
Rosmarinic Acid Attenuates Testicular Damage via Modulating Oxidative Stress and Apoptosis in Streptozotocin-Induced Diabetic Albino Mice

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
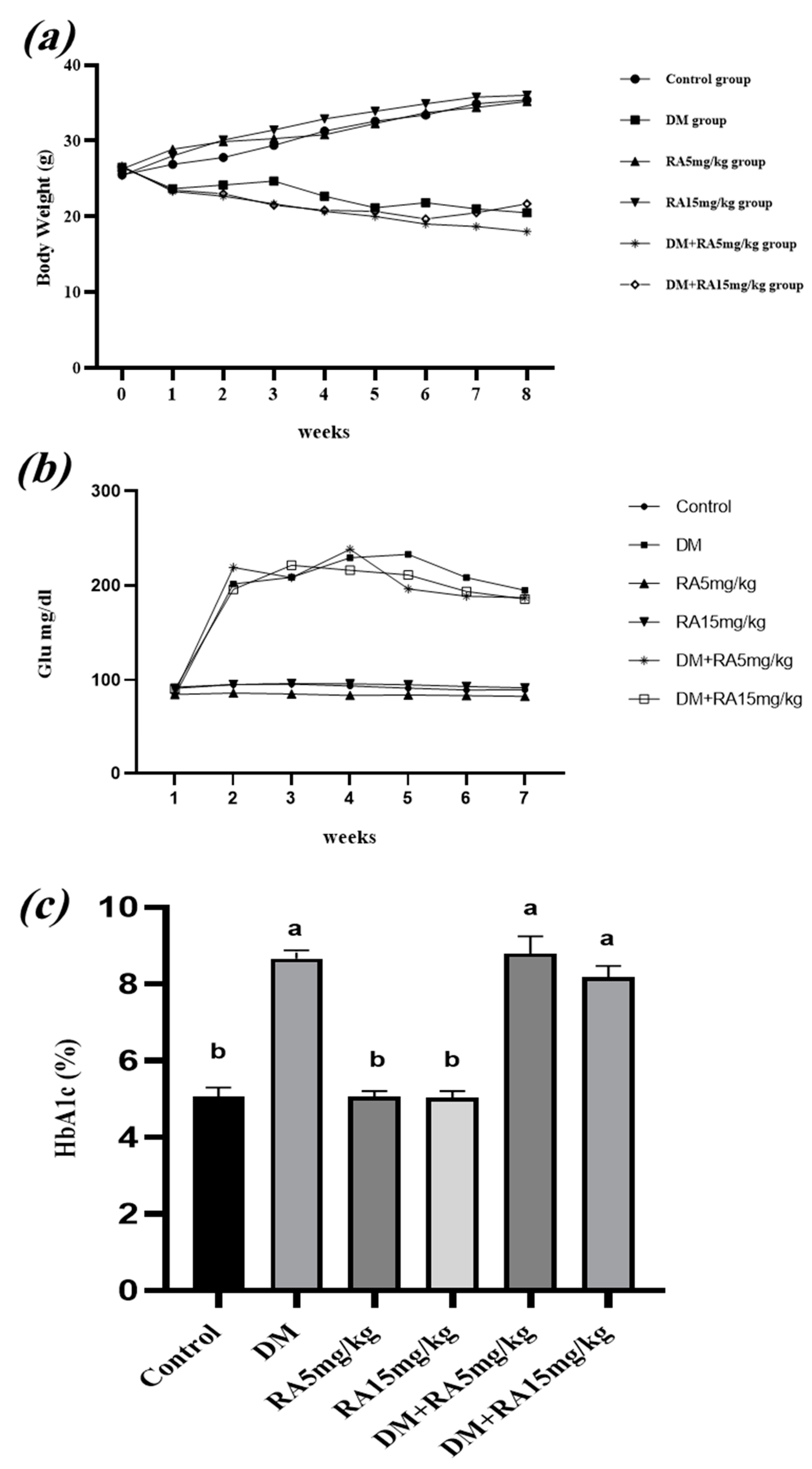
2.1. Effects of RA on Body Weight, FBG Levels, and HbA1c in Diabetic Mice
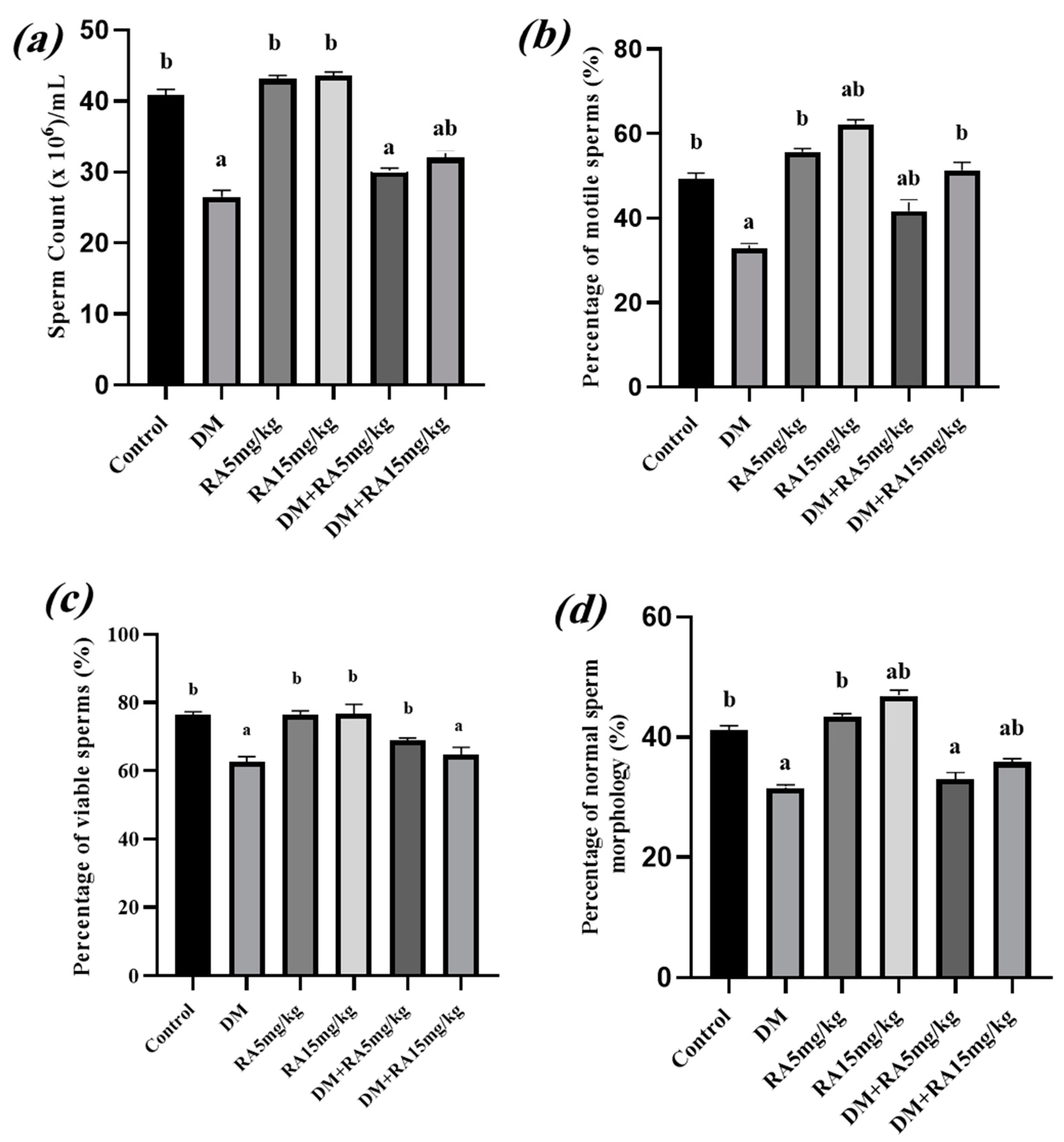
2.2. Effects of RA on Sperm Parameters in Diabetic Mice
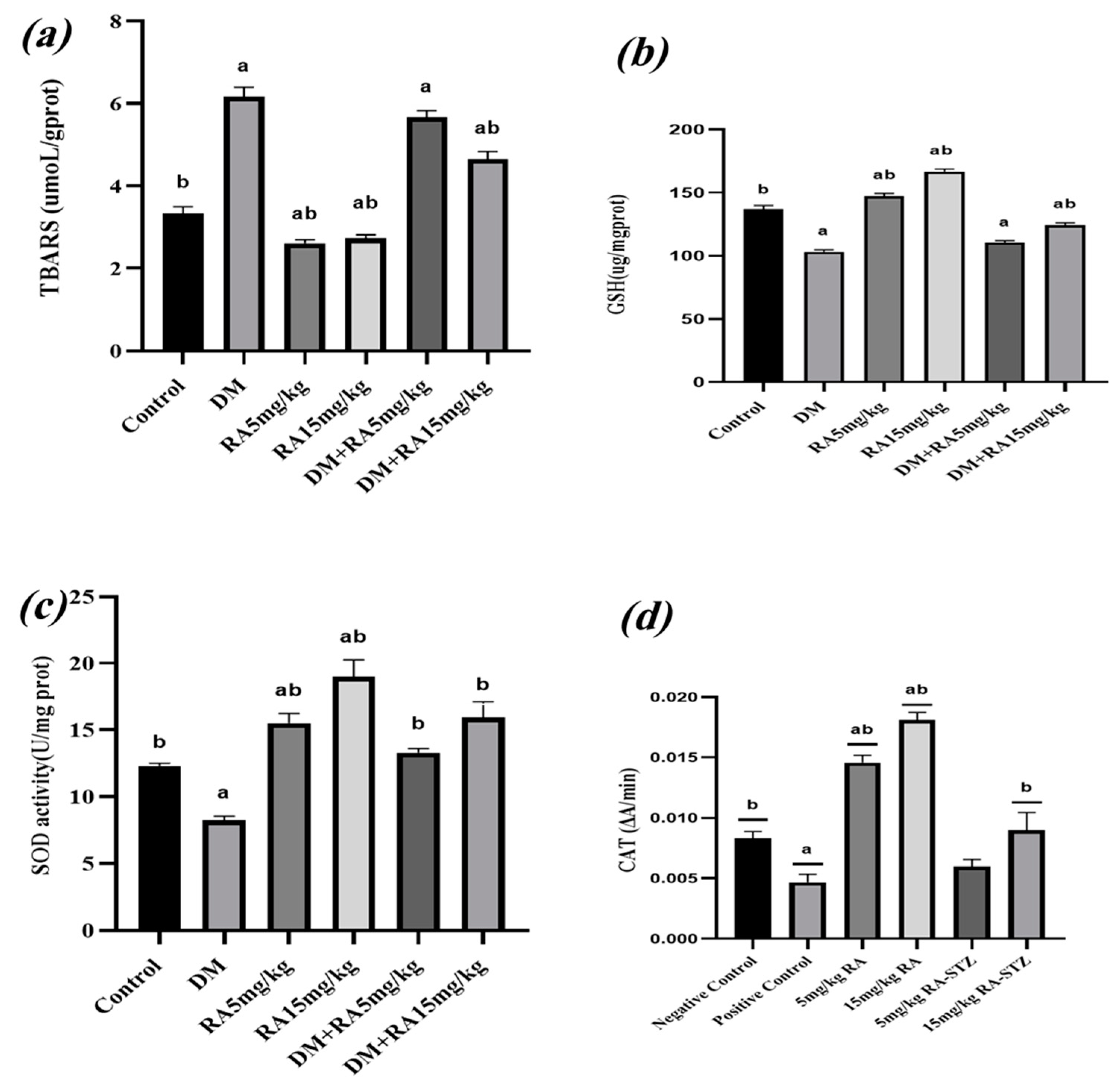
2.3. RA Mitigates Oxidative Stress in the Testes of Diabetic Mice
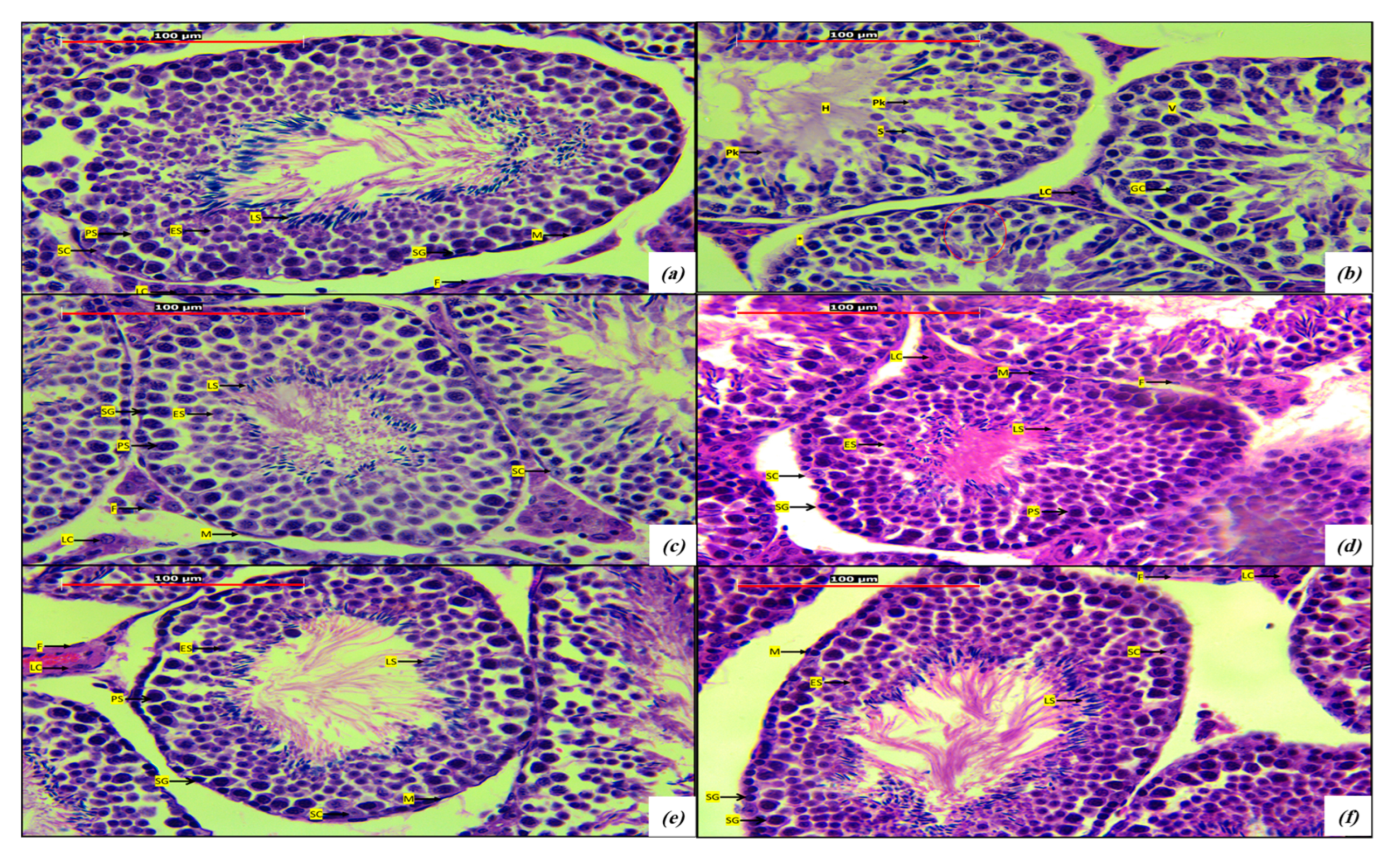
2.4. Histological Studies
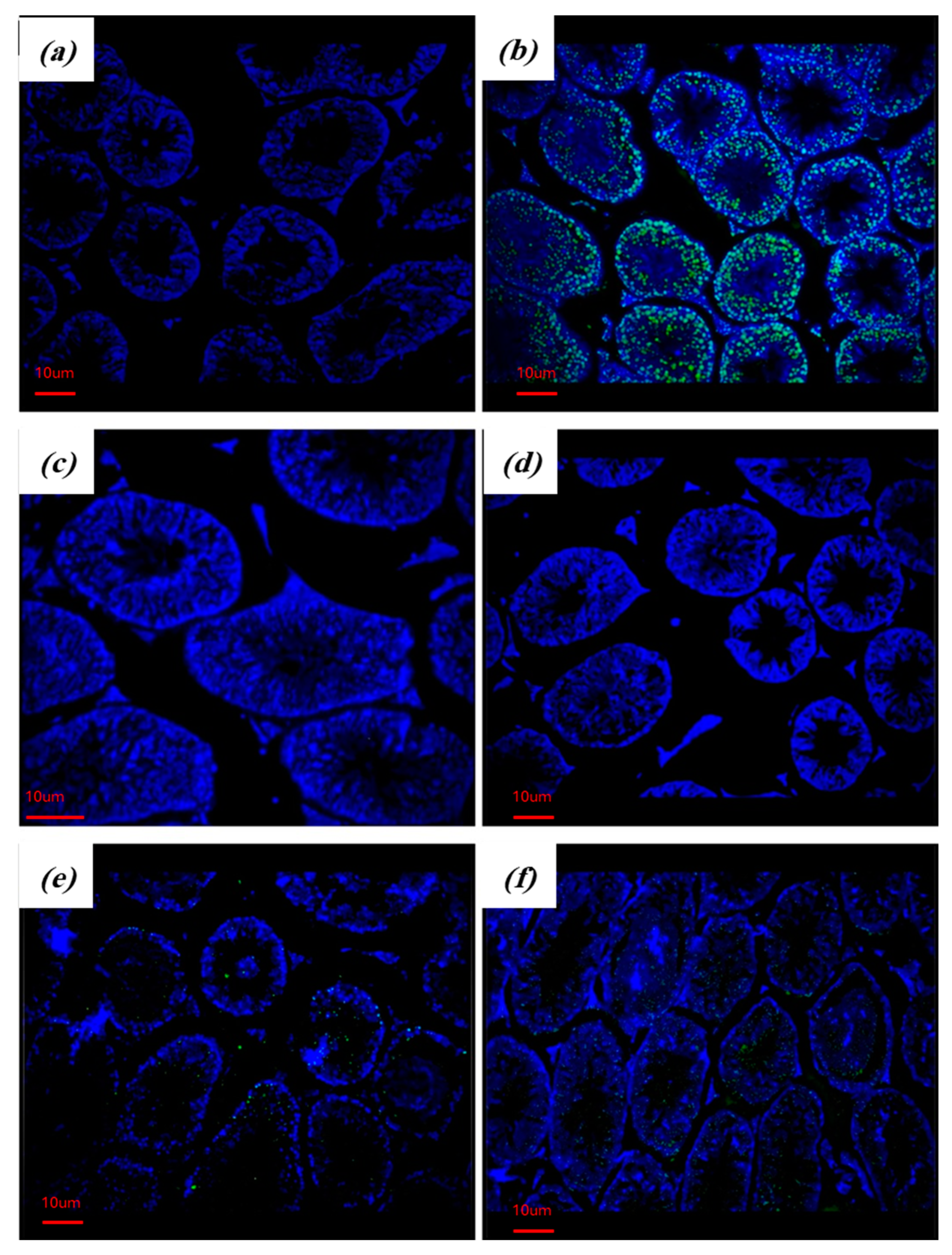
2.5. RA Attenuates Apoptosis in the Testes of Diabetic Mice
3. Discussion
4. Materials and Methods
4.1. Experimental Animals
4.2. Chemicals
4.3. Induction of DM
4.4. Experimental Groups
4.5. Animal Treatment and Sacrifice
4.6. Sperm Parameters
4.7. Glycated HemoglobinA1c (HbA1c)
4.8. Testicular Oxidative Stress Markers and Antioxidants
4.9. TUNEL Assay
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Asmat, U.; Abad, K.; Ismail, K. Diabetes mellitus and oxidative stress-A concise review. Saudi Pharm. J. 2016, 24, 547–553. [Google Scholar] [CrossRef] [PubMed]
- Lotfy, M.; Adeghate, J.; Kalasz, H.; Singh, J.; Adeghate, E. Chronic Complications of Diabetes Mellitus: A Mini Review. Curr. Diabetes Rev. 2017, 13, 3–10. [Google Scholar] [CrossRef]
- Hurrle, S.; Hsu, W.H. The etiology of oxidative stress in insulin resistance. Biomed. J. 2017, 40, 257–262. [Google Scholar] [CrossRef]
- Redza-Dutordoir, M.; Averill-Bates, D.A. Activation of apoptosis signalling pathways by reactive oxygen species. Biochim. Biophys. Acta 2016, 1863, 2977–2992. [Google Scholar] [CrossRef]
- Lam, P.L.; Wong, R.S.; Lam, K.H.; Hung, L.K.; Wong, M.M.; Yung, L.H.; Ho, Y.W.; Wong, W.Y.; Hau, D.K.; Gambari, R.; et al. The role of reactive oxygen species in the biological activity of antimicrobial agents: An updated mini review. Chem. Biol. Interact. 2020, 320, 109023. [Google Scholar] [CrossRef]
- Kaltsas, A. Oxidative Stress and Male Infertility: The Protective Role of Antioxidants. Medicina 2023, 59, 1769. [Google Scholar] [CrossRef]
- D‘Cruz, S.C.; Vaithinathan, S.; Jubendradass, R.; Mathur, P.P. Effects of plants and plant products on the testis. Asian J. Androl. 2010, 12, 468–479. [Google Scholar] [CrossRef] [PubMed]
- Nasim, N.; Sandeep, I.S.; Mohanty, S. Plant-derived natural products for drug discovery: Current approaches and prospects. Nucleus 2022, 65, 399–411. [Google Scholar] [CrossRef] [PubMed]
- Ngo, Y.L.; Lau, C.H.; Chua, L.S. Review on rosmarinic acid extraction, fractionation and its anti-diabetic potential. Food Chem. Toxicol. 2018, 121, 687–700. [Google Scholar] [CrossRef]
- Petersen, M.; Abdullah, Y.; Benner, J.; Eberle, D.; Gehlen, K.; Hücherig, S.; Janiak, V.; Kim, K.H.; Sander, M.; Weitzel, C.; et al. Evolution of rosmarinic acid biosynthesis. Phytochemistry 2009, 70, 1663–1679. [Google Scholar] [CrossRef]
- Petersen, M.; Simmonds, M.S. Rosmarinic acid. Phytochemistry 2003, 62, 121–125. [Google Scholar] [CrossRef] [PubMed]
- Al-Alami, Z.M.; Shraideh, Z.A.; Taha, M.O. Rosmarinic acid reverses the effects of metronidazole-induced infertility in male albino rats. Reprod. Fertil. Dev. 2017, 29, 1910–1920. [Google Scholar] [CrossRef] [PubMed]
- Raisi, A.; Davoodi, F.; Mohammadi, R. Protective effects of rosmarinic acid on testicular torsion-detorsion in an animal model. Iran. J. Vet. Surg. 2022, 17, 80–86. [Google Scholar] [CrossRef]
- Facondo, P.; Di Lodovico, E.; Delbarba, A.; Anelli, V.; Pezzaioli, L.C.; Filippini, E.; Cappelli, C.; Corona, G.; Ferlin, A. The impact of diabetes mellitus type 1 on male fertility: Systematic review and meta-analysis. Andrology 2022, 10, 426–440. [Google Scholar] [CrossRef] [PubMed]
- Condorelli, R.A.; La Vignera, S.; Mongioì, L.M.; Alamo, A.; Calogero, A.E. Diabetes Mellitus and Infertility: Different Pathophysiological Effects in Type 1 and Type 2 on Sperm Function. Front. Endocrinol. 2018, 9, 268. [Google Scholar] [CrossRef] [PubMed]
- Papachristoforou, E.; Lambadiari, V.; Maratou, E.; Makrilakis, K. Association of Glycemic Indices (Hyperglycemia, Glucose Variability, and Hypoglycemia) with Oxidative Stress and Diabetic Complications. J. Diabetes Res. 2020, 2020, 7489795. [Google Scholar] [CrossRef] [PubMed]
- Furman, B.L. Streptozotocin-Induced Diabetic Models in Mice and Rats. Curr. Protoc. Pharmacol. 2015, 70, 5–47. [Google Scholar] [CrossRef] [PubMed]
- Szkudelski, T. The mechanism of alloxan and streptozotocin action in B cells of the rat pancreas. Physiol. Res. 2001, 50, 537–546. [Google Scholar] [PubMed]
- Cummings, D.E.; Overduin, J.; Foster-Schubert, K.E. Gastric bypass for obesity: Mechanisms of weight loss and diabetes resolution. J. Clin. Endocrinol. Metab. 2004, 89, 2608–2615. [Google Scholar] [CrossRef]
- He, L.; He, T.; Farrar, S.; Ji, L.; Liu, T.; Ma, X. Antioxidants Maintain Cellular Redox Homeostasis by Elimination of Reactive Oxygen Species. Cell Physiol. Biochem. 2017, 44, 532–553. [Google Scholar] [CrossRef]
- Georgy, G.S.; Maher, O.W. Ellagic acid and rosmarinic acid attenuate doxorubicin-induced testicular injury in rats. J. Biochem. Mol. Toxicol. 2017, 31, e21937. [Google Scholar] [CrossRef]
- Nna, V.U.; Bakar, A.B.A.; Ahmad, A.; Mohamed, M. Diabetes-induced testicular oxidative stress, inflammation, and caspase-dependent apoptosis: The protective role of metformin. Arch. Physiol. Biochem. 2020, 126, 377–388. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.L.; Xu, Y.; Zhang, S.P.; Hou, J.; Zhu, H.B. Effect of rosmarinic acid on experimental diabetic nephropathy. Basic. Clin. Pharmacol. Toxicol. 2012, 110, 390–395. [Google Scholar] [CrossRef]
- Sotnikova, R.; Okruhlicova, L.; Vlkovicova, J.; Navarova, J.; Gajdacova, B.; Pivackova, L.; Fialova, S.; Krenek, P. Rosmarinic acid administration attenuates diabetes-induced vascular dysfunction of the rat aorta. J. Pharm. Pharmacol. 2013, 65, 713–723. [Google Scholar] [CrossRef]
- Domitrović, R.; Potočnjak, I.; Crnčević-Orlić, Z.; Škoda, M. Nephroprotective activities of rosmarinic acid against cisplatin-induced kidney injury in mice. Food Chem. Toxicol. 2014, 66, 321–328. [Google Scholar] [CrossRef] [PubMed]
- Tavafi, M.; Ahmadvand, H. Effect of rosmarinic acid on inhibition of gentamicin induced nephrotoxicity in rats. Tissue Cell 2011, 43, 392–397. [Google Scholar] [CrossRef]
- Alkam, T.; Nitta, A.; Mizoguchi, H.; Itoh, A.; Nabeshima, T. A natural scavenger of peroxynitrites, rosmarinic acid, protects against impairment of memory induced by Abeta(25-35). Behav. Brain Res. 2007, 180, 139–145. [Google Scholar] [CrossRef]
- Domitrović, R.; Skoda, M.; Vasiljev Marchesi, V.; Cvijanović, O.; Pernjak Pugel, E.; Stefan, M.B. Rosmarinic acid ameliorates acute liver damage and fibrogenesis in carbon tetrachloride-intoxicated mice. Food Chem. Toxicol. 2013, 51, 370–378. [Google Scholar] [CrossRef] [PubMed]
- Cameron, D.F.; Murray, F.T.; Drylie, D.D. Interstitial compartment pathology and spermatogenic disruption in testes from impotent diabetic men. Anat. Rec. 1985, 213, 53–62. [Google Scholar] [CrossRef]
- Sisman, A.R.; Kiray, M.; Camsari, U.M.; Evren, M.; Ates, M.; Baykara, B.; Aksu, I.; Guvendi, G.; Uysal, N. Potential novel biomarkers for diabetic testicular damage in streptozotocin-induced diabetic rats: Nerve growth factor Beta and vascular endothelial growth factor. Dis. Markers 2014, 2014, 108106. [Google Scholar] [CrossRef]
- Zha, W.; Bai, Y.; Xu, L.; Liu, Y.; Yang, Z.; Gao, H.; Li, J. Curcumin Attenuates Testicular Injury in Rats with Streptozotocin-Induced Diabetes. Biomed. Res. Int. 2018, 2018, 7468019. [Google Scholar] [CrossRef] [PubMed]
- Sevgi, K.; Tepe, B.; Sarikurkcu, C. Antioxidant and DNA damage protection potentials of selected phenolic acids. Food Chem. Toxicol. 2015, 77, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Khalaf, A.A.; Hassanen, E.I.; Ibrahim, M.A.; Tohamy, A.F.; Aboseada, M.A.; Hassan, H.M.; Zaki, A.R. Rosmarinic acid attenuates chromium-induced hepatic and renal oxidative damage and DNA damage in rats. J. Biochem. Mol. Toxicol. 2020, 34, e22579. [Google Scholar] [CrossRef]
- Rajesh, M.; Mukhopadhyay, P.; Bátkai, S.; Patel, V.; Saito, K.; Matsumoto, S.; Kashiwaya, Y.; Horváth, B.; Mukhopadhyay, B.; Becker, L.; et al. Cannabidiol attenuates cardiac dysfunction, oxidative stress, fibrosis, and inflammatory and cell death signaling pathways in diabetic cardiomyopathy. J. Am. Coll. Cardiol. 2010, 56, 2115–2125. [Google Scholar] [CrossRef]
- Althunibat, O.Y.; Al-Mustafa, A.H.; Tarawneh, K.; Khleifat, K.M.; Ridzwan, B.; Qaralleh, H.N. Protective role of Punica granatum L. peel extract against oxidative damage in experimental diabetic rats. Process Biochem. 2010, 45, 581–585. [Google Scholar] [CrossRef]
- Ikhlas, S.; Ahmad, M. Acute and sub-acute bisphenol-B exposures adversely affect sperm count and quality in adolescent male mice. Chemosphere 2020, 242, 125286. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-khawaldeh, O.; Al-Alami, Z.M.; Althunibat, O.Y.; Abuamara, T.M.M.; Mihdawi, A.; Abukhalil, M.H. Rosmarinic Acid Attenuates Testicular Damage via Modulating Oxidative Stress and Apoptosis in Streptozotocin-Induced Diabetic Albino Mice. Stresses 2024, 4, 505-517. https://doi.org/10.3390/stresses4030032
Al-khawaldeh O, Al-Alami ZM, Althunibat OY, Abuamara TMM, Mihdawi A, Abukhalil MH. Rosmarinic Acid Attenuates Testicular Damage via Modulating Oxidative Stress and Apoptosis in Streptozotocin-Induced Diabetic Albino Mice. Stresses. 2024; 4(3):505-517. https://doi.org/10.3390/stresses4030032
Chicago/Turabian StyleAl-khawaldeh, Omar, Zina M. Al-Alami, Osama Y. Althunibat, Tamer M. M. Abuamara, Afnan Mihdawi, and Mohammad H. Abukhalil. 2024. "Rosmarinic Acid Attenuates Testicular Damage via Modulating Oxidative Stress and Apoptosis in Streptozotocin-Induced Diabetic Albino Mice" Stresses 4, no. 3: 505-517. https://doi.org/10.3390/stresses4030032
APA StyleAl-khawaldeh, O., Al-Alami, Z. M., Althunibat, O. Y., Abuamara, T. M. M., Mihdawi, A., & Abukhalil, M. H. (2024). Rosmarinic Acid Attenuates Testicular Damage via Modulating Oxidative Stress and Apoptosis in Streptozotocin-Induced Diabetic Albino Mice. Stresses, 4(3), 505-517. https://doi.org/10.3390/stresses4030032