

Insights into Key Biometric, Physiological and Biochemical Markers of Magnesium (Mg) Deficiency Stress in the Halophyte Cakile maritima

Abstract
1. Introduction
2. Results
2.1. Biometric and Physiological Markers of Mg Deficiency Stress in C. maritima
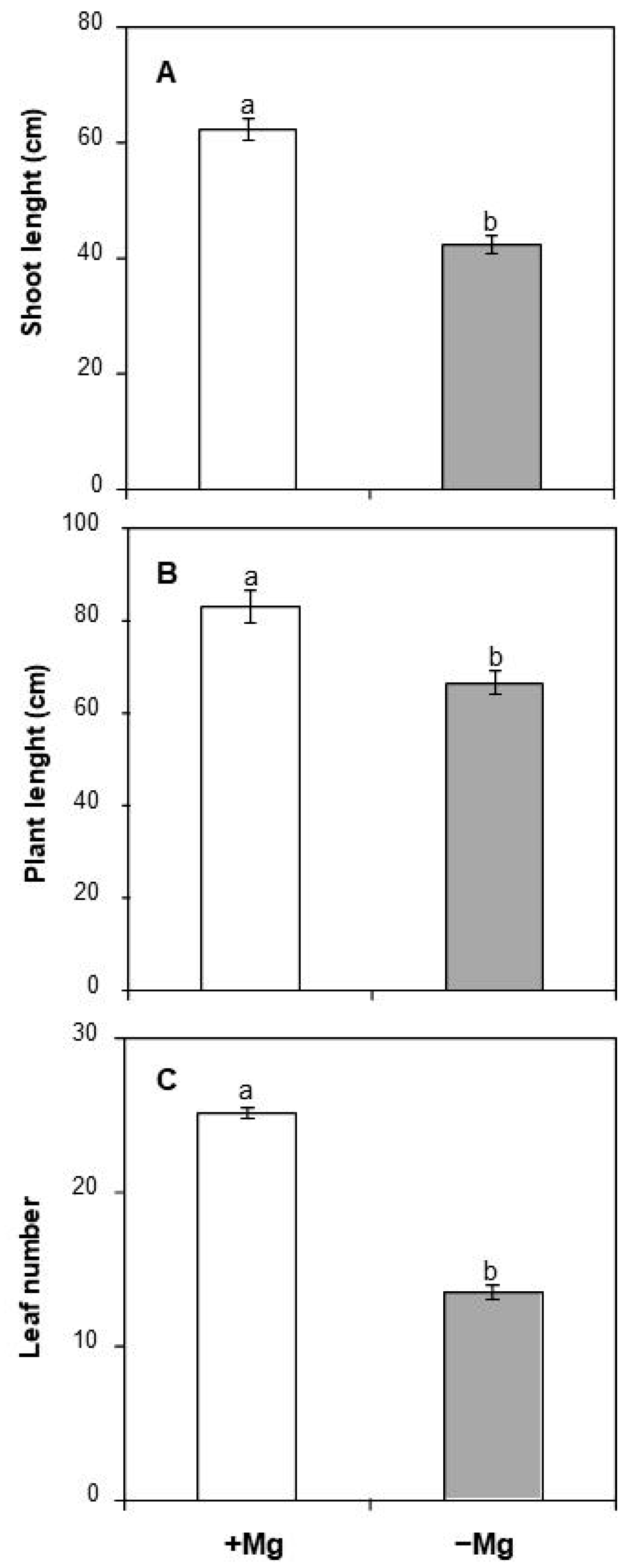
2.1.1. Shoot Length, Plant Length and Total Leaf Number
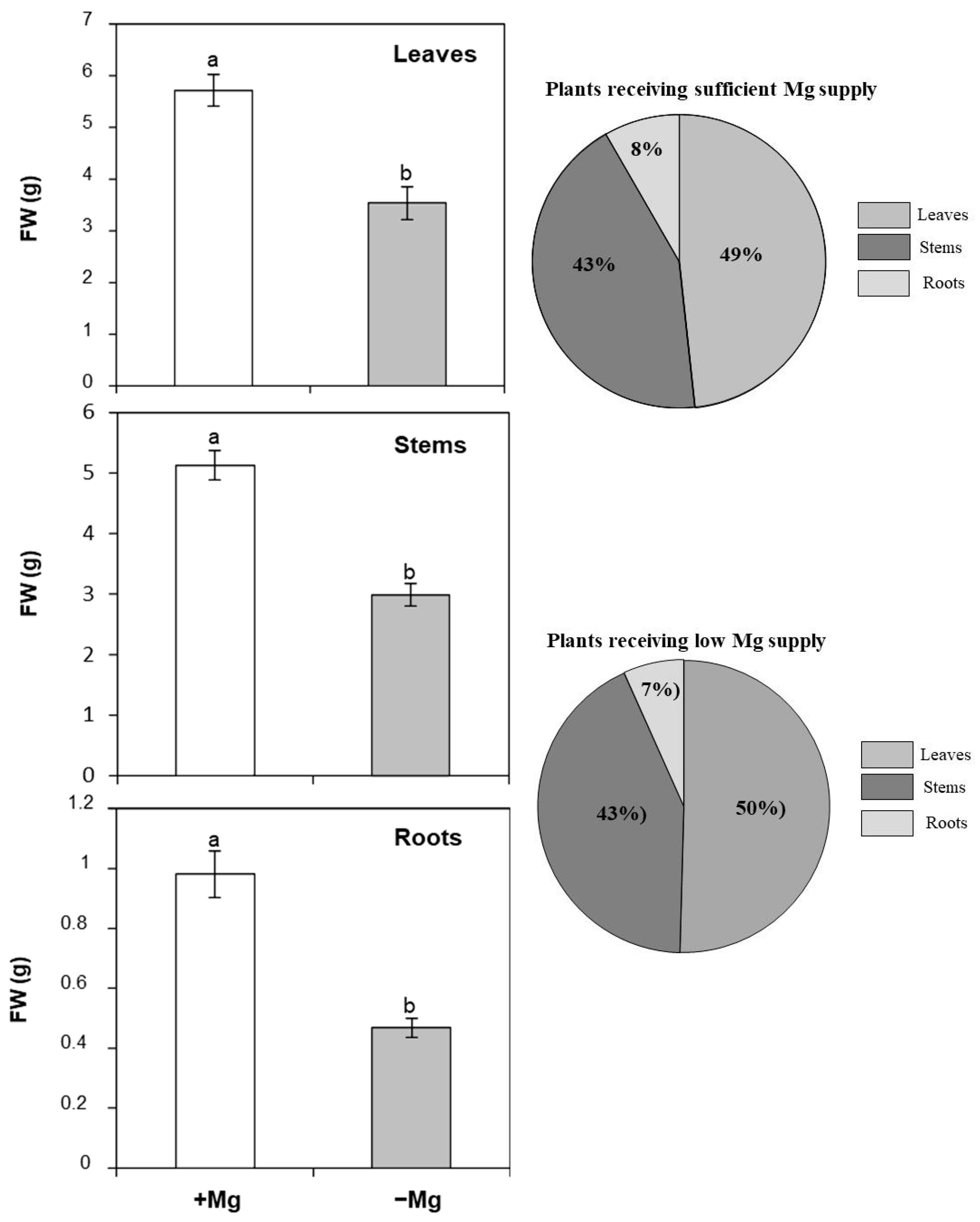
2.1.2. Leaf, Stem, and Root Biomass Accumulation
2.1.3. Tolerance Stress Indices
2.2. Effect of Mg Deficiency on Chlorophyll Content
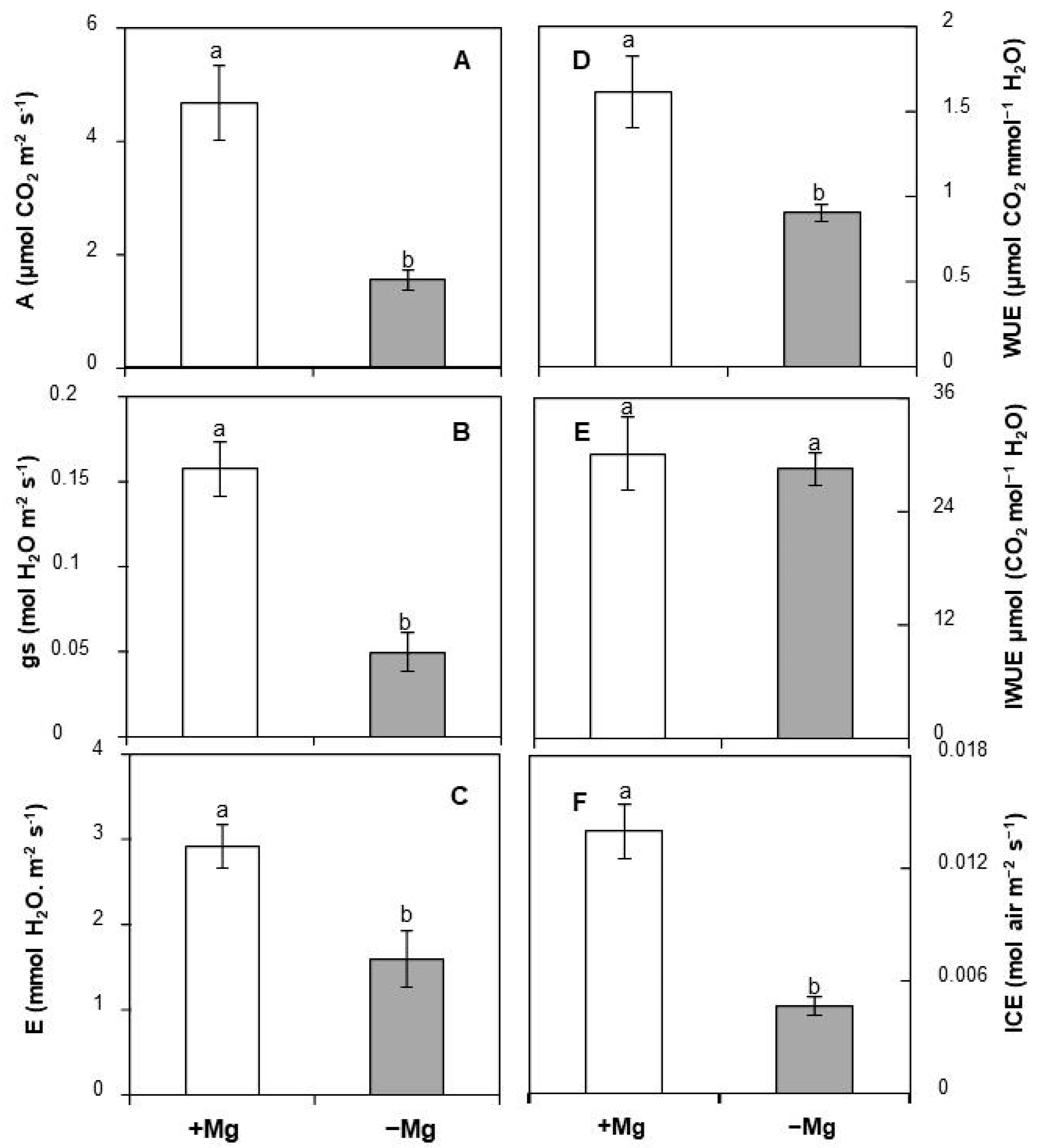
2.3. Effect of Mg Deficiency on Leaf Gas Exchange
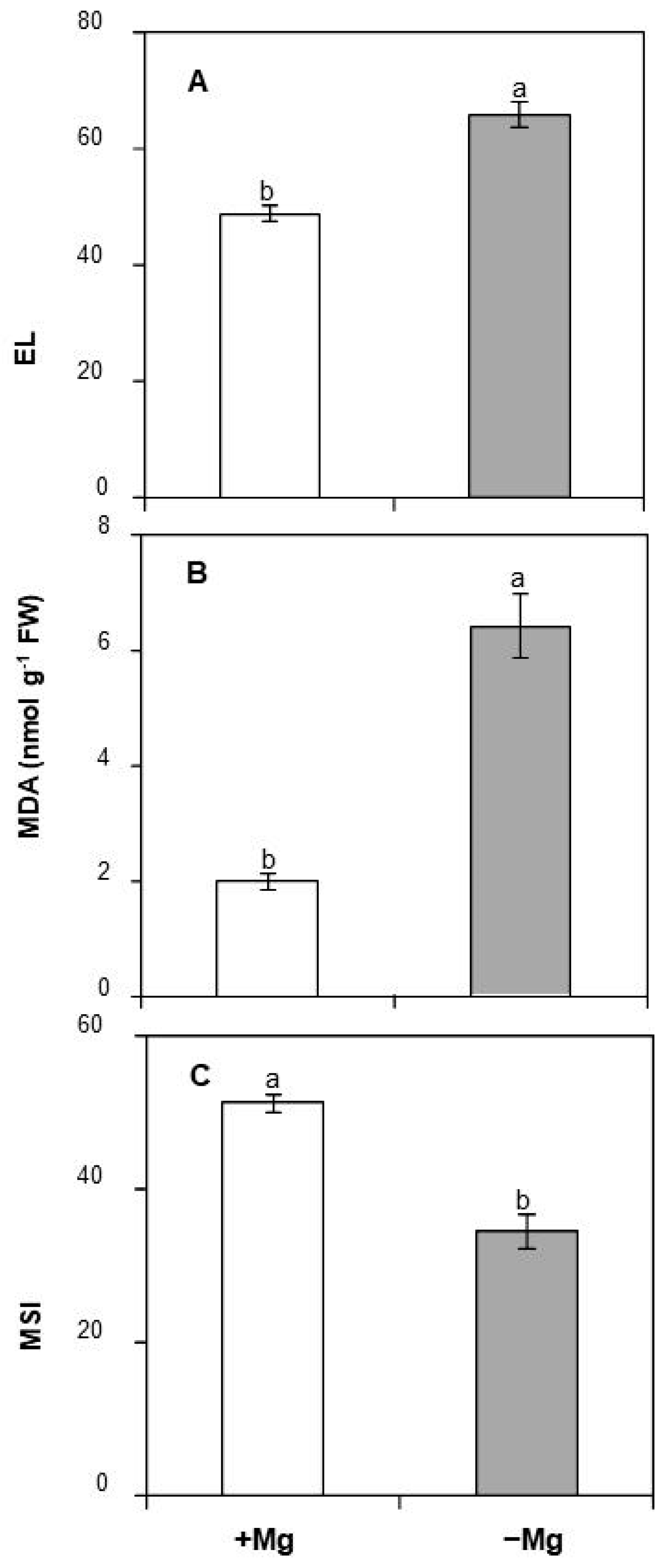
2.4. Effect of Mg Deficiency on Some Indicators of Oxidative Damage
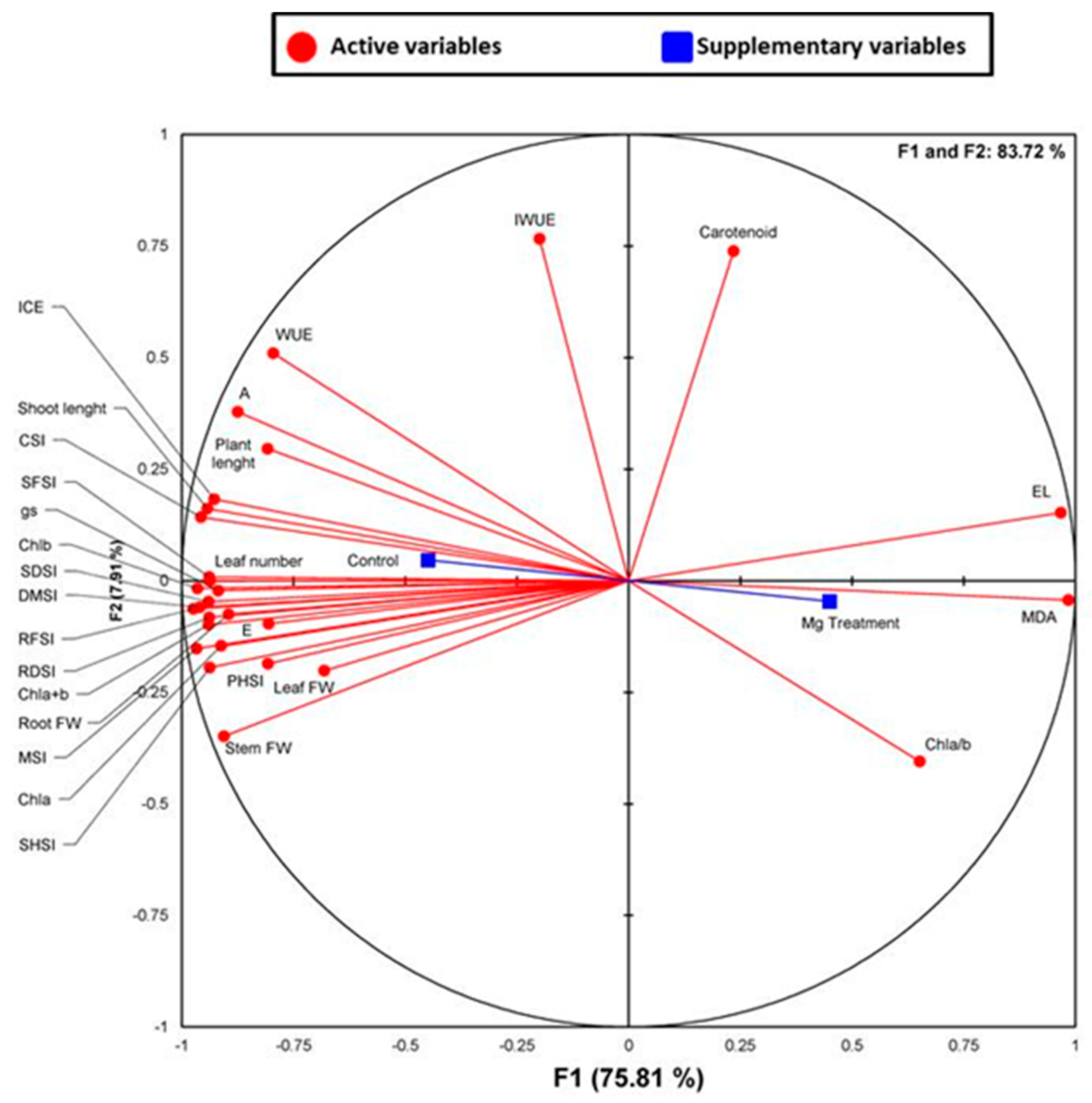
2.5. Correlation Analysis and Principal Component Analysis (PCA)
3. Discussion
4. Materials and Methods
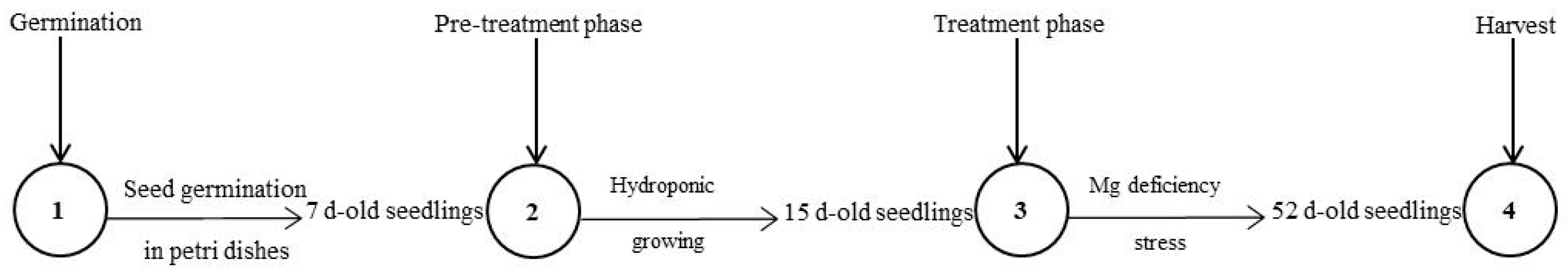
4.1. Plant Material and Growth Conditions
4.2. Physiological Stress Markers
4.2.1. Photosynthetic Pigment Analysis
4.2.2. Chlorophyll Stability Index (CSI)
4.2.3. Gas Exchange Analysis
4.2.4. Water Use Efficiency (WUE), Intrinsic Water Use Efficiency (IWUE), and Instantaneous Carboxylation Efficiency (ICE)
4.3. Biochemical Stress Markers
4.3.1. Relative Electrolyte Leakage (EL)
4.3.2. Membrane Stability Index (MSI)
4.3.3. Lipid Peroxidation
4.4. Physiological Stress Tolerance Indices
4.5. Statistical Analysis
4.6. Principal Component Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chaudhry, A.H.; Nayab, S.; Hussain, S.B.; Ali, M.; Pan, Z. Current understandings on magnesium deficiency and future outlooks for sustainable agriculture. Int. J. Mol. Sci. 2021, 22, 1819. [Google Scholar] [CrossRef] [PubMed]
- Cakmak, I.; Yazici, A.M. Magnesium: A forgotten element in crop production. Better Crops 2010, 94, 23–25. [Google Scholar]
- Guo, W. Magnesium homeostasis mechanisms and magnesium use efficiency in plants. In Plant Macronutrient Use Efficiency; Academic Press: Cambridge, MA, USA, 2017; pp. 197–213. [Google Scholar]
- Baloch, F.S.; Nadeem, M.A.; Sönmez, F.; Habyarimana, E.; Mustafa, Z.; Karaköy, T.; Cömertpay, G.; Alsaleh, A.; Çiftçi, V.; Sun, S.; et al. Magnesium-a forgotten element: Phenotypic variation and genome wide association study in Turkish Common Bean Germplasm. Front. Genet. 2022, 13, 848663. [Google Scholar] [CrossRef] [PubMed]
- Tränkner, M.; Tavakol, E.; Jákli, B. Functioning of potassium and magnesium in photosynthesis, photosynthate translocation and photoprotection. Physiol. Plant. 2018, 163, 414–431. [Google Scholar] [CrossRef] [PubMed]
- Gransee, A.; Führs, H. Magnesium mobility in soils as a challenge for soil and plant analysis, magnesium fertilization and root uptake under adverse growth conditions. Plant Soil 2013, 368, 5–21. [Google Scholar] [CrossRef]
- Niu, Y.; Jin, G.; Zhang, Y.S. Root development under control of magnesium availability. Plant Signal Behav. 2014, 9, e29720. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Ackah, M.; Wang, L.; Amoako, F.K.; Shi, Y.; Essoh, L.G.; Li, J.; Zhang, Q.; Li, H.; Zhao, W. Magnesium Nutrient Application Induces Metabolomics and Physiological Responses in Mulberry (Morus alba). Int. J. Mol. Sci. 2023, 24, 9650. [Google Scholar] [CrossRef] [PubMed]
- Billard, V.; Maillard, A.; Coquet, L.; Jouenne, T.; Cruz, F.; Garcia-Mina, J.M.; Yvin, J.C.; Ourry, A.; Etienne, P. Mg deficiency affects leaf Mg remobilization and the proteome in Brassica napus. Plant Physiol. Biochem. 2016, 107, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Ishfaq, M.; Wang, Y.; Yan, M.; Wang, Z.; Wu, L.; Li, C.; Li, X. Physiological essence of magnesium in plants and its widespread deficiency in the farming system of China. Front. Plant Sci. 2022, 13, 802274. [Google Scholar] [CrossRef]
- Zhang, B.; Cakmak, I.; Feng, J.; Yu, C.; Chen, X.; Xie, D.; Wu, L.; Song, Z.; Cao, J.; He, Y. Magnesium deficiency reduced the yield and seed germination in wax gourd by affecting the carbohydrate translocation. Front. Plant Sci. 2020, 11, 797. [Google Scholar] [CrossRef]
- Omar, M.M.; Abdrabou, H.A.; Elghamry, A.M. Response of lettuce plant grown on sandy soil to organic and inorganic amendments. J. Soil Sci. Agric. Eng. 2022, 13, 33–38. [Google Scholar] [CrossRef]
- Qu, S.; Li, H.; Zhang, X.; Gao, J.; Ma, R.; Ma, L.; Ma, J. Effects of Magnesium Imbalance on Root Growth and Nutrient Absorption in Different Genotypes of Vegetable Crops. Plants 2023, 12, 3518. [Google Scholar] [CrossRef]
- Peng, Y.Y.; Liao, L.L.; Liu, S.; Nie, M.M.; Li, J.; Zhang, L.D.; Ma, J.F.; Chen, Z.C. Magnesium deficiency triggers SGR–mediated chlorophyll degradation for magnesium remobilization. Plant Physiol. 2019, 181, 262–275. [Google Scholar] [CrossRef] [PubMed]
- Ye, X.; Chen, X.F.; Deng, C.L.; Yang, L.T.; Lai, N.W.; Guo, J.X.; Chen, L.S. Magnesium-deficiency effects on pigments, photosynthesis and photosynthetic electron transport of leaves, and nutrients of leaf blades and veins in Citrus sinensis seedlings. Plants 2019, 8, 389. [Google Scholar] [CrossRef] [PubMed]
- Mao, D.; Chen, J.; Tian, L.; Liu, Z.; Yang, L.; Tang, R.; Li, J.; Lu, C.; Yang, Y.; Shi, J.; et al. Arabidopsis transporter MGT6 mediates magnesium uptake and is required for growth under magnesium limitation. Plant Cell 2014, 26, 2234–2248. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.W.; Mao, D.D.; Yang, L.; Qi, J.L.; Zhang, X.X.; Tang, Q.L.; Li, Y.P.; Tang, R.J.; Luan, S. Magnesium transporter MGT6 plays an essential role in maintaining magnesium homeostasis and regulating high magnesium tolerance in Arabidopsis. Front. Plant Sci. 2018, 9, 274. [Google Scholar] [CrossRef]
- Ogura, T.; Kobayashi, N.I.; Hermans, C.; Ichihashi, Y.; Shibata, A.; Shirasu, K.; Aoki, N.; Sugita, R.; Ogawa, T.; Suzuki, H.; et al. Short-term magnesium deficiency triggers nutrient retranslocation in Arabidopsis thaliana. Front. Plant Sci. 2020, 11, 563. [Google Scholar] [CrossRef]
- Wang, Z.; Hassan, M.U.; Nadeem, F.; Wu, L.; Zhang, F.; Li, X. Magnesium fertilization improves crop yield in most production systems: A meta-analysis. Front. Plant Sci. 2020, 10, 495191. [Google Scholar] [CrossRef]
- Farhat, N.; Ivanov, A.G.; Krol, M.; Rabhi, M.; Smaoui, A.; Abdelly, C.; Hüner, N.P. Preferential damaging effects of limited magnesium bioavailability on photosystem I in Sulla carnosa plants. Planta 2015, 241, 1189–1206. [Google Scholar] [CrossRef]
- Chen, Z.C.; Peng, W.T.; Li, J.; Liao, H. Functional dissection and transport mechanism of magnesium in plants. Semin. Cell Dev. Biol. 2018, 74, 142–152. [Google Scholar] [CrossRef]
- Meng, X.; Bai, S.; Wang, S.; Pan, Y.; Chen, K.; Xie, K.; Wang, M.; Guo, S. The sensitivity of photosynthesis to magnesium deficiency differs between rice (Oryza sativa L.) and cucumber (Cucumis sativus L.). Front. Plant Sci. 2023, 14, 1164866. [Google Scholar] [CrossRef] [PubMed]
- Czarnocka, W.; Karpiński, S. Friend or foe? Reactive oxygen species production, scavenging and signaling in plant response to environmental stresses. Free Radic. Biol. Med. 2018, 122, 4–20. [Google Scholar] [CrossRef] [PubMed]
- Farhat, N.; Rabhi, M.; Krol, M.; Barhoumi, Z.; Ivanov, A.G.; McCarthy, A.; Abdelly, C.; Smaoui, A.; Hüner, N.P. Starch and sugar accumulation in Sulla carnosa leaves upon Mg2+ starvation. Acta Physiol. Plant. 2014, 36, 2157–2165. [Google Scholar] [CrossRef]
- Debez, A.; Rejeb, K.B.; Ghars, M.A.; Gandour, M.; Megdiche, W.; Hamed, K.B.; Amor, N.B.; Brown, S.C.; Savouré, A.; Abdelly, C. Ecophysiological and genomic analysis of salt tolerance of Cakile maritima. Environ. Exp. Bot. 2013, 92, 64–72. [Google Scholar] [CrossRef]
- Arbelet-Bonnin, D.; Ben-Hamed-Louati, I.; Laurenti, P.; Abdelly, C.; Ben-Hamed, K.; Bouteau, F. Cakile maritima, a promising model for halophyte studies and a putative cash crop for saline agriculture. Adv. Agron. 2019, 155, 45–78. [Google Scholar]
- Clausing, G.; Vickers, K.; Kadereit, J.W. Historical biogeography in linear system: Genetic variation of Sea Rocket (Cakile maritima) and Sea Holly (Eryngium maritimum) along European coasts. Mol. Ecol. 2000, 9, 1823–1833. [Google Scholar] [CrossRef] [PubMed]
- Amor, N.B.; Jiménez, A.; Megdiche, W.; Lundqvist, M.; Sevilla, F.; Abdelly, C. Response of antioxidant systems to NaCl stress in the halophyte Cakile maritima. Physiol. Plant. 2006, 126, 446–457. [Google Scholar] [CrossRef]
- Placines, C.; Castañeda-Loaiza, V.; João Rodrigues, M.; Pereira, C.G.; Stefanucci, A.; Mollica, A.; Zengin, G.; Llorent-Martínez, E.J.; Castilho, P.C.; Custódio, A.L. Phenolic profile, toxicity, enzyme inhibition, in silico studies, and antioxidant properties of Cakile maritima Scop. (Brassicaceae) from Southern Portugal. Plants 2020, 9, 142. [Google Scholar] [CrossRef] [PubMed]
- Farhat, N.; Belghith, I.; Senkler, J.; Hichri, S.; Abdelly, C.; Braun, H.P.; Debez, A. Recovery aptitude of the halophyte Cakile maritima upon water deficit stress release is sustained by extensive modulation of the leaf proteome. Ecotoxicol. Environ. Saf. 2019, 179, 198–211. [Google Scholar] [CrossRef]
- Farhat, N.; Kouas, W.; Braun, H.P.; Debez, A. Stability of thylakoid protein complexes and preserving photosynthetic efficiency are crucial for the successful recovery of the halophyte Cakile maritima from high salinity. Plant Physiol. Biochem. 2021, 166, 177–190. [Google Scholar] [CrossRef]
- Houmani, H.; Debez, A.; Freitas-Silva, L.D.; Abdelly, C.; Palma, J.M.; Corpas, F.J. Potassium (K+) starvation-induced oxidative stress triggers a general boost of antioxidant and NADPH-generating systems in the halophyte Cakile maritima. Antioxidants 2022, 11, 401. [Google Scholar] [CrossRef] [PubMed]
- Houmani, H.; Palma, J.M.; Corpas, F.J. High Salinity Stimulates the Adaptive Response to Potassium Deficiency Through the Antioxidant and the NADPH-Generating Systems in the Roots and Leaves of the Halophyte Cakile maritima. J. Plant Growth Regul. 2022, 42, 6286–6306. [Google Scholar] [CrossRef]
- Houmani, H.; Rodríguez-Ruiz, M.; Palma, J.M.; Abdelly, C.; Corpas, F.J. Modulation of superoxide dismutase (SOD) isozymes by organ development and high long-term salinity in the halophyte Cakile maritima. Protoplasma 2016, 253, 885–894. [Google Scholar] [CrossRef]
- Houmani, H.; Corpas, F.J. Can nutrients act as signals under abiotic stress? Plant Physiol. Biochemistry 2023, 206, 108313. [Google Scholar] [CrossRef] [PubMed]
- Soltabayeva, A.; Ongaltay, A.; Omondi, J.O.; Srivastava, S. Morphological, physiological and molecular markers for salt-stressed plants. Plants 2021, 10, 243. [Google Scholar] [CrossRef]
- Li, D.; Ma, W.; Wei, J.; Mao, Y.; Peng, Z.; Zhang, J.; Kong, X.; Han, Q.; Fan, W.; Yang, Y.; et al. Magnesium promotes root growth and increases aluminum tolerance via modulation of nitric oxide production in Arabidopsis. Plant Soil 2020, 457, 83–95. [Google Scholar] [CrossRef]
- Koch, M.; Busse, M.; Naumann, M.; Jákli, B.; Smit, I.; Cakmak, I.; Hermans, C.; Pawelzik, E. Differential effects of varied potassium and magnesium nutrition on production and partitioning of photoassimilates in potato plants. Physiol. Plant 2019, 166, 921–935. [Google Scholar] [CrossRef]
- Koch, M.; Winkelmann, M.K.; Hasler, M.; Pawelzik, E.; Naumann, M. Root growth in light of changing magnesium distribution and transport between source and sink tissues in potato (Solanum tuberosum L.). Sci. Rep. 2020, 10, 8796. [Google Scholar] [CrossRef]
- Ishfaq, M.; Zhong, Y.; Wang, Y.; Li, X. Magnesium limitation leads to transcriptional down-tuning of auxin synthesis, transport, and signaling in the tomato root. Front. Plant Sci. 2021, 12, 802399. [Google Scholar] [CrossRef]
- Jia, Y.; Xu, H.; Wang, Y.; Ye, X.; Lai, N.; Huang, Z.; Yang, L.; Li, Y.; Chen, L.S.; Guo, J. Differences in morphological and physiological features of citrus seedlings are related to Mg transport from the parent to branch organs. BMC Plant Biol. 2021, 21, 239. [Google Scholar] [CrossRef]
- Schneider, G.; Lindqvist, Y.; Branden, C.I. Rubisco—Structure and mechanism. Annu. Rev. Biophys. Biomol. Struct. 1992, 21, 119–143. [Google Scholar] [CrossRef] [PubMed]
- Jaghdani, S.J.; Jahns, P.; Tränkner, M. Mg deficiency induces photo-oxidative stress primarily by limiting CO2 assimilation and not by limiting photosynthetic light utilization. Plant Sci. 2021, 302, 110751. [Google Scholar] [CrossRef]
- Farhat, N.; Elkhouni, A.; Zorrig, W.; Smaoui, A.; Abdelly, C.; Rabhi, M. Effects of magnesium deficiency on photosynthesis and carbohydrate partitioning. Acta Physiol. Plant. 2016, 38, 145. [Google Scholar] [CrossRef]
- El-Ezz, S.F.A.; Al-Harbi, N.A.; Al-Qahtani, S.M.; Allam, H.M.; Abdein, M.A.; Abdelgawad, Z.A. A Comparison of the effects of several foliar forms of magnesium fertilization on ‘Superior Seedless’ (Vitis vinifera L.) in saline soils. Coatings 2022, 12, 201. [Google Scholar] [CrossRef]
- Pang, J.J.; Shin, J.S.; Li, S.Y. The Catalytic Role of RuBisCO for in situ CO2 recycling in Escherichia coli. Front. Bioeng. Biotechnol. 2020, 8, 543807. [Google Scholar] [CrossRef] [PubMed]
- Jamali Jaghdani, S.; Jahns, P.; Tränkner, M. The impact of magnesium deficiency on photosynthesis and photoprotection in Spinacia oleracea. Plant Stress 2021, 2, 100040. [Google Scholar] [CrossRef]
- Pena-Rojas, K.; Aranda, X.; Fleck, I. Stomatal limitation to CO2 assimilation and down-regulation of photosynthesis in Quercus ilex resprouts in response to slowly imposed drought. Tree Physiol. 2004, 24, 813–822. [Google Scholar] [CrossRef] [PubMed]
- Santos, E.F.; Mateus, N.S.; Rabêlo, F.H.S.; Macedo, F.G.; Lavres, J. Diagnosing early disorders in Jatropha curcas to calcium, magnesium and sulfur deficiency. J. Plant Nutr. 2020, 43, 1604–1616. [Google Scholar] [CrossRef]
- Benslima, W.; Zorrig, W.; Bagues, M.; Abdelly, C.; Hafsi, C. Silicon mitigates potassium deficiency in Hordeum vulgare by improving growth and photosynthetic activity but not through polyphenol accumulation and the related antioxidant potential. Plant Soil 2022, 477, 153–170. [Google Scholar] [CrossRef]
- Xu, B.; Long, Y.; Feng, X.; Zhu, X.; Sai, N.; Chirkova, L.; Betts, A.; Herrmann, J.; Edwards, E.J.; Okamoto, M.; et al. GABA signalling modulates stomatal opening to enhance plant water use efficiency and drought resilience. Nat. Commun. 2021, 12, 1952. [Google Scholar] [CrossRef]
- Waraich, E.A.; Ahmad, R.; Ashraf, M.Y.; Saifullah; Ahmad, M. Improving agricultural water use efficiency by nutrient management in crop plants. Acta Agric. Scand Sect. B-Soil Plant Sci. 2011, 61, 291–304. [Google Scholar] [CrossRef]
- Wang, P.; Grimm, B. Connecting chlorophyll metabolism with accumulation of the photosynthetic apparatus. Trends Plant Sci. 2021, 26, 484–495. [Google Scholar] [CrossRef] [PubMed]
- Hörtensteiner, S. Chlorophyll degradation during senescence. Annu. Rev. Plant Biol. 2006, 57, 55–77. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Okazawa, A.; Fukusaki, E.; Kobayashi, A. Removal of magnesium by Mg-dechelatase is a major step in the chlorophyll-degrading pathway in Ginkgo biloba in the process of autumnal tints. Z. Naturforsch. C. 2000, 55, 923–926. [Google Scholar] [CrossRef] [PubMed]
- Hermans, C.; Johnson, G.N.; Strasser, R.J.; Verbruggen, N. Physiological characterisation of magnesium deficiency in sugar beet: Acclimation to low magnesium differentially affects photosystems I and II. Planta 2004, 220, 344–355. [Google Scholar] [CrossRef] [PubMed]
- Hermans, C.; Bourgis, F.; Faucher, M.; Strasser, R.J.; Delrot, S.; Verbruggen, N. Magnesium deficiency in sugar beets alters sugar partitioning and phloem loading in young mature leaves. Planta 2005, 220, 541–549. [Google Scholar] [CrossRef] [PubMed]
- Hermans, C.; Verbruggen, N. Physiological characterization of Mg deficiency in Arabidopsis thaliana. J. Exp. Bot. 2005, 56, 2153–2161. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Li, J.; Yu, Y.; Kou, X.; Periakaruppan, R.; Chen, X.; Li, X. STAY-GREEN and light-harvesting complex II chlorophyll a/b binding protein are involved in albinism of a novel albino tea germplasm ‘Huabai 1’. Sci. Hortic. 2022, 293, 110653. [Google Scholar] [CrossRef]
- Zhao, Y.; Kong, H.; Guo, Y.; Zou, Z. Light-harvesting chlorophyll a/b-binding protein-coding genes in jatropha and the comparison with castor, cassava and arabidopsis. PeerJ 2020, 8, e8465. [Google Scholar] [CrossRef]
- Shimoda, Y.; Ito, H.; Tanaka, A. Arabidopsis STAY-GREEN, Mendel’s green cotyledon gene, encodes magnesium-dechelatase. Plant Cell 2016, 28, 2147–2160. [Google Scholar] [CrossRef]
- Tanoi, K.; Kobayashi, N.I. Leaf senescence by magnesium deficiency. Plants 2015, 4, 756–772. [Google Scholar] [CrossRef]
- Tewari, R.K.; Yadav, N.; Gupta, R.; Kumar, P. Oxidative stress under macronutrient deficiency in plants. J. Soil Sci. Plant Nutr. 2021, 21, 832–859. [Google Scholar] [CrossRef]
- Tewari, R.K.; Kumar, P.; Sharma, P.N. Magnesium deficiency induced oxidative stress and antioxidant responses in mulberry plants. Sci. Hortic. 2006, 108, 7–14. [Google Scholar] [CrossRef]
- Tränkner, M.; Jaghdani, S.J. Minimum magnesium concentrations for photosynthetic efficiency in wheat and sunflower seedlings. Plant Physiol. Biochem. 2019, 144, 234–243. [Google Scholar] [CrossRef]
- Cai, Y.T.; Zhang, H.; Qi, Y.P.; Ye, X.; Huang, Z.R.; Guo, J.X.; Chen, L.S.; Yang, L.T. Responses of reactive oxygen species and methylglyoxal metabolisms to magnesium-deficiency differ greatly among the roots, upper and lower leaves of Citrus sinensis. BMC Plant Biol. 2019, 9, 76. [Google Scholar]
- Houmani, H.; Rodriguez-Ruiz, M.; Palma, J.M.; Corpas, F.J. Mechanical wounding promotes local and long distance response in the halophyte Cakile maritima through the involvement of the ROS and RNS metabolism. Nitric Oxide 2018, 74, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Arnon, D.T. Copper enzymes in isolated chloroplasts. Polyphenol oxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Sairam, R.K.; Deshmukh, P.S.; Shukla., D.S. Tolerance of Drought and Temperature Stress in Relation to Increased Antioxidant Enzyme Activity in Wheat. J. Agron. Crop Sci. 1997, 178, 171–178. [Google Scholar] [CrossRef]
- Sairam, R.K.; Srivastava, G.C. Changes in antioxidant activity in sub-cellular fractions of tolerant and susceptible wheat genotypes in response to long term salt stress. Plant Sci. 2002, 162, 897–904. [Google Scholar] [CrossRef]
- Maghsoudi, K.; Emam, Y.; Pessarakli, M. Effect of silicon on photosynthetic gas exchange, photosynthetic pigments, cell membrane stability and relative water content of different wheat cultivars under drought stress conditions. J. Plant Nutr. 2016, 39, 1001–1015. [Google Scholar] [CrossRef]
- Draper, H.H.; Hadley, M. [43] Malondialdehyde determination as index of lipid Peroxidation. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1990; Volume 186, pp. 421–431. [Google Scholar]
- Kausar, A.; Ashraf, M.Y.; Ali, I.; Niaz, M.; Abbass, Q. Evaluation of sorghum varieties/lines for salt tolerance using physiological indices as screening tool. Pak. J. Bot. 2012, 44, 47–52. [Google Scholar]
- Majeed, S.; Nawaz, F.; Naeem, M.; Ashraf, M.Y. Effect of exogenous nitric oxide on sulfur and nitrate assimilation pathway enzymes in maize (Zea mays L.) under drought stress. Acta Physiol. Plant. 2018, 40, 206. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| +Mg | −Mg | |
|---|---|---|
| SHSI (%) | 100 ± 0.00 a | 70.33 ± 3.97 b |
| PHSI (%) | 100 ± 0.00 a | 83.16 ± 5.10 b |
| SFSI (%) | 100 ± 0.00 a | 58.43 ± 3.80 b |
| RFSI (%) | 100 ± 0.00 a | 50.08 ± 5.30 b |
| SDSI (%) | 100 ± 0.00 a | 68.61 ± 2.23 b |
| RDSI (%) | 100 ± 0.00 a | 55.36 ± 6.58 b |
| DMSI(%) | 100 ± 0.00 a | 66.92 ± 1.83 b |
| +Mg | −Mg | |
|---|---|---|
| Chla (mg g−1 FW) | 0.39 ± 0.03 a | 0.18 ± 0.01 b |
| Chlb (mg g−1 FW) | 0.22 ± 0.01 a | 0.09 ± 0.01 b |
| Chla+Chlb (mg g−1 FW) | 0.61 ± 0.04 a | 0.27 ± 0.02 b |
| Car (mg g−1 FW) | 0.023 ± 0.002 a | 0.025 ± 0.001 a |
| Chla/Chlb | 1.81 ± 0.12 a | 2.09 ± 0.08 a |
| CSI (%) | 100 ± 0.00 a | 44.51 ± 4.54 b |
| Treatment Variable | +Mg | −Mg |
|---|---|---|
| Shoot lenght | 0.90 *** | −0.90 *** |
| Plant lenght | 0.69 ** | −0.69 ** |
| Leaf number | 0.98 *** | −0.98 *** |
| Leaf FW | 0.77 *** | −0.77 *** |
| Stem FW | 0.87 *** | −0.87 *** |
| Root FW | 0.84 *** | −0.84 *** |
| SHSI | 0.89 *** | −0.89 *** |
| PHSI | 0.67 ** | −0.67 ** |
| SFSI RFSI | 0.95 *** 0.92 *** | −0.95 *** −0.92 *** |
| SDSI | 0.96 *** | −0.96 *** |
| RDSI | 0.86 *** | −0.86 *** |
| DMSI | 0.97 *** | −0.97 *** |
| Chla | 0.93 *** | −0.93 *** |
| Chlb | 0.98 *** | −0.98 *** |
| Chla+Chlb | 0.96 *** | −0.96 *** |
| Car | −0.263 | 0.263 |
| Chla/b | −0.631 | 0.631 |
| CSI | 0.98 *** | −0.98 *** |
| EL | −0.94 *** | 0.94 *** |
| MDA | −0.95 *** | 0.95 *** |
| MSI | 0.94 *** | −0.94 *** |
| A | 0.85 *** | −0.85 *** |
| gs | 0.89 *** | −0.89 *** |
| E | 0.75 * | −0.75 * |
| WUE | 0.76 * | −0.76 * |
| IWUE | 0.133 | −0.133 |
| ICE | 0.91 *** | −0.91 *** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Houmani, H.; Hidri, R.; Farhat, N.; Debez, A. Insights into Key Biometric, Physiological and Biochemical Markers of Magnesium (Mg) Deficiency Stress in the Halophyte Cakile maritima. Stresses 2024, 4, 342-357. https://doi.org/10.3390/stresses4020022
Houmani H, Hidri R, Farhat N, Debez A. Insights into Key Biometric, Physiological and Biochemical Markers of Magnesium (Mg) Deficiency Stress in the Halophyte Cakile maritima. Stresses. 2024; 4(2):342-357. https://doi.org/10.3390/stresses4020022
Chicago/Turabian StyleHoumani, Hayet, Rabaa Hidri, Nèjia Farhat, and Ahmed Debez. 2024. "Insights into Key Biometric, Physiological and Biochemical Markers of Magnesium (Mg) Deficiency Stress in the Halophyte Cakile maritima" Stresses 4, no. 2: 342-357. https://doi.org/10.3390/stresses4020022
APA StyleHoumani, H., Hidri, R., Farhat, N., & Debez, A. (2024). Insights into Key Biometric, Physiological and Biochemical Markers of Magnesium (Mg) Deficiency Stress in the Halophyte Cakile maritima. Stresses, 4(2), 342-357. https://doi.org/10.3390/stresses4020022