
Do Morphological Variations in Sclerotinia sclerotiorum Strains Indicate Differences in Aggressiveness?


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
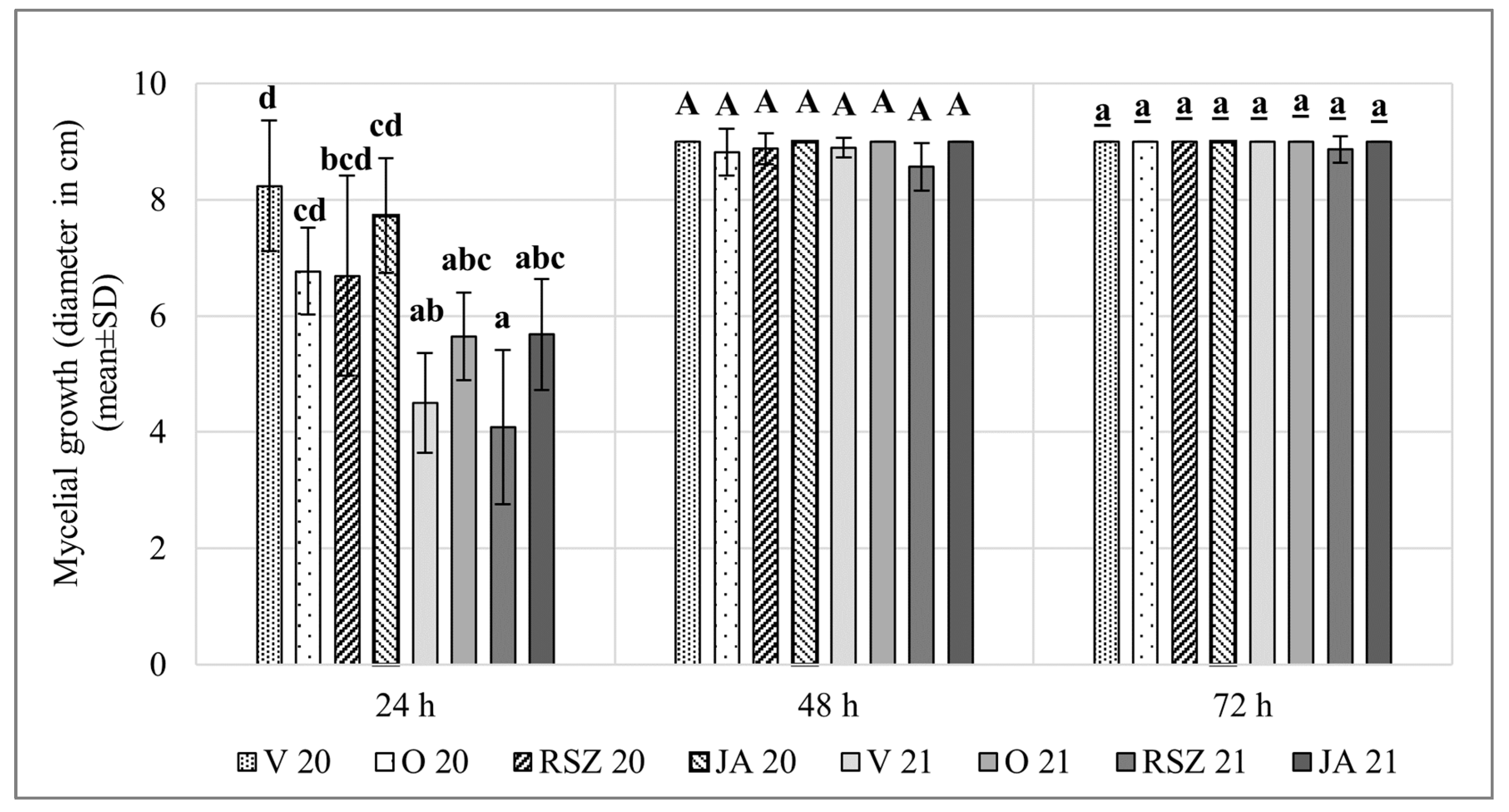
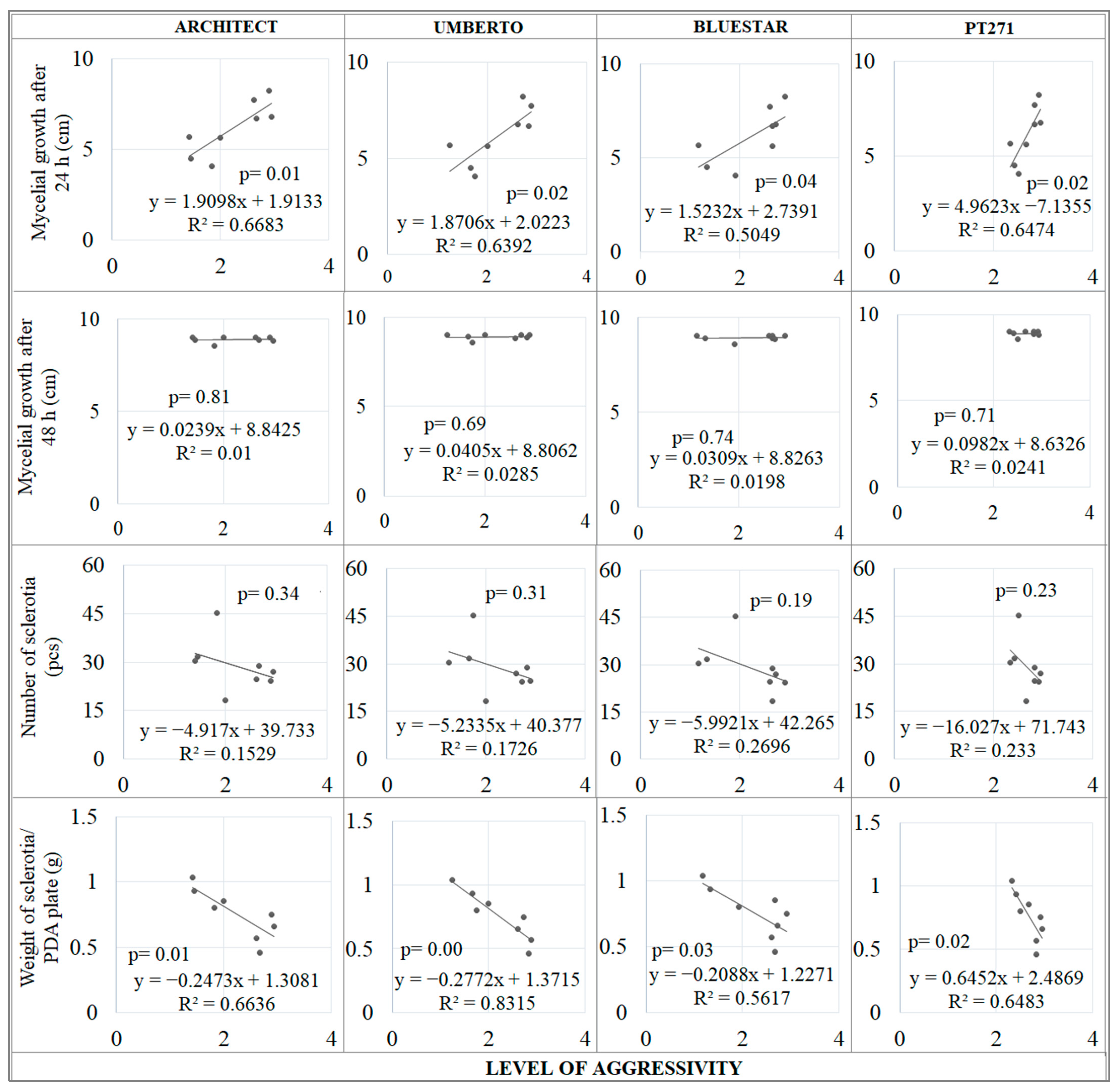
2.1. Mycelial Growth
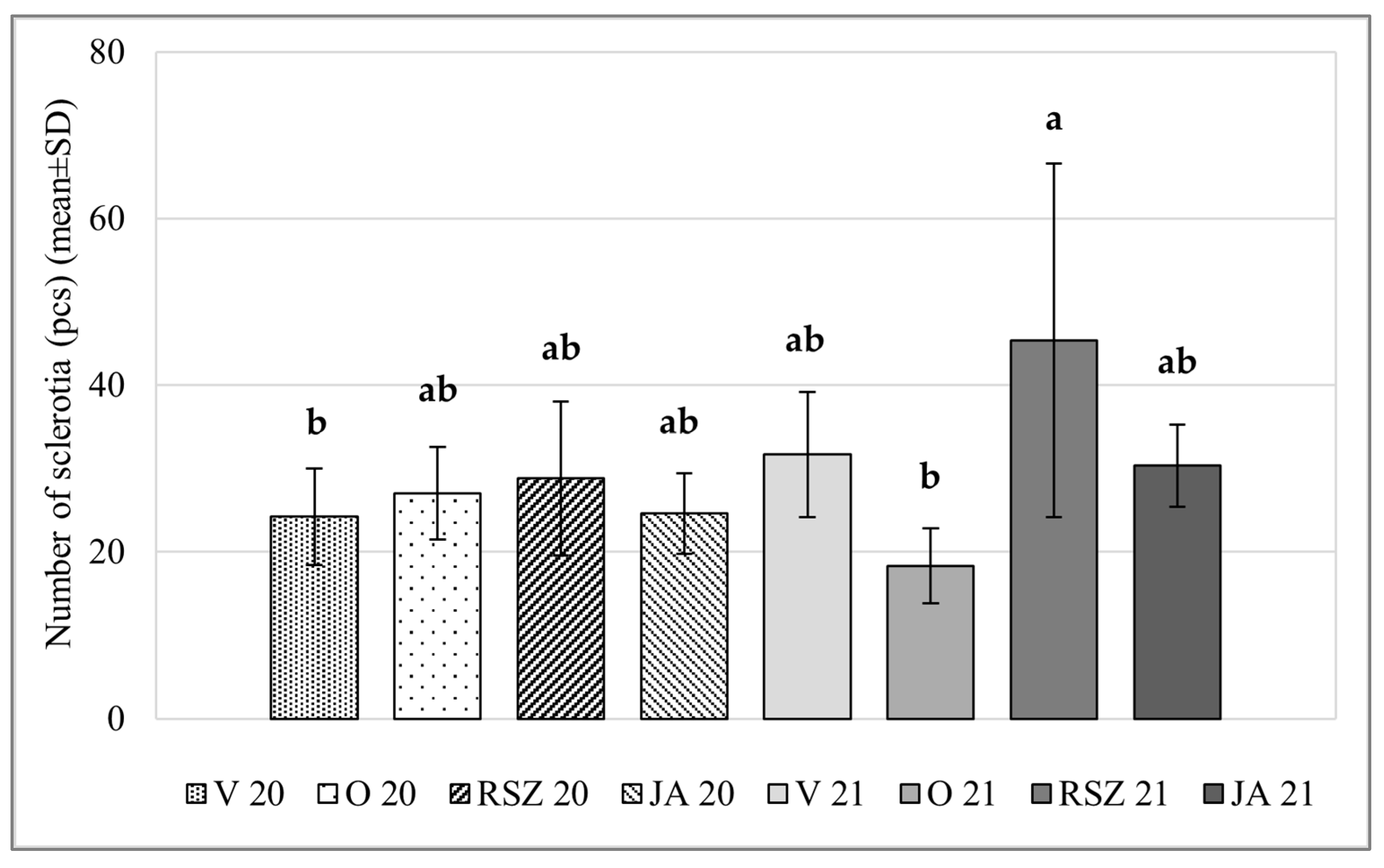
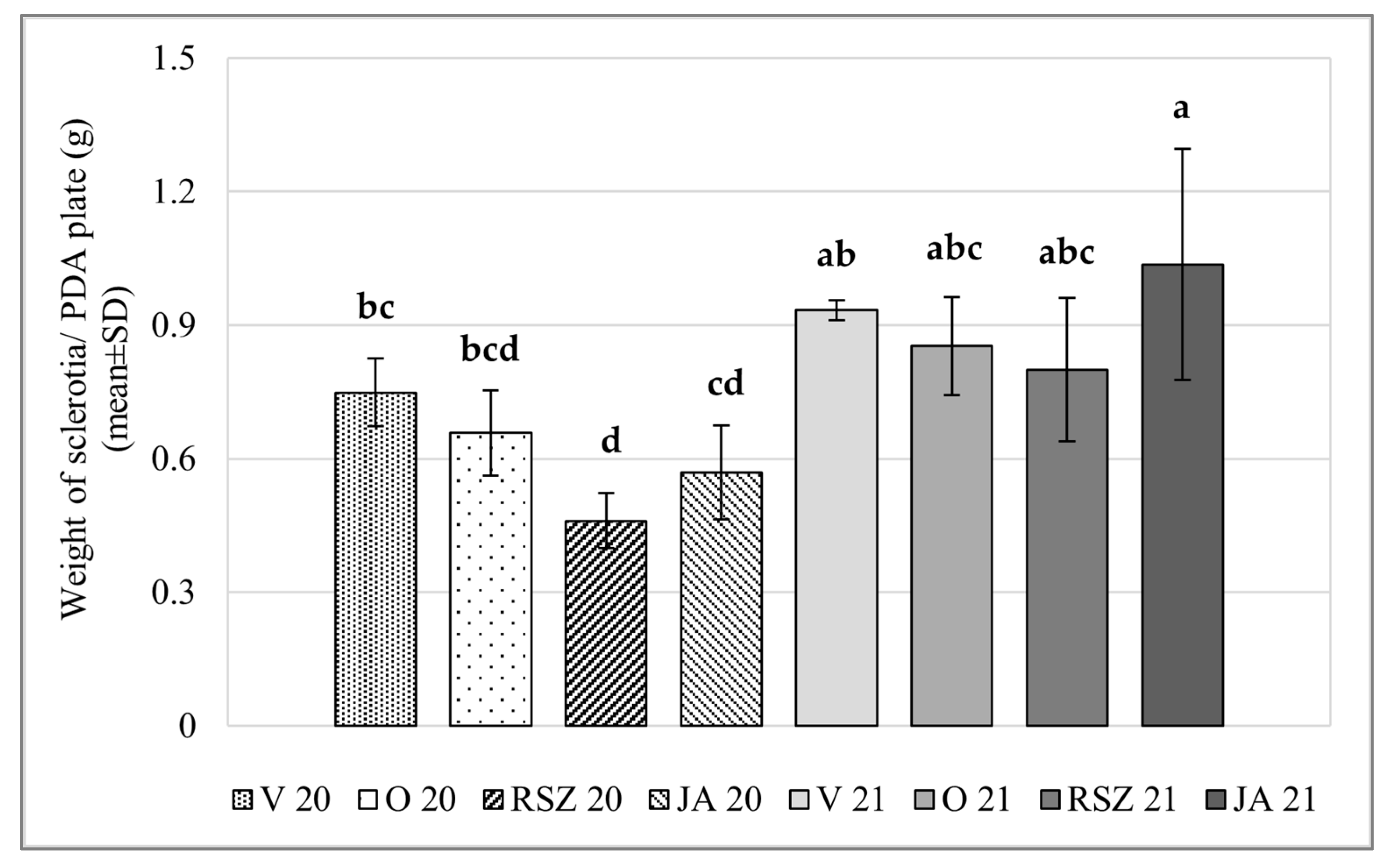
2.2. Sclerotial Production
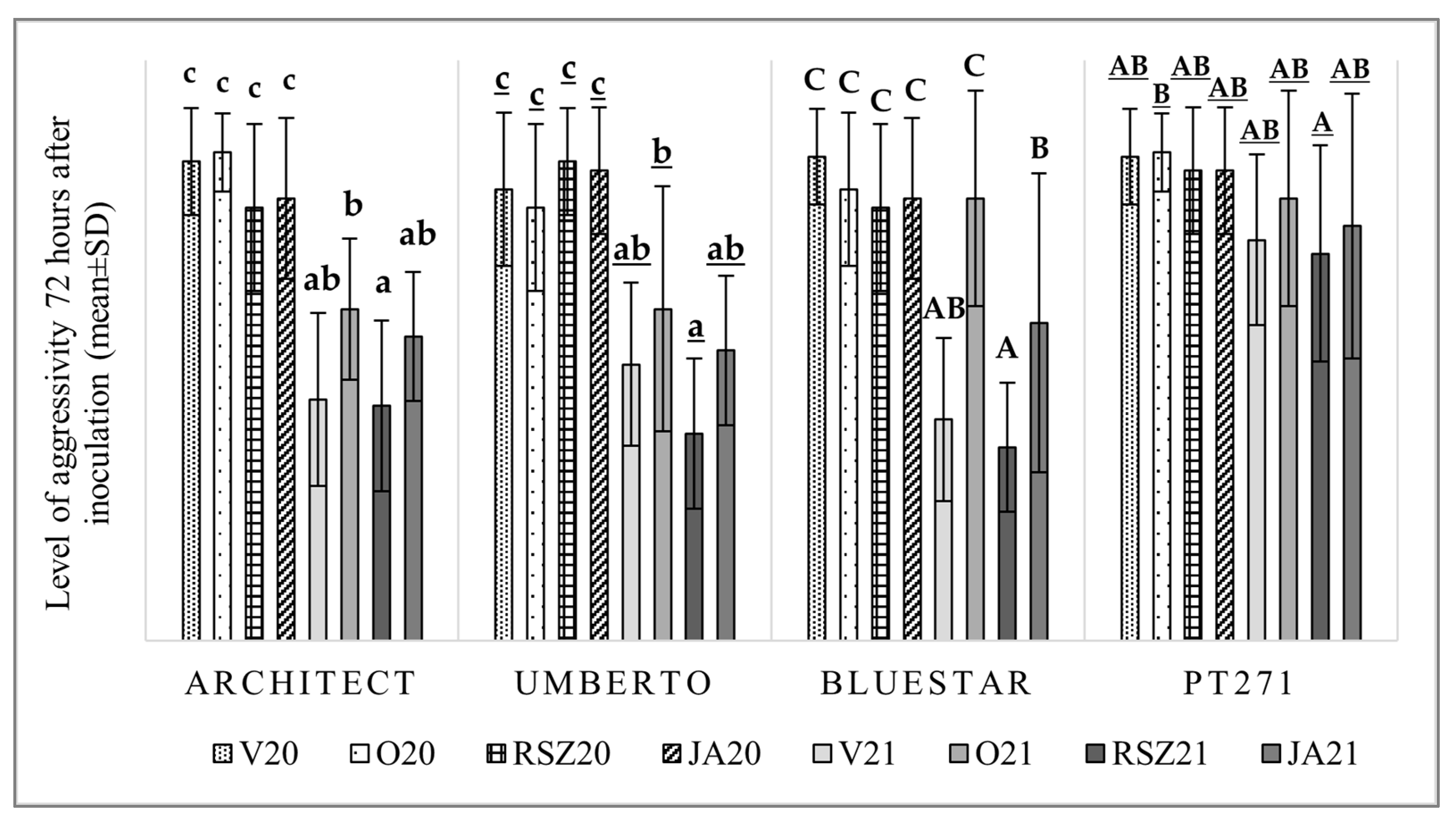
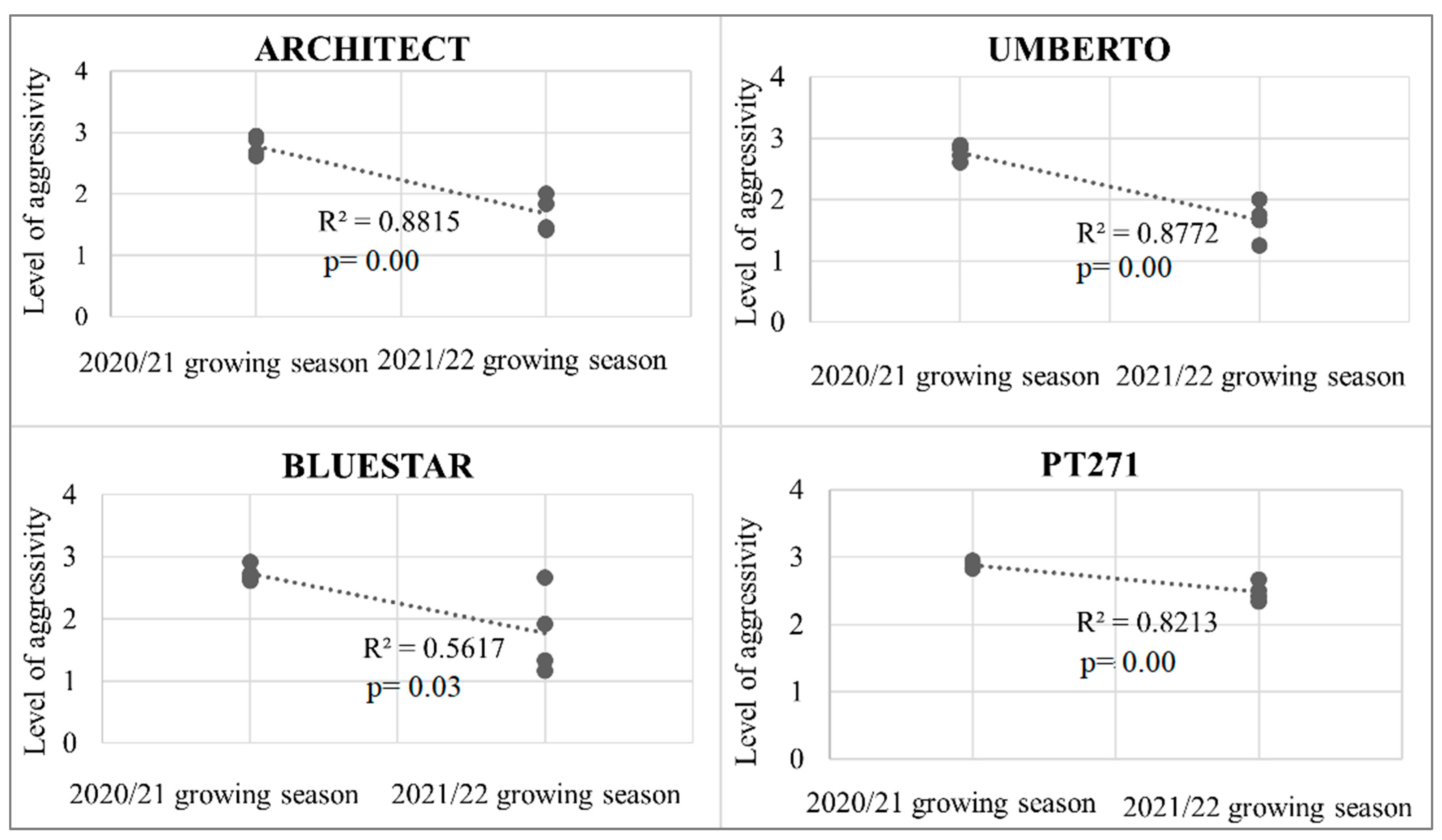
2.3. Aggressivity on OSR
2.4. Regression Analysis
3. Materials and Methods
3.1. Sample Collection
3.2. Subcultures of Isolate
3.3. In Vitro Experiment
3.4. Statistical and Regression Analyses
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Crops and Livestock Products. Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 3 May 2023).
- Oilseeds and Protein Crops Production in EU in 2011–2023. Available online: https://agridata.ec.europa.eu/extensions/DashboardCereals/OilseedProduction.html# (accessed on 3 May 2023).
- Zală, C.R.; Cristea, S.; Radu, E.; Manole, M.S.; Bălaşu, V.A.; Clinciu, E.C. Research regarding the response of rapeseed hybrids to pathogens. Sci. Pap. Ser. A Agron. 2012, LV, 265–268. [Google Scholar]
- Fitt, B.D.; Hu, B.C.; Li, Z.Q.; Liu, S.Y.; Lange, R.M.; Kharbanda, P.D.; Butterworth, M.H.; White, R.P. Strategies to prevent spread of Leptosphaeria maculans (phoma stem canker) onto oilseed rape crops in China; costs and benefits. Plant Pathol. 2008, 57, 652–664. [Google Scholar] [CrossRef]
- Sharma, P.; Meena, P.; Verma, P.; Saharan, G.; Mehta, N.; Singh, D.; Kumar, A. Sclerotinia sclerotiorum (lib) de Bary causing sclerotinia rot in oilseed Brassicas: A review. J. Oilseed Brassica 2016, 1, 1–44. [Google Scholar]
- Alizadeh, N.; Babai-Ahary, A.; Assadi, Y.; Valizadeh, M.; Passebaneslam, B. The effects of Sclerotinia stem rot of oilseed rape on the production and quality of extracted oil. Isfahan Univ. Technol. J. Crop Prod. Process. 2006, 10, 485–495. [Google Scholar]
- Boland, G.J.; Hall, R. Index of plant hosts of Sclerotinia sclerotiorum. Can. J. Plant Pathol. 1994, 16, 93–108. [Google Scholar] [CrossRef]
- Bolton, M.D.; Thomma, B.P.H.J.; Nelson, B.D. Sclerotinia sclerotiorum (Lib.) de Bary: Biology and molecular traits of a cosmopolitan pathogen. Mol. Plant Pathol. 2006, 7, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Managing Sclerotinia Stem Rot in Canola. Department of Agriculture and Food. Australian Government. Available online: https://www.agric.wa.gov.au/canola/managing-sclerotinia-stem-rot-canola (accessed on 3 May 2023).
- Erental, A.; Dickman, M.B.; Yarden, O. Sclerotial development in Sclerotinia sclerotiorum: Awakening molecular analysis of a “Dormant” structure. Fungal Biol. Rev. 2008, 22, 6–16. [Google Scholar] [CrossRef]
- Adams, P.B.; Ayers, W.A. Ecology of Sclerotinia species. Phytopathology 1979, 69, 896–899. [Google Scholar] [CrossRef]
- Taylor, A.; Coventry, E.; Handy, C.; West, J.S.; Young, C.S.; Clarkson, J.P. Inoculum potential of Sclerotinia sclerotiorum sclerotia depends on isolate and host plant. Plant Pathol. 2018, 67, 1286–1295. [Google Scholar] [CrossRef]
- Ben-Yephet, Y.; Genizi, A.; Siti, E. Sclerotial survival and apothecial production by Sclerotinia sclerotiorum following outbreaks of lettuce drop. Phytopathology 1993, 83, 509–513. [Google Scholar] [CrossRef]
- Harvey, I.C.; Foley, L.M.; Saville, D.J. Survival and germination of shallow-buried sclerotia of Sclerotinia sclerotiorum in pastures in Canterbury. N. Z. J. Agric. Res. 1995, 38, 279–284. [Google Scholar] [CrossRef]
- Abawi, G.S.; Grogan, R.G. Epidemiology of diseases caused by Sclerotinia species. Phytopathology 1979, 69, 899–904. [Google Scholar] [CrossRef]
- Lane, D.; Denton-Giles, M.; Derbyshire, M.; Kamphuis, L.G. Abiotic conditions governing the myceliogenic germination of Sclerotinia sclerotiorum allowing the basal infection of Brassica napus. Australas. Plant Pathol. 2019, 48, 85–91. [Google Scholar] [CrossRef]
- Hao, J.J.; Subbarao, K.V.; Duniway, J.M. Germination of Sclerotinia minor and S. sclerotiorum sclerotia under various soil moisture and temperature combinations. Phytopathology 2003, 93, 443–450. [Google Scholar] [CrossRef]
- Heran, A.; McCartney, H.A.; Li, Q. The effect of petal characteristics, inoculum density and environmental factors on infection of oilseed rape by Sclerotinia sclerotiorum. In Proceedings of the 10th International Rapeseed Congress 1999, Canberra, Australia, 26–29 September 1999; Wratten, N., Salisbury, P.A., Eds.; The Regional Institute Ltd.: Gosford, Australia. Available online: http://www.regional.org.au/au/gcirc (accessed on 4 June 2024).
- Purdy, L. Sclerotinia sclerotiorum: History, diseases and symptomatology, host range, geographic distribution, and impact. Phytopathology 1979, 69, 875–880. [Google Scholar] [CrossRef]
- Huang, H.C.; Kozub, G.C. Germination of immature and mature sclerotia of Sclerotinia sclerotiorum. Bot. Bull. Acad. Sin. 1994, 35, 243–247. [Google Scholar]
- Adams, P.B.; Ayers, W.A. Histological and physiological aspects of infection of sclerotia of two Sclerotinia species by two mycoparasites. Phytopathology 1983, 73, 1072–1076. [Google Scholar] [CrossRef]
- Huang, H.C. Induction of myceliogenic germination of sclerotia of Sclerotinia sclerotiorum by exposure to sub-freezing temperatures. Plant Pathol. 1991, 40, 621–625. [Google Scholar] [CrossRef]
- Matheron, M.E.; Porchas, M. Influence of soil temperature and moisture on eruptive germination and viability of sclerotia of Sclerotinia minor and S. sclerotiorum. Plant Dis. 2005, 89, 50–54. [Google Scholar]
- Fischl, G. Fungal diseases of sunflowers. In Diseases of Field Crops; Horváth, J., Ed.; Mezőgaza Kiadó Press: Budapest, Hungary, 1995; pp. 110–124. (In Hungarian) [Google Scholar]
- The Growth Stages of Oilseed Rape. Available online: https://ahdb.org.uk/knowledge-library/the-growth-stages-of-oilseed-rape (accessed on 16 January 2024).
- Hegedus, D.D.; Rimmer, S.R. Sclerotinia sclerotiorum: When “to be or not to be” a pathogen? FEMS Microbiol. Lett. 2005, 251, 177–184. [Google Scholar] [CrossRef]
- Mathew, F.; Harveson, R.; Block, C.; Gulya, T.; Ryley, M.; Thompson, S.; Markell, S. Symptoms and signs. Phytopathol. News 2022. [Google Scholar] [CrossRef]
- Riou, C.; Freyssinet, G.; Fevre, M. Production of cell wall-degrading enzymes by the phytopathogenic fungus Sclerotinia sclerotiorum. Appl. Environ. Microbiol. 1991, 57, 1478–1484. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, M.; Wang, Y.; Li, R.; Fernando, W.D. Mycelial compatibility group and pathogenicity variation of Sclerotinia sclerotiorum populations in sunflower from China, Canada and England. Plant Pathol. J. 2008, 7, 131–139. [Google Scholar] [CrossRef]
- Barzman, M.; Barberi, P.; Birch, N.; Boonekamp, P.; Dachbrodt-Saaydeh, S.; Graf, B.; Hommel, B.; Jensen, J.E.; Kiss, J.; Kudsk, P.; et al. Eight principles of Integrated Pest Management. Agronomy for sustainable development. 2015, 35, 1199–1215. [Google Scholar] [CrossRef]
- Effect of Cultural Control on Rapeseed Stem Rot (Sclerotinia sclerotiorum) in Brassica napus. The Regional Institute Online Publishing. Available online: http://www.regional.org.au/au/gcirc/3/520.htm (accessed on 3 May 2023).
- Masirevic, S.; Gulya, T.J. Sclerotinia and Phomopsis—Two devastating sunflower pathogens. Field Crops Res. 1992, 30, 271–300. [Google Scholar] [CrossRef]
- Szlávik, S.z.; Gergely, L.; Zalka, A.; Orlóciné Debreceni, Á.; Bartus, E. Evaluation of the Hungarian State registered winter rape cultivars for susceptibility to Sclerotinia sclerotiorum. In Proceedings of the 9. Tiszántúli Növényvédelmi Fórum, Debrecen, Hungary, 20–21 October 2004; pp. 130–133. (In Hungarian). [Google Scholar]
- Abro, M.A.; Syed, R.N.; Yasmin, A.; Abro, S.; Kumar, A.; Nicot, P.C. Different nitrogen fertilization regimes influences the susceptibility of lettuce plants to Botrytis cinerea and Sclerotinia sclerotiorum. Sci. Int. 2014, 26, 325–329. [Google Scholar]
- Ni, L.; Punja, Z.K. Effects of a foliar fertilizer containing boron on the development of Sclerotinia stem rot (Sclerotinia sclerotiorum) on canola (Brassica napus L.) leaves. J. Phytopathol. 2020, 168, 47–55. [Google Scholar] [CrossRef]
- Verkley, G.J.M.; Dukik, K.; Renfurm, R.; Göker, M.; Stielow, J.B. Novel genera and species of coniothyrium-like fungi in Montagnulaceae (Ascomycota). Persoonia 2014, 32, 25–51. [Google Scholar] [CrossRef] [PubMed]
- Turóczi, G.y. Biological control against plant pathogens. In Biological Plant Protection and Its Role in Hungary; Darvas, B., Ed.; Tudományos Akadémia Press: Budapest, Hungary, 2008; pp. 100–151. (In Hungarian) [Google Scholar]
- Pálinkás, Z.; Perczel, M.; Szénási, Á.; Dorner, Z.; Kiss, J.; Bán, R. Integrated protection of sunflowers. Növényvédelem 2018, 79, 483–504. (In Hungarian) [Google Scholar]
- Tu, J. Mycoparasitism by Coniothyrium minitans on Sclerotinia sclerotiorum and its Effect on Sclerotial Germination. J. Phtyopathology 1984, 109, 261–268. [Google Scholar] [CrossRef]
- Agrios, G.N. Plant Pathology; Elsevier Academic Press: San Diego, CA, USA, 2005. [Google Scholar]
- Vleugels, T.; Baert, J.; Van Bockstaele, E. Morphological and pathogenic characterization of genetically diverse Sclerotinia isolates from European red clover crops (Trifolium pratense L.). J. Phytopathol. 2013, 161, 254–262. [Google Scholar] [CrossRef]
- Irani, H.; Javan-Nikkhah, M.; İbrahimov, A.Ş.; Heydari, A. Diversity of aggressiveness of Sclerotinia sclerotiorum (Lib.) de Bary populations in oil plants fields of north and northeast of Iran. Iran. J. Plant Pathol. 2015, 51, 1–10. [Google Scholar]
- Kohli, Y.; Morrall, R.A.A.; Anderson, J.B.; Kohn, L.M. Local and trans-Canadian clonal distribution of Sclerotinia sclerotiorum on canola. Phytopathology 1992, 82, 875–880. [Google Scholar] [CrossRef]
- Template Europe Map. Available online: https://enfo.hu/index.php/node/12175 (accessed on 9 May 2024).
- Dhingra, O.D.; Sinclair, J.B. Basic Plant Pathology Methods; CRC Press Inc.: Boca Raton, FL, USA, 1985; p. 355. [Google Scholar]
- Clarkson, J.P.; Staveley, J.; Phelps, K.; Young, C.S.; Whipps, J.M. Ascospore release and survival in Sclerotinia sclerotiorum. Mycol. Res. 2003, 107, 213–222. [Google Scholar] [CrossRef]
- Rathi, A.S.; Jattan, M.; Punia, R.; Singh, S.; Kumar, P.; Avtar, R. Morphological and molecular diversity of Sclerotinia sclerotiorum infecting Indian mustard. Indian. Phytopathol. 2018, 71, 407–413. [Google Scholar] [CrossRef]
- Kull, L.S.; Vuong, T.D.; Powers, K.S.; Eskridge, K.M.; Steadman, J.R.; Hartman, G.L. Evaluation of resistance screening methods for Sclerotinia stem rot of soybean and dry bean. Plant Dis. 2003, 87, 1471–1476. [Google Scholar] [CrossRef]
- Garg, H.; Kohn, L.M.; Andrew, M.; Li, H.; Sivasithamparam, K.; Barbetti, M.J. Pathogenicity of morphologically different isolates of Sclerotinia sclerotiorum with Brassica napus and B. juncea genotypes. European J. Plant Pathol. 2010, 126, 305–315. [Google Scholar] [CrossRef]
- Reza Ahmadi, M.; Javan-Nik-Khah, M.; Aghajani, M.A.; Ghobakhloo, M. Morphological variability among Sclerotinia sclerotiorum populations associated with stem rot of important crops and weeds. World Appl. Sci. J. 2012, 20, 1561–1564. [Google Scholar] [CrossRef]
- Ghasolia, R.P.; Shivpuri, A. Morphological and pathogenic variability in rapeseed and mustard isolates of Sclerotinia sclerotiorum. Indian Phytopathol. 2011, 60, 76–81. [Google Scholar]
- Pawan, K.; Rathi, A.S.; Singh, J.K.; Berwal, M.K.; Mukesh, K.; Anil, K.; Dhiraj, S. Cultural, morphological and pathogenic diversity analysis of Sclerotinia sclerotiorum causing Sclerotinia rot in Indian mustard. Indian J. Ecol. 2016, 43, 463–472. [Google Scholar]
- Morrall, R.A.A.; Duczek, L.J.; Sheard, J.W. Variations and correlations within and between morphology, pathogenicity, and pectolytic enzyme activity in Sclerotinia from Saskatchewan. Can. J. Bot. 1972, 50, 767–786. [Google Scholar] [CrossRef]
- Kyryk, M.; Pikovskyi, M.; Kolesnichenko, O.; Borodai, V.; Markovska, O.; Dudchenko, V.; Solomiyciuk, M. Sclerotic productivity, mycelial compatibility, and pathogenicity of the isolates of the fungus Sclerotinia sclerotiorum (Lib.) de Bary. Ukr. J. Ecol. 2021, 11, 21–27. [Google Scholar] [CrossRef]
- Ge, X.T.; You, M.P.; Barbetti, M.J. Virulence differences among Sclerotinia sclerotiorum isolates determines host cotyledon resistance responses in Brassicaceae genotypes. Eur. J. Plant Pathol. 2015, 143, 527–541. [Google Scholar] [CrossRef]
- Denton-Giles, M.; Derbyshire, M.C.; Khentry, Y.; Buchwaldt, L.; Kamphuis, L.G. Partial stem resistance in Brassica napus to highly aggressive and genetically diverse Sclerotinia sclerotiorum isolates from Australia. Can. J. Plant Pathol. 2018, 40, 551–561. [Google Scholar] [CrossRef]
- Pratt, R.G.; Rowe, D.E. Comparative pathogenicity of isolates of Sclerotinia trifoliorum and S. sclerotiorum on alfalfa cultivars. Plant Dis. 1995, 79, 474–477. [Google Scholar] [CrossRef]
- Alavi, N.; Dalili, A. Evaluation of resistance in rapeseed lines against Sclerotinia rot (Sclerotinia sclerotiorum). J. Oilseed Brassica 2016, 1, 32–39. [Google Scholar]
- Taylor, A.; Coventry, E.; Jones, J.E.; Clarkson, J.P. Resistance to a highly aggressive isolate of Sclerotinia sclerotiorum in a Brassica napus diversity set. Plant Pathol. 2015, 64, 932–940. [Google Scholar] [CrossRef]
- Otto-Hanson, L.; Steadman, J.R.; Higgins, R.; Eskridge, K.M. Variation in Sclerotinia sclerotiorum bean isolates from multisite resistance screening locations. Plant Dis. 2011, 95, 1370–1377. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vizi, R.; Kiss, J.; Turóczi, G.; Dobra, N.; Pálinkás, Z. Do Morphological Variations in Sclerotinia sclerotiorum Strains Indicate Differences in Aggressiveness? Stresses 2024, 4, 367-379. https://doi.org/10.3390/stresses4020024
Vizi R, Kiss J, Turóczi G, Dobra N, Pálinkás Z. Do Morphological Variations in Sclerotinia sclerotiorum Strains Indicate Differences in Aggressiveness? Stresses. 2024; 4(2):367-379. https://doi.org/10.3390/stresses4020024
Chicago/Turabian StyleVizi, Ramóna, József Kiss, György Turóczi, Nóra Dobra, and Zoltán Pálinkás. 2024. "Do Morphological Variations in Sclerotinia sclerotiorum Strains Indicate Differences in Aggressiveness?" Stresses 4, no. 2: 367-379. https://doi.org/10.3390/stresses4020024
APA StyleVizi, R., Kiss, J., Turóczi, G., Dobra, N., & Pálinkás, Z. (2024). Do Morphological Variations in Sclerotinia sclerotiorum Strains Indicate Differences in Aggressiveness? Stresses, 4(2), 367-379. https://doi.org/10.3390/stresses4020024