 17 superior genotypes were tall or intermediate for plant height. The differences for this trait were within 7–8 cm. The semidwarf stature trait has yet to be introduced into local cultivars and breeding material. The grain size, as measured by TKW, varied among the superior genotypes from 29.5 g (Bayterek 15) to 42.1 g (Lutescens 2231). However, the five highest-yielding genotypes with productivity exceeding 2420 kg/ha had an average TKW between 35.1 and 36.4 g.
Table 6.
Agronomic traits of high-yielding spring wheat germplasm from three maturity groups evaluated at four sites in 2022–2023.
Representation of the germplasm from five breeding programs in the highest-yielding material was as follows: Karagandy AES contributed eight genotypes; Kaz. Grain SPC—four; Karabalyk AES—three; North Kazakhstan AES and Aktobe AES—one each. Overall, the superior performance of Karagandy AES materials observed when comparing the average yield of germplasm from different breeding programs was supported by a larger representation of its material in the top yielders.
4. Discussion
Kazakhstan’s spring wheat production faces challenges associated with climate change, resulting in stagnating yields [2]. Enhancement of production technologies and cultivation of resilient varieties is a high priority to realize productivity gains and reduce the negative effects of climate change. More than 130 spring wheat varieties have been officially released in the country, although probably only 25–30 are cultivated on a commercial scale. A substantial area of spring wheat is cultivated using locally bred Kazakh cultivars, which are under pressure from the growing number of Russian and foreign cultivars being adopted by producers. Kazakhstan’s strategy is to strengthen local breeding and seed production to rely on locally bred cultivars as an important component of food security [15].
The current study evaluated the performance of spring bread wheat germplasm originating from five breeding programs to identify differences, elucidate the underlying factors, and provide guidance for future research. One important study outcome is the high degree of genotypes x environment (G x E) interaction most likely related to weather and soil conditions of the breeding sites. Abiotic and biotic stresses contribute to differential performance of spring wheat germplasm at different breeding sites [16]. High G x E raises a question of breeding strategy for broad versus specific adaptation. The Kazakhstan official state system [3] releases cultivars for individual administrative regions like Akmola, Kostanay, and North Kazakhstan in the main spring wheat zone. However, some unique cultivars occupy over 1 million ha across all the regions while others are grown on a limited scale in specific environments. There is value in additional studies and analysis using long-term multilocational data to identify main G x E drivers and breeding direction for broad versus specific adaptation.
Adaptation and agronomic performance of the germplasm from five breeding programs tested at four breeding sites can be summarized as follows: (a) Karagandy AES germplasm was better performing for grain yield not only at its “home” location but also at other sites and across sites; (b) Karabalyk AES material was a few days later heading, having the highest number of grains per spike and high yield at “home” and other sites; (c) Kaz. Grain SPC material had average grain yield but contributed two out of three top-yielding genotypes; (d) North Kazakhstan AES material was a few days later heading with a high number of grains per spike and average grain yield; (e) Aktobe AES germplasm was competitive for grain yield at North Kazakhstan AES and had the overall highest TKW. There is complementarity among the germplasm developed at different breeding programs.
The study underlined the deficiency of the germplasm for two important traits: semidwarf plant stature and rust resistance (both stem and leaf rust). These traits are highly important in the region due to the intensification of production and increasing pressure of diseases [17,18]. The lack of locally bred short-stature (60–80) wheat varieties has been demonstrated by the recent successful introduction of European cultivars with better response to inputs and higher rainfall. A growing number of farmers are equipped with modern sowing and harvesting machinery which slows down while harvesting tall varieties. The modern field technology requires shorter wheat cultivars. So far, none of the local spring wheat varieties released for Northern Kazakhstan possess the dwarfing genes and demonstrate short stature [19]. Newly released European cultivars can be used in crossing programs to reduce height. Sources of resistance to leaf and stem rust are available, pathogen populations have been studied, and genetics of resistance have been published [18].
The genetic diversity of spring wheat germplasm in this study, judged by variation in 20 KASP markers, was similar to diversity in other regions and countries [20,21]. The material from North Kazakhstan AES was by far the most diverse, taking into account common parameters such as the number of effective alleles and Nei’s unbiased diversity index.
Out of twenty KASP markers used, only seven demonstrated an association with the genes having a significant effect on days to heading and grain yield. The magnitude and direction of these effects varied depending on the experimental site and year. Some KASP markers initially selected for linkage with genes affecting other traits were effective for the same traits but also other agronomic traits, showing their multiple effects. Marker ipbb_SH_223 was validated in this study, having a consistent effect on number of days to heading and grain yield at one site. An important tendency was observed: the alleles with negative effects on grain yield at a specific site were in lower frequency in the germplasm developed at that site. This demonstrates the reliability of the markers and their potential for use in breeding programs, along with other spring wheat markers identified for Kazakhstan conditions [4,8,9,10,11,22]. Further actions may involve further validation of these markers and their use in practical breeding.
Several underlying factors affect the relative adaptation and performance of the germplasm from five Kazakh breeding programs. The main study hypothesis was that spring wheat germplasm developed at a particular breeding program would be better adapted to the conditions where it was developed. The analysis of air temperature, precipitation, and spring wheat grain yield over the last ten years revealed differences and similarities between the breeding programs. Kaz. Grain SPC and Karagandy AES were quite similar, but their germplasm performance differed for yield and other traits. A high degree of genotype × environment interaction was observed in the study, and even the performance of the material at the same site in consecutive years differed. This is an important consideration for spring wheat breeding in the region [5]. The clustering of the breeding programs’ germplasm according to KASP marker diversity differed from the clustering based on weather traits and grain yield. There is no clear relation between germplasm adaptation, performance, and diversity and the ecological and agronomic variables of the site where the germplasm was developed. Similar and even larger-scale cooperative breeding nurseries are employed in North America for winter wheat like South and Northern Regional Performance Nurseries [23]. They have been used for evaluation of adaptation, quality, and specific traits including molecular markers. However, they have not been studied for factors contributing to adaptation and performance of individual breeding programs.
The important factors driving germplasm performance are related to the breeding program’s strategy, methodology, parentage used, selection procedures, experimental design, phenotyping precision and scale, generation advance speed, and the use of genomic and other tools and approaches, making the breeding programs productive and resulting in successful cultivars [24,25]. Five breeding programs participating in this study employ conventional wheat improvement, with limited or no use of modern phenomic and genomic tools. Hence, their germplasm performance and success depend more on the breeder’s intuition and application of robust screening procedures.
5. Conclusions
The current spring wheat breeding framework in Kazakhstan requires fundamental revision to remain competitive. Firstly, the application of modern tools and approaches requires rapid introduction and adoption. These may include marker-assisted selection and the first steps have been undertaken already, including in this study. Genomic selection is to be designed and tested in the breeding programs. Phenotyping precision and scale using modern physiological approaches and tools is worth consideration. The first speed breeding platform has been established in Kazakhstan and needs integration into practical wheat breeding. Secondly, this study demonstrated the power of multilocation testing in characterizing environments and genotypes. Closer cooperation between the programs is well justified, targeting coordinated crossing programs, speed breeding, and other cooperative tools. Thirdly, the utilization of novel genetic resources, especially in addressing challenging traits, is well justified. Recent cultivar Aina from Karabalyk AES is a good example. It was developed through a shuttle breeding program with CIMMYT, utilizing semidwarf disease-resistant parents. Aina proved resistant to stem rust during the 2018 epidemic and was high-yielding, reaching over 1 million hectares by 2024. International cooperation with advanced research and breeding institutions in the country and abroad is crucial for maintaining competitiveness and advancing genetic progress.
The potential of Kazakhstan in spring wheat production and its contribution to national and regional food security can only be utilized with modern and relevant cultivars developed by competitive local programs.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/crops5050063/s1. Table S1: List of material used in the study. Table S2: Agronomic traits across experimental sites in 2022–2023; Table S3: ANOVA results for grain yield.; Table S4: Mean agronomic traits of spring wheat germplasm originating from different breeding programs across experimental sites in 2022 and 2023; Table S5. SNP alleles frequency in spring wheat germplasm originating from five breeding programs.
Author Contributions
Conceptualization, T.S., Y.T. and A.M. (Alexey Morgounov); methodology, A.A., L.Z. and A.M. (Alexey Morgounov); software, D.T.; formal analysis, D.T., A.A. and S.A.; investigation, A.A., S.A., A.B., V.C., E.F., Y.K., A.M. (Akerke Maulenbay), G.S., S.S., V.T., A.T. and L.Z.; resources, T.S. and Y.T.; data curation, Y.K. and A.M. (Alexey Morgounov); writing—original draft preparation, A.M. (Alexey Morgounov); writing—review and editing, T.S. and Y.T.; funding acquisition, T.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research has been funded by the Ministry of Agriculture of the Republic of Kazakhstan BR24892821 “Breeding, seed production of grain crops to increase the potential of productivity, quality, stress resistance in various soil-climatic zones of Kazakhstan”.
Data Availability Statement
The study data is available from the corresponding author upon request.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| AES | Agricultural Experimental Station |
| SPC | Scientific Production Center |
References
- Anonymous. Eighth National Communication and Fifth Biennial Report of the Republic of Kazakhstan to the UN Framework Convention on Climate Change; Ministry of Ecology, Geology and Natural Resources: Astana, Kazakhstan, 2022; 473p. [Google Scholar]
- World Bank. Republic of Kazakhstan: Climate Adaptation Options and Opportunities in the Agriculture Sector; World Bank Group: Washington, DC, USA, 2024; 75p. [Google Scholar]
- Azhgaliev, T.B. State Register of Breeding Achievements Recommended for Use in the Republic of Kazakhstan; Ministry of Agriculture: Nursultan, Kazakhstan, 2022; pp. 7–12. [Google Scholar]
- Morgounov, A.; Babkenov, A.; Ben, C.; Chudinov, V.; Dolinny, Y.; Dreisigacker, S.; Fedorenko, E.; Gentzbittel, L.; Rasheed, A.; Savin, T. Molecular markers help with breeding for agronomic traits of spring wheat in Kazakhstan and Siberia. Genes 2024, 15, 86. [Google Scholar] [CrossRef] [PubMed]
- Tajibayev, D.; Mukin, K.; Babkenov, A.; Chudinov, V.; Dababat, A.A.; Jiyenbayeva, K.; Kenenbayev, S.; Savin, T.; Shamanin, V.; Tagayev, K. Exploring the agronomic performance and molecular characterization of diverse spring durum wheat germplasm in Kazakhstan. Agronomy 2023, 13, 1955. [Google Scholar] [CrossRef]
- Pask, A.J.D.; Pietragalla, J.; Mullan, D.M.; Reynolds, M.P. Physiological Breeding II: A Field Guide to Wheat Phenotyping; CIMMYT: Ciudad de México, Mexico, 2012; 140p. [Google Scholar]
- Dellaporta, S.L.; Wood, J.; Hicks, J.B. A plant DNA minipreparation: Version II. Plant Mol. Biol. Rep. 1983, 1, 19–21. [Google Scholar] [CrossRef]
- Turuspekov, Y.; Baibulatova, A.; Yermekbayev, K.; Tokhetova, L.; Chudinov, V.; Sereda, G.; Ganal, M.W.; Griffiths, S.; Abugalieva, S. GWAS for plant growth stages and yield components in spring wheat (Triticum aestivum L.) harvested in three regions of Kazakhstan. BMC Plant. Biol. 2017, 17, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Amalova, A.; Abugalieva, S.; Chudinov, V.; Sereda, G.; Tokhetova, L.; Abdikhalyk, A.; Turuspekov, Y. QTL mapping of agronomic traits in wheat using the UK Avalon× Cadenza reference mapping population grown in Kazakhstan. PeerJ 2021, 9, 10733. [Google Scholar] [CrossRef] [PubMed]
- Amalova, A.; Abugalieva, S.; Babkenov, A.; Babkenova, S.; Turuspekov, Y. Genome-wide association study of yield components in spring wheat collection harvested under two water regimes in Northern Kazakhstan. PeerJ 2021, 9, 11857. [Google Scholar] [CrossRef] [PubMed]
- Amalova, A.; Yermekbayev, K.; Griffiths, S.; Abugalieva, S.; Babkenov, A.; Fedorenko, E.; Abugalieva, A.; Turuspekov, Y. Identification of quantitative trait loci of agronomic traits in bread wheat using a Pamyati Azieva × Paragon mapping population harvested in three regions of Kazakhstan. PeerJ 2022, 10, 14324. [Google Scholar] [CrossRef] [PubMed]
- Semagn, K.; Babu, R.; Hearne, S.; Olsen, M. Single nucleotide polymorphism genotyping using Kompetitive Allele Specific PCR (KASP): Overview of the technology and its application in crop improvement. Mol. Breed. 2014, 33, 1–14. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017; 245p. [Google Scholar]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research—An update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [PubMed]
- Mukhametzhanov, A.; Zholaman, R. Economic analysis of spring soft wheat seed production in North Kazakhstan region. Sci. Horiz. 2023, 26, 92–100. [Google Scholar] [CrossRef]
- Kushenbekova, A.K.; Mukhomedyarova, A.S. Productivity of spring wheat varieties depending on climatic conditions of the dry steppe zone of western Kazakhstan. Sci. Edu. 2022, 66, 33–40. [Google Scholar]
- Goncharov, N.P. Scientific support to plant breeding and seed production in Siberia in the XXI century. Vavilov J. Genet. Breed. 2021, 25, 448–459. [Google Scholar] [CrossRef] [PubMed]
- Kelbin, V.N.; Skolotneva, E.S.; Shamanin, V.P.; Salina, E.A. Diversity of stem rust resistance in modern Siberian bread wheat (Triticum aestivum) germplasm. Plant Breed. 2022, 141, 194–203. [Google Scholar] [CrossRef]
- Bazilova, D.S.; Dolinnyi, Y.Y.; Ivanova, G.N. Initial material for breeding of spring soft wheat in the conditions of northern Kazakhstan. KazNAU Res. Results 2022, 94, 37–46. [Google Scholar]
- Ren, J.; Sun, D.; Chen, L.; You, F.M.; Wang, J.; Peng, Y.; Nevo, E.; Sun, D.; Luo, M.-C.; Peng, J. Genetic diversity revealed by single nucleotide polymorphism markers in a worldwide germplasm collection of durum wheat. Int. J. Mol. Sci. 2013, 14, 7061–7088. [Google Scholar] [CrossRef] [PubMed]
- Semagn, K.; Iqbal, M.; Alachiotis, N.; N’Diaye, A.; Pozniak, C.; Spaner, D. Genetic diversity and selective sweeps in historical and modern Canadian spring wheat cultivars using the 90K SNP array. Sci. Rep. 2021, 11, 23773. [Google Scholar] [CrossRef] [PubMed]
- Genievskaya, Y.; Abugalieva, S.; Rsaliyev, A.; Yskakova, G.; Turuspekov, Y. QTL mapping for seedling and adult plant re-sistance to leaf and stem rusts in Pamyati Azieva × Paragon mapping population of bread wheat. Agronomy 2020, 10, 1285. [Google Scholar] [CrossRef]
- Boehm, J.D.; Masterson, S.; Palmer, N.; Cai, X.; Miguez, F. Genetic improvement of winter wheat (Triticum aestivum L.) grain yield in the Northern Great Plains of North America, 1959–2021. Crop Sci. 2023, 63, 3236–3249. [Google Scholar] [CrossRef]
- Bapela, T.; Shimelis, H.; Tsilo, T.J.; Mathew, I. Genetic improvement of wheat for drought tolerance: Progress, challenges and opportunities. Plants 2022, 11, 1331. [Google Scholar] [CrossRef] [PubMed]
- Aidarbekova, T.J.; Syzdykova, G.T.; Malitskaya, N.V.; Nurgaziyev, R.E.; Husainov, A.T.; Zhabayeva, M.U.; Makhanova, S.K.; Shoykin, O.D. Comparative assessment of spring soft wheat lines (Triticum aestivum L.) in the steppe zone of the north Kazakhstan region. Agric. Biol. 2022, 57, 66–80. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).