
The Morphological and Ecogeographic Characterization of the Musa L. Collection in the Gene Bank of INIAP, Ecuador

 ,
,  ,
,  ,
,  , ,
, ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Morphological Characterization
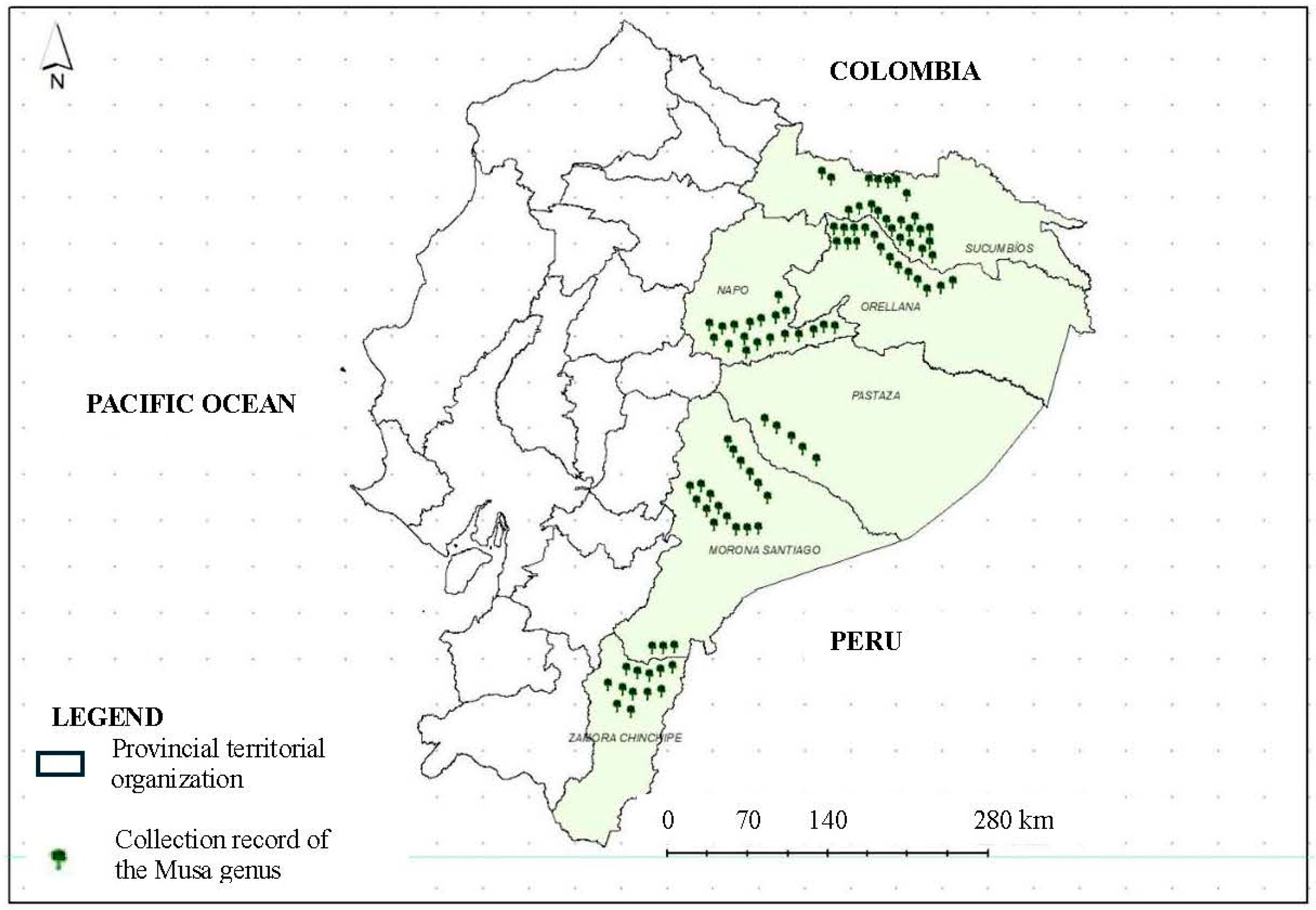
2.2. Ecogeographic Characterization
3. Results
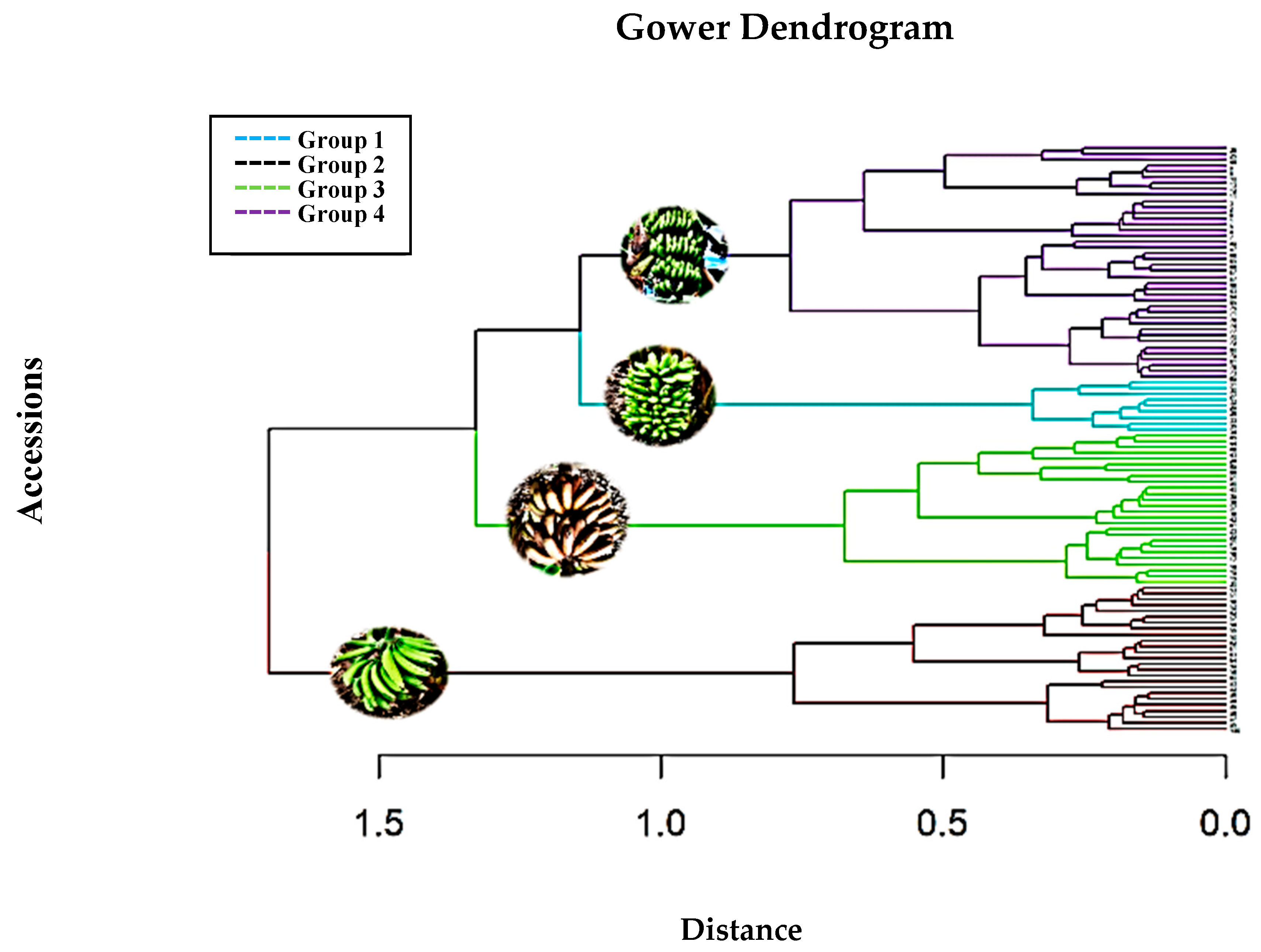
3.1. Grouping of Accessions Based on Morphological Variables
3.2. Combined Analysis of Qualitative and Quantitative Variables
3.3. Grouping of Accessions Based on Quantitative Morphological Characters
3.4. Grouping of Accessions Based on Qualitative Morphological Characters
3.4.1. Plant and Pseudostem
3.4.2. Leaves
3.4.3. ♂ Flower
3.4.4. Bracts
3.4.5. ♀ Flower
3.4.6. Raceme
3.4.7. Fruit
3.5. Discriminating Capacity of Descriptors
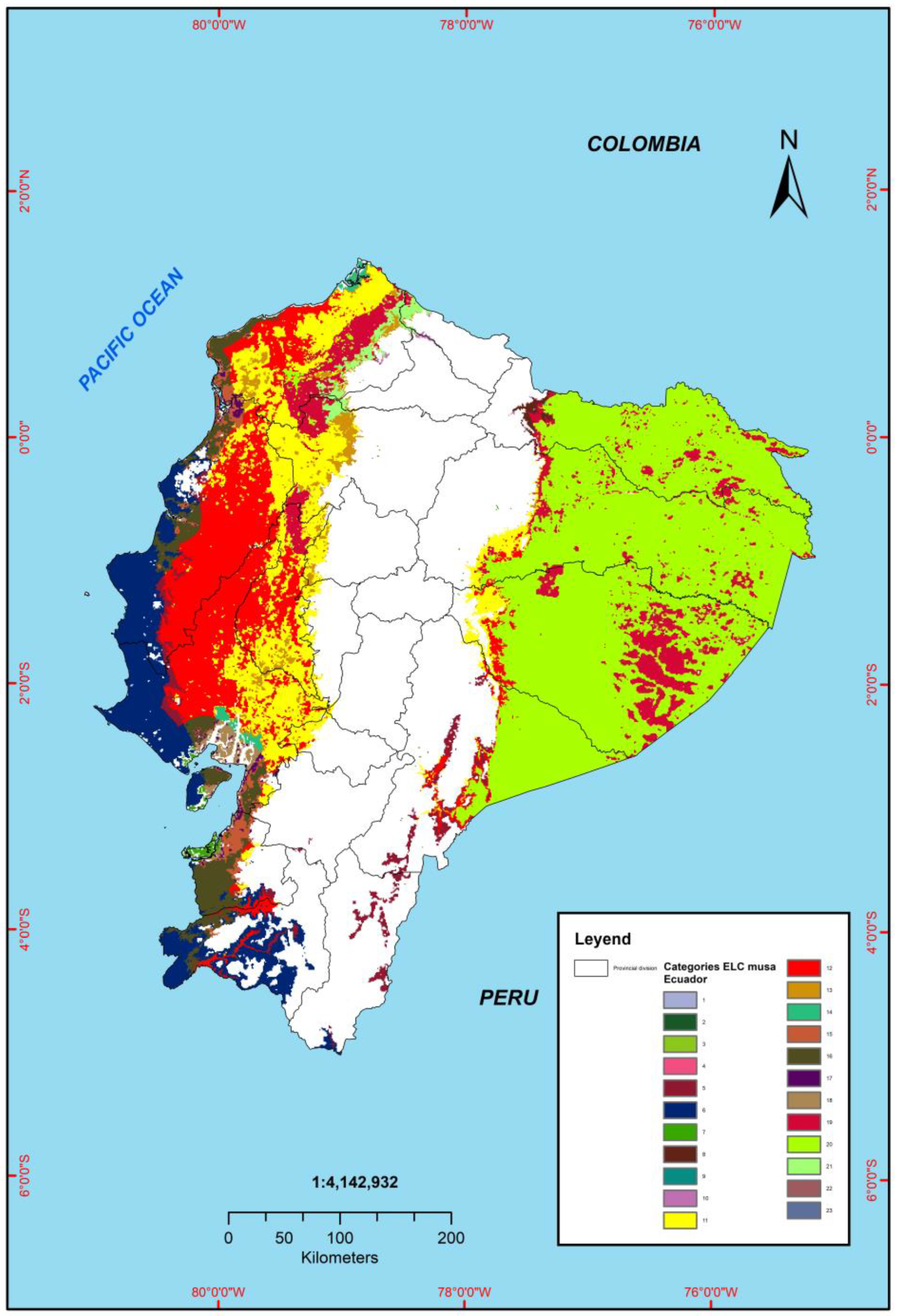
3.6. Evaluation of the Ecogeographic Diversity of Musa Using the ELC Map
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De Langhe, E.; Vrydaghs, L.; de Maret, P.; Perrier, X.; Denham, T. Why Bananas Matter: An introduction to the history of banana domestication. Ethnobot. Res. Appl. 2009, 7, 165–177. [Google Scholar] [CrossRef]
- Campos, H.; Caligari, P.D.; Brown, A.; Tumuhimbise, R.; Amah, D.; Uwimana, B.; Swennen, R. Bananas and plantains (Musa spp.). In Genetic Improvement of Tropical Crops; Campos, H., Caligari, P.D.S., Eds.; Springer International Publishing AG: Berlin/Heidelberg, Germany, 2017; pp. 219–240. [Google Scholar]
- Vásquez, R.; Romero, A.; Figueroa, J. Paquete Tecnológico para el Cultivo de Plátano; Gobierno del Estado de Colima: Colima, Mexico, 2005; N° 001; 72p. [Google Scholar]
- Cayón, S.D.G. (Ed.) Post Cosecha y Agroindustria del Plátano en el eje Cafetero de Colombia; CORPOICA: Cali, Colombia, 2000; 145p, ISBN 9589688500. [Google Scholar]
- Debnath, S.; Khan, A.A.; Das, A.; Murmu, I.; Khan, A.; Mandal, K.K. Genetic Diversity in Banana. In Genetic Diversity in Horticultural Plants. Sustainable Development and Biodiversity; Nandwani, D., Ed.; Springer: Cham, Switzerland, 2019; Volume 22, pp. 217–241. [Google Scholar] [CrossRef]
- Robinson, J.; Galán Saúco, V. Banana and Plantains, 2nd ed.; CAB International: Cambridge, UK, 2010; 297p, ISBN 978-1-84593-658-7. [Google Scholar]
- Power, R.C.; Güldemann, T.; Crowther, A.; Boivin, N. Asian Crop Dispersal in Africa and Late Holocene Human Adaptation to Tropical Environments. J. World Prehist. 2019, 32, 353–392. [Google Scholar] [CrossRef]
- Salas-Pascual, M.; Cáceres-Lorenzo, T. The Dispersal of Bananas (Musa spp.) to the Americas in the Sixteenth Century. Econ. Bot. 2022, 76, 354–367. [Google Scholar] [CrossRef]
- Guerrero, S. Características Morfométricas de Cultivares de Musáceas Establecidos en la Finca Experimental “La Maria”. Pregrade Project Thesis. Quevedo State Technical University: Quevedo, Ecuador, 2016. 93p. Available online: https://repositorio.uteq.edu.ec/handle/43000/3259 (accessed on 22 February 2025).
- MAG [Ministerio de Agricultura y Ganadería]. Boletín Situacional Cultivo de Banano. 2022. Available online: http://sipa.agricultura.gob.ec/index.php/platano/boletines-situacionales-platano-ecuador (accessed on 22 February 2025).
- INEC [Instituto Nacional de Estadística y Censos]. Boletín Técnico Encuesta de Superficie y Producción Agropecuaria Continua (ESPAC). 2023. Available online: https://www.ecuadorencifras.gob.ec/estadisticas-agropecuarias-2/ (accessed on 22 February 2025).
- Engels, J.M.M.; Visser, L. (Eds.) Guía para el manejo eficaz de un banco de germoplasma. In Manuales para Bancos de Germoplasma; Bioversity International: Rome, Italy, 2007; Volume 6, 192p, Available online: https://cropgenebank.sgrp.cgiar.org/images/file/learning_space/genebankmanual6_spa.pdf (accessed on 22 February 2025).
- Travez Guanotuña, X.D. Creación de Bancos Locales de Semillas Ancestrales como Estrategia para Enfrentar el Cambio Climático en la Parroquia de Chugchilán e Isinliví. Thesis Ing. Ambiental [BSc Environment]. Universidad Técnica de Cotopaxi: Latacunga, Ecuador, 2024. 94p. Available online: https://repoadmin.utc.edu.ec/bitstreams/054a25a5-8f31-431c-9adb-56801ac4dbad/download (accessed on 24 February 2025).
- MGIS [Musa Germplasm Information System]. 2023. Available online: https://www.crop-diversity.org/mgis/ (accessed on 28 May 2025).
- Cedeño Cedeño, L.M. El Boom Bananero y la Movilidad Humana en la Sierra Centro Ecuatoriana en el Período 1948-1965. Bachelor’s Thesis, Universidad Central del Ecuador, Quito, Ecuador, 2017; 103p. Repositorio UCE. Available online: http://www.dspace.uce.edu.ec/handle/25000/13992 (accessed on 24 February 2025).
- Jiffy Growing Solutions. ¿Cuáles Son los Desafios del Cultivo de Plátano? Available online: https://jiffygroup.com/es/noticias/cuales-son-los-desafios-del-cultivo-de-platano/Torres (accessed on 6 May 2025).
- Torres Jaramillo, L.A.; Centanaro Quiroz, P.H.; Raffo Folleco, L.A.; Nava Luzardo, J.C. Factores Limitantes del Desarrollo Agroecológico en el Cultivo de Banano (Musa AAA), Provincia del Guayas, Ecuador. Rev. Técnica Fac. Ing. Univ. Zulia 2023, 46, e234601. [Google Scholar] [CrossRef]
- Hernandez Villarreal, A.E. Caracterización morfológica de recursos fitogenéticos. Rev. Bio Cienc. 2013, 2, 113–118. Available online: https://revistabiociencias.uan.edu.mx/index.php/BIOCIENCIAS/article/view/41/133 (accessed on 24 February 2025).
- Santo-Pineda, U.; Torres-Vargas, L.; Santamaria-Guerra, J.; Thomas, G.; Montezuma, V. Caracterización morfológica de 21 Accesiones de musáceas colectadas en la Comarca Ngäbe-Buglé, Panamá. Cienc. Agropecu. 2024, 38, 7–26. Available online: http://revistacienciaagropecuaria.ac.pa/index.php/ciencia-agropecuaria/article/view/624 (accessed on 24 February 2025).
- Parra-Quijano, M.; Iriondo, J.M.; Torres, E. Review. Applications of ecogeography and geographic information systems in conservation and utilization of plant genetic resources. Span J. Agric. Res. 2012, 10, 419–429. [Google Scholar] [CrossRef]
- Elbehri, A.; Calberto, G.; Staver, C.; Hospido, A.; Roibas, L.; Skully, D.; Siles, P.; Arguello, J.; Sotomayor, I.; Bustamante, A. Ecuador’s Banana Sector Under Climate Change—An Economic and Biophysical Assessment to Promote a Sustainable and Climate-Compatible Strategy; Food and Agriculture Organization of the United Nations [FAO]: Rome, Italy, 2016; 164p, ISBN 978-92-5-109249-1. [Google Scholar]
- Cedeño-Zambrano, J.R.; García-Párraga, J.V.; Solórzano-Cobeña, C.M.; Jiménez-Flores, L.A.; Ulloa-Cortazar, S.M.; López-Mejía, F.X.; Avellán-Vásquez, L.E.; Bracho-Bravo, B.; Sánchez-Urdaneta, A.B. Fertilización con magnesio en plátano Barraganete (Musa AAB) Ecuador. Granja Rev. Cienc. Vida 2022, 35, 8–19. Available online: https://lagranja.ups.edu.ec/pdf/granja/fertilizante_platano_esp.pdf (accessed on 26 February 2025). [CrossRef]
- Torres, S.J.P.; Cedeño, F.J.A.; Segura, R. Estado actual del mejoramiento genético en el cultivo del banano (Musa AAA, subgrupo Cavendish), y plátano (Musa AAB). Acorbat Rev. Tecnol. Cienc. 2024, 1, 2. [Google Scholar] [CrossRef]
- Beaton, K.; Mazadza, A.; Chikwambi, Z. Identificación de cultivares locales de banano (Musa spp.) en Zimbabue mediante morfología y secuenciación genómica. J. Genet. Eng. Biotechnol. 2023, 21, 118. [Google Scholar] [CrossRef]
- Monteros-Altamirano, A.; Tacán, M.; Peña Monserrate, G.R.; Tapia, B.C.; Paredes, N.; Lima, L. Guía para el Manejo y Conservación de Recursos Fitogenéticos en Ecuador: Protocolos; INIAP, Estación Experimental Santa Catalina, Departamento Nacional de Recursos Fitogenéticos: Mejía, Ecuador, 2018; Volume 432, pp. 1–104, Publicación Miscelánea; Available online: https://repositorio.iniap.gob.ec/bitstream/41000/4889/1/iniapscpm432.pdf (accessed on 26 February 2025).
- Asociación de Exportadores de Banano [AEBE]. Guía Culinaria del Banano—Banana Culinary Guide; Imprenta Mariscal: Guayaquil, Ecuador, 2021; 111p, ISBN 978-9942-962-09-6. Available online: https://www.aebe.com.ec/_files/ugd/f4cd67_21aef8d33457473f997eea1dc297f638.pdf?index=true (accessed on 26 February 2025).
- FAO. Banano. In Análisis del Mercado 2023; FAO: Rome, Italy, 2023; 10p, Available online: https://openknowledge.fao.org/server/api/core/bitstreams/318cda0c-dae9-4c24-a8cb-44afd40b90c3/content (accessed on 26 February 2025).
- León Ajila, J.P.; Espinosa Aguilar, M.A.; Carvajal Romero, H.R.; Quezada Campoverde, J. Análisis de la producción y comercialización de banano en la provincia de El Oro en el periodo 2018–2022. Cienc. Lat. 2023, 7, 7494–7507. [Google Scholar] [CrossRef]
- Tapia, C.; Paredes, N.; Lima, L. Representatividad de la diversidad del género Musa en el Ecuador. Repos. Digit. INIAP 2019, 6, 53–58. [Google Scholar]
- FAO [Food and Agriculture Organization of the United Nations]. Medium-Term Outlook: Prospects for World Production and Trade in Bananas and Tropical Fruits 2019–2028; FAO: Rome, Italy, 2020; Available online: https://www.fao.org/home/en/ (accessed on 26 February 2025).
- IPGRI/INIBAP/CIRAD [International Plant Genetic Resources Institute (IPGRI); International Network for the Improvement of Banana and Plantain (INIBAP); Centre de Cooperation Internationale en Recherche Agronomique pour le Developpement (CIRAD)]. Descriptores para el Banano (Musa spp.); International Plant Genetic Resources Institute: Rome Italy, 1996; 55p, Available online: https://alliancebioversityciat.org/es/node/3726 (accessed on 26 February 2025).
- Royal Horticultural Society (RHS). The RSH Colour Chart, 6th ed.; Royal Horticultural Society (RHS): London, UK, 2007; Available online: https://www.rhs.org.uk/ (accessed on 26 February 2025).
- Infostat. Infostat—Software Estadístico. 2020. Available online: https://www.infostat.com.ar/ (accessed on 28 February 2025).
- Flores, C.; Salomé, D. Estudio de la Diversidad Morfológica y Eco-Geográfica en tres Cultivos Andinos del Ecuador: El caso del Tarwi (Lupinus mutabilis Sweet.), Jícama (Smallanthus sonchifolius (Poepp. & Endl.) H.Robinson), y Miso (Mirabilis expansa Ruiz & Pav. Standley.); Universidad de Santiago de Compostela: España, Spain, 2021; Available online: http://hdl.handle.net/10347/27229 (accessed on 28 February 2025).
- Parra-Quijano, M.; Iriondo, J.M.; Torres, M.E.; López, F.; Maxted, N.; Kell, S.P. Capftogen3: A Toolbox for the Conservation and Promotion of the Use of Agricultural Biodiversity; Universidad Nacional de Colombia: Bogotá, Colombia, 2021; p. 45194. Available online: https://repositorio.unal.edu.co/handle/unal/85787 (accessed on 28 February 2025).
- Crespo, C.F.; Guanochanga, J. Diseño de un Proceso para la Obtención de Pulpa Congelada a Partir de Orito (Musa acuminata AA). Rev. Cienc. Tecnol. 2022, 22. [Google Scholar] [CrossRef]
- Jiménez-Esparza, L.O.; Decker-Campuzano, F.E.; González-Parra, M.M.; Mera-Andrade, R. Abonos orgánicos una alternativa en el desarrollo de cormos de orito (Musa acuminata AA). J. Selva Andina Biosph. 2019, 7, 54–62. [Google Scholar] [CrossRef]
- Nadal-Medina, R.; Manzo-Sánchez, G.; Orozco-Romero, J.; Orozco-Santos, M.; Guzmán-González, S. Diversidad genética de bananos y plátanos (Musa spp.) determinada mediante marcadores rapd/Genetic diversity of bananas and plantains (Musa spp.) determined by rapd markers. Rev. Fitotec. Mex. 2009, 32, 1–7. [Google Scholar]
- Ukwueze, C.K.; Oselebe, H.O.; Igwe, D.O.; Mekonnen, T.B. Morphological characterization and genetic diversity assessment of accessions of Musa spp. Res. Sq. 2024, preprint. [Google Scholar] [CrossRef]
- Christelová, P.; De Langhe, E.; Hřibová, E.; Čížková, J.; Sardos, J.; Hušáková, M.; Van den Houwe, I.; Sutanto, A.; Kepler, A.K.; Swennen, R.; et al. Molecular and cytological characterization of the global Musa germplasm collection provides insights into the treasure of banana diversity. Biodivers. Conserv. 2017, 26, 801–824. [Google Scholar] [CrossRef]
- Sampangi-Ramaiah, M.H.; Ravishankar, K.V.; Shivashankar, K.S.; Roy, T.K.; Rekha, A.; Hunashikatti, L.R. Developmental changes in the composition of leaf cuticular wax of banana influenced by wax biosynthesis gene expression: A case study in Musa acuminata and Musa balbisiana. Acta Physiol. Plant 2019, 41, 141. [Google Scholar] [CrossRef]
- Chaves-Barrantes, N.F.; Gutiérrez-Soto, M.V. Respuestas al estrés por calor en los cultivos. II. Tolerancia y tratamiento agronómico. Agron. Mesoam. 2016, 28, 255–271. [Google Scholar] [CrossRef]
- Caicedo Arana, Á. Caracterización y Evaluación Morfológica, Física y Química de Introducciones del Banco de Germoplasma de Musáceas en el Centro de Investigación Corpoica Palmira. Master’s Thesis, Universidad Nacional de Colombia, Palmira, Colombia, 2015. Available online: https://repositorio.unal.edu.co/bitstream/handle/unal/58986/Alvaro_Caicedo_Arana.pdf?sequence=1&isAllowed=y (accessed on 12 March 2025).
- Deras, M.; Durán, L.F.; Mercadal, L.; Rivera, M. Avances del proyecto evaluación y diseminación de híbridos de Musa con resistencia a Sigatoka Negra. In Informe Técnico Programa de Banano y Plátano 2003; FHIA, Ed.; Federación Hondureña de Investigaciones Agrícolas: La Lima, Honduras, 2004; pp. 45–58. [Google Scholar]
- Aldana Leyva, F.; Fernández Martínez, O.; García-Águila, L.; Sarría, Z.; Hurtado Ribalta, O. Respuesta agronómica de plantas de banano cultivar ‘FHIA-17’ (Musa AAAA) obtenidas por cultivo de tejidos y por propagación agámica. Biotecnol. Veg. 2020, 20, 83–91. [Google Scholar]
- Dela Cruz, F.S.; Gueco, L.S.; Damasco, O.P.; Huelgas, V.C.; dela Cueva, F.M.; Dizon, T.O.; Sison, M.L.J.; Banasihan, I.G.; Sinohin, V.O.; Molina, A.B., Jr. Farmers’ Handbook on Introduced and Local Banana Cultivars in the Philippines; Bioversity International: Rome, Italy, 2008; 68p, Available online: https://www.researchgate.net/publication/317932686_Farmer%27s_handbook_on_introduced_and_local_banana_cultivars_in_the_Philippines (accessed on 28 February 2025).
- FHIA [La Fundación Hondureña de Investigación Agricola]. ‘FHIA-17’ Programa de Bananos y Plátanos; Instituto Patria: La Lima, Honduras, 2016; Available online: http://www.fhia.org.hn (accessed on 20 April 2019).
- Torres-Cabrera, D.; García-Águila, L.; Bermúdez-Caraballoso, I.; Sarría, Z.; Hurtado Ribalta, O.; Delgado, E.; Pérez, A.; Fernández Martínez, O. Respuesta morfo-agronómica y organoléptica de cinco cultivares de banano (Musa spp.) en condiciones de campo. Biotecnol. Veg. 2020, 20, 43–50. [Google Scholar]
- Hernández, Y.; Marín, M.; García, J. Respuesta en el rendimiento del plátano (Musa AAB cv. Hartón) en función de la nutrición mineral y su ciclo fenológico Parte, I. Crecimiento y producción. Rev. Fac. Agron. 2007, 24, 607–626. [Google Scholar]
- Brenes-Gamboa, S. Production and quality parameters of three banana cultivars FHIA-17, FHIA-25 and Yangambi. Agron. Mesoam. 2017, 28, 719–733. [Google Scholar] [CrossRef]
- Rodríguez, C.; Cayón, G.D.; Mira, J.J. Effect of number of functional leaves at flowering on yield of banana Grand Naine (Musa AAA Simmonds). Rev. Fac. Nac. Agron. Medellín 2012, 65, 6591–6597. [Google Scholar]
- Martínez, A.M.; Cayón, D.G. Dinámica del crecimiento del banano (Musa AAA, Simmons cvs. ‘Grande naine’ y Valery). Rev. Fac. Nal. Agr. Medellín 2011, 64, 6055–6064. [Google Scholar]
- Chundawat, B.S.; Patel, N.L.; Dave, S.K.; Tikka, S.B.S. Quantitative variability for yield and other characters in banana. Indian J. Hortic. 1988, 45, 8–12. [Google Scholar]
- Nayar, N.K.; Lyla, K.R.; Mathew, V. Genetic variability in dessert type of banana. Indian J. Agric. Sci. 1979, 49, 414–416. [Google Scholar]
- Rodríguez Cabriales, J.C. Estimación de la Variación Somaclonal y la Inducida por Rayos Gamma en Banano (Musa spp.): Determinación de Índices para el Proceso de Selección. Ph.D. Thesis, Universidad Autónoma de Nuevo León, San Nicolás de los Garza, Mexico, 2000. Available online: https://cdigital.dgb.uanl.mx/te/1080110328.PDF (accessed on 28 February 2025).
- Hoyos-Leyva, J.D.; Jaramillo-Jiménez, P.A.; Giraldo-Toro, A.; Dufour, D.; Sánchez, T.; Lucas-Aguirre, J.C. Caracterización física, morfológica y evaluación de las curvas de empastamiento de musáceas (Musa spp.). Acta Agronómica 2012, 61, 214–229. Available online: http://www.scielo.org.co/scielo.php?script=sci_arttext&pid=S0120-28122012000300003&lng=en&tlng=es (accessed on 11 March 2025).
- Smith, M.K.; Langdon, P.W.; Pegg, K.G.; Daniells, J.W. Growth, yield and Fusarium wilt resistance of six FHIA tetraploid bananas (Musa spp.) grown in the Australian subtropics. Sci. Hortic. 2014, 170, 176–181. [Google Scholar] [CrossRef]
- Vuylsteke, D.R.; Ortiz, R.; Shaun, R.; Ferris, B.; Crouch, J.H. Plantain improvement. Plant Breed. Rev. 1997, 14, 267–320. [Google Scholar]
- Buitrago-Bitar, M.A.; Enríquez-Valencia, A.L.; Londoño-Caicedo, J.M.; Muñoz-Flórez, J.E.; Villegas-Estrada, B.; Santana-Fonseca, G.E. Molecular and morphological characterization of Musa spp. (Zingiberales: Musaceae) cultivars. Boletín Científico Cent. Mus. Mus. Historia Natural 2020, 24, 33–47. [Google Scholar] [CrossRef]
- Dufour, D.; Giraldo, A.; Gibert, O.; Sánchez, T.; Reynes, M.; González, A.; Fernández, A.; Díaz, A. Propiedades físico-químicas y funcionales de los bananos de postre, plátanos de cocción y FHIA híbridos: Preferencia varietal de los consumidores en Colombia. In Proceedings of the CD-Proceedings, Acorbat 2008, XVIII International Meeting, Guayaquil, Ecuador, 11–14 November 2008; Borja, J.S., Nogales, C., Orrantia, C., Paladines, R., Quimi, V., Tazan, L., Eds.; p. 33. Available online: https://agritrop.cirad.fr/547229/ (accessed on 28 February 2025).
- Gibert, O.; Dufour, D.; Giraldo, A.; Sánchez, T.; Reynes, M.; Pain, J.-P.; González, A.; Fernández, A.; Díaz, A. Differentiation between cooking bananas and dessert bananas. 1. Morphological and compositional characterization of cultivated Colombian Musaceae (Musa sp.) in relation to consumer preferences. J. Agric. Food Chem. 2009, 57, 7857–7869. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, M.E.; Jarvis, A.; Stalker, H.T.; Williams, D.E.; Guarino, L.; Valls, J.F.M.; Pittman, R.N.; Simpson, C.E.; Bramel, P.J. Biogeography of wild Arachis (Leguminosae): Distribution and environmental characterization. Biodivers. Conserv. 2005, 14, 1777–1798. [Google Scholar] [CrossRef]
- Wang, G.; Zhou, G.; Yang, L.; Li, Z. Distribution, species diversity and life-form spectra of plant communities along an altitudinal gradient in the northern slopes of Qilianshan Mountains, Gansu, China. Plant Ecol. 2003, 165, 169–181. [Google Scholar] [CrossRef]
- Williams, C.L.; Hargrove, W.W.; Liebman, M.; James, D.E. Agro-ecoregionalization of Iowa using multivariate geographical clustering. Agric. Ecosyst. Environ. 2008, 123, 161–174. [Google Scholar] [CrossRef]
- Parra-Quijano, M.; Iriondo, J.M.; Torres, E. Ecogeographical land characterization maps as tool for assessing plant adaptation and their implications in agrobiodiversity studies. Genet. Resour. Crop. Evol. 2011, 59, 205–217. [Google Scholar] [CrossRef]
- Lobo, M.G.; Rojas, F.J.F. Biology and postharvest physiology of banana. In Handbook of Banana Production, Postharvest Science, Processing Technology, and Nutrition; Siddiq, M., Ahmed, J., Lobo, M.G., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2020; pp. 19–44. [Google Scholar] [CrossRef]
- Lima Neto, A.J.; Deus, J.A.L.; Rodrigues Filho, V.A.; Natale, W.; Parent, L.E. Nutrient diagnosis of fertigated “Prata” and “Cavendish” banana (Musa spp.) at plot-scale. Plants 2020, 9, 1467. [Google Scholar] [CrossRef]
- Solly, E.F.; Weber, V.; Zimmermann, S.; Walthert, L.; Hagedorn, F.; Schmidt, M.W.I. A Critical Evaluation of the Relationship Between the Effective Cation Exchange Capacity and Soil Organic Carbon Content in Swiss Forest Soils. Front. For. Glob. Change 2020, 3, 98. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Descriptor | Unit of Measurement | Descriptor | Unit of Measurement |
|---|---|---|---|
| Pseudostem Height | m | Number of fruits (middle hand) | n |
| Number of suckers | n | Length of pealed fruits | cm |
| Length of a leaf blade (lamina) | m | Raceme weight | kg |
| Width of a leaf blade (lamina) | cm | Number of hands raceme−1 | n |
| Length of the ♀ peduncle | cm | Number of fruits | n |
| Length of the ♂ peduncle | cm | External length of the fruit | cm |
| Diameter of the ♀ peduncle | cm | Fruit diameter | cm |
| Number of green leaves at flowering | n | Weight of the fruit | kg |
| Number of green leaves at harvest | n |
| Descriptor | State | Descriptor | State |
|---|---|---|---|
| Leaf Habit | Erect | Pseudostem aspects | Thin |
| Normal | Normal | ||
| Decumbent | Robust | ||
| Color of the pseudostem | Yellow green | Underlying color of the pseudostem | Yellow green |
| Grayish orange | Grayish orange | ||
| Grayish red | Grayish red | ||
| Grayish violet | Grayish violet | ||
| Red | Purple red | ||
| Grayish brown | |||
| Pigmentation of the inner peel | Yellow green | Wax on the fruits | Very little or no visible signs of wax Little wax Waxy Very waxy |
| Grayish orange | |||
| Purple | |||
| Grayish violet | |||
| Purple red | |||
| Sucker development | Taller than the mother plant More than 3/4 the height of the parent plant Between 1/4 and 3/4 the size of the mother plant Inhibited | Sucker emergence | Distant from mother plant (>50 cm from mother plant) Near the mother plant (vertical growth) Near the mother plant (very inclined) |
| Spots at the base of the petiole | Few | Spot color | Brown |
| Small spots | Grayish orange | ||
| Large spots | Grayish red | ||
| Extensive pigmentation | Grayish violet | ||
| No pigmentation | |||
| 3rd Leaf petiole canal | Open with winged margins | Petiole margin | Winged and wavy Winged and not narrowed by the pseudostem Winged and narrowed by the pseudostem Not winged and narrowed by the pseudostem Not winged and not narrowed by the pseudostem |
| Open with erect margins | |||
| Narrow with erect margins | |||
| Margins twisted inwards | |||
| Overlapping margin | |||
| Petiole margin color | Green yellow Green Grayish red Grayish violet Purple red | Color of the abaxial surface of the bract | Absent |
| Purple | |||
| Grayish orange | |||
| Grayish red | |||
| Grayish violet | |||
| Appearance of the pseudostem | Opaque (waxy) | Color of the top surface of the leaf blade | Greenish yellow |
| Shiny (not waxy) | Green | ||
| Appearance of the upper side of the leaf blade | Opaque | Color of the lower side of the leaf blade | Greenish yellow |
| Shiny | Green | ||
| Appearance of the leaf blade | Opaque | Vein color on the adaxial leaf surface | Greenish yellow |
| Shiny | Green | ||
| Dorsal surface color of the cigar leaf | Green yellow Green | Peel adhesion | Detaches easily It does not come off easily |
| Shape of the leaf blade basis | Both rounded One rounded and one sharp Both sharp | Stain on the sheath of suckers | Spotless Small or narrow spots Large purple spots |
| Pubescence of the peduncle | Glabrous (lacking hair and glands) Little pubescent Very pubescent/short hairs (like touching velvet) Very pubescent/long hairs (>2 mm) | Position of the raceme | Vertically pendular Slightly inclined Oblique at 45° Horizontal Erect |
| Rachis type | Truncated, without a bract scar below the last fruit hand | Seed surface | Smooth Wrinkled |
| Present and the male bud may be degenerated or persistent | |||
| Appearance of the floral rachis | Absent Naked Sterile flower (in one or a few coats, the rachis is bare afterwards) Male flower or persistent bract above the male bud (the rachis is bare above) Sterile or male flower and presence of bracts with brushstrokes (along the entire rachis) Sterile or male flower on the entire rachis, without a persistent bract Small cluster of neutral/hermaphrodite flowers just above the male bud Without sterile or male flower on the entire rachis, without a persistent bract Sterile or male flowers from the middle rachis, with persistent bracts or others | Color of the adaxial side of the bract | Absent Grayish yellow Grayish orange Grayish red Grayish violet Red Purple red Red orange Orange |
| Shape of the male bud | Absent | Shape of the apex of the bracts | Absent |
| Cone shaped | Sharp | ||
| Lanceolate | Slightly pointed | ||
| Intermediate | Intermediate | ||
| Ovoid | Obtuse | ||
| Rounded | Obtuse and cracked | ||
| Type of male bud | Normal (present) Degenerated before maturity (e.g., ‘false horn’ banana) As a ‘true horn’ plantain (absent) | Imbrication of bracts | Absent |
| Old bracts cover the apex | |||
| Young bracts lightly cover it | |||
| Young bracts cover it | |||
| Position of the rachis (spine) | Absent Vertically pendular Inclined With a bend Horizontal Erect | Compound tepal color | Absent Yellow Yellow orange White Yellow white Purple red |
| Attenuated coloration of the bract base | Absent Discontinuous coloration towards the insertion with the rachis (pigmented zone at the level of the bract base) Homogeneous coloring (pigmentation is uniform and continuous to the base) | Color of the compound tepal locule | Absent Yellow Yellow orange |
| Pigmentation of the compound tepal | Absent Very little or no visible sign of pigmentation Splattered with “rust” Presence of pink color | Anther color | Absent Yellow Yellow orange White Grayish yellow Grayish orange |
| Bract behavior before falling | Absent Enrolled (rolls up) Not enrolled (does not curl) | Pigmentation of the ovary | Absent Very little or no visible sign of pigmentation Red/violet |
| Fruit cross-section | Pronounced edges Weakly pronounced edges Rounded | Floral vestige at the apex of the fruit | Without a trace of flowers Persistent style Prominent style base |
| Presence of wax on the bract | Absent Very little or no visible signs of wax Very little wax Waxy Very waxy | Style shape | Absent Straight With a curve below the stigma Curve at the base With two curves |
| Stigma color | Absent Yellow Yellow orange Grayish orange Grayish yellow Yellow white | Color of the ovary basis | Absent Yellow Orange White Greenish yellow Greenish white |
| Appearance of the free tepal | Absent Fold below the apex More or less smooth Very folded under the apex (corrugated) | Fruit shape | Straight (slight curve) Straight in the distal part Curved (a very marked curve) S-curve (double curve) |
| Fruit apex | Pointed Long pointed Truncated | Seed shape | Flattened Angular (more or less pyramidal) Globular (spherical) Rounded (but not completely spherical) |
| In bottleneck Rounded | |||
| Color of the unripe peel | Green yellow Green Grayish orange | Color of the ripe peel | Yellow Yellow orange Grayish orange |
| Color of the pulp at maturity | Yellow Yellow orange Orange White Yellow white Orange white Grayish yellow Grayish orange | Color of the immature pulp | Yellow Yellow orange Orange White Yellow white Orange white Grayish yellow |
| Predominant flavor | Astringent (like cooked bananas) Mild Sweet (like Cavendish) Sweetened (like ‘Pisang Mas’) Sweet and sour (like apples) |
| Group 1 | INIAP-ECU-21792 c, INIAP-ECU-21775 c, INIAP-ECU-21732 c, INIAP-ECU-21795 c, INIAP-ECU-29044 c, INIAP-ECU-29045 c, INIAP-ECU-29046 c, INIAP-ECU-21807 c, INIAP-ECU-21749 c, INIAP-ECU-29047 c, INIAP-ECU-29048 c, INIAP-ECU-29049 c, INIAP-ECU-29050 c, INIAP-ECU-29051 c, INIAP-ECU-29052 c, INIAP-ECU-21748 c, INIAP-ECU-29064 c, INIAP-ECU-29065 c, INIAP-ECU-29066 c, INIAP-ECU-29067 c, INIAP-ECU-29068 c, INIAP-ECU-29069 c, INIAP-ECU-21803 c, INIAP-ECU-29070 c, and INIAP-ECU-29071 c |
| Group 2 | INIAP-ECU-21763 b, INIAP-ECU-21760 b, INIAP-ECU-21753 b, INIAP-ECU- INIAP-ECU-29034 b, INIAP-ECU-29035 b, INIAP-ECU-29036 b, INIAP-ECU-29037 b, INIAP-ECU-29038 b, and INIAP-ECU-29039 b |
| Group 3 | INIAP-ECU-21776 d, INIAP-ECU-21754 d, INIAP-ECU-21744 d, INIAP-ECU-21736 d, INIAP-ECU-29029 d, INIAP-ECU-21804 d, INIAP-ECU-29030 d, INIAP-ECU-29031 d, INIAP-ECU-21782 d, 21730 d, INIAP-ECU-29032 d, INIAP-ECU-21777 d, INIAP-ECU-21783 d, INIAP-ECU-29040 d, INIAP-ECU-29041 d, INIAP-ECU-21798 d, INIAP-ECU-21813 d, INIAP-ECU-29042 d, INIAP-ECU-21745 d, INIAP-ECU-21758 d, INIAP-ECU-21820 d, INIAP-ECU-29043 d, INIAP-ECU-21751 c, INIAP-ECU-21789 c, INIAP-ECU-29062 c, and INIAP-ECU-29063 c |
| Group 4 | INIAP-ECU-21769 a, INIAP-ECU-21766 a, INIAP-ECU-21781 a, INIAP-ECU-21791 a, 21,733 a, 29,026 a, 29,027 a, 21,762 a, 21,740 a, 21,734 a, 29,028 a, 21,800 a, 21,817 a, 29,033 a, 21,774 a, INIAP-ECU-21818 d, INIAP-ECU-21801 c, INIAP-ECU-21752 c, INIAP-ECU-21786 c, INIAP-ECU-21799 c, INIAP-ECU-21811 c, INIAP-ECU-29053 c, INIAP-ECU-21767 c, INIAP-ECU-21765 c, INIAP-ECU-21738 c, INIAP-ECU-21779 c, INIAP-ECU-21787 c, INIAP-ECU-29054 c, INIAP-ECU-29055 c, INIAP-ECU-21793 c, INIAP-ECU-21814 c, INIAP-ECU-29056 c, INIAP-ECU-29057 c, INIAP-ECU-29058 c, INIAP-ECU-29059 c, INIAP-ECU-21741 c, INIAP-ECU-21784 c, INIAP-ECU-21815 c, INIAP-ECU-29060 c, and INIAP-ECU-29061 c |
| Characteristic | Group 1 | Group 2 | Group 3 | Group 4 |
|---|---|---|---|---|
| Pseudo-stem height (m) | 3.29 | 3.25 | 4.05 | 3.62 |
| Number of suckers | 2 | 3 | 2 | 2 |
| Length of the leaf blade (lamina) (m) | 1.92 | 2.54 | 2.70 | 2.26 |
| Width of the leaf blade (lamina) (cm) | 77.03 | 68.51 | 85.89 | 78.17 |
| Length of the petiole (cm) | 38.92 | 39.08 | 48.67 | 40.31 |
| Number of green leaves at flowering | 10 | 10 | 10 | 9 |
| Number of green leaves at harvest | 4 | 6 | 4 | 3 |
| Length of the peduncle (cm) | 46.44 | 50.88 | 54.26 | 47.06 |
| Diameter of the peduncle (cm) | 5.55 | 5.1 | 6.54 | 6.05 |
| Weight of the raceme (kg) | 11.57 | 10.99 | 24.93 | 16.78 |
| Number of hands | 6 | 7 | 8 | 8 |
| Number of fruits (average hand) | 8 | 20 | 15 | 14 |
| Number of fruits raceme−1 | 36 | 143 | 117 | 107 |
| Length of peeled fruits (cm) | 19.4 | 8.34 | 13.31 | 13.42 |
| Length of unpeeled fruits (cm) | 32.13 | 13.08 | 22.07 | 23.4 |
| Diameter of the fruit (cm) | 4.42 | 3.24 | 4.21 | 3.44 |
| Weight of the fruit (kg) | 0.32 | 0.07 | 0.19 | 0.14 |
| Qualitative Characteristics | Group 1 | Group 2 | Group 3 | Group 4 | |
|---|---|---|---|---|---|
| Leaf Habit 1 | Erect | 1% | 1% | ||
| Normal | 25% | 8% | 24% | 38% | |
| Decumbent | 2% | 1% | |||
| Sucker development 1 | Higher than 3/4 of the mother plant’s height | 1% | |||
| Between 1/4 and 3/4 the size of the mother plant | 24% | 9% | 26% | 40% | |
| Color of the pseudostem 2 | Yellow green | 24% | 9% | 17% | 37% |
| Grayish orange | 1% | ||||
| Grayish red | 1% | ||||
| Grayish violet | 1% | ||||
| Red | 1% | ||||
| Grayish brown | 8% | ||||
| Appearance of the pseudostem 2 | Opaque (waxy) | 25% | 9% | 25% | 40% |
| Shiny (not waxy) | 1% | ||||
| Pigmentation of the inner peel 2 | Yellow green | 1% | |||
| Grayish orange | 1% | 1% | |||
| Purple | 1% | 15% | 2% | ||
| Grayish violet | 8% | 9% | 6% | 11% | |
| Purple red | 15% | 4% | 26% | ||
| Presence of wax on the fruit bract 2 | Very little or no visible signs of wax | 7% | 2% | ||
| Little wax | 19% | 2% | 2% | 22% | |
| Waxy | 6% | 18% | 17% | ||
| Very waxy | 4% | 1% | |||
| Spots at the base of the petiole 3 | Few | 8% | 13% | 3% | |
| Small spots | 16% | 9% | 5% | 18% | |
| Large spots½ | 8% | 15% | |||
| Extensive pigmentation | 1% | 4% | |||
| Spot color 3 | Brown | 4% | 12% | 18% | |
| Grayish orange | 9% | 3% | 3% | ||
| Grayish red | 1% | ||||
| Grayish violet | 21% | 11% | 18% | ||
| 3rd Leaf petiole canal 3 | Open with winged margins | 1% | |||
| Open with erect margins | 2% | 15% | 12% | ||
| Narrow with erect margins | 23% | 9% | 10% | 27% | |
| Margins twisted inwards | 1% | ||||
| Vein color on the adaxial leaf surface 3 | Greenish yellow | 23% | 1% | 16% | 37% |
| Green | 2% | 8% | 10% | 3% | |
| Dorsal surface color of the cigar leaf 3 | Greenish yellow | 24% | 9% | 24% | 39% |
| Green | 1% | 2% | 1% | ||
| Type of male bud 4 | Normal (present) | 2% | 9% | 26% | 40% |
| Degenerated before maturity (e.g., ‘false horn’ banana) | 14% | ||||
| As a ‘true horn’ plantain (absent) | 9% | ||||
| Shape of the male bud 4 | Absent | 9% | |||
| Cone shaped | 6% | ||||
| Lanceolate | 16% | ||||
| Intermediate | 9% | 15% | 17% | ||
| Ovoid | 1% | 22% | |||
| Rounded | 4% | 1% | |||
| Shape of the apex of the bracts 5 | Absent | 9% | |||
| Sharp | 7% | ||||
| Slightly pointed | 7% | 5% | 8% | 4% | |
| Intermediate | 2% | 4% | 14% | 30% | |
| Obtuse | 3% | 2% | |||
| Obtuse and cracked | 1% | 4% | |||
| Imbrication of bracts 5 | Absent | 9% | |||
| Old bracts cover the apex | |||||
| Young bracts lightly cover it | 16% | 13% | 27% | ||
| Young bracts cover it | 9% | 13% | 13% | ||
| Color of the adaxial side of the bract 5 | Absent | 9% | |||
| Grayish yellow | 2% | ||||
| Grayish orange | 6% | 1% | |||
| Grayish red | 1% | 17% | 7% | ||
| Grayish violet | 13% | 6% | 24% | ||
| Red | 1% | 2% | 4% | ||
| Purple red | 1% | 1% | |||
| Red orange | 1% | 3% | |||
| Orange | 1% | ||||
| Attenuated coloration of the bract base 5 | Absent | 9% | |||
| Discontinuous coloration towards the insertion with the rachis (pigmented zone at the level of the bract base) | 9% | 15% | |||
| Homogeneous coloring (pigmentation is uniform and continuous to the base) | 16% | 26% | 25% | ||
| Bract behavior before falling 5 | Absent | 9% | |||
| Enrolled (rolls up) | 9% | 24% | 18% | ||
| Not enrolled (does not curl) | 16% | 2% | 22% | ||
| Presence of wax on the bract 5 | Absent | 9% | |||
| Very little or no visible signs of wax | |||||
| Very little wax | 6% | 9% | 11% | ||
| Waxy | 10% | 22% | 28% | ||
| Very waxy | 4% | 1% | |||
| Compound tepal color 6 | Absent | 9% | |||
| Yellow | 1% | 3% | |||
| Yellow orange | 1% | ||||
| White | 9% | 8% | 21% | 19% | |
| Yellow white | 6% | 1% | 3% | 17% | |
| Purple red | 1% | 1% | |||
| Pigmentation of the compound tepal 6 | Absent | 9% | |||
| Very little or no visible sign of pigmentation | 6% | 9% | 5% | 34% | |
| Splattered with “rust” | 10% | 1% | 5% | ||
| Presence of pink color | 20% | 1% | |||
| Color of compound tepal locule 6 | Absent | 9% | |||
| Yellow | 9% | 5% | 16% | 20% | |
| Yellow orange | 7% | 4% | 10% | 20% | |
| Appearance of the free tepal 6 | Absent | 9% | |||
| Fold below the apex | 16% | 9% | 26% | 19% | |
| More or less smooth | 3% | ||||
| Very folded under the apex (corrugated) | 18% | ||||
| Stigma color 6 | Absent | 9% | |||
| Yellow | 1% | 1% | 4% | 6% | |
| Yellow orange | 1% | 5% | 2% | 14% | |
| Grayish orange | 1% | 6% | 3% | ||
| Grayish yellow | 9% | 3% | 4% | 3% | |
| Yellow white | 4% | 10% | 14% | ||
| Color of the ovary basis 6 | Absent | 9% | |||
| Yellow | 1% | 2% | |||
| Orange | 1% | ||||
| White | 2% | 1% | 6% | 6% | |
| Greenish yellow | 6% | 2% | 5% | 30% | |
| Greenish white | 8% | 6% | 14% | 1% | |
| Pigmentation of the ovary 6 | Absent | 9% | |||
| Very little or no visible sign of pigmentation | 16% | 9% | 6% | 39% | |
| Red/violet | 20% | 1% | |||
| Position of the raceme 7 | Vertically pendular | 19% | 4% | 20% | 24% |
| Slightly inclined | 6% | 14% | |||
| Oblique at 45° | 1% | ||||
| Horizontal | 6% | 5% | |||
| Erect | 1% | ||||
| Rachis type 7 | Truncated, without a bract scar below the last fruit hand | 9% | |||
| Present and the male bud may be degenerated or persistent | 16% | 9% | 26% | 40% | |
| Position of the rachis (spine) 7 | Absent | 9% | |||
| Vertically pendular | 8% | 1% | 26% | 36% | |
| Inclined | 8% | ||||
| With a bend | 8% | 4% | |||
| Appearance of the floral rachis 7 | Male flower or persistent bract above the male bud (the rachis is bare above) | 10% | 6% | ||
| Male flower or persistent bract above the male bud (the rachis is bare above) | 9% | 14% | 12% | ||
| Sterile or male flowers throughout the rachis, without persistent bracts | 16% | ||||
| Small cluster of sterile flowers just above the male bud | 2% | 21% | |||
| No sterile or male flowers on the entire rachis, without persistent bracts | 9% | ||||
| Sterile or male flowers from the middle rachis, with persistent bracts | 1% | ||||
| Fruit shape 8 | Straight (slight curve) | 1% | 9% | 14% | 5% |
| Straight in the distal part | 24% | 4% | 22% | ||
| Curved (a very marked curve) | 8% | 13% | |||
| Fruit cross-section 8 | Pronounced edges | 25% | 5% | 22% | |
| Weakly pronounced edges | 21% | 18% | |||
| Rounded | 9% | ||||
| Fruit apex 8 | Pointed | 3% | 1% | ||
| Long pointed | 25% | 4% | 21% | ||
| Truncated | 18% | 18% | |||
| In bottleneck | 1% | ||||
| Rounded | 9% | ||||
| Floral vestige at the apex of the fruit 8 | Without a trace of flowers | 22% | 16% | ||
| Persistent style | 9% | 2% | |||
| Prominent style base | 25% | 4% | 22% | ||
| Color of the unripe peel 8 | Greenish yellow | 24% | 9% | 16% | 38% |
| Green | 1% | 1% | 2% | ||
| Grayish orange | 9% | ||||
| Color of the ripe peel 8 | Yellow | 24% | 9% | 15% | 38% |
| Yellow orange | 1% | 2% | 2% | ||
| Grayish orange | 9% | ||||
| Peel adhesion 8 | Detaches easily | 9% | 23% | 17% | |
| It does not come off easily | 25% | 3% | 23% | ||
| Color of the immature pulp 8 | Yellow | 1% | 6% | ||
| Yellow orange | 3% | 1% | 4% | ||
| Orange | 6% | 2% | 2% | ||
| White | 4% | 3% | 8% | ||
| Yellow white | 4% | 5% | 1% | ||
| Orange-white | 11% | 15% | 17% | ||
| Grayish yellow | 5% | 2% | |||
| Color of the pulp at maturity 8 | Yellow | 3% | 2% | 11% | |
| Yellow orange | 17% | 4% | 10% | 20 | |
| Orange | 7% | 1% | |||
| White | 1% | 1% | |||
| Yellow white | 1% | 3% | 4% | ||
| Orange-white | 2% | ||||
| Grayish yellow | 1% | 9% | 1% | ||
| Grayish orange | 1% | 1% | |||
| Predominant flavor 8 | Astringent (like cooked bananas) | ||||
| Mild | 22% | 3% | |||
| Sweet (like Cavendish) | 14% | ||||
| Sweetened (like ‘Pisang Mas’) | 9% | ||||
| Sweet and sour (like apples) | 25% | 4% | 23% |
| Variable | X2 | Coefficient (P) | Cramer (V) |
|---|---|---|---|
| Fruit cross-section | 137.46 ** | 3.4496 × 10−27 | 0.83 |
| Fruit apex | 143.52 ** | 1.1639 × 10−24 | 0.69 |
| Type of male bud | 149.34 ** | 3.2612 × 10−24 | 0.71 |
| Floral vestige at the apex of the fruit | 122.51 ** | 4.841 × 10−24 | 0.78 |
| Appearance of the rachis | 138.31 ** | 4.9674 × 10−22 | 0.68 |
| Pigmentation of the compound tepal | 113.16 ** | 3.3409 × 10−20 | 0.61 |
| Pigmentation of the ovary | 93.16 ** | 6.6632 × 10−18 | 0.68 |
| Type of male bud | 89.61 ** | 3.6505 × 10−17 | 0.67 |
| Bract behavior before falling | 69.20 ** | 5.9569 × 10−13 | 0.59 |
| Fruit shape | 62.67 ** | 1.2863 × 10−11 | 0.56 |
| Shape of the apex of the bracts | 77.31 ** | 2.1613 × 10−10 | 0.51 |
| Imbrication of bracts | 54.79 ** | 5.0999 × 10−10 | 0.52 |
| Color of the ovary basis | 67.80 ** | 1.0983 × 10−8 | 0.48 |
| Stigma color | 62.42 ** | 9.6134 × 10−8 | 0.46 |
| Spots at the base of the petiole | 41.61 ** | 3.8757 × 10−6 | 0.37 |
| Color of the immature pulp | 57.73 ** | 4.7351 × 10−6 | 0.44 |
| Color of the ripe peel | 29.79 ** | 4.3139 × 10−5 | 0.39 |
| Color of the unripe peel | 28.72 ** | 6.884 × 10−5 | 0.38 |
| Color of pseudostem | 29.42 ** | 0.01420494 | 0.31 |
| Seed shape | 8.22 ** | 0.04170303 | 0.29 |
| Category | Annual Temperature | Apparent Density | Cation Exchange Capacity (CEC) |
|---|---|---|---|
| 4 | 23.99 °C ± 1.62 | 0.99 g cm−3 ± 0.08 | 45.79 cmol kg−1 ± 15.53 |
| 9 | 22.36 °C ± 4.96 | 1.32 g cm−3 ± 0.01 | 17.43 cmol kg−1 ± 1.97 |
| 10 | 22 °C ± 4. 40 | 1.13 g cm−3 ± 0.03 | 27.17 cmol kg−1 ± 4.27 |
| 22 | 25.01 °C ± 6.01 | 1.12 g cm−3 ± 0.07 | 29.63 cmol kg−1 ± 7.19 |
| 23 | 25.32 °C ± 1.42 | 0.86 g cm−3 ± 0.07 | 29.39 cmol kg−1 ± 8.53 |
| 12 | 24.75 °C ± 7.70 | 1.39 g cm−3 ± 0.04 | 18.46 cmol kg−1 ± 4.24 |
| 11 | 24.34 °C ± 12.56 | 1.28 g cm−3 ± 0.04 | 21.07 cmol kg−1 ± 5.23 |
| 6 | 23.46 °C ± 13.02 | 1.42 g/cm3 ± 0.04 | 16.78 cmol kg−1 ± 3.96 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Poaquiza, N.A.; Vásconez, R.A.; Tandazo, L.L.; Monteros-Altamirano, Á.; Bastidas, C.T.; Franklin, S.M.; Sørensen, M.; Andrade, N.P. The Morphological and Ecogeographic Characterization of the Musa L. Collection in the Gene Bank of INIAP, Ecuador. Crops 2025, 5, 34. https://doi.org/10.3390/crops5030034
Poaquiza NA, Vásconez RA, Tandazo LL, Monteros-Altamirano Á, Bastidas CT, Franklin SM, Sørensen M, Andrade NP. The Morphological and Ecogeographic Characterization of the Musa L. Collection in the Gene Bank of INIAP, Ecuador. Crops. 2025; 5(3):34. https://doi.org/10.3390/crops5030034
Chicago/Turabian StylePoaquiza, Nelly Avalos, Ramiro Acurio Vásconez, Luis Lima Tandazo, Álvaro Monteros-Altamirano, César Tapia Bastidas, Sigcha Morales Franklin, Marten Sørensen, and Nelly Paredes Andrade. 2025. "The Morphological and Ecogeographic Characterization of the Musa L. Collection in the Gene Bank of INIAP, Ecuador" Crops 5, no. 3: 34. https://doi.org/10.3390/crops5030034
APA StylePoaquiza, N. A., Vásconez, R. A., Tandazo, L. L., Monteros-Altamirano, Á., Bastidas, C. T., Franklin, S. M., Sørensen, M., & Andrade, N. P. (2025). The Morphological and Ecogeographic Characterization of the Musa L. Collection in the Gene Bank of INIAP, Ecuador. Crops, 5(3), 34. https://doi.org/10.3390/crops5030034