
Boron–Vicinal Diol Xanthophyll Complexes as Emerging Photoprotective Adjuvants



Abstract
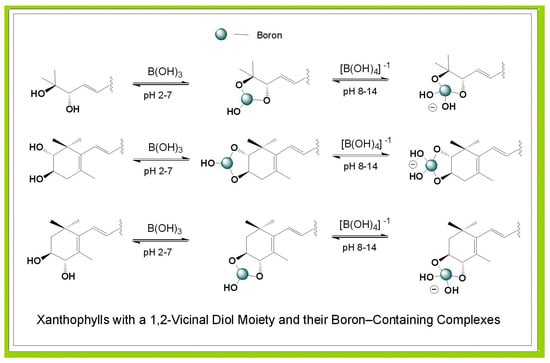
1. Introduction
2. Boron Sequestration by Bacteria
3. Detection of Boron as Complex with Carotenoids
4. Stereochemistry of 1,2-Diol Xanthophylls
5. Functions of 1,2-Diol Xanthophylls
6. Xanthophylls with cis-1,2-Diol Configuration
7. Boron Complexes of Xanthophylls as Structural Elements of Biological Membranes in Bacteria
8. Challenges, Limitations, and Future Perspectives
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brar, G.; Dhaliwal, A.; Brar, A.S.; Sreedevi, M.; Ahmadi, Y.; Irfan, M.; Abarca-Pineda, Y.A. A comprehensive review of the role of UV radiation in photoaging processes between different types of skin. Cureus 2025, 17, e81109. [Google Scholar] [CrossRef]
- Yu, S.L.; Lee, S.K. Ultraviolet radiation: DNA damage, repair, and human disorders. Mol. Cell. Toxicol. 2017, 13, 21–28. [Google Scholar] [CrossRef]
- Ichihashi, M.; Ueda, M.; Budiyanto, A.; Bito, T.; Oka, M.; Fukunaga, M.; Horikawa, T. UV-induced skin damage. Toxicology 2003, 189, 21–39. [Google Scholar] [CrossRef] [PubMed]
- Matsui, Y.; Shimizu, T. The role of macrophage migration inhibitory factor and its homolog D-dopachrome tautomerase in ultraviolet radiation-induced carcinogenesis: New insights into skin cancer mechanisms. Photodermatol. Photoimmunol. Photomed. 2025, 41, e70046. [Google Scholar] [CrossRef]
- Yu, Z.W.; Zheng, M.; Fan, H.Y.; Liang, X.H.; Tang, Y.L. Ultraviolet (UV) radiation: A double-edged sword in cancer development and therapy. Mol. Biomed. 2024, 5, 49. [Google Scholar] [CrossRef]
- Karimi, M.; Sadeghi, E.; Bigdeli, S.K.; Zahedifar, M. Optical properties, singlet oxygen, and free radical production ability with different UV irradiations and antimicrobial inhibitors against various bacterial species of ZnO:Eu nanoparticles. Radiat. Phys. Chem. 2023, 212, 111132. [Google Scholar] [CrossRef]
- Wang, P.; Wang, L.; Xiao, R.; Qiu, S.; Cao, J.; Fu, Y.; Wang, Z. New evidence for the involvement of superoxide and singlet oxygen in UV-activated peroxydisulfate system under acidic conditions. Chem. Eng. J. 2025, 505, 159531. [Google Scholar] [CrossRef]
- Miranda, J.A.; Cruz, Y.F.D.; Girão, Í.C.; Souza, F.J.J.D.; Oliveira, W.N.D.; Alencar, É.D.N.; Egito, E.S.T.D. Beyond traditional sunscreens: A review of liposomal-based systems for photoprotection. Pharmaceutics 2024, 16, 661. [Google Scholar] [CrossRef]
- Dharmalingam, V.; Patrick, A.P.R.; Venkatesan, V.; Subramani, N. Bioprospecting of lithophytic microalgae for photoprotective compounds against UV-B radiation: A review. Bioresour. Technol. Rep. 2026, 12, 102527. [Google Scholar] [CrossRef]
- Verma, A.; Zanoletti, A.; Kareem, K.Y.; Adelodun, B.; Kumar, P.; Ajibade, F.O.; Dwivedi, A. Skin protection from solar ultraviolet radiation using natural compounds: A review. Environ. Chem. Lett. 2024, 22, 273–295. [Google Scholar] [CrossRef]
- Řezanka, T.; Temina, M.; Tolstikov, A.G.; Dembitsky, V.M. Natural microbial UV radiation filters—Mycosporine-like amino acids. Folia Microbiol. 2004, 49, 339–352. [Google Scholar] [CrossRef]
- Thomas, S.E.; Johnson, E.J. Xanthophylls. Adv. Nutr. 2018, 9, 160–162. [Google Scholar] [CrossRef]
- Jackson, H.; Braun, C.L.; Ernst, H. The chemistry of novel xanthophyll carotenoids. Am. J. Cardiol. 2008, 101, S50–S57. [Google Scholar] [CrossRef]
- Zaripheh, S.; Erdman, J.W., Jr. Factors that influence the bioavailability of xanthophylls. J. Nutr. 2002, 132, 531S–534S. [Google Scholar] [CrossRef]
- Goss, R.; Jakob, T. Regulation and function of xanthophyll cycle-dependent photoprotection in algae. Photosynth. Res. 2010, 106, 103–122. [Google Scholar] [CrossRef] [PubMed]
- Giossi, C.; Cartaxana, P.; Cruz, S. Photoprotective role of neoxanthin in plants and algae. Molecules 2020, 25, 4617. [Google Scholar] [CrossRef]
- Rastogi, R.P.; Richa; Sinha, R.P.; Singh, S.P.; Häder, D.P. Photoprotective compounds from marine organisms. J. Ind. Microbiol. Biotechnol. 2010, 37, 537–558. [Google Scholar] [CrossRef] [PubMed]
- Bhosale, P.; Bernstein, P.S. Microbial xanthophylls. Appl. Microbiol. Biotechnol. 2005, 68, 445–455. [Google Scholar] [CrossRef]
- Tyman, J.H.P. The chemistry of some natural colourants. Stud. Nat. Prod. Chem. 1997, 20, 719–788. [Google Scholar]
- Dembitsky, V.M.; Terent’ev, A.O.; Baranin, S.V.; Gursky, M.E. Aromatic compounds and their fascinating boron complexes as potential quorum sensing molecules. Vietnam J. Chem. 2025, 63, 883–911. [Google Scholar] [CrossRef]
- Dembitsky, V.M.; Baranin, S.V.; Terent’ev, A.O. 1,2-Diol xanthophylls and their boron complexes. MarineMedicine 2025, 1, 11. [Google Scholar] [CrossRef]
- Iga, D.P. Carotenoid structures, an illustration of a new kind of symmetry in chemistry. Chem. Res. J. 2021, 6, 20–48. [Google Scholar]
- Jerković, I.; Kuś, P.M. Terpenes in honey: Occurrence, origin and their role as chemical biomarkers. RSC Adv. 2014, 4, 31710–31728. [Google Scholar] [CrossRef]
- Khurana, A.L. Food analysis on silica-bound HPLC phases. Crit. Rev. Food Sci. Nutr. 1990, 29, 197–235. [Google Scholar] [CrossRef] [PubMed]
- Smaoui, S.; Barkallah, M.; Ben Hlima, H.; Fendri, I.; Mousavi Khaneghah, A.; Michaud, P.; Abdelkafi, S. Microalgae xanthophylls: From biosynthesis pathway and production techniques to encapsulation development. Foods 2021, 10, 2835. [Google Scholar] [CrossRef]
- Saini, D.K.; Pabbi, S.; Shukla, P. Cyanobacterial pigments: Perspectives and biotechnological approaches. Food Chem. Toxicol. 2018, 120, 616–624. [Google Scholar] [CrossRef]
- Bailey, S.; Grossman, A. Photoprotection in cyanobacteria: Regulation of light harvesting. Photochem. Photobiol. 2008, 84, 1410–1420. [Google Scholar] [CrossRef]
- MacIntyre, H.L.; Kana, T.M.; Anning, T.; Geider, R.J. Photoacclimation of photosynthesis irradiance response curves and photosynthetic pigments in microalgae and cyanobacteria. J. Phycol. 2002, 38, 17–38. [Google Scholar] [CrossRef]
- Kim, K.C.; Kim, N.I.; Jiang, T.; Kim, J.C.; Kang, C.I. Boron recovery from salt lake brine, seawater, and wastewater—A review. Hydrometallurgy 2023, 218, 106062. [Google Scholar]
- Bulut, G.; Aydın, Ş.B.; Perek, K.T.; Arslan, F. Enrichment of boron using physical and chemical methods: A review. Miner. Process. Extr. Metall. Rev. 2025, 47, 426–443. [Google Scholar] [CrossRef]
- Dembitsky, V.M.; Terent’ev, A.O.; Baranin, S.V. Life with Boron: Microbial Boron-Binding Siderophores, Adaptation, and Function. Microbiol. Res. 2026, 17, 57. [Google Scholar] [CrossRef]
- Zhu, S.; Khan, M.S.; Xiao, K.; Chang, C.M.; Zhao, Q.; Chung, T.S.; Chen, S.B. Brick-and-cement structured polyamide membranes enabling to selectively separate boron from brackish water and real seawater permeate. J. Membr. Sci. 2026, 26, 125198. [Google Scholar] [CrossRef]
- Mutlu-Salmanli, O.; Koyuncu, I. Boron removal and recovery from water and wastewater. Rev. Environ. Sci. Biotechnol. 2022, 21, 635–664. [Google Scholar] [CrossRef]
- Sharkh, B.A.; Al-Amoudi, A.A.; Farooque, M.; Fellows, C.M.; Ihm, S.; Lee, S.; Voutchkov, N. Seawater desalination concentrate—A new frontier for sustainable mining of valuable minerals. npj Clean Water 2022, 5, 9. [Google Scholar] [CrossRef]
- Xuemin, L.; Xu, W.; Yuanliang, Y.; Dabin, Y.; Xingjian, L.; Pufeng, Z.; Bingli, L. Pegmatite evolution and mineralization: Insights from tourmaline geochemistry and boron isotopes. Ore Geol. Rev. 2026, 190, 107156. [Google Scholar] [CrossRef]
- Kang, N.; Zhou, W.; Qi, Z.; Li, Y.; Wang, Z.; Li, Q.; Lv, K. Recent progress of natural mineral materials in environmental remediation. Catalysts 2022, 12, 996. [Google Scholar] [CrossRef]
- Halvacı, E.; İkballı, D.; Özengül, A.; Sevimli, B.; Tumeh, A.A.; Güzel, Z.; Şen, F. The role of boron in new generation technologies and sustainable future. Int. J. Boron Sci. Nanotechnol. 2024, 1, 86–102. [Google Scholar]
- Micus, J.L.; Altman, I.; Pantoya, M.L. Fundamental calorimetric measurements of boron oxidation from dry powder suspensions. Chem. Eng. J. 2026, 18, 172628. [Google Scholar] [CrossRef]
- Guo, S.; Mao, B.; Tang, X.; Zhang, Q.; Zhao, J.; Chen, W.; Cui, S. Autoinducer-2 as a universal language in microbial consortia: Decoding molecular mechanisms, ecological impacts, and application. Gut Microbes 2026, 18, 2615494. [Google Scholar] [CrossRef]
- Konaklieva, M.I.; Plotkin, B.J. Activity of organoboron compounds against biofilm-forming pathogens. Antibiotics 2024, 13, 929. [Google Scholar] [CrossRef]
- Dembitsky, V.M.; Terent’ev, A.O.; Stolbov, L.A.; Pogodin, P.V.; Filimonov, D.A.; Poroikov, V.V. Salicylic acid and its boron complexes as quorum sensing molecules. Mol. Pharm. 2025, 22, 6499–6509. [Google Scholar] [CrossRef]
- Dembitsky, V.M.; Terent’ev, A.O.; Baranin, S.V. Boronosteroids as potential antitumor drugs: A review. Tumor Discov. 2025, 5, 14–33. [Google Scholar] [CrossRef]
- Dembitsky, V.M.; Terent’ev, A.O.; Gursky, M.E.; Baranin, S.V. Fascinating and intriguing biomolecules: The chemistry of boron complexes with carbohydrates, glycolipids, and steroids. Vietnam J. Chem. 2025, in press. [Google Scholar] [CrossRef]
- Dembitsky, V.M.; Al Quntar, A.A.A.; Srebnik, M. Natural and synthetic small boron-containing molecules as potential inhibitors of bacterial and fungal quorum sensing. Chem. Rev. 2011, 111, 209–237. [Google Scholar] [CrossRef]
- Dembitsky, V.M.; Smoum, R.; Al-Quntar, A.A.; Ali, H.A.; Pergament, I.; Srebnik, M. Natural occurrence of boron-containing compounds in plants, algae and microorganisms. Plant Sci. 2002, 163, 931–942. [Google Scholar] [CrossRef]
- Ahmed, I.; Zulfiqar, S.; Ali, M.; Abbas, S.; Fujiwara, T. Boron-tolerant bacteria in bioremediation: Mechanisms, applications, and prospects for environmental sustainability. Sustain. Microbiol. 2026, 3, qvaf034. [Google Scholar] [CrossRef]
- Shireen, F.; Nawaz, M.A.; Chen, C.; Zhang, Q.; Zheng, Z.; Sohail, H.; Bie, Z. Boron: Functions and approaches to enhance its availability in plants for sustainable agriculture. Int. J. Mol. Sci. 2018, 19, 1856. [Google Scholar] [CrossRef]
- Biţă, A.; Scorei, I.R.; Bălşeanu, T.A.; Ciocîlteu, M.V.; Bejenaru, C.; Radu, A.; Benner, S.A. New insights into boron essentiality in humans and animals. Int. J. Mol. Sci. 2022, 23, 9147. [Google Scholar] [CrossRef]
- Coulthurst, S.J.; Whitehead, N.A.; Welch, M.; Salmond, G.P.C. Can boron get bacteria talking? Trends Biochem. Sci. 2002, 27, 217–219. [Google Scholar] [CrossRef]
- Huang, Z.; Bai, L.; Liu, J.; Luo, Y. Boron-containing compounds as antimicrobial agents to tackle drug-resistant bacteria. Pharm. Front. 2024, 6, e336–e354. [Google Scholar] [CrossRef]
- Miwa, H.; Fujiwara, T. Isolation and identification of boron-accumulating bacteria from contaminated soils and active sludge. Soil. Sci. Plant Nutr. 2009, 55, 643–646. [Google Scholar] [CrossRef]
- Lima, S.; Matinha-Cardoso, J.; Giner-Lamia, J.; Couto, N.; Pacheco, C.C.; Florencio, F.J.; Oliveira, P. Extracellular vesicles as an alternative copper-secretion mechanism in bacteria. J. Hazard. Mater. 2022, 431, 128594. [Google Scholar] [CrossRef]
- Akai, M.; Onai, K.; Kusano, M.; Sato, M.; Redestig, H.; Toyooka, K.; Uozumi, N. Plasma membrane aquaporin AqpZ protein is essential for glucose metabolism during photomixotrophic growth of Synechocystis sp. PCC 6803. J. Biol. Chem. 2011, 286, 25224–25235. [Google Scholar] [CrossRef]
- Mateo, P.; Bonilla, I.; Fernandez-Valiente, E.; Sanchez-Maeso, E. Essentiality of boron for dinitrogen fixation in Anabaena sp. PCC 7119. Plant Physiol. 1986, 81, 430–433. [Google Scholar] [CrossRef]
- Abreu, I.; Orús, I.; Bolaños, L.; Bonilla, I. The interaction of boron with glycolipids is required to increase tolerance to stresses in Anabaena PCC 7120. Phytochemistry 2014, 106, 55–60. [Google Scholar] [CrossRef]
- Bonilla, I.; Bolaños, L.; Mateo, P. Interaction of boron and calcium in the cyanobacteria Anabaena and Synechococcus. Physiol. Plant. 1995, 94, 31–36. [Google Scholar] [CrossRef]
- Niemczyk, E.; Pogrzeba, J.; Adamczyk-Woźniak, A.; Lipok, J. Boronic acids of pharmaceutical importance affect the growth and photosynthetic apparatus of cyanobacteria in a dose-dependent manner. Toxins 2020, 12, 793. [Google Scholar] [CrossRef]
- Morsi, H.H.; Gharieb, M.M.; Abd El-Monem, A.M.; Doman, K.M. The influence of nutrient manipulation on growth and cultivation constituents of Anabaena variabilis. Egypt. J. Phycol. 2023, 24, 54–77. [Google Scholar] [CrossRef]
- Bonilla, I.; Garcia-Gonzalez, M.; Mateo, P. Boron requirement in cyanobacteria: Its possible role in the early evolution of photosynthetic organisms. Plant Physiol. 1990, 94, 1554–1560. [Google Scholar] [CrossRef]
- Dembitsky, V.M.; Rezanka, T. Metabolites produced by nitrogen-fixing Nostoc species. Folia Microbiol. 2005, 50, 363–391. [Google Scholar] [CrossRef]
- Řezanka, T.; Dor, I.; Prell, A.; Dembitsky, V.M. Fatty acid composition of six freshwater wild cyanobacterial species. Folia Microbiol. 2003, 48, 71–75. [Google Scholar] [CrossRef]
- Temina, M.; Rezankova, H.; Rezanka, T.; Dembitsky, V.M. Diversity of the fatty acids of the Nostoc species and their statistical analysis. Microbiol. Res. 2007, 162, 308–321. [Google Scholar] [CrossRef]
- Dembitsky, V.M.; Shkrob, I.; Dor, I. Separation and identification of hydrocarbons and other volatile compounds from cultured blue-green alga Nostoc sp. by gas chromatography–mass spectrometry using serially coupled capillary columns with consecutive nonpolar and semipolar stationary phases. J. Chromatogr. A 1999, 862, 221–229. [Google Scholar] [CrossRef]
- Dembitsky, V.M. Hydrobiological aspects of fatty acids: Unique, rare, and unusual fatty acids incorporated into linear and cyclic lipopeptides and their biological activity. Hydrobiology 2022, 1, 331–432. [Google Scholar] [CrossRef]
- Eyster, C. Necessity of boron for Nostoc muscorum. Nature 1952, 170, 755. [Google Scholar] [CrossRef]
- Taştan, B.E.; Bakir, B.; Dönmez, G. Boron bio-mining by high boron-tolerant native microalgae in Turkey and boron toxicity in the aquatic environment. Water Sci. Technol. 2023, 87, 2490–2503. [Google Scholar] [CrossRef]
- Deyhle, A.; Hodge, V.; Lewin, R.A. 33Boron in diatoms. J. Phycol. 2003, 39, 12–13. [Google Scholar] [CrossRef]
- Bala, S.; Garg, D.; Phutela, U.G.; Kaur, M.; Bhatia, S. Oscillatoria sancta cultivation using fruit and vegetable waste formulated media and its potential as a functional food: Assessment of cultivation optimization. Mol. Biotechnol. 2023, 1–19. [Google Scholar] [CrossRef]
- Singh, A.D.; Singh, G.P. Molybdenum stress and its effect on growth and biopigment profile of blue green alga Oscillatoria agardhii. Int. J. Sci. Technol. Res. 2019, 8, 789–792. [Google Scholar]
- Aoki, J.; Koshikawa, R.; Asayama, M. Recent progress in the cyanobacterial products and applications of phycocyanins. World J. Microbiol. Biotechnol. 2025, 41, 84. [Google Scholar] [CrossRef]
- Eberly, J.O.; Ely, R.L. Photosynthetic accumulation of carbon storage compounds under CO2 enrichment by the thermophilic cyanobacterium Thermosynechococcus elongatus. J. Ind. Microbiol. Biotechnol. 2012, 39, 843–850. [Google Scholar] [CrossRef]
- Eberly, J.O. Analysis of the Thermophilic cyanobacterium Thermosynechococcus elongatus as a Model Organism for Carbon Sequestration, Biofuel, and Biomaterial Production. Ph.D. Thesis, Oregon State University, Corvallis, OR, USA, 2010. [Google Scholar]
- Ohkubo, S.; Miyashita, H. A niche for cyanobacteria producing chlorophyll f within a microbial mat. ISME J. 2017, 11, 2368–2378. [Google Scholar] [CrossRef]
- Wang, B.; Zhang, Y.; Minteer, S.D. Renewable electron-driven bioinorganic nitrogen fixation: A superior route toward green ammonia? Energy Environ. Sci. 2023, 16, 404–420. [Google Scholar] [CrossRef]
- Mondal, N.; Dutta, S.; Chatterjee, S.; Sarkar, J.; Mondal, M.; Roy, C.; Ghosh, W. Aquificae overcomes competition by archaeal thermophiles, and crowding by bacterial mesophiles, to dominate the boiling vent-water of a Trans-Himalayan sulfur-borax spring. PLoS ONE 2024, 19, e0310595. [Google Scholar] [CrossRef]
- Hajiyeva, S.; Cankilic, M.Y.; Sariozlu, N.Y.; Student, P. Production, large-scale extraction, and purification of phycocyanin by different cyanobacteria isolated from various environments. Biointerface Res. Appl. Chem. 2024, 14, 91. [Google Scholar] [CrossRef]
- Falkiewicz-Dulik, M.; Janda, K.; Wypych, G. Handbook of Material Biodegradation, Biodeterioration, and Biostabilization; Elsevier: Amsterdam, The Netherlands, 2015. [Google Scholar]
- Hayes, P.R. Studies on marine flavobacteria. Microbiology 1963, 30, 1–19. [Google Scholar] [CrossRef]
- Weerasinghe, A.J.; Amin, S.A.; Barker, R.A.; Othman, T.; Romano, A.N.; Parker Siburt, C.J.; Crumbliss, A.L. Borate as a synergistic anion for Marinobacter algicola ferric binding protein, FbpA: A role for boron in iron transport in marine life. J. Am. Chem. Soc. 2013, 135, 14504–14507. [Google Scholar] [CrossRef]
- Ahmed, I.; Yokota, A.; Fujiwara, T. Chimaereicella boritolerans sp. nov., a boron-tolerant and alkaliphilic bacterium of the family Flavobacteriaceae isolated from soil. Int. J. Syst. Evol. Microbiol. 2007, 57, 986–992. [Google Scholar] [CrossRef][Green Version]
- De Carvalho, C.C.; Fernandes, P. Production of metabolites as bacterial responses to the marine environment. Mar. Drugs 2010, 8, 705–727. [Google Scholar] [CrossRef]
- Strigul, N.; Vaccari, L.; Galdun, C.; Wazne, M.; Liu, X.; Christodoulatos, C.; Jasinkiewicz, K. Acute toxicity of boron, titanium dioxide, and aluminum nanoparticles to Daphnia magna and Vibrio fischeri. Desalination 2009, 248, 771–782. [Google Scholar] [CrossRef]
- Sayin, Z.; Ucan, U.S.; Sakmanoglu, A. Antibacterial and antibiofilm effects of boron on different bacteria. Biol. Trace Elem. Res. 2016, 173, 241–246. [Google Scholar] [CrossRef]
- Yaylacı, E.U. Antibacterial effects of boric acid against aquatic pathogens. J. Anatol. Environ. Anim. Sci. 2021, 6, 240–244. [Google Scholar] [CrossRef]
- Nakagawa, Y.; Hanaoka, H.; Kobayashi, M.; Miyoshi, K.; Miwa, K.; Fujiwara, T. Cell-type specificity of the expression of OsBOR1, a rice efflux boron transporter gene, is regulated in response to boron availability for efficient boron uptake and xylem loading. Plant Cell 2007, 19, 2624–2635. [Google Scholar] [CrossRef]
- Sharma, H.; Sharma, A.; Rajput, R.; Sidhu, S.; Dhillon, H.; Verma, P.C.; Upadhyay, S.K. Molecular characterization, evolutionary analysis, and expression profiling of BOR genes in important cereals. Plants 2022, 11, 911. [Google Scholar] [CrossRef]
- Zhang, Q.; Chen, H.; He, M.; Zhao, Z.; Cai, H.; Ding, G.; Xu, F. The boron transporter BnaC4.BOR1;1c is critical for inflorescence development and fertility under boron limitation in Brassica napus. Plant Cell Environ. 2017, 40, 1819–1833. [Google Scholar] [CrossRef]
- Takano, J.; Miwa, K.; Fujiwara, T. Boron transport mechanisms: Collaboration of channels and transporters. Trends Plant Sci. 2008, 13, 451–457. [Google Scholar] [CrossRef]
- Herrera-Rodríguez, M.B.; González-Fontes, A.; Rexach, J.; Camacho-Cristóbal, J.J.; Maldonado, J.M.; Navarro-Gochicoa, M.T. Role of boron in vascular plants and response mechanisms to boron stresses. Plant Stress 2010, 4, 115–122. [Google Scholar]
- Pandey, N. Update on boron—Physiological responses and homeostasis in plants. In Physiology of Nutrition and Environmental Stresses on Crop Productivity; Scientific Publishers: Jodhpur, India, 2014; p. 28. [Google Scholar]
- Wang, G. Molecular Mechanisms of Boron Toxicity Tolerance in Plants; Louisiana State University and Agricultural & Mechanical College: Baton Rouge, LA, USA, 2020. [Google Scholar]
- Lopalco, A.; Lopedota, A.A.; Laquintana, V.; Denora, N.; Stella, V.J. Boric acid, a Lewis acid with unique and unusual properties: Formulation implications. J. Pharm. Sci. 2020, 109, 2375–2386. [Google Scholar] [CrossRef]
- Weser, U. Chemistry and structure of some borate polyol compounds of biochemical interest. In Structure and Bonding; Jørgensen, C.K., Neilands, J.B., Nyholm, R.S., Reinen, D., Williams, R.J.P., Eds.; Springer: Berlin/Heidelberg, Germany, 1967; Volume 2. [Google Scholar]
- Sah, R.N.; Brown, P.H. Boron determination—A review of analytical methods. Microchem. J. 1997, 56, 285–304. [Google Scholar] [CrossRef]
- Türker, A.; Türker, A.R. A critical review on the determination of boron in various matrices. J. Boron 2019, 4, 31–38. [Google Scholar]
- Farhat, A.; Ahmad, F.; Arafat, H. Analytical techniques for boron quantification supporting desalination processes: A review. Desalination 2013, 310, 9–17. [Google Scholar] [CrossRef]
- Carrano, C.J.; Schellenberg, S.; Amin, S.A.; Green, D.H.; Küpper, F.C. Boron and marine life: A new look at an enigmatic bioelement. Mar. Biotechnol. 2009, 11, 431–440. [Google Scholar] [CrossRef]
- Hatcher, J.T.; Wilcox, L.V. Colorimetric determination of boron using carmine. Anal. Chem. 1950, 22, 567–569. [Google Scholar] [CrossRef]
- Smith, W.C., Jr.; Goudie, A.J.; Sivertson, J.N. Colorimetric determination of trace quantities of boric acid in biological materials. Anal. Chem. 1955, 27, 295–297. [Google Scholar] [CrossRef]
- Hill, W.H.; Merrill, J.M.; Palm, B.J. Direct determination of boranes by the carmine method. Am. Ind. Hyg. Assoc. J. 1958, 19, 461–463. [Google Scholar] [CrossRef] [PubMed]
- Pappin, B.; Kiefel, M.J.; Houston, T.A. Boron–carbohydrate interactions. In Carbohydrates—Comprehensive Studies on Glycobiology and Glycotechnology; IntechOpen: London, UK, 2012; pp. 37–54. [Google Scholar]
- Ferrier, R.J. Carbohydrate boronates. In Advances in Carbohydrate Chemistry and Biochemistry; Academic Press: New York, NY, USA, 1978; Volume 35, pp. 31–80. [Google Scholar]
- Lee, D. Boron–Diol Interactions as the Basis for Novel Catalytic Transformations. Doctoral Dissertation, University of Toronto, Toronto, ON, USA, 2014. [Google Scholar]
- Henderson, W.G.; How, M.J.; Kennedy, G.R.; Mooney, E.F. The interconversion of aqueous boron species and the interaction of borate with diols: A 11B NMR study. Carbohydr. Res. 1973, 28, 1–12. [Google Scholar]
- Gertsev, V.V.; Komissarov, S.A. Thermal degradation of carbohydrates, cellulose and their esters with boric acid. Polym. Sci. USSR 1972, 14, 511–516. [Google Scholar]
- Böeseken, J. The use of boric acid for the determination of the configuration of carbohydrates. In Advances in Carbohydrate Chemistry; Academic Press: New York, NY, USA, 1949; Volume 4, pp. 189–210. [Google Scholar]
- Weedon, B.C.L. Stereochemistry. In Carotenoids; Birkhäuser: Basel, Switzerland, 1971; pp. 267–323. [Google Scholar]
- Roy, S.; Dora, K.C.; Kumar, S.; Saklani, P.; Muthukumar, A.; Ozogul, F.; Harisankar, K.C.; Mutum, R.D.; Celine Hilda Mary, S.; Surasani, V.K.R.; et al. A critical review on technical advances and multifaceted role of carotenoids in human health with special emphasis on metabolic diseases. Phytochem. Rev. 2026, 25, 1067–1092. [Google Scholar] [CrossRef]
- Walton, T.J.; Britton, G.; Goodwin, T.W. Biosynthesis of xanthophylls in higher plants: Stereochemistry of hydroxylation at C-3. Biochem. J. 1969, 112, 383. [Google Scholar] [CrossRef]
- Eugster, C.H. New carotenoid structures and stereochemistry. In Carotenoid Chemistry and Biochemistry; Pergamon: Oxford, UK, 1982; pp. 1–26. [Google Scholar]
- Zechmeister, L.; Lemmon, R.M. Contribution to the stereochemistry of cryptoxanthin and zeaxanthin. J. Am. Chem. Soc. 1944, 66, 317–322. [Google Scholar] [CrossRef]
- Sujak, A.; Mazurek, P.; Gruszecki, W.I. Xanthophyll pigments lutein and zeaxanthin in lipid multibilayers formed with dimyristoylphosphatidylcholine. J. Photochem. Photobiol. B 2002, 68, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Zechmeister, L. Cis–trans isomerization and stereochemistry of carotenoids and diphenyl-polyenes. Chem. Rev. 1944, 34, 267–344. [Google Scholar]
- Niyogi, K.K.; Björkman, O.; Grossman, A.R. The roles of specific xanthophylls in photoprotection. Proc. Natl. Acad. Sci. USA 1997, 94, 14162–14167. [Google Scholar]
- Cazzaniga, S.; Bressan, M.; Carbonera, D.; Agostini, A.; Dall’Osto, L. Differential roles of carotenes and xanthophylls in photosystem I photoprotection. Biochemistry 2016, 55, 3636–3649. [Google Scholar] [CrossRef]
- Kholili, U.; Wicaksono, A.B.; Hidayat, A.A.; Bintoro, U.Y.; Soetjipto, S.; Aryati, A.; Defianto, M.Z.F.; Miftahussurur, M. Xanthophyll-Rich Extracts from Garcinia dulcis Pulp as Potential Anti-Hepatocellular Carcinoma Functional Food. Nutrients 2026, 18, 670. [Google Scholar] [CrossRef]
- Baroli, I.; Niyogi, K.K. Molecular genetics of xanthophyll-dependent photoprotection in green algae and plants. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2000, 355, 1385–1394. [Google Scholar] [CrossRef]
- Dall’Osto, L.; Fiore, A.; Cazzaniga, S.; Giuliano, G.; Bassi, R. Different roles of α- and β-branch xanthophylls in photosystem assembly and photoprotection. J. Biol. Chem. 2007, 282, 35056–35068. [Google Scholar] [CrossRef]
- Esteban, R.; Olano, J.M.; Castresana, J.; Fernández-Marín, B.; Hernández, A.; Becerril, J.M.; García-Plazaola, J.I. Distribution and evolutionary trends of photoprotective isoprenoids (xanthophylls and tocopherols) within the plant kingdom. Physiol. Plant. 2009, 135, 379–389. [Google Scholar] [CrossRef]
- Demmig-Adams, B.; Adams, W.W. Light stress and photoprotection related to the xanthophyll cycle. In Causes of Photooxidative Stress and Amelioration of Defense Systems in Plants; CRC Press: Boca Raton, FL, USA, 2019; pp. 105–126. [Google Scholar]
- Emiliani, J.; D’Andrea, L.; Falcone Ferreyra, M.L.; Maulión, E.; Rodriguez, E.; Rodriguez-Concepción, M.; Casati, P. A role for β,β-xanthophylls in Arabidopsis UV-B photoprotection. J. Exp. Bot. 2018, 69, 4921–4933. [Google Scholar] [CrossRef] [PubMed]
- Takaichi, S.; Maoka, T.; Masamoto, K. Myxoxanthophyll in Synechocystis sp. PCC 6803 is myxol 2′-dimethyl-fucoside, (3R,2′S)-myxol 2′-(2,4-di-O-methyl-α-L-fucoside), not rhamnoside. Plant Cell Physiol. 2001, 42, 756–762. [Google Scholar] [CrossRef] [PubMed]
- Takaichi, S.; Mochimaru, M.; Maoka, T. Presence of free myxol and 4-hydroxymyxol and absence of myxol glycosides in Anabaena variabilis ATCC 29413, and proposal of a biosynthetic pathway of carotenoids. Plant Cell Physiol. 2006, 47, 211–216. [Google Scholar] [CrossRef]
- Shindo, K.; Kikuta, K.; Suzuki, A.; Katsuta, A.; Kasai, H.; Yasumoto-Hirose, M.; Takaichi, S. Rare carotenoids, (3R)-saproxanthin and (3R,2′S)-myxol, isolated from novel marine bacteria (Flavobacteriaceae) and their antioxidative activities. Appl. Microbiol. Biotechnol. 2007, 74, 1350–1357. [Google Scholar] [CrossRef]
- Wan, L.L.; Zhuo, R.Y.; He, D.Y.; Li, J.X.; Sun, C.; Chen, C.; Xu, L. Salinimicrobium aquimaris sp. nov., a novel Flavobacteriaceae member isolated from seawater in the tidal region of Danzhou, Hainan Island. Intern. J. Syst. Evol. Microbiol. 2026, 76, 007064. [Google Scholar] [CrossRef]
- Teramoto, M.; Onodera, K.-I.; Moriyama, H.; Komatsu, A.; Akakabe, M.; Nishijima, M. Aurantiacicella marina gen. nov., sp. nov., a myxol-producing bacterium from surface seawater. Int. J. Syst. Evol. Microbiol. 2016, 66, 248–254. [Google Scholar] [CrossRef]
- Manh, H.D.; Matsuo, Y.; Katsuta, A.; Matsuda, S.; Shizuri, Y.; Kasai, H. Robiginitalea myxolifaciens sp. nov., a novel myxol-producing bacterium isolated from marine sediment, and emended description of the genus Robiginitalea. Int. J. Syst. Evol. Microbiol. 2008, 58, 1660–1664. [Google Scholar] [CrossRef] [PubMed]
- Misawa, N. Carotenoid β-ring hydroxylase and ketolase from marine bacteria—Promiscuous enzymes for synthesizing functional xanthophylls. Mar. Drugs 2011, 9, 757–771. [Google Scholar] [CrossRef]
- Mochimaru, M.; Masukawa, H.; Maoka, T.; Mohamed, H.E.; Vermaas, W.F.; Takaichi, S. Substrate specificities and availability of fucosyltransferase and β-carotene hydroxylase for myxol 2′-fucoside synthesis in Anabaena sp. strain PCC 7120 compared with Synechocystis sp. strain PCC 6803. J. Bacteriol. 2008, 190, 6726–6733. [Google Scholar] [CrossRef]
- Mai, T.T. A Study of Cyanobacteria Metabolites, Exopolysaccharides and Carotenoids, in Context of Applied Phycology. Doctoral Dissertation, New Mexico State University, Las Cruces, NM, USA, 2021. [Google Scholar]
- Takaichi, S.; Maoka, T.; Mochimaru, M. Unique carotenoids in the terrestrial cyanobacterium Nostoc commune NIES-24: 2-Hydroxymyxol 2′-fucoside, nostoxanthin and canthaxanthin. Curr. Microbiol. 2009, 59, 413–419. [Google Scholar] [CrossRef]
- Takaichi, S.; Mochimaru, M.; Maoka, T.; Katoh, H. Myxol and 4-ketomyxol 2′-fucosides, not rhamnosides, from Anabaena sp. PCC 7120 and Nostoc punctiforme PCC 73102, and proposal for the biosynthetic pathway of carotenoids. Plant Cell Physiol. 2005, 46, 497–504. [Google Scholar] [CrossRef]
- Madhour, A.; Anke, H.; Mucci, A.; Davoli, P.; Weber, R.W. Biosynthesis of the xanthophyll plectaniaxanthin as a stress response in the red yeast Dioszegia (Tremellales, Heterobasidiomycetes, Fungi). Phytochemistry 2005, 66, 2617–2626. [Google Scholar] [CrossRef]
- Bae, M.; Lee, T.H.; Yokoyama, H.; Boettger, H.G.; Chichester, C.O. The occurrence of plectaniaxanthin in Cryptococcus laurentii. Phytochemistry 1971, 10, 625–629. [Google Scholar] [CrossRef]
- Røonneberg, H.; Borch, G.; Buchecker, R.; Arpin, N.; Liaaen-Jensen, S. Chirality of plectaniaxanthin. Phytochemistry 1982, 21, 2087–2090. [Google Scholar] [CrossRef]
- Liu, I.; Yokoyama, H.; Simpson, K.L.; Chichester, C.O. Isolation and identification of 2-hydroxyplectaniaxanthin from Rhodotorula aurantiaca. Phytochemistry 1973, 12, 2953–2956. [Google Scholar] [CrossRef]
- Takaichi, S.; Maoka, T.; Takasaki, K.; Hanada, S. Carotenoids of Gemmatimonas aurantiaca (Gemmatimonadetes): Identification of a novel carotenoid, deoxyoscillol 2-rhamnoside, and proposed biosynthetic pathway of oscillol 2,2′-dirhamnoside. Microbiology 2010, 156, 757–763. [Google Scholar] [CrossRef] [PubMed]
- Foss, P.; Skulberg, O.M.; Kilaas, L.; Liaaen-Jensen, S. The carbohydrate moieties bound to the carotenoids myxol and oscillol and their chemosystematic applications. Phytochemistry 1986, 25, 1127–1132. [Google Scholar] [CrossRef]
- Tsuchiya, T.; Takaichi, S.; Misawa, N.; Maoka, T.; Miyashita, H.; Mimuro, M. The cyanobacterium Gloeobacter violaceus PCC 7421 uses bacterial-type phytoene desaturase in carotenoid biosynthesis. FEBS Lett. 2005, 579, 2125–2129. [Google Scholar] [CrossRef] [PubMed]
- Aakermann, T.; Skulberg, O.M. A comparison of the carotenoids of strains of Oscillatoria and Spirulina (Cyanobacteria). Biochem. Syst. Ecol. 1992, 20, 761–769. [Google Scholar] [CrossRef]
- Francis, G.W.; Hertzberg, S.; Andersen, K.; Liaaen-Jensen, S. New carotenoid glycosides from Oscillatoria limosa. Phytochemistry 1970, 9, 629–635. [Google Scholar] [CrossRef]
- Hertzberg, S.; Liaaen-Jensen, S.; Siegelman, H.W. The carotenoids of blue-green algae. Phytochemistry 1971, 10, 3121–3127. [Google Scholar] [CrossRef]
- Takaichi, S.; Mochimaru, M. Carotenoids, their diversity and carotenogenesis in cyanobacteria. In Handbook on Cyanobacteria: Biochemistry, Biotechnology and Applications; Nova Science Publishers: New York, NY, USA, 2009; pp. 399–428. [Google Scholar]
- Takaichi, S. Carotenoids in phototrophic microalgae: Distributions and biosynthesis. In Pigments from Microalgae Handbook; Springer International Publishing: Cham, Switzerland, 2020; pp. 19–41. [Google Scholar]
- Barahona, S.; Yuivar, Y.; Socias, G.; Alcaíno, J.; Cifuentes, V.; Baeza, M. Identification and characterization of yeasts isolated from sedimentary rocks of Union Glacier at the Antarctica. Extremophiles 2016, 20, 479–491. [Google Scholar] [CrossRef]
- López, G.D.; Álvarez-Rivera, G.; Carazzone, C.; Ibáñez, E.; Leidy, C.; Cifuentes, A. Bacterial carotenoids: Extraction, characterization, and applications. Crit. Rev. Anal. Chem. 2023, 53, 1239–1262. [Google Scholar] [CrossRef]
- Dufossé, L. Microbial pigments from bacteria, yeasts, fungi, and microalgae for the food and feed industries. In Natural and Artificial Flavoring Agents and Food Dyes; Academic Press: London, UK, 2018; pp. 113–132. [Google Scholar]
- Dufossé, L. Back to nature, microbial production of pigments and colorants for food use. In Advances in Food and Nutrition Research; Academic Press: London, UK, 2022; Volume 102, pp. 93–122. [Google Scholar]
- Martín, J.F.; Gudiña, E.; Barredo, J.L. Conversion of β-carotene into astaxanthin: Two separate enzymes or a bifunctional hydroxylase–ketolase protein? Microb. Cell Fact. 2008, 7, 3. [Google Scholar]
- Ojima, K.; Breitenbach, J.; Visser, H.; Setoguchi, Y.; Tabata, K.; Hoshino, T.; Sandmann, G. Cloning of the astaxanthin synthase gene from Xanthophyllomyces dendrorhous (Phaffia rhodozyma) and its assignment as a β-carotene 3-hydroxylase/4-ketolase. Mol. Genet. Genom. 2006, 275, 148–158. [Google Scholar] [CrossRef] [PubMed]
- Pollmann, H.; Breitenbach, J.; Wolff, H.; Bode, H.B.; Sandmann, G. Combinatorial biosynthesis of novel multi-hydroxy carotenoids in the red yeast Xanthophyllomyces dendrorhous. J. Fungi 2017, 3, 9. [Google Scholar] [CrossRef] [PubMed]
- Del Toro-Sánchez, L.; Sánchez, S.; Ortiz, M.A.; Villanueva, S.; Lugo-Cervantes, E. Generation of aroma compounds from Ditaxis heterantha by Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2006, 72, 155–162. [Google Scholar] [CrossRef]
- Linden, H. Carotenoid hydroxylase from Haematococcus pluvialis: cDNA sequence, regulation and functional complementation. Biochim. Biophys. Acta Gene Struct. Expr. 1999, 1446, 203–212. [Google Scholar]
- Osawa, A.; Harada, H.; Choi, S.K.; Misawa, N.; Shindo, K. Production of caloxanthin 3′-β-D-glucoside, zeaxanthin 3,3′-β-D-diglucoside, and nostoxanthin in a recombinant Escherichia coli expressing system harboring seven carotenoid biosynthesis genes, including crtX and crtG. Phytochemistry 2011, 72, 711–716. [Google Scholar]
- Buchecker, R.; Liaaen-Jensen, S.; Borch, G.; Siegelman, H.W. Carotenoids of Anacystis nidulans, structures of caloxanthin and nostoxanthin. Phytochemistry 1976, 15, 1015–1018. [Google Scholar] [CrossRef]
- Kosourov, S.; Murukesan, G.; Jokela, J.; Allahverdiyeva, Y. Carotenoid biosynthesis in Calothrix sp. 336/3: Composition of carotenoids on full medium, during diazotrophic growth and after long-term H2 photoproduction. Plant Cell Physiol. 2016, 57, 2269–2282. [Google Scholar] [CrossRef]
- Hager, A.T.; Stransky, H. Das Carotinoidmuster und die Verbreitung des lichtinduzierten Xanthophyllcyclus in verschiedenen Algenklassen. Arch. Mikrobiol. 1970, 71, 132–163. [Google Scholar] [CrossRef]
- Yokoyama, A.; Miki, W.; Izumida, H.; Shizuri, Y. New trihydroxy-keto-carotenoids isolated from an astaxanthin-producing marine bacterium. Biosci. Biotechnol. Biochem. 1996, 60, 200–203. [Google Scholar] [CrossRef]
- Asker, D. Isolation and characterization of a novel, highly selective astaxanthin-producing marine bacterium. J. Agric. Food Chem. 2017, 65, 9101–9109. [Google Scholar] [CrossRef]
- Asker, D.; Awad, T.S.; Beppu, T.; Ueda, K. Screening and profiling of natural ketocarotenoids from environmental aquatic bacterial isolates. Food Chem. 2018, 253, 247–254. [Google Scholar] [CrossRef]
- Asker, D. High throughput screening and profiling of high-value carotenoids from a wide diversity of bacteria in surface seawater. Food Chem. 2018, 261, 103–111. [Google Scholar] [CrossRef]
- De Ridder, E.; Vandamme, P.; Willems, A. Carotenoid biosynthesis in bacteria: The crt gene products and their functional roles in the carotenogenic pathways. Crit. Rev. Microbiol. 2025, 52, 64–83. [Google Scholar] [CrossRef]
- Kathiresan, A.; Kim, H.S.; Li, C.; Dong, K.; Srinivasan, S.; Lee, S.S. Litorerythrobacter xanthomarinus gen. nov., sp. nov., a novel marine bacterium with distinct phenotypic traits from tidal mudflat sediment. Antonie Leeuwenhoek 2026, 119, 34. [Google Scholar] [CrossRef]
- Liu, H.; Tan, K.S.; Zhang, X.; Zhang, H.; Cheng, D.; Ting, Y.; Zheng, H. Comparison of gut microbiota between golden and brown noble scallop Chlamys nobilis and its association with carotenoids. Front. Microbiol. 2020, 11, 36. [Google Scholar] [CrossRef]
- Bjerkeng, B.; Hatlen, B.; Jobling, M. Astaxanthin and its metabolites idoxanthin and crustaxanthin in flesh, skin, and gonads of sexually immature and maturing Arctic charr (Salvelinus alpinus L.). Comp. Biochem. Physiol. B 2000, 125, 395–404. [Google Scholar]
- Schiedt, K.; Mayer, H.; Vecchi, M.; Glinz, E.; Storebakken, T. Metabolism of carotenoids in salmonids. Part 2. Distribution and absolute configuration of idoxanthin in various organs and tissues of one Atlantic salmon (Salmo salar L.) fed with astaxanthin. Helv. Chim. Acta 1988, 71, 881–886. [Google Scholar] [CrossRef]
- Aas, G.H.; Bjerkeng, B.; Hatlen, B.; Storebakken, T. Idoxanthin, a major carotenoid in the flesh of Arctic charr (Salvelinus alpinus) fed diets containing astaxanthin. Aquaculture 1997, 150, 135–142. [Google Scholar] [CrossRef]
- Märki-Fischer, E.; Eugster, C.H. Das Carotinoidspektrum der Antheren und Petalen von Lilium tigrinum cv. ‘Red Night’. Helv. Chim. Acta 1985, 68, 1708–1715. [Google Scholar] [CrossRef]
- Goodwin, T.W. Distribution of carotenoids. In Methods in Enzymology; Academic Press: New York, NY, USA, 1992; Volume 213, pp. 167–172. [Google Scholar]
- Eugster, C.H. Recent progress in carotenoid structures. In Carotenoids: Chemistry and Biology; Springer: Berlin/Heidelberg, Germany, 1990; pp. 1–20. [Google Scholar]
- Jensen, S.L. The carotenoid group of natural products. In Second Supplements to the 2nd Edition of Rodd’s Chemistry of Carbon Compounds; Elsevier: Amsterdam, The Netherlands, 1991; pp. 57–101. [Google Scholar]
- Rodríguez-Sáiz, M.; de la Fuente, J.L.; Barredo, J.L. Xanthophyllomyces dendrorhous for the industrial production of astaxanthin. Appl. Microbiol. Biotechnol. 2010, 88, 645–658. [Google Scholar] [CrossRef]
- Barredo, J.L.; García-Estrada, C.; Kosalkova, K.; Barreiro, C. Biosynthesis of astaxanthin as a main carotenoid in the heterobasidiomycetous yeast Xanthophyllomyces dendrorhous. J. Fungi 2017, 3, 44. [Google Scholar] [CrossRef]
- Visser, H.; Sandmann, G.; Verdoes, J.C. Xanthophylls in fungi: Metabolic engineering of the astaxanthin biosynthetic pathway in Xanthophyllomyces dendrorhous. In Microbial Processes and Products; Humana Press: Totowa, NJ, USA, 2005; pp. 257–272. [Google Scholar]
- Huang, R.; Wang, J.; Ding, R.; Liu, Y.; Su, J.; Chen, Z.; Fan, Y.; Li, F.; Wang, L.; Liu, X.; et al. A conserved E3 ubiquitin lig-ase rewires carotenoid, sterol, and lipid metabolism in Xanthophyllomyces dendrorhous. Proc. Natl. Acad. Sci. USA 2026, 123, e2530496123. [Google Scholar]
- Kikukawa, H.; Okaya, T.; Maoka, T.; Miyazaki, M.; Murofushi, K.; Kato, T.; Hara, K.Y. Carotenoid nostoxanthin production by Sphingomonas sp. SG73 isolated from deep sea sediment. Mar. Drugs 2021, 19, 274. [Google Scholar] [CrossRef]
- Jiang, L.; Seo, J.; Peng, Y.; Jeon, D.; Lee, J.H.; Kim, C.Y.; Lee, J. Sphingomonas nostoxanthinifaciens sp. nov., a nostoxanthin-producing bacterium alleviating salt stress in Arabidopsis seedlings via reactive oxygen species scavenging. Front. Microbiol. 2023, 14, 1101150. [Google Scholar]
- Raman, J.; Kim, J.S.; Ko, Y.J.; Kim, S.J. Nostoxanthin biosynthesis by Sphingomonas species (COS14-R2): Isolation, identification, and optimization of culture conditions. Curr. Microbiol. 2024, 81, 453. [Google Scholar] [CrossRef]
- Czygan, F.C. Zum Vorkommen von Crustaxanthin (3,3′,4,4′-Tetraoxi-β-carotin) und Phoenicopteron (4-Oxo-α-carotin) in Aplanosporen von Haematococcus pluvialis. Flora 1968, 159, 339–345. [Google Scholar] [CrossRef]
- Czeczuga, B.; Czeczuga-Semeniuk, E.; Semeniuk, A. Carotenoids and carotenoproteins in Asellus aquaticus L. (Crustacea: Isopoda). Folia Biol. 2005, 53, 109–114. [Google Scholar] [CrossRef][Green Version]
- Strain, H.H.; Benton, F.L.; Grandolfo, M.C.; Aitzetmüller, K.; Svec, W.; Katz, J. Heteroxanthin, diatoxanthin and diadinoxanthin from Tribonema aequale. Phytochemistry 1970, 9, 2561–2565. [Google Scholar] [CrossRef]
- Nitsche, H. Heteroxanthin in Euglena gracilis. Arch. Mikrobiol. 1973, 90, 151–155. [Google Scholar] [CrossRef]
- Strain, H.H.; Aitzetmüller, K.; Svec, W.A.; Katz, J.J. Structure of heteroxanthin, a unique xanthophyll from the Xanthophyceae (Heterokontae). J. Chem. Soc. D Chem. Commun. 1970, 14, 876–877. [Google Scholar] [CrossRef]
- Borodina, A.V. Features of carotenoid profile in Black Sea bivalve mollusks. J. Evol. Biochem. Physiol. 2022, 58, 943–954. [Google Scholar] [CrossRef]
- Tan, K.; Zhang, H.; Zheng, H. Carotenoid content and composition: A special focus on commercially important fish and shellfish. Crit. Rev. Food Sci. Nutr. 2024, 64, 544–561. [Google Scholar] [CrossRef]
- Mohd Hassan, N.; Yusof, N.A.; Yahaya, A.F.; Mohd Rozali, N.N.; Othman, R. Carotenoids of Capsicum fruits: Pigment profile and health-promoting functional attributes. Antioxidants 2019, 8, 469. [Google Scholar] [CrossRef]
- del Rocío Gómez-García, M.; Ochoa-Alejo, N. Biochemistry and molecular biology of carotenoid biosynthesis in chili peppers (Capsicum spp.). Int. J. Mol. Sci. 2013, 14, 19025–19053. [Google Scholar] [CrossRef]
- Sandmann, G. Generation of stable homozygous transformants of diploid yeasts such as Xanthophyllomyces dendrorhous. Appl. Microbiol. Biotechnol. 2022, 106, 4921–4927. [Google Scholar] [CrossRef]
- Sandmann, G. Carotenoids and their biosynthesis in fungi. Molecules 2022, 27, 1431. [Google Scholar] [CrossRef]
- Shahi, R.; Lohith Kumar, N.; Mishra, A. Microbial Carotenoids: Biosynthesis Pathways, Produc-tion Strategies, and Their Industrial Applications. In Biotechnological Solutions for a Sustainable Future: From Soil Health to Industrial Applications; Springer: Singapore, 2026; Volume 55, pp. 279–305. [Google Scholar]
- Johnson, E.A. Phaffia rhodozyma: Colorful odyssey. Int. Microbiol. 2003, 6, 169–174. [Google Scholar] [CrossRef] [PubMed]
- Tavares, D.Q.; Santos, G.C.; Mangussi, I.M.A.S.; Vital, L.R.; Nascimento, L.M.; Oliveira, C.R. Xanthophylls: Potential benefits in protecting against UV burns. Braz. J. Biol. 2025, 85, e288662. [Google Scholar] [CrossRef] [PubMed]
- Rezanka, T.; Palyzová, A. Yeast need not be used only in beer production. I. The carotenoid astaxanthin produced by yeast. Kvas. Prum. 2020, 66, 208–214. [Google Scholar]
- Haugan, J.A. Algal carotenoids 54. Carotenoids of brown algae (Phaeophyceae). Biochem. Syst. Ecol. 1994, 22, 31–41. [Google Scholar] [CrossRef]
- Nagy, V.; Agocs, A.; Turcsi, E.; Molnár, P.; Szabó, Z.; Deli, J. Latoxanthin, a minor carotenoid isolated from the fruits of yellow paprika (Capsicum annuum var. lycopersiciforme flavum). Tetrahedron Lett. 2007, 48, 9012–9014. [Google Scholar] [CrossRef]
- Takaichi, S. Distribution, biosynthesis, and function of carotenoids in oxygenic phototrophic algae. Mar. Drugs 2025, 23, 62. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Ashihara, H.; Waller, G.R. Purine and purine alkaloid metabolism in Camellia and Coffea plants. Phytochemistry 1992, 31, 2575–2584. [Google Scholar] [CrossRef]
- Maoka, T.; Hashimoto, K.; Akimoto, N.; Fujiwara, Y. Structures of five new carotenoids from the oyster Crassostrea gigas. J. Nat. Prod. 2001, 64, 578–581. [Google Scholar] [CrossRef]
- Maoka, T.; Fujiwara, Y.; Hashimoto, K.; Akimoto, N. Structures of new carotenoids with a 3,4-dihydroxy-γ-end group from the oyster Crassostrea gigas. Chem. Pharm. Bull. 2005, 53, 1207–1209. [Google Scholar] [CrossRef]
- Dembitsky, V.M.; Maoka, T. Allenic and cumulenic lipids. Prog. Lipid Res. 2007, 46, 328–375. [Google Scholar] [CrossRef] [PubMed]
- Maoka, T.; Akimoto, N.; Yim, M.-J.; Hosokawa, M.; Miyashita, K. A new C37-skeletal carotenoid from the clam Paphia amabillis. J. Agric. Food Chem. 2008, 56, 12069–12072. [Google Scholar] [CrossRef]
- Maoka, T.; Fujiwara, Y.; Hashimoto, K. Carotenoids in three species of corbicula clams, Corbicula japonica, Corbicula sandai, and Corbicula sp. (Chinese freshwater corbicula clam). J. Agric. Food Chem. 2005, 53, 8357–8364. [Google Scholar] [CrossRef]
- Maoka, T.; Fujiwara, Y.; Hashimoto, K.; Akimoto, N. Structure of new carotenoids from the corbicula clam Corbicula japonica. J. Nat. Prod. 2005, 68, 1341–1344. [Google Scholar] [CrossRef]
- Siddiq, A.; Dembitsky, V.M. Acetylenic anticancer agents. Anti-Cancer Agents Med. Chem. 2008, 8, 132–170. [Google Scholar] [CrossRef]
- Kantha, S.S. Carotenoids of edible molluscs: A review. J. Food Biochem. 1989, 13, 429–442. [Google Scholar] [CrossRef]
- Dembitsky, V.M.; Levitsky, D.O.; Gloriozova, T.A.; Poroikov, V.V. Acetylenic aquatic anticancer agents and related compounds. Nat. Prod. Commun. 2006, 1, 773–811. [Google Scholar] [CrossRef]
- Maoka, T. Recent progress in structural studies of carotenoids in animals and plants. Arch. Biochem. Biophys. 2009, 483, 191–195. [Google Scholar] [CrossRef] [PubMed]
- Dembitsky, V.M. Anticancer activity of natural and synthetic acetylenic lipids. Lipids 2006, 41, 883–924. [Google Scholar] [CrossRef]
- Tsushima, M.; Maoka, T.; Matsuno, T. Structure of carotenoids with 5,6-dihydro-β-end group from the spindle shell Fusinus perplexus. J. Nat. Prod. 2001, 64, 1139–1142. [Google Scholar] [CrossRef]
- Maoka, T. Carotenoids in marine animals. Mar. Drugs 2011, 9, 278–293. [Google Scholar] [CrossRef]
- Dembitsky, V.M.; Rosenberg, G.S.; Zanfera, V.M. The Evolutionary Pathway to the Biomembrane: The Role of Low Molecular Weight Polyols in the Formation of the Protomembrane; IEVRB RAS Editorial Department: Moscow, Russia, 2023; 128p. [Google Scholar]
- Dembitsky, V.M. Ether lipids of the organic world: Formation and biotransformation. In Fats for the Future; Cambie, R.Q., Ed.; Ellis Harwood Series in Food Science and Technology; Van Nostrand Reinhold/Avi: London, UK, 1989; pp. 173–189. [Google Scholar]
- Deamer, D. Origins of life research: The conundrum between laboratory and field simulations of messy environments. Life 2022, 12, 1429. [Google Scholar] [CrossRef]
- Peretó, J. Prebiotic chemistry that led to life. In Handbook of Astrobiology; CRC Press: Boca Raton, FL, USA, 2019; pp. 219–233. [Google Scholar]
- Gómez-Márquez, J. The origin of life and cellular systems: A continuum from prebiotic chemistry to biodiversity. Life 2025, 15, 1745. [Google Scholar] [CrossRef]
- Anton, O.; Stirnemann, G. Computational studies of prebiotic chemistry at the age of machine learning: From recent breakthroughs to future revolutions. ChemSystemsChem 2026, 8, e00057. [Google Scholar] [CrossRef]
- Dembitsky, V.M.; Terent’ev, A.O.; Baranin, S.V.; Scorei, R.I. Life with Boron: Steroid Architecture and the Chemistry of Marine Boronosteroids. Mar. Drugs 2026, 24, 113. [Google Scholar] [CrossRef] [PubMed]
- Dembitsky, V.M.; Terent’ev, A.O.; Baranin, S.V.; Scorei, I.R. Boron’s Double Edge—Antibiotics, Toxins, and the Fine Line Between Them. Molecules 2026, 31, 1021. [Google Scholar] [CrossRef]
- Dembitsky, V.M.; Terent’ev, A.O.; Baranin, S.V. Functional Plasticity of Microbial Siderophores in Iron- and Boron-Rich Niches. Appl. Microbiol. 2026, 6, 50. [Google Scholar] [CrossRef]
- Le Blanc, O.H., Jr. Tetraphenylborate conductance through lipid bilayer membranes. Biochim. Biophys. Acta (BBA)-Biomembr. 1969, 193, 350–360. [Google Scholar] [CrossRef]
- Bakowsky, H.; Richter, T.; Kneuer, C.; Hoekstra, D.; Rothe, U.; Bendas, G.; Ehrhardt, C.; Bakowsky, U. Adhesion characteristics and stability assessment of lectin-modified liposomes for site-specific drug delivery. Biochim. Biophys. Acta (BBA)-Biomembr. 2008, 1778, 242–249. [Google Scholar] [CrossRef]
- Mautner-Culetto, A.; Huhn, M.; Schwarz, S.; Tian, L.; Hamballer, M.; Afonin, S.; Martinac, B.; Buth, G.; Watts, A.; Weinschenk, S.; et al. Role of the lipid matrix in the action of local anesthetics. Biochim. Biophys. Acta (BBA)-Biomembr. 2026, 1868, 184504. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Feature | Description | Functional Consequence | Level of Evidence |
|---|---|---|---|
| Boron coordination (diol binding) | Formation of cyclic borate esters with xanthophyll hydroxyl groups | Alters electronic structure and polarity | Established (model compounds) |
| Trigonal ↔ tetrahedral transition | pH-dependent boron speciation | Dynamic modulation of binding strength and geometry | Established (boron chemistry), inferred (xanthophylls) |
| Conjugated system perturbation | Interaction with π-electron system of carotenoids | Potential shift in absorption spectra and excited-state behavior | Hypothesized |
| Complex reversibility | Labile borate ester formation in aqueous environments | Enables adaptive or transient photoprotection | Established (general), untested in membranes |
| Property | Expected Role of Xanthophylls | Additional Effect of Boron Complexation | Biological Implication |
|---|---|---|---|
| Membrane rigidity | Orientation across lipid bilayer stabilizes structure | Enhanced cross-linking or anchoring via boron bridges | Increased membrane robustness |
| Lipid packing | Ordering of surrounding lipids | Modulation of packing density through complex formation | Altered permeability and fluidity |
| Light absorption | Dissipation of excess energy | Modified absorption/emission properties | Improved photoprotection |
| Reactive oxygen species (ROS) quenching | Scavenging and energy dissipation | Possible enhancement or modulation via boron coordination | Reduced oxidative damage (hypothetical) |
| System | Membrane Type | Xanthophyll Role | Potential Role of Boron Complexes | Evidence Status |
|---|---|---|---|---|
| Archaea (extreme environments) | Isoprenoid ether lipids | Structural stabilization, stress resistance | Additional photostabilization under high UV/salinity | Hypothetical |
| Bacteria | Glycerolipid membranes | Photoprotection (in phototrophs) | Modulation of membrane properties and light response | Hypothetical |
| Photosynthetic organisms | Thylakoid membranes | Non-photochemical quenching (NPQ) | Possible tuning of energy dissipation pathways | Speculative |
| Model systems (liposomes) | Artificial membranes | Controlled study systems | Platform to test boron–xanthophyll interactions | Experimentally accessible |
| Question | Significance | Suggested Approach |
|---|---|---|
| Do boron–xanthophyll complexes form in vivo? | Establishes biological relevance | Spectroscopy (11B NMR), imaging, isotope labeling |
| What are the photophysical changes upon complexation? | Determines photoprotective mechanism | Ultrafast spectroscopy, fluorescence studies |
| How stable are these complexes in membranes? | Defines functional viability | Model membrane experiments, varying pH/salinity |
| Can boron availability regulate membrane behavior? | Links chemistry to physiology | Controlled biological studies, boron modulation |
| Are there evolutionary advantages in extremophiles? | Supports ecological relevance | Comparative studies across taxa |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Share and Cite
Dembitsky, V.M.; Terent’ev, A.O. Boron–Vicinal Diol Xanthophyll Complexes as Emerging Photoprotective Adjuvants. Photochem 2026, 6, 22. https://doi.org/10.3390/photochem6020022
Dembitsky VM, Terent’ev AO. Boron–Vicinal Diol Xanthophyll Complexes as Emerging Photoprotective Adjuvants. Photochem. 2026; 6(2):22. https://doi.org/10.3390/photochem6020022
Chicago/Turabian StyleDembitsky, Valery M., and Alexander O. Terent’ev. 2026. "Boron–Vicinal Diol Xanthophyll Complexes as Emerging Photoprotective Adjuvants" Photochem 6, no. 2: 22. https://doi.org/10.3390/photochem6020022
APA StyleDembitsky, V. M., & Terent’ev, A. O. (2026). Boron–Vicinal Diol Xanthophyll Complexes as Emerging Photoprotective Adjuvants. Photochem, 6(2), 22. https://doi.org/10.3390/photochem6020022