
Functional and Qualitative Metabolic Compounds in the Twigs of the Deciduous Mistletoe Loranthus europaeus Jacq.

 ,
,  ,
,
Abstract
:1. Introduction
2. Results
2.1. Secondary Metabolites
2.2. Other Components
3. Discussion
4. Materials and Methods
4.1. Research Site
4.2. Collection of Specimens
4.3. Extraction of Plant Material for HRMS Analysis
4.4. LC-HRMS-Based Analysis and Dereplication
4.5. Soluble Sugar and Starch Content
4.6. Chlorophyll Content
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Balle, S.; Dandy, J.E.; Gilmour, J.S.L.; Holltum, R.E.; Stearn, W.T.; Thoday, D. Loranthus. Taxon 1960, 9, 208–210. Available online: https://www.jstor.org/stable/1216271 (accessed on 10 October 2023).
- Snogerup, S.; Snogerup, B.B. Loranthus L. In Flora Hellenica; Strid, A., Tan, K., Eds.; Koeltz: Königstein, Germany, 1997; Volume 1, p. 67. [Google Scholar]
- Plants of the World Online. Available online: https://powo.science.kew.org/taxon/urn:lsid:ipni.org:names:549404-1 (accessed on 10 October 2023).
- Briggs, J. Loranthus europaeus, new to Britain, and notes on Viscum album, both new to Kew Gardens. Haustorium 2009, 56, 2–3. [Google Scholar]
- Glatzel, G. Mineral nutrition and water relations of hemiparasitic mistletoes: A question of partitioning. Experiments with Loranthus europaeus on Quercus petraea and Quercus robur. Oecologia 1983, 56, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Schulze, E.-D.; Turner, N.C.; Glatzel, G. Carbon, water and nutrient relations of two mistletoes and their hosts: A hypothesis. Plant Cell Environ. 1984, 7, 239–299. [Google Scholar] [CrossRef]
- Griffith, J. The Oak: Its Natural History, Antiquity, and Folk-Lore. Nature 1913, 90, 589–590. [Google Scholar] [CrossRef]
- Kubov, M.; Fleischer, P., Jr.; Rozkošný, J.; Kurjak, D.; Konôpková, A.; Galko, J.; Húdoková, H.; Lalík, M.; Rell, S.; Pittner, J.; et al. Drought or severe drought? Hemiparasitic yellow mistletoe (Loranthus europaeus) amplifies drought stress in sessile oak trees (Quercus petraea) by altering water status and physiological responses. Water 2020, 12, 2985. [Google Scholar] [CrossRef]
- Harvala, E.; Exner, J.; Becker, H. Flavonoids of Loranthus europaeus. J. Nat. Prod. 1984, 47, 1054–1055. [Google Scholar] [CrossRef]
- Krasylenko, Y.A.; Gleb, R.Y.; Volutsa, O.D. Loranthus europaeus (Loranthaceae) in Ukraine: An overview of distribution patterns and hosts. Ukr. Bot. J. 2019, 76, 406–417. [Google Scholar] [CrossRef]
- Leroy, T.; Plomion, C.; Kremer, A. Oak symbolism in the light of genomics. New Phytol. 2020, 226, 1012–1017. [Google Scholar] [CrossRef]
- Barnht, R.K.; Steinmetz, S. Chambers Dictionary of Etymology; Chambers Harrap Publishers Ltd.: Edinburgh, UK, 2008. [Google Scholar]
- Riddle, J.M. History as a tool in identifying ‘new’ old drugs. Adv. Exp. Med. Biol. 2002, 505, 89–94. [Google Scholar] [CrossRef]
- Paine, L.K.; Harrison, H.C. Mistletoe: Its role in horticulture and human life. HortTechnology 1992, 2, 324–330. [Google Scholar] [CrossRef]
- Hejcman, M.; Hejcmanová, P.; Stejskalová, M.; Pavlů, V. Nutritive value of winter-collected annual twigs of main European woody species, mistletoe and ivy and its possible consequences for winter foddering of livestock in prehistory. Holocene 2014, 24, 659–667. [Google Scholar] [CrossRef]
- Brooks, R.A. Discolor Aura. Reflections on the golden bough. Amer. J. Philol. 1953, 74, 260–280. [Google Scholar] [CrossRef]
- Krappe, A.H. The myth of Balder: A study in comparative mythology. Folklore 1923, 34, 184–215. [Google Scholar] [CrossRef]
- Kanner, L. Mistletoe, Magic and Medicine. Bull. Hist. Med. 1939, 7, 875–936. [Google Scholar]
- Nickrent, D.L.; Malécot, V.; Vidal-Russell, R.; Der, J.P. A revised classification of Santalales. Taxon 2010, 59, 538–558. [Google Scholar] [CrossRef]
- Griffis, F.C. Mistletoe myth develops industry. J. Geogr. 1956, 55, 251–254. [Google Scholar] [CrossRef]
- Wolf-Rottkay, W.H. Balder and the Mistletoe: A Note. Scand. Stud. 1967, 39, 340–344. [Google Scholar]
- Danser, B.H. The Loranthaceae of French Indochina and Siam. Bull. Jard. Bot. Buitenzorg 1938, 16, 1–63. [Google Scholar]
- Nickrent, D.L.; Su, H.J.; Lin, R.Z.; Devkota, M.P.; Hu, J.M.; Glatzel, G. Examining the needle in the haystack: Evolutionary relationships in the mistletoe genus Loranthus (Loranthaceae). Syst. Bot. 2021, 46, 403–415. [Google Scholar] [CrossRef]
- Theophrastus. De Causis Plantarum; Einarson, B.; Link, G.K.K., Translators; Harvard University Press: Cambridge, MA, USA, 1976 and 1990; Volumes 1–3. [Google Scholar]
- Theophrastus. Enquiry into Plants; Hort, A.F., Translator; Harvard University Press: Cambridge, MA, USA, 1916 and 1926; Volumes 1–2. [Google Scholar]
- Theophrastus. The Compiled Works; Kactos: Athens, Greece, 1998; Volumes 1–7. [Google Scholar]
- Thanos, C.A. Theophrastus on oaks. Bot. Chron. 2005, 18, 29–36. [Google Scholar]
- Lack, H.W.; Mabberley, D.J. The Flora Graeca Story, Sibthorp, Bauer and Hawkins in the Levant; Oxford University Press: Oxford, UK, 1999. [Google Scholar]
- Katsarou, A.; Rhizopoulou, S.; Kefalas, P. Antioxidant potential of the aerial tissues of the mistletoe Loranthus europaeus Jacq. Rec. Nat. Prod. 2012, 6, 394–397. [Google Scholar]
- Chimona, C.; Papadopoulou, S.; Kolyva, F.; Mina, M.; Rhizopoulou, S. From Biodiversity to Musketry: Detection of Plant Diversity in Pre-Industrial Peloponnese during the Flora Graeca Expedition. Life 2022, 12, 1957. [Google Scholar] [CrossRef] [PubMed]
- Zebec, M.; Idžojtić, M. Hosts and distribution of yellow mistletoe, Loranthus europaeus Jacq. in Croatia. Hladnikia 2006, 19, 41–46. [Google Scholar]
- Benabderrahim, M.A.; Elfalleh, W.; Sarikurkcu, C.; Sarikurkcu, R.B. Biological activities and phytochemical composition of organs from Loranthus europaeus. Ind. Crops Prod. 2019, 141, 111772. [Google Scholar] [CrossRef]
- Kumal, K.; Pant, D.R.; Aryal, B.; Tripathi, G.R.; Joshi, G.P. Phytochemical and antioxidant properties of traditionally used mistletoes in Nepal. Sci. World 2021, 14, 83–89. [Google Scholar] [CrossRef]
- Ambrosio, R.L.; Gratino, L.; Mirino, S.; Cocca, E.; Pollio, A.; Anastasio, A.; Palmieri, G.; Balestrieri, M.; Genovese, A.; Goglittino, M. The bactericidal activity of protein extracts from Loranthus europaeus berries: A natural resource of bioactive compounds. Antibiotics 2020, 9, 47. [Google Scholar] [CrossRef] [PubMed]
- Eliáš, P.; Huzulák, J. Contribution to the study of water relationships between hemiparasite (Loranthus europaeus L.) and its host (Quercus cerris L.). Acta Bot. Slov. Acad. Sci. Slovacae 1978, 4, 265–276. [Google Scholar]
- Gebauer, R.; Volařík, D.; Urban, J. Quercus pubescens and its hemiparasite Loranthus europaeus: Nutrient dynamics of leaves and twigs. Acta Physiol. Plant. 2012, 34, 1801–1809. [Google Scholar] [CrossRef]
- Kubíček, J.; Špinlerová, Z.; Michalko, R.; Vrška, T.; Matula, R. Temporal dynamics and size effects of mistletoe (Loranthus europaeus Jacq.) infection in an oak forest. Austrian J. For. Sci. 2018, 135, 119–135. [Google Scholar]
- Gebauer, R.; Albrechtová, P.; Plichta, R.; Volařík, D. The comparative xylem structure and function of petioles and twigs of mistletoe Loranthus europaeus and its host Quercus pubescence. Trees 2019, 33, 933–942. [Google Scholar] [CrossRef]
- Bernhofer, C.; Gay, L.W. Evapotranspiration from an oak forest infested by mistletoe. Agric. For. Meteorol. 1989, 48, 205–223. [Google Scholar] [CrossRef]
- Doležal, J.; Lehečková, E.; Sohar, K.; Altman, J. Oak decline induced by mistletoe, competition and climate change: A case study from central Europe. Preslia 2016, 88, 323–346. [Google Scholar]
- Barlow, B.A. Loranthaceae. In Flora Malesiana-Series 1, Spermatophyta; Noordhoff-Kolff: Djakarta, Indonesia, 1997; Volume 13, pp. 209–401. [Google Scholar]
- Shavvon, R.S.; Mehrvarz, S.S.; Golmohammadi, N. Evidence from micromorphology and gross morphology of the genus Loranthus (Loranthaceae) in Iran. Turk. J. Bot. 2012, 36, 655–666. [Google Scholar] [CrossRef]
- Gadotti, V.M.; Santos, A.R.; Meyre-Silva, C.; Schmeling, L.O.; Machado, C.; Liz, F.H.; Filho, V.C. Antinociceptive action of the extract and the flavonoid quercitrin isolated from Bauhinia microstachya leaves. J. Pharm. Pharmacol. 2005, 57, 1345–1351. [Google Scholar] [CrossRef]
- Del Bubba, M.; Checchini, L.; Chiuminatto, U.; Doumett, S.; Fibbi, D.; Giordani, E. Liquid chromatographic/electrospray ionization tandem mass spectrometric study of polyphenolic composition of four cultivars of Fragaria vesca L. berries and their comparative evaluation. J. Mass Spectrom. 2012, 47, 1207–1220. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Fang, S.; Yin, Z.; Fu, X.; Shang, X.; Yang, W.; Yang, H. Chemical fingerprint and multicomponent quantitative analysis for the quality evaluation of Cyclocarya paliurus leaves by HPLC–Q–TOF–MS. Molecules 2017, 22, 1927. [Google Scholar] [CrossRef] [PubMed]
- Menezes, R.D.P.; Bessa, M.A.D.S.; Siqueira, C.D.P.; Teixeira, S.C.; Ferro, E.A.V.; Martins, M.M.; Cunha, L.C.S.; Martins, C.H.G. Antimicrobial, Antivirulence, and Antiparasitic Potential of Capsicum chinense Jacq. Extracts and Their Isolated Compound Capsaicin. Antibiotics 2022, 11, 1154. [Google Scholar] [CrossRef] [PubMed]
- Shi, F.; Xie, L.; Lin, Q.; Tong, C.; Fu, Q.; Xu, J.; Xiao, J.; Shi, S. Profiling of tyrosinase inhibitors in mango leaves for a sustainable agro-industry. Food Chem. 2020, 312, 126042. [Google Scholar] [CrossRef]
- Seukep, J.A.; Zhang, Y.L.; Xu, Y.B.; Guo, M.Q. In vitro antibacterial and antiproliferative potential of Echinops lanceolatus Mattf. (Asteraceae) and identification of potential bioactive compounds. Pharmaceuticals 2020, 13, 59. [Google Scholar] [CrossRef]
- Xia, C.; Deng, J.; Pan, Y.; Lin, C.; Zhu, Y.; Xiang, Z.; Li, W.; Chen, J.; Zhang, Y.; Zhu, B.; et al. Comprehensive profiling of macamides and fatty acid derivatives in maca with different postharvest drying processes using UPLC-QTOF-MS. ACS Omega 2021, 6, 24484–24492. [Google Scholar] [CrossRef]
- Strassburg, K.; Huijbrechts, A.M.; Kortekaas, K.A.; Lindeman, J.H.; Pedersen, T.L.; Dane, A.; Berger, R.; Brenkman, A.; Hankemeier, T.; van Duynhoven, J.; et al. Quantitative profiling of oxylipins through comprehensive LC-MS/MS analysis: Application in cardiac surgery. Anal. Bioanal. Chem. 2012, 404, 1413–1426. [Google Scholar] [CrossRef] [PubMed]
- Pecio, Ł.; Kozachok, S.; Saber, F.R.; Garcia-Marti, M.; El-Amier, Y.; Mahrous, E.A.; Swiątek, L.; Boguszewska, A.; Skiba, A.; Elosaily, A.H.; et al. Metabolic profiling of Ochradenus baccatus Delile. utilizing UHPLC-HRESIMS in relation to the in vitro biological investigations. Food Chem. 2023, 412, 135587. [Google Scholar] [CrossRef]
- Tao, Y.; Jiang, Y.; Li, W.; Cai, B. Rapid magnetic solid-phase extraction combined with ultra-high performance liquid chromatography and quadrupole-time-of-flight mass spectrometry for analysis of thrombin binders from a crude extract and injection of Erigeron breviscapus. RSC Adv. 2016, 6, 34782–34790. [Google Scholar] [CrossRef]
- Gläser, P.; Dawid, C.; Meister, S.; Bader-Mittermaier, S.; Schott, M.; Eisner, P.; Hofmann, T. Molecularization of bitter off-taste compounds in pea-protein isolates (Pisum sativum L.). J. Agric. Food Chem. 2020, 68, 10374–10387. [Google Scholar] [CrossRef] [PubMed]
- Levison, B.S.; Zhang, R.; Wang, Z.; Fu, X.; DiDonato, J.A.; Hazen, S.L. Quantification of fatty acid oxidation products using online high-performance liquid chromatography tandem mass spectrometry. Free Radic. Biol. Med. 2013, 59, 2–13. [Google Scholar] [CrossRef] [PubMed]
- Saeed, M.M.; Fernández-Ochoa, Á.; Saber, F.R.; Sayed, R.H.; Cádiz-Gurrea, M.D.L.L.; Elmotayam, A.K.; Leyva-Jiménez, F.-J.; Segura-Carretero, A.; Nadeem, R.I. The Potential Neuroprotective Effect of Cyperus esculentus L. Extract in Scopolamine-Induced Cognitive Impairment in Rats: Extensive Biological and Metabolomics Approaches. Molecules 2022, 27, 7118. [Google Scholar] [CrossRef] [PubMed]
- Grati, W.; Samet, S.; Bouzayani, B.; Ayachi, A.; Treilhou, M.; Téné, N.; Mezghani-Jarraya, R. HESI-MS/MS Analysis of phenolic compounds from Calendula aegyptiaca fruits extracts and evaluation of their antioxidant activities. Molecules 2022, 27, 2314. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Qin, Y.; Li, H.; Peng, H.; Chen, H.; Xie, H.R.; Deng, Z. Rapid characterization of chemical constituents in Radix tetrastigma, a functional herbal mixture, before and after metabolism and their antioxidant/antiproliferative activities. J. Funct. Foods 2015, 18, 300–318. [Google Scholar] [CrossRef]
- El-Shazly, M.A.; Hamed, A.A.; Kabary, H.A.; Ghareeb, M.A. LC-MS/MS profiling, antibiofilm, antimicrobial and bacterial growth kinetic studies of Pluchea dioscoridis extracts. Acta Chromatogr. 2022, 34, 338–350. [Google Scholar] [CrossRef]
- Yerlikaya, S.; Baloglu, M.C.; Diuzheva, A.; Jekő, J.; Cziáky, Z.; Zengin, G. Investigation of chemical profile, biological properties of Lotus corniculatus L. extracts and their apoptotic-autophagic effects on breast cancer cells. J. Pharm. Biomed. Anal. 2019, 174, 286–299. [Google Scholar] [CrossRef]
- Qian, X.Y.; Nie, L.X.; Zhao, H.; Dai, Z.; Ma, S.C.; Liu, J.M.; Kuang, Y.H. Discovery and molecular elucidation of the anti-influenza material basis of Banlangen granules based on biological activities and ultra-high performance liquid chromatography coupled with quadrupole-orbitrap mass spectrometry. J. Ethnopharmacol. 2022, 298, 115683. [Google Scholar] [CrossRef] [PubMed]
- Eid, H.M.; Ouchfoun, M.; Saleem, A.; Guerrero-Analco, J.A.; Walshe-Roussel, B.; Musallam, L.; Rapinski, M.; Cuerrier, A.; Martineau, L.C.; Arnason, J.T.; et al. A combination of (+)-catechin and (−)-epicatechin underlies the in vitro adipogenic action of Labrador tea (Rhododendron groenlandicum), an antidiabetic medicinal plant of the Eastern James Bay Cree pharmacopeia. J. Ethnopharmacol. 2016, 178, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Shin, D.W.; Kim, S.N.; Lee, S.M.; Lee, W.; Song, M.J.; Park, S.M.; Lee, T.R.; Baik, J.-H.; Kim, H.K.; Hong, J.-H.; et al. Catechin promotes adipocyte differentiation in human bone marrow mesenchymal stem cells through PPARγ transactivation. Biochem. Pharmacol. 2009, 77, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Blount, J.W.; Ferruzzi, M.; Raftery, D.; Pasinetti, G.M.; Dixon, R.A. Enzymatic synthesis of substituted epicatechins for bioactivity studies in neurological disorders. Biochem. Biophys. Res. Commun. 2012, 417, 457–461. [Google Scholar] [CrossRef] [PubMed]
- He, F.; Pan, Q.H.; Shi, Y.; Duan, C.Q. Biosynthesis and genetic regulation of proanthocyanidins in plants. Molecules 2008, 13, 2674–2703. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Li, W.; Son, Y.O.; Sun, L.; Lu, J.; Kim, D.; Wang, X.; Yao, H.; Wang, L.; Pratheeshkumar, P.; et al. Quercitrin protects skin from UVB-induced oxidative damage. Toxicol. Appl. Pharmacol. 2013, 269, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Li, G.; Sun, C.; Peng, F.; Yu, L.; Chen, Y.; Tan, Y.; Cao, X.; Tang, Y.; Xie, X.; et al. Chemistry, pharmacokinetics, pharmacological activities, and toxicity of quercitrin. Phytother. Res. 2020, 36, 1545–1575. [Google Scholar] [CrossRef]
- Aljeboory, A.A.; Al-Zubaidi, N.M.A. Loranthus europaeus is an alternative medicine in treatment of cyst and mouth inflammation resulted from chemotherapy of breast cancer. Int. J. Drug Deliv. Technol. 2020, 10, 183–186. [Google Scholar] [CrossRef]
- Nazzaro-Porro, M. Azelaic acid. JAAD 1987, 17, 1033–1041. [Google Scholar] [CrossRef]
- Gollnick, H.P.; Graupe, K.; Zaumseil, R.P. Azelaic acid 15% gel in the treatment of acne vulgaris. Combined results of two double-blind clinical comparative studies. J. Dtsch. Dermatol. Ges 2004, 2, 841–847. [Google Scholar] [CrossRef]
- Ajiboye, B.O.; Diayi, A.; Agunbiade, S.O.; Akinyemi, A.J.; Adewale, O.B.; Ojo, O.A. Ameliorating activity of polyphenolic-rich extracts of Basella rubra L. leaves on pancreatic β-cell dysfunction in streptozotocin-induced diabetic rats. J. Complement. Integr. Med. 2021, 19, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Lim, Y.C.; Rajabalaya, R.; Lee, S.H.F.; Tennakoon, K.U.; Le, Q.V.; Idris, A.; Zulkipli, I.N.; Keasberry, N.; David, S.R. Parasitic mistletoes of the genera Scurrula and Viscum: From bench to bedside. Molecules 2016, 21, 1048. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.S.; Kwon, O.W.; Kim, S.Y.; Choi, S.U.; Kim, K.H.; Lee, K.R. Five new oxylipins from Chaenomeles sinensis. Lipids 2014, 49, 1151–1159. [Google Scholar] [CrossRef] [PubMed]
- He, M.; Qin, C.-X.; Wang, X.; Ding, N.-Z. Plant Unsaturated Fatty Acids: Biosynthesis and Regulation. Front. Plant Sci. 2020, 11, 390. [Google Scholar] [CrossRef] [PubMed]
- Coniglio, S.; Shumskaya, M.; Vassiliou, E. Unsaturated Fatty Acids and Their Immunomodulatory Properties. Biology 2023, 12, 279. [Google Scholar] [CrossRef] [PubMed]
- Zhukov, A.V.; Shumskaya, M. Very-long-chain fatty acids (VLCFAs) in plant response to stress. Funct. Plant Biol. 2020, 47, 695–703. [Google Scholar] [CrossRef]
- Meletiou-Christou, M.S.; Rhizopoulou, S. Leaf functional traits of four evergreen species growing in Mediterranean environmental conditions. Acta Physiol. Plant. 2017, 39, 34. [Google Scholar] [CrossRef]
- Pfanz, H.; Aschan, G. The existence of bark photosynthesis in woody plants and its significance for the overall carbon gain. An eco-physiological approach. Prog. Bot 2001, 62, 477–510. [Google Scholar]
- Liu, J.; Gu, L.; Yu, Y.; Ju, G.; Sun, Z. Stem photosynthesis of twig and its contribution to new organ development in cutting seedlings of Salix matsudana Koidz. Forests 2018, 9, 207. [Google Scholar] [CrossRef]
- Amutenya, A.; Kwembeya, E.; Shikangalah, R.; Tsvuura, Z. Photosynthesis, chlorophyll content and water potential of a mistletoe-host pair in a semi-arid savanna. S. Afr. J. Bot. 2023, 163, 311–315. [Google Scholar] [CrossRef]
- Mosblech, L.; Feussner, I.; Heilmann, I. Oxylipins: Structurally diverse metabolites from fatty acid oxidation. Plant Physiol. Biochem. 2009, 47, 511–517. [Google Scholar] [CrossRef] [PubMed]
- Savchenko, T.; Kolla, V.A.; Wang, C.Q.; Nasafi, Z.; Hicks, D.R.; Phadungchob, B.; Chehab, W.E.; Brandizzi, F.; Froehlich, J.; Dehesh, K. Functional convergence of oxylipin and abscisic acid pathways controls stomatal closure in response to drought. Plant Physiol. 2014, 164, 1151–1160. [Google Scholar] [CrossRef]
- Milanović, S.; Janković-Tomanić, M.; Kostić, I.; Kostić, M.; Morina, F.; Živanović, B.; Lazarević, J. Behavioural and physiological plasticity of gypsy moth larvae to host plant switching. Entomol. Exp. Applic. 2016, 158, 152–162. [Google Scholar] [CrossRef]
- Wolkerstorfer, S.V.; Wonisch, A.; Stankova, T.; Tsvetkova, N.; Tausz, M. Seasonal variations of gas exchange, photosynthetic pigments, and antioxidants in Turkey oak (Quercus cerris L.) and Hungarian oak (Quercus frainetto Ten.) of different age. Trees 2011, 25, 1043–1052. [Google Scholar] [CrossRef]
- Pavlović, P.; Kostić, O.; Jarić, S.; Gajić, G.; Pavlović, D.; Pavlović, M.; Mitrović, M. The effects of leaf litter chemistry and anatomical traits on the litter decomposition rate of Quercus frainetto Ten. and Quercus cerris L. in situ. Arch. Biol. Sci. 2020, 72, 543–553. [Google Scholar] [CrossRef]
- Kitikidou, K.; Milios, E.; Tsirekis, E.; Pipinis, E.; Stampoulidis, A. Site quality assessment of degraded Quercus frainetto stands in central Greece. Iforest-Biogeosciences For. 2015, 8, 53–58. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.T.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Buysse, J.A.N.; Merckx, R. An improved colorimetric method to quantify sugar content of plant tissue. J. Exp. Bot. 1993, 44, 1627–1629. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Buschmann, C. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Buschmann, C. Chlorophylls and carotenoids: Measurement and characterization by UV–VIS spectroscopy. Curr. Protoc. Food Anal. Chem 2001, 1, F4.3.1–F4.3.8. [Google Scholar] [CrossRef]
- Wullschleger, S.D.; Norby, R.J.; Hendrix, D.L. Carbon exchange rates, chlorophyll content, and carbohydrate status of two forest tree species exposed to carbon dioxide enrichment. Tree Physiol. 1992, 10, 21–31. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
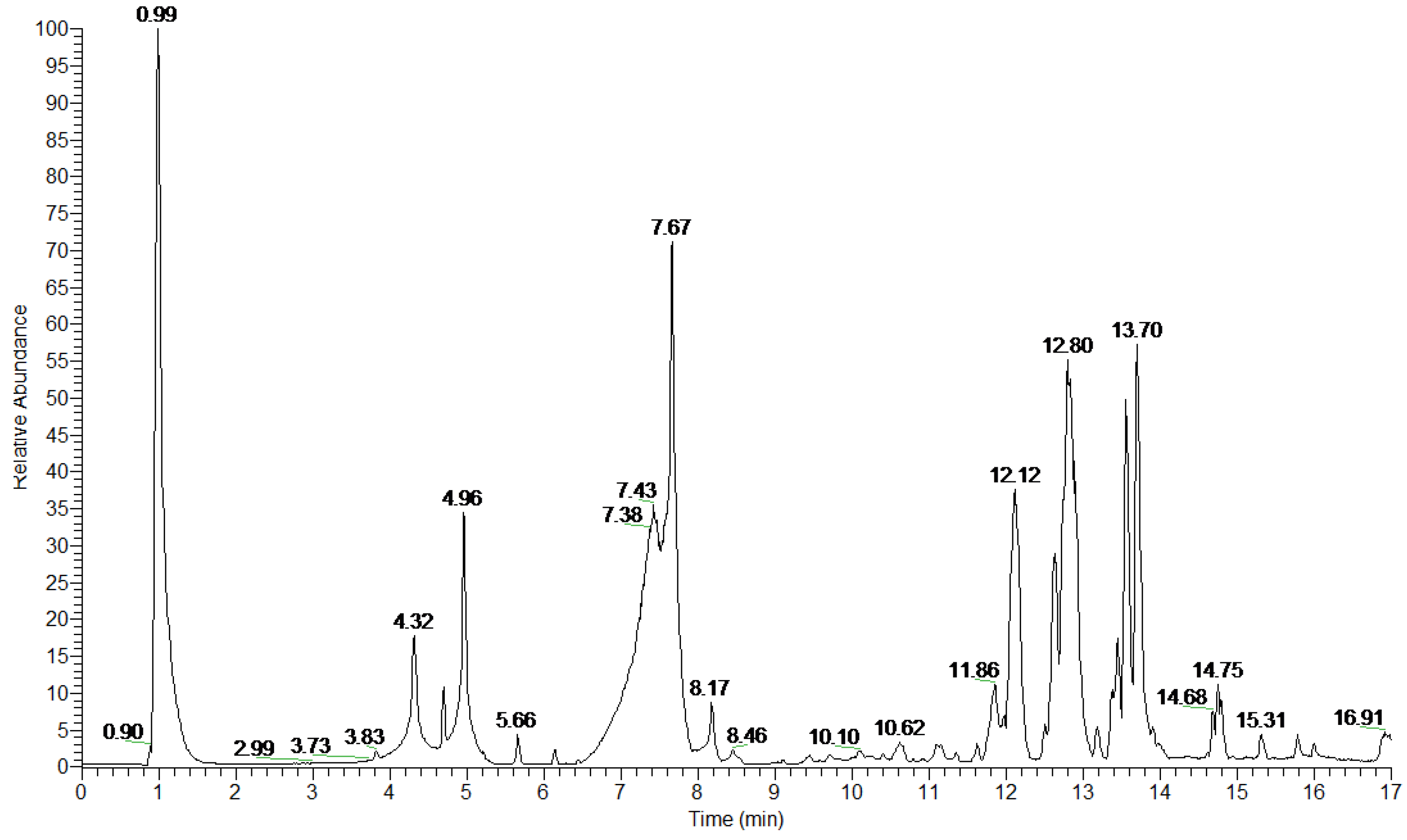
| Peak Number | Retention Time (min) | Proposed Compound | Molecular Formula, Elemental Composition | m/z Detected [M—H] | Delta Δ (ppm) | Ring and Double Bond Equivalent RDBeq. | HRMS/MS |
|---|---|---|---|---|---|---|---|
| 1 | 4.32 | Catechin 1 | C15H14O6 | 289.0715 | −0.904 | 9.5 | 245.0821 (100) 205.0505 (38) 179.0350 (15) |
| 2 | 4.70 | Procyanidin dimer type B isomer 1 | C30H26O12 | 577.1346 | −0.952 | 18.5 | 451.1031 (27) 425.0877 (100) 407.0772 (52) 289.0718 (20) |
| 3 | 4.96 | Epicatechin 1 | C15H14O6 | 289.0714 | −1.250 | 9.5 | 245.0818 (100) 205.0505 (38) 179.0350 (16) |
| 4 | 5.66 | Procyanidin dimer type B isomer 1 | C30H26O12 | 577.1348 | −0.605 | 18.5 | 451.1031 (29) 425.0876 (100) 407.0771 (53) 289.0717 (22) |
| 5 | 6.15 | Quercitrin (Quercetin-3-O-rhamnoside) 2 | C21H20O11 | 447.0930 | −0.636 | 12.5 | 301.0355 (100) 300.0278 (30) |
| 6 | 7.43 | Rhamnetin-3-O-pentoside 3 | C22H22O11 | 461.1084 | −1.159 | 12.5 | 315.0507 (100) 314.0431 (33) |
| 7 | 7.67 | Isorhamnetin-3-O-pentoside 3 | C22H22O11 | 461.1083 | −1.376 | 12.5 | 315.0506 (100) 314.0433 (33) |
| 8 | 8.17 | Rhamnocitrin pentoside | C22H22O10 | 445.1137 | −0.719 | 12.5 | 299.0560 (100) |
| 9 | 8.46 | Pinellic acid 4 | C18H34O5 | 329.2332 | −0.448 | 2.5 | 311.2228 (49) 229.1445 (91) 211.1340 (64) 171.1027 (100) |
| 10 | 9.45 | Hydroxy-oxo-octadecatrienoic acid isomer 5 | C18H28O4 | 307.1915 | −0.057 | 5.5 | 289.1807 (100) 260.9319 (41) 235.1910 (67) 185.1183 (39) |
| 11 | 10.62 | Hydroxy-oxo-octadecatrienoic acid isomer 5 | C18H28O4 | 307.1915 | −0.057 | 5.5 | 289.1808 (100) 260.9320 (8) 245.1910 (10) |
| 12 | 11.11 | Dihydroxyoctadecadienoic acid isomer 6 | C18H32O4 | 311.2227 | −0.266 | 3.5 | 293.2119 (100) 275.2015 (33) 201.1130 (41) 171.1025 (86) |
| 13 | 11.34 | Dihydroxyoctadecadienoic acid isomer 6 | C18H32O4 | 311.2227 | −0.266 | 3.5 | 293.2119 (100) 171.1025 (47) |
| 14 | 11.63 | Thapsic acid 7 | C16H30O4 | 285.2073 | 0.587 | 2.5 | 267.1963 (66) 223.2065 (100) |
| 15 | 11.86 | Oxo-octadecadienoic acid isomer 8,9 | C18H30O3 | 293.2122 | −0.062 | 4.5 | 275.2015 (100) 235.1701 (11) 211.1338 (30) 183.1390 (14) 171.1026 (15) |
| 16 | 11.97 | Oxo-octadecadienoic acid isomer 8 | C18H30O3 | 293.2121 | −0.403 | 4.5 | 275.2016 (100) 195.1390 (59) |
| 17 | 12.12 | α-Licanic acid (Oxo-octadecatrienoic acid) | C18H28O3 | 291.1962 | −1.264 | 5.5 | 273.1858 (100) 247.2065 (81) 185.1182 (34) |
| 18 | 12.63 | Hydroxy-octadecadienoic acid 8 | C18H32O3 | 295.2274 | −1.586 | 3.5 | 277.2171 (100) 195.1389 (31) 171.1026 (55) |
| 19 | 12.80 | Oxo-octadecadienoic acid isomer 10 | C18H30O3 | 293.2118 | −1.426 | 4.5 | 275.2017 (27) 249.2223 (100) 197.1183 (27) 185.1283 (76) |
| 20 | 13.45 | Ricinoleic acid 11 | C18H34O3 | 297.2432 | −1.070 | 2.5 | 279.2327 (100) 171.1026 (19) 155.1076 (68) |
| 21 | 13.56 | Hydroxy-octadecadienoic acid isomer 12 | C18H32O3 | 295.2276 | −0.908 | 3.5 | 251.2379 (100) 185.1183 (38) 151.1128 (85) |
| 22 | 13.70 | Hydroxy-octadecadienoic acid isomer 12 | C18H32O3 | 295.2276 | −0.908 | 3.5 | 277.2171 (31) 251.2327 (100) 155.1440 (78) |
| 23 | 14.64 | Hydroxyeicosadie-noic acid | C20H36O3 | 323.2593 | 0.408 | 3.5 | 305.2488 (22) 279.2692 (40) 179.1441 (51) |
| 24 | 14.75 | Unknown compound | C26H40O6 | 447.2745 | −1.369 | 7.5 | - |
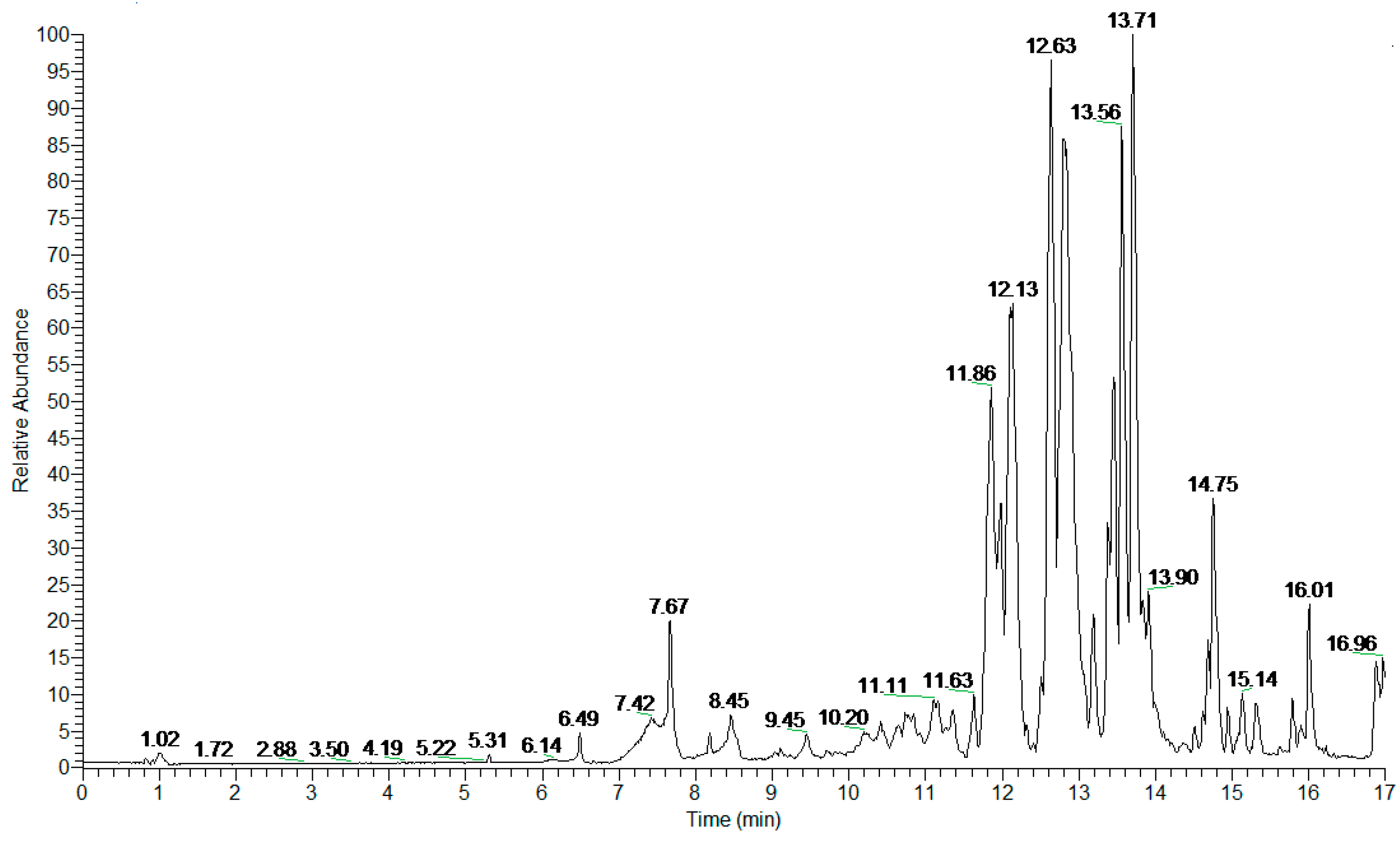
| Peak Number | Retention Time (min) | Proposed Compound | Molecular Formula, Elemental Composition | m/z Detected [M—H] | Delta, Δ (ppm) | Ring and Double Bond Equivalent RDBeq. | HRMS/MS |
|---|---|---|---|---|---|---|---|
| 1 | 6.49 | Azelaic acid 13 | C9H16O4 | 187.0975 | −0.044 | 2.5 | 125.0970 (100) |
| 2 | 7.42 | Rhamnetin-3-O-pentoside 3 | C22H22O11 | 461.1084 | −1.159 | 12.5 | 315.0506 (100) 314.0429 (34) |
| 3 | 7.67 | Isorhamnetin-3-O-pentoside 3 | C22H22O11 | 461.1082 | −1.29 | 12.5 | 315.0505 (100) 314.0430 (34) |
| 4 | 8.18 | Rhamnocitrin pentoside | C22H22O10 | 445.1135 | −1.168 | 12.5 | 299.0560 (100) |
| 5 | 8.48 | Pinellic acid 4 | C18H34O5 | 329.2332 | −0.448 | 2.5 | 311.2224 (49) 229.1442 (91) 211.1337 (64) 171.1025 (100) |
| 6 | 9.45 | Hydroxy-oxo-octadecatrienoic acid isomer 5 | C18H28O4 | 307.1915 | 0.057 | 5.5 | 289.1806 (100) 260.9318 (37) 235.1337 (73) 185.1182 (43) |
| 7 | 10.20 | Linolenic acid 13-hydroperoxide 14 | C18H30O4 | 309.2070 | −0.429 | 4.5 | 291.1965 (100) |
| 8 | 10.75 | Dihydroxyoctadecadienoic acid isomer 6 | C18H32O4 | 311.2227 | −0.266 | 3.5 | 293.2120 (100) 275.2016 (6) 201.1131 (10) |
| 9 | 11.11 | Dihydroxyoctadecadienoic acid isomer 6 | C18H32O4 | 311.2227 | −0.266 | 3.5 | 293.2119 (100) 275.2015 (32) 201.1130 (35) 171.1025 (75) |
| 10 | 11.34 | Dihydroxyoctadecadienoic acid isomer 6 | C18H32O4 | 311.2227 | −0.266 | 3.5 | 293.2119 (100) 275.2015 (22) 171.1025 (45) |
| 11 | 11.63 | Thapsic acid 7 | C16H30O4 | 285.2075 | 0.061 | 2.5 | 267.1966 (67) 223.2067 (100) |
| 12 | 11.86 | Oxo-octadecadienoic acid isomer 8,9 | C18H30O3 | 293.2118 | −1.426 | 4.5 | 275.2015 (100) 235.1702 (14) 211.1319 (28) 183.1390 (13) 171.1026 (15) |
| 13 | 11.95 | Oxo-octadecadienoic acid isomer 8 | C18H30O3 | 293.2118 | −1.426 | 4.5 | 275.2013 (100) 211.1338 (19) 195.1389 (37) |
| 14 | 12.12 | α-Licanic acid (Oxo-octadecatrienoic acid) | C18H28O3 | 291.1964 | −0.577 | 5.5 | 273.1859 (99) 247.2066 (100) 219.1390 (29) 185.1182 (24) |
| 15 | 12.63 | Hydroxy-octadecadienoic acid isomer 8 | C18H32O3 | 295.2276 | −0.908 | 3.5 | 277.2170 (100) 195.1389 (41) 171.1026 (47) |
| 16 | 12.80 | Oxo-octadecadienoic acid isomer 10 | C18H30O3 | 293.2118 | −1.085 | 4.5 | 275.2015 (26) 249.2222 (100) 185.1183 (52) |
| 17 | 13.17 | Oxo-octadecadienoic acid isomer 10 | C18H30O3 | 293.2121 | −0.403 | 4.5 | 275.2015 (11) 249.2223 (12) 197.1182 (35) 185.1182 (100) |
| 18 | 13.45 | Ricinoleic acid 11 | C18H34O3 | 297.2431 | −1.407 | 2.5 | - |
| 19 | 13.56 | Hydroxy-octadecadienoic acid isomer 12 | C18H32O3 | 295.2277 | −0.569 | 3.5 | 251.2378 (100) 151.1127 (77) 141.1284 (43) 125.0907 (55) |
| 20 | 13.71 | Hydroxy-octadecadienoic acid isomer 12 | C18H32O3 | 295.2276 | −0.908 | 3.5 | 277.2170 (30) 251.2376 (100) 155.1439 (77) |
| 21 | 13.90 | Hydroxy-octadecadienoic acid isomer 12 | C18H32O3 | 295.2276 | −0.908 | 3.5 | 251.2376 (100) 155.1440 (54) 151.1127 (29) |
| 22 | 14.68 | Hydroxyeicosadienoic acid | C20H36O3 | 323.2589 | −0.830 | 3.5 | 305.2486 (27) 279.2693 (52) 179.1442 (62) |
| 23 | 14.75 | Unknown compound | C26H40O6 | 447.2745 | −1.592 | 7.5 | - |
| 24 | 15.14 | Linolenic acid 15 | C18H30O2 | 277.2173 | −0.013 | 4.5 | 275.2015 (21) 259.2066 (30) 233.2273 (100) |
| 25 | 15.69 | Unknown compound | C22H42O4 | 369.3007 | −0.902 | 2.5 | 351.2901 (33) 307.3004 (100) |
| 26 | 16.00 | Linoleic acid 15 | C18H32O2 | 279.2325 | −1.624 | 3.5 | 279.2329 (80) 261.2219 (100) |
| Substance | Twigs’ Content |
|---|---|
| Chlorophyll a + b | 0.50 ± 0.45 mg g−1 |
| Carotenoids | 0.23 ± 0.02 mg g−1 |
| Soluble sugars | 142.13 ± 2.32 mg g−1 |
| Starch | 33.62 ± 0.89 mg g−1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bampali, A.; Karoutzou, O.; Katsarou, A.; Haralampidis, K.; Skaltsounis, L.A.; Rhizopoulou, S. Functional and Qualitative Metabolic Compounds in the Twigs of the Deciduous Mistletoe Loranthus europaeus Jacq. Stresses 2024, 4, 14-27. https://doi.org/10.3390/stresses4010002
Bampali A, Karoutzou O, Katsarou A, Haralampidis K, Skaltsounis LA, Rhizopoulou S. Functional and Qualitative Metabolic Compounds in the Twigs of the Deciduous Mistletoe Loranthus europaeus Jacq. Stresses. 2024; 4(1):14-27. https://doi.org/10.3390/stresses4010002
Chicago/Turabian StyleBampali, Anthi, Olga Karoutzou, Alexandra Katsarou, Kosmas Haralampidis, Leandros A. Skaltsounis, and Sophia Rhizopoulou. 2024. "Functional and Qualitative Metabolic Compounds in the Twigs of the Deciduous Mistletoe Loranthus europaeus Jacq." Stresses 4, no. 1: 14-27. https://doi.org/10.3390/stresses4010002
APA StyleBampali, A., Karoutzou, O., Katsarou, A., Haralampidis, K., Skaltsounis, L. A., & Rhizopoulou, S. (2024). Functional and Qualitative Metabolic Compounds in the Twigs of the Deciduous Mistletoe Loranthus europaeus Jacq. Stresses, 4(1), 14-27. https://doi.org/10.3390/stresses4010002