
Elicitation of Fruit Fungi Infection and Its Protective Response to Improve the Postharvest Quality of Fruits





Abstract
1. Introduction
2. Pathogens, Their Entry, and Transmission
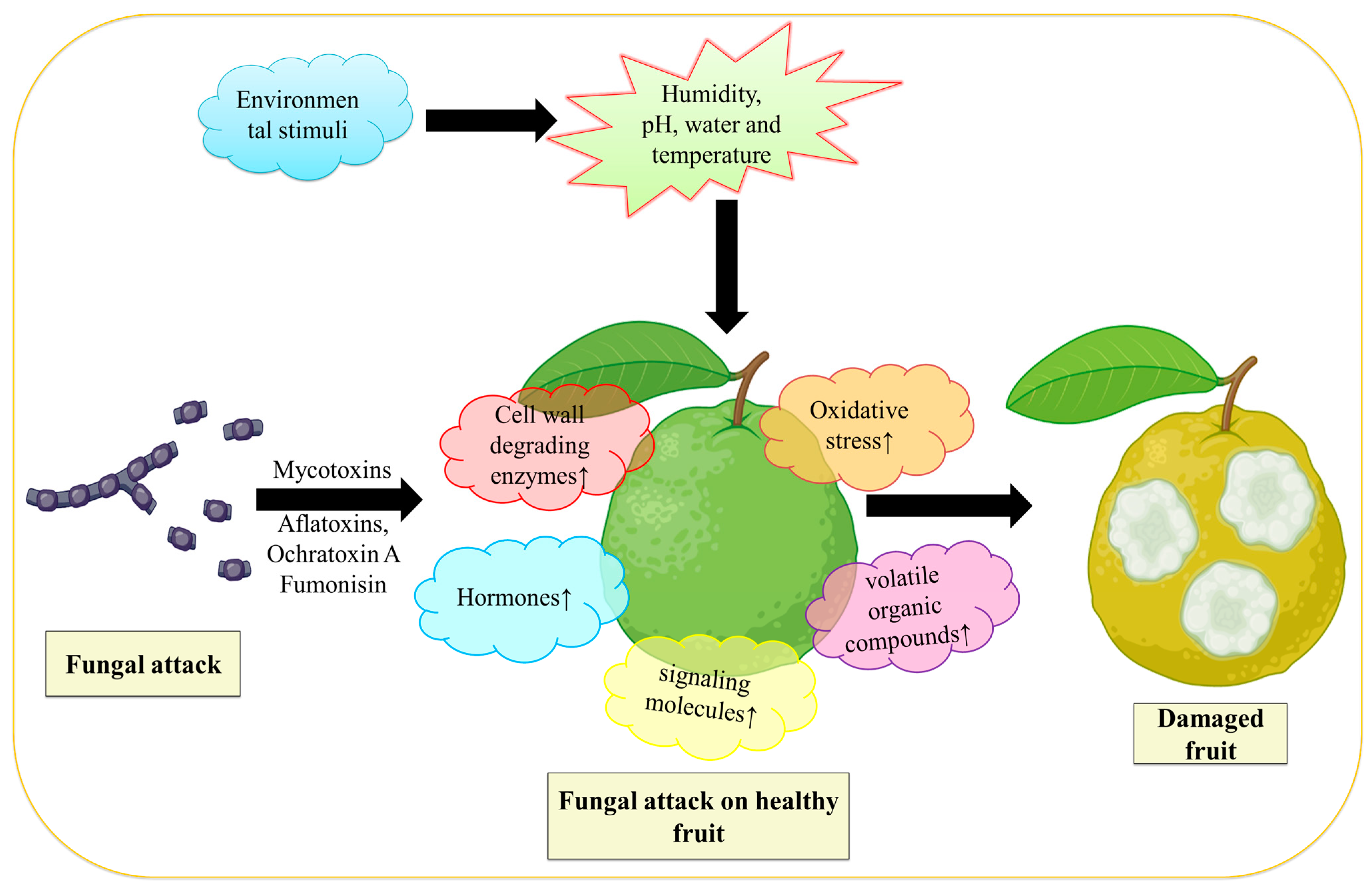
3. Postharvest Fungal Infection

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Host Crops | Species | Common Names | Symptoms | References |
|---|---|---|---|---|
| Stone fruits and Pome fruits | Alternaria spp. | Alternaria rot | Grey, black, and/or green spore colonies and sunken lesions; over-ripe softer fruits and spots on fruits. | [55] |
| Blueberries | Botrytis cinerea | Gray mold | Tissue softening. | [56] |
| Guava | Botrytis cinerea | Grey mold | Soft rotting; water-soaked parenchyma tissues occur and prolific grey conidiophores with collapsed. | [57] |
| Guava, Citrus species, pome fruits, and mango | Penicillium italicum | Blue mold | Fruit decay; green or blue mold; softens fruits and causes quick senescence. | [58] |
| Citrus | Penicillium digitatum | Green mold | Fruit decay. | [59] |
| Papaya | Stagonosporopsis caricae | Dry or Black rot | Damages fruit peel and causes fruit decay. | [60] |
| Apricots, apples, peaches, pears fruits | Monilia fructiocola | Brown rot | Brown rot; cankers; blossom blight; fruit rots. | [61] |
| Apples, mangoes, avocados, bananas, and pear fruits | Colletotrichum spp. | Anthracnose | Microbial decay; brown lesions. | [62] |
| Variety of papaya (Maradol, Golden, shahi, and Caribbean red) | Colletotrichum okinawense, C. plurivorum, C. capsici, C. truncatum, C. gloeosporioides C. magnum; and C. fructicola | Anthracnose | Brown lesions; microbial decay. | [60] |
| Papaya | Rhizopus stolonifera | Rhizopus rot | Microbial decay, brown lesions on the fruit surface. | [63] |
4. Environmental Influence on Fungal Infection
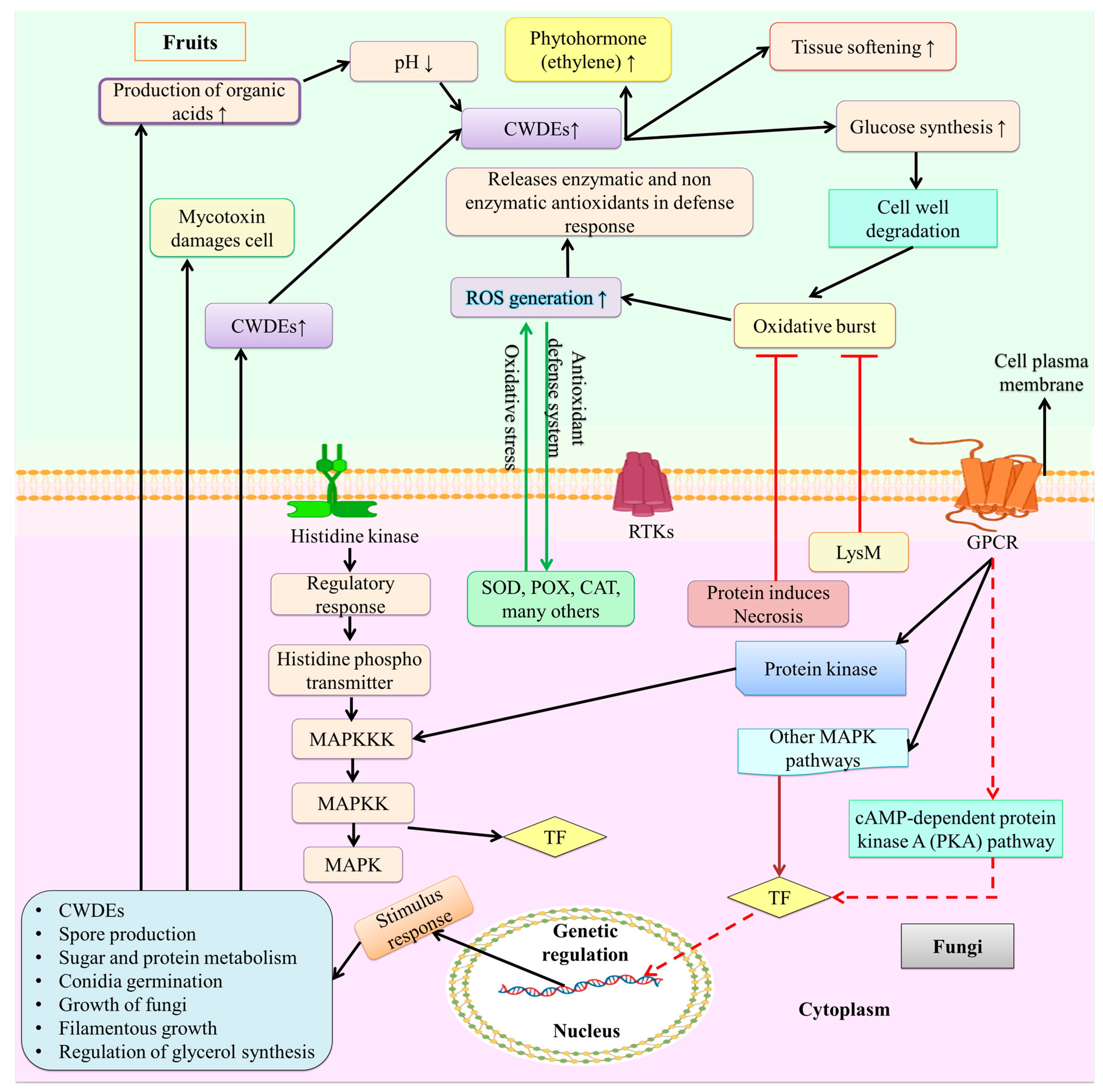
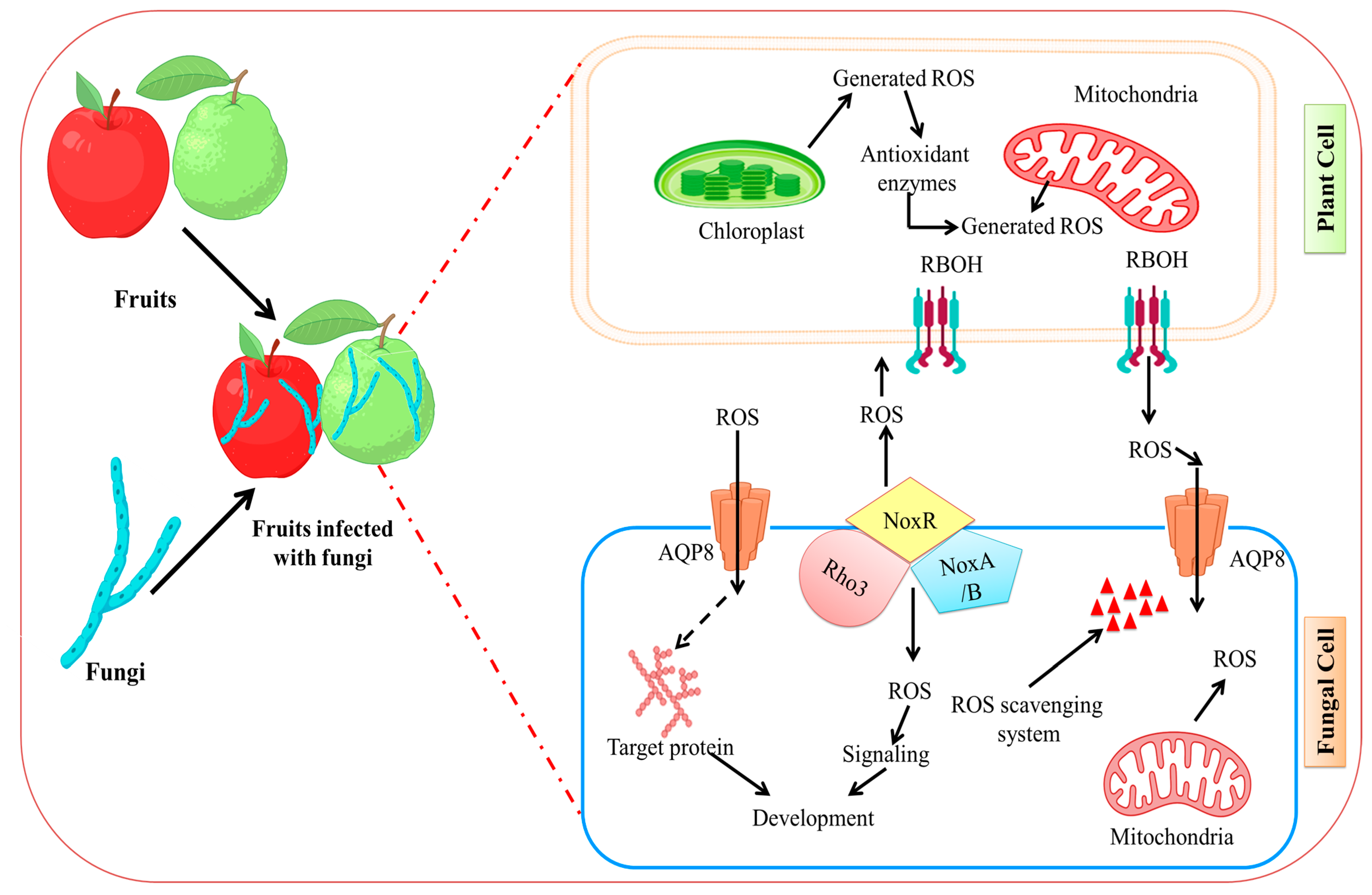
5. Fungal Infection Physicological and Physical Process
6. Molecular Mechanism of Infection
Gene Regulation
7. Modern Methods for Securing Fruit Yield
7.1. Use of Cell Wall, Membrane Degrading Enzymes, and Antifungal Proteins (AFP)
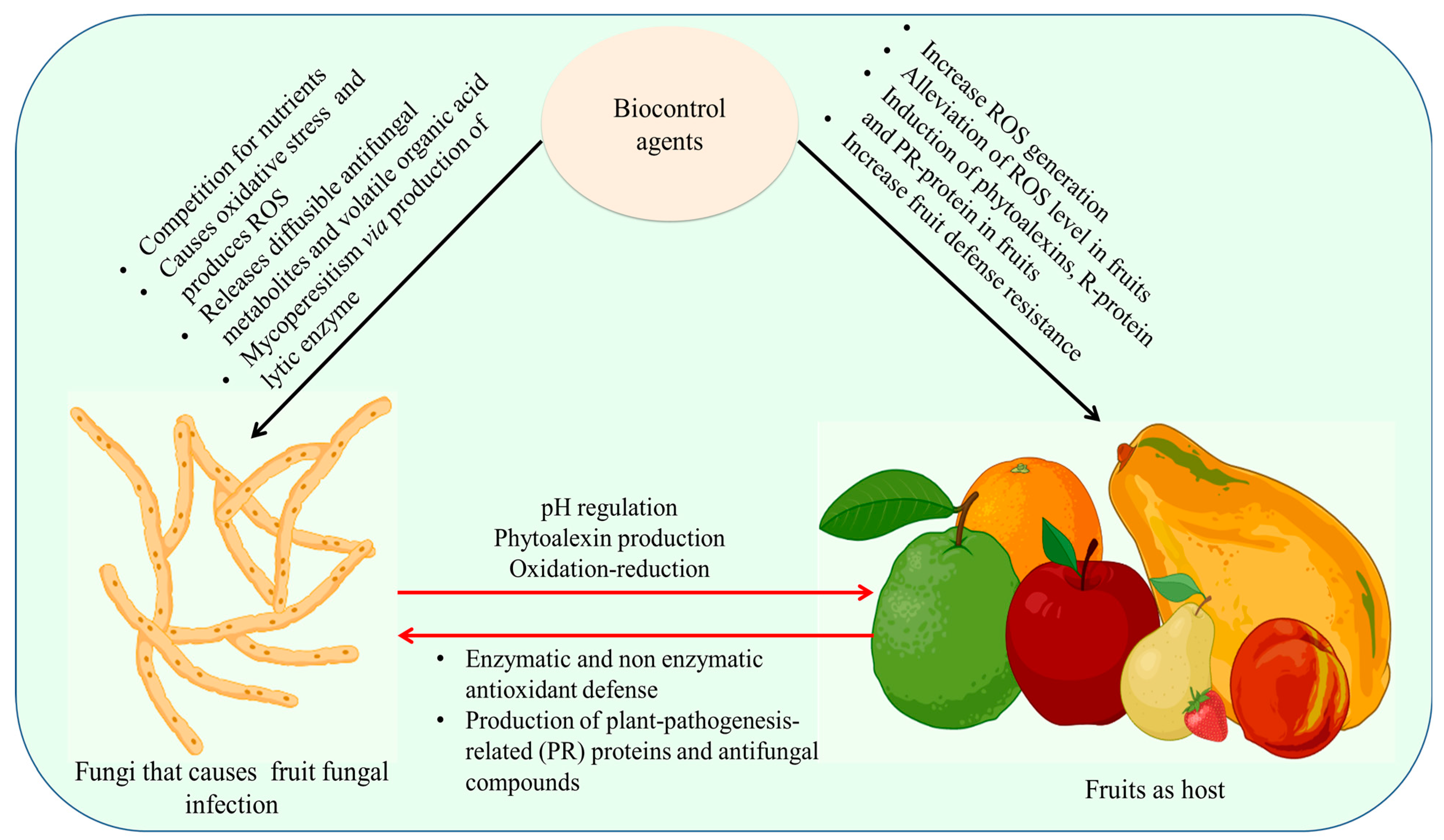
7.2. Use of Bio-Inhibitors
7.3. Use of Natural Molecules—Plant Extracts, Essential Oils, Other Solvent Extracted Plant Molecules, and Molecules of Animal Origin
7.4. Use of Hot Water Treatment (HWT)
7.5. Use of Irradiation
7.6. Use of Microbial Antagonists
8. Future Perspectives
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Semwal, P.; Painuli, S.; Jamloki, A.; Rauf, A.; Rahman, M.M.; Olatunde, A.; Hemeg, H.A.; Abu-Izneid, T.; Naz, S.; Punia Bangar, S.; et al. Himalayan Wild Fruits as a Strong Source of Nutraceuticals, Therapeutics, Food and Nutrition Security. Food Rev. Int. 2022. [Google Scholar] [CrossRef]
- Gâtlan, A.-M.; Gutt, G. Sea Buckthorn in Plant Based Diets. An Analytical Approach of Sea Buckthorn Fruits Composition: Nutritional Value, Applications, and Health Benefits. Int. J. Environ. Res. Public Health 2021, 18, 8986. [Google Scholar] [CrossRef]
- Rong, S.; Liao, Y.; Zhou, J.; Yang, W.; Yang, Y. Comparison of Dietary Guidelines among 96 Countries Worldwide. Trends Food Sci. Technol. 2021, 109, 219–229. [Google Scholar] [CrossRef]
- Jaglan, P.; Buttar, H.S.; Al-bawareed, O.A.; Chibisov, S. Potential Health Benefits of Selected Fruits: Apples, Blueberries, Grapes, Guavas, Mangos, Pomegranates, and Tomatoes. In Functional Foods and Nutraceuticals in Metabolic and Non-Communicable Diseases; Academic Press: Cambridge, MA, USA, 2022; pp. 359–370. [Google Scholar] [CrossRef]
- Vicente, A.R.; Manganaris, G.A.; Darre, M.; Ortiz, C.M.; Sozzi, G.O.; Crisosto, C.H. Compositional Determinants of Fruit and Vegetable Quality and Nutritional Value. In Postharvest Handling: A Systems Approach; Academic Press: Cambridge, MA, USA, 2022; pp. 565–619. [Google Scholar] [CrossRef]
- Da Silva, L.M.R.; de Figueiredo, E.A.T.; Ricardo, N.M.P.S.; Vieira, I.G.P.; de Figueiredo, R.W.; Brasil, I.M.; Gomes, C.L. Quantification of Bioactive Compounds in Pulps and By-Products of Tropical Fruits from Brazil. Food Chem. 2014, 143, 398–404. [Google Scholar] [CrossRef]
- World Health Organization. Food Safety. Available online: https://www.who.int/health-topics/food-safety (accessed on 29 November 2022).
- UNEP DTU Partnership and United Nations Environment Programme. Reducing Consumer Food Waste Using Green and Digital Technologies. Copenhagen and Nairobi. Copyright IEA. 2021. Available online: https://unepccc.org/wp-content/uploads/2022/03/reducing-consumer-food-waste-using-green-and-digital-technologies.pdf (accessed on 22 November 2022).
- World Health Organization. Safety Evaluation of Certain Contaminants in Food: Prepared by the Seventy-Second Meeting of the Joint FAO/WHO Expert Committee on Food Additives (JECFA); World Health Organization: Geneva, Switzerland; Food and Agriculture Organization of the United Nations: Rome, Italy, 2011. [Google Scholar]
- Pirozzi, A.; Ferrari, G.; Donsì, F. The Use of Nanocellulose in Edible Coatings for the Preservation of Perishable Fruits and Vegetables. Coatings 2021, 11, 990. [Google Scholar] [CrossRef]
- Mostafa, A.M.; Kumar, S.A.; Meraj, T.; Rauf, H.T.; Alnuaim, A.A.; Alkhayyal, M.A. Guava Disease Detection Using Deep Convolutional Neural Networks: A Case Study of Guava Plants. Appl. Sci. 2021, 12, 239. [Google Scholar] [CrossRef]
- Petrasch, S.; Silva, C.J.; Mesquida-Pesci, S.D.; Gallegos, K.; van den Abeele, C.; Papin, V.; Fernandez-Acero, F.J.; Knapp, S.J.; Blanco-Ulate, B. Infection Strategies Deployed by Botrytis cinerea, Fusarium acuminatum, and Rhizopus stolonifera as a Function of Tomato Fruit Ripening Stage. Front. Plant Sci. 2019, 10, 223. [Google Scholar] [CrossRef]
- Luciano-Rosario, D.; Keller, N.P.; Jurick, W.M., II. Penicillium expansum: Biology, Omics, and Management Tools for a Global Postharvest Pathogen Causing Blue Mould of Pome Fruit. Mol. Plant Pathol. 2020, 21, 1391–1404. [Google Scholar] [CrossRef]
- Li, X.; Yang, S.; Zhang, M.; Yang, Y.; Peng, L. Identification of Pathogenicity-Related Effector Proteins and the Role of PIWSC1 in the Virulence of Penicillium italicum on Citrus Fruits. J. Fungi 2022, 8, 646. [Google Scholar] [CrossRef]
- Liu, S.; Du, Y.; Zhang, D.; Yang, F.; He, X.; Long, C. Aluminum Sulfate Inhibits Green Mold by Inducing Chitinase Activity of Penicillium digitatum and Enzyme Activity of Citrus Fruit. Food Control 2022, 136, 108854. [Google Scholar] [CrossRef]
- Dong, H.; Cheng, L.; Tan, J.; Zheng, K.; Jiang, Y. Effects of Chitosan Coating on Quality and Shelf Life of Peeled Litchi Fruit. J. Food Eng. 2004, 64, 355–358. [Google Scholar] [CrossRef]
- Nath, V.; Lal, N.; Singh, S.K.; Pandey, S.; Prakash, K. Seventy Five Years of Research and Development in Litchi. Int. J. Innov. Hortic. 2022, 11, 47–61. [Google Scholar] [CrossRef]
- Johansen, K.S. Lytic Polysaccharide Monooxygenases: The Microbial Power Tool for Lignocellulose Degradation. Trends Plant Sci. 2016, 21, 926–936. [Google Scholar] [CrossRef]
- Wu, Y.; Yin, C.; Huang, R.; He, M.; Duan, X.; Jiang, Y.; Li, T. Enhanced Resistance in ‘Shatang’ Mandarin Fruit against Penicillium italicum Caused by 2-Methoxy-1,4-Naphthoquinone. Physiol. Mol. Plant Pathol. 2022, 119, 101828. [Google Scholar] [CrossRef]
- Tang, N.; Chen, N.; Hu, N.; Deng, W.; Chen, Z.; Li, Z. Comparative Metabolomics and Transcriptomic Profiling Reveal the Mechanism of Fruit Quality Deterioration and the Resistance of Citrus Fruit against Penicillium digitatum. Postharvest Biol. Technol. 2018, 145, 61–73. [Google Scholar] [CrossRef]
- Graça, A.; Esteves, E.; Nunes, C.; Abadias, M.; Quintas, C. Microbiological Quality and Safety of Minimally Processed Fruits in the Marketplace of Southern Portugal. Food Control 2017, 73, 775–783. [Google Scholar] [CrossRef]
- Rodrigues, M.H.P.; Furlong, E.B. Fungal Diseases and Natural Defense Mechanisms of Tomatoes (Solanum lycopersicum): A Review. Physiol. Mol. Plant Pathol. 2022, 122, 101906. [Google Scholar] [CrossRef]
- Mushtaq, M. Extraction of Fruit Juice: An Overview. In Fruit Juices: Extraction, Composition, Quality and Analysis; Academic Press: Cambridge, MA, USA, 2018; pp. 131–159. [Google Scholar] [CrossRef]
- Mahato, D.K.; Kamle, M.; Sharma, B.; Pandhi, S.; Devi, S.; Dhawan, K.; Selvakumar, R.; Mishra, D.; Kumar, A.; Arora, S.; et al. Patulin in Food: A Mycotoxin Concern for Human Health and Its Management Strategies. Toxicon 2021, 198, 12–23. [Google Scholar] [CrossRef]
- Taniwaki, M.H.; Pitt, J.I.; Magan, N. Aspergillus Species and Mycotoxins: Occurrence and Importance in Major Food Commodities. Curr. Opin. Food Sci. 2018, 23, 38–43. [Google Scholar] [CrossRef]
- Marwa, A.Y.; Embaby, E.M.; Abeer, A.F. Control of the Toxigenic Fungi Affecting Fig Fruits Quality. Egypt. J. Chem. 2022, 65, 339–347. [Google Scholar] [CrossRef]
- Wang, Y.; Nie, J.; Yan, Z.; Li, Z.; Cheng, Y.; Farooq, S. Multi-Mycotoxin Exposure and Risk Assessments for Chinese Consumption of Nuts and Dried Fruits. J. Integr. Agric. 2018, 17, 1676–1690. [Google Scholar] [CrossRef]
- Asghar, M.A.; Ahmed, F.; Kamal, M.; Khan, S.; Aghar, M.A. Effectiveness of Citrus Fruit Peel as a Biosorbent for the Mitigation of Aflatoxins In Vitro. Food Addit. Contam. Part A 2022, 39, 1987–2001. [Google Scholar] [CrossRef]
- Nikolchina, I.; Rodrigues, P. A Preliminary Study on Mycobiota and Ochratoxin a Contamination in Commercial Palm Dates (Phoenix dactylifera). Mycotoxin Res. 2021, 37, 215–220. [Google Scholar] [CrossRef]
- The Top Food Loss and Waste Statistics of 2022. Available online: https://www.postharvest.com/blog/top-food-waste-statistics-of-2021/ (accessed on 22 November 2022).
- Alkan, N.; Fortes, A.M. Insights into Molecular and Metabolic Events Associated with Fruit Response to Post-Harvest Fungal Pathogens. Front. Plant Sci. 2015, 6, 889. [Google Scholar] [CrossRef]
- Cesarino, I. Killing Me Softly: A Pathogen Accelerates Fruit Ripening and Softening to Cause Disease. Plant Physiol. 2022, 191, 21–23. [Google Scholar] [CrossRef]
- Welke, J.E. Fungal and Mycotoxin Problems in Grape Juice and Wine Industries. Curr. Opin. Food Sci. 2019, 29, 7–13. [Google Scholar] [CrossRef]
- Filippovich, S.Y.; Bachurina, G.P. Antifungal Surfaces. Appl. Biochem. Microbiol. 2022, 58, 507–517. [Google Scholar] [CrossRef]
- Li, D.; Zhang, X.; Gu, X.; Zhang, Q.; Zhao, L.; Zheng, X.; Zhang, H. The Infection of Grapes by Talaromyces rugulosus O1 and the Role of Cell Wall-Degrading Enzymes and Ochratoxin A in the Infection. Physiol. Mol. Plant Pathol. 2019, 106, 263–269. [Google Scholar] [CrossRef]
- Apaliya, M.T.; Zhang, H.; Yang, Q.; Zheng, X.; Zhao, L.; Kwaw, E.; Mahunu, G.K. Hanseniaspora uvarum Enhanced with Trehalose Induced Defense-Related Enzyme Activities and Relative Genes Expression Levels against Aspergillus tubingensis in Table Grapes. Postharvest Biol. Technol. 2017, 132, 162–170. [Google Scholar] [CrossRef]
- Wenneker, M.; Thomma, B.P.H.J. Latent Postharvest Pathogens of Pome Fruit and Their Management: From Single Measures to a Systems Intervention Approach. Eur. J. Plant Pathol. 2020, 156, 663–681. [Google Scholar] [CrossRef]
- Arya, G.C.; Sarkar, S.; Manasherova, E.; Aharoni, A.; Cohen, H. The Plant Cuticle: An Ancient Guardian Barrier Set Against Long-Standing Rivals. Front. Plant Sci. 2021, 12, 663165. [Google Scholar] [CrossRef] [PubMed]
- Esterio, M.; Osorio-Navarro, C.; Carreras, C.; Azócar, M.; Copier, C.; Estrada, V.; Rubilar, M.; Auger, J. Botrytis prunorum Associated to Vitis vinifera Blossom Blight in Chile. Plant Dis. 2020, 104, 2324–2329. [Google Scholar] [CrossRef] [PubMed]
- Machota, R., Jr.; Bortoli, L.C.; Cavalcanti, F.R.; Botton, M.; Grützmacher, A.D. Assessment of Injuries Caused by Anastrephafraterculus (Wied.) (Diptera: Tephritidae) on the Incidence of Bunch Rot Diseases in Table Grape. Neotrop. Entomol. 2016, 45, 361–368. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Muñoz, R.; Heredia, A.; Domínguez, E. The Role of Cuticle in Fruit Shelf-Life. Curr. Opin. Biotechnol. 2022, 78, 102802. [Google Scholar] [CrossRef] [PubMed]
- Jianying, F.; Bianyu, Y.; Xin, L.; Dong, T.; Weisong, M. Evaluation on Risks of Sustainable Supply Chain Based on Optimized BP Neural Networks in Fresh Grape Industry. Comput. Electron. Agric. 2021, 183, 105988. [Google Scholar] [CrossRef]
- Zhao, P.; Ndayambaje, J.P.; Liu, X.; Xia, X. Microbial Spoilage of Fruits: A Review on Causes and Prevention Methods. Food Rev. Int. 2020, 38, 225–246. [Google Scholar] [CrossRef]
- Kankam, F.; Larbi-Koranteng, S.; Adomako, J.; Kwowura Kwodaga, J.; Boatey Akpatsu, I.; Danso, Y.; Nortaa Kunedeb Sowley, E. Anthracnose Disease of Mango: Epidemiology, Impact and Management Options. In Current and Emerging Challenges in the Diseases of Trees [Working Title]; InTech Open: London, UK, 2022. [Google Scholar] [CrossRef]
- Strano, M.C.; Altieri, G.; Allegra, M.; Di Renzo, G.C.; Paterna, G.; Matera, A.; Genovese, F. Postharvest Technologies of Fresh Citrus Fruit: Advances and Recent Developments for the Loss Reduction during Handling and Storage. Horticulturae 2022, 8, 612. [Google Scholar] [CrossRef]
- Prusky, D.; Alkan, N.; Mengiste, T.; Fluhr, R. Quiescent and Necrotrophic Lifestyle Choice during Postharvest Disease Development. Annu. Rev. Phytopathol. 2013, 51, 155–176. [Google Scholar] [CrossRef]
- Diskin, S.; Feygenberg, O.; Maurer, D.; Droby, S.; Prusky, D.; Alkan, N. Microbiome Alterations Are Correlated with Occurrence of Postharvest Stem-End Rot in Mango Fruit. Phytobiomes J. 2017, 1, 117–127. [Google Scholar] [CrossRef]
- Moreira, V.; Ferronato, B.; de Benedetti, F.; González-Barrios, P.; Mondino, P.; Alaniz, S. Incidence of Colletotrichum Latent Infections during Olive Fruit Development under Uruguayan Environmental Conditions. Int. J. Pest Manag. 2022, 68, 286–294. [Google Scholar] [CrossRef]
- Rhouma, A.; Hajji-Hedfi, L.; Ben Othmen, S.; Kumari Shah, K.; Matrood, A.A.A.; Okon, O.G.; Pant, D. Strawberry Grey Mould, a Devastating Disease Caused by the Airborne Fungal Pathogen Botrytis cinerea. Egypt. J. Phytopathol. 2022, 50, 44–50. [Google Scholar] [CrossRef]
- Navarro, B.L.; Edwards Molina, J.P.; Nogueira, A.F., Jr. Penetration by Botryosphaeriaceae Species in Avocado, Guava and Persimmon Fruit during Postharvest. J. Phytopathol. 2021, 170, 57–68. [Google Scholar] [CrossRef]
- Embaby, E.; Hassan, M.K. Decay of guava fruit (Psidium guajava Linn.) quality caused by some mold fungi. Int. J. Agric. Technol. 2015, 11, 713–730. [Google Scholar]
- Alkan, N.; Friedlander, G.; Ment, D.; Prusky, D.; Fluhr, R. Simultaneous transcriptome analysis of Colletotrichum gloeosporioides and tomato fruit pathosystem reveals novel fungal pathogenicity and fruit defense strategies. New Phytol. 2014, 205, 801–815. [Google Scholar] [CrossRef] [PubMed]
- Williamson, B.; Tudzynski, B.; Tudzynski, P.; Van Kan, J.A.L. Botrytis cinerea: The Cause of Grey Mould Disease. Mol. Plant Pathol. 2007, 8, 561–580. [Google Scholar] [CrossRef] [PubMed]
- Cantu, D.; Vicente, A.R.; Greve, L.C.; Dewey, F.M.; Bennett, A.B.; Labavitch, J.M.; Powell, A.L.T. The Intersection between Cell Wall Disassembly, Ripening, and Fruit Susceptibility to Botrytis cinerea. Proc. Natl. Acad. Sci. USA 2008, 105, 859–864. [Google Scholar] [CrossRef]
- Chen, D.; Förster, H.; Adaskaveg, J.E. Baseline Sensitivities of Major Citrus, Pome, and Stone Fruits Postharvest Pathogens to Natamycin and Estimation of the Resistance Potential in Penicillium digitatum. Plant Dis. 2021, 105, 2114–2121. [Google Scholar] [CrossRef]
- Saito, S.; Wang, F.; Xiao, C.-L. Natamycin as a Postharvest Treatment to Control Gray Mold on Stored Blueberry Fruit Caused by Multi-Fungicide Resistant Botrytis cinerea. Postharvest Biol. Technol. 2022, 187, 111862. [Google Scholar] [CrossRef]
- Abdel-Rahim, I.R.; Abo-Elyousr, K.A.M. Using of Endophytic Saccharomycopsis fibuligera and Thyme Oil for Management of Gray Mold Rot of Guava Fruits. Biol. Control 2017, 110, 124–131. [Google Scholar] [CrossRef]
- Schirra, M.; D’Aquino, S.; Cabras, P.; Angioni, A. Control of Postharvest Diseases of Fruit by Heat and Fungicides: Efficacy, Residue Levels, and Residue Persistence. A Review. J. Agric. Food Chem. 2011, 59, 8531–8542. [Google Scholar] [CrossRef]
- Papoutsis, K.; Mathioudakis, M.M.; Hasperué, J.H.; Ziogas, V. Non-Chemical Treatments for Preventing the Postharvest Fungal Rotting of Citrus Caused by Penicillium digitatum (Green Mold) and Penicillium italicum (Blue Mold). Trends Food Sci. Technol. 2019, 86, 479–491. [Google Scholar] [CrossRef]
- Rodrigues, J.P.; de Souza Coelho, C.C.; Soares, A.G.; Freitas-Silva, O. Current Technologies to Control Fungal Diseases in Postharvest Papaya (Carica papaya L.). Biocatal. Agric. Biotechnol. 2021, 36, 102128. [Google Scholar] [CrossRef]
- Yin, L.; Zhang, S.; Du, J.; Wang, X.; Xu, W.; Luo, C. Monilinia fructicola on Loquat: An Old Pathogen Invading a New Host. J. Integr. Agric. 2021, 20, 2009–2014. [Google Scholar] [CrossRef]
- Ciofini, A.; Negrini, F.; Baroncelli, R.; Baraldi, E. Management of Post-Harvest Anthracnose: Current Approaches and Future Perspectives. Plants 2022, 11, 1856. [Google Scholar] [CrossRef]
- Cruz-Lachica, I.; Marquez-Zequera, I.; Allende-Molar, R.; Sañudo-Barajas, J.A.; Leon-Felix, J.; Ley-Lopez, N.; Garcia-Estrada, R.S. Diversity of Mucoralean Fungi in Soils of Papaya (Carica papaya L.) Producing Regions in Mexico. Fungal Biol. 2018, 122, 810–816. [Google Scholar] [CrossRef]
- Ciliberti, N.; Fermaud, M.; Roudet, J.; Languasco, L.; Rossi, V. Environmental Effects on the Production of Botrytis cinerea Conidia on Different Media, Grape Bunch Trash, and Mature Berries. Aust. J. Grape Wine Res. 2016, 22, 262–270. [Google Scholar] [CrossRef]
- Jiao, W.; Liu, X.; Li, Y.; Li, B.; Du, Y.; Zhang, Z.; Chen, Q.; Fu, M. Organic Acid, a Virulence Factor for Pathogenic Fungi, Causing Postharvest Decay in Fruits. Mol. Plant Pathol. 2021, 23, 304–312. [Google Scholar] [CrossRef]
- Li, H.; Zhang, Y.; Gao, C.; Gao, Q.; Cheng, Y.; Zhao, M.; Guan, J. Mycotoxin Production and the Relationship between Microbial Diversity and Mycotoxins in Pyrus bretschneideri Rehd Cv. Huangguan Pear. Toxins 2022, 14, 699. [Google Scholar] [CrossRef]
- He, Y.; Cox, R.J. The Molecular Steps of Citrinin Biosynthesis in Fungi. Chem. Sci. 2016, 7, 2119–2127. [Google Scholar] [CrossRef]
- Costa, J.; Rodríguez, R.; Garcia-Cela, E.; Medina, A.; Magan, N.; Lima, N.; Battilani, P.; Santos, C. Overview of Fungi and Mycotoxin Contamination in Capsicum Pepper and in Its Derivatives. Toxins 2019, 11, 27. [Google Scholar] [CrossRef]
- Caceres, I.; Al Khoury, A.; El Khoury, R.; Lorber, S.; Oswald, I.P.; El Khoury, A.; Atoui, A.; Puel, O.; Bailly, J.-D. Aflatoxin Biosynthesis and Genetic Regulation: A Review. Toxins 2020, 12, 150. [Google Scholar] [CrossRef]
- Ghuffar, S.; Irshad, G.; Ahmed, M.Z.; Zeshan, M.A.; Ali, R.; ul Haq, E.; Anwaar, H.A.; Abdullah, A.; Ahmad, F.; Haque, K. First Report of Aspergillus flavus Causing Fruit Rot of Grapes (Vitis vinifera) in Pakistan. Plant Dis. 2020, 104, 3062. [Google Scholar] [CrossRef]
- Troncoso-Rojas, R.; Tiznado-Hernández, M.E. Alternaria alternata (Black Rot, Black Spot). In Postharvest Decay: Control Strategies; Academic Press: Cambridge, MA, USA, 2014; pp. 147–187. [Google Scholar] [CrossRef]
- Wenderoth, M.; Garganese, F.; Schmidt-Heydt, M.; Soukup, S.T.; Ippolito, A.; Sanzani, S.M.; Fischer, R. Alternariol as Virulence and Colonization Factor of Alternaria alternata during Plant Infection. Mol. Microbiol. 2019, 112, 131–146. [Google Scholar] [CrossRef] [PubMed]
- Jurick, W.M., II; Kou, L.P.; Gaskins, V.L.; Luo, Y.G. First Report of Alternaria alternata Causing Postharvest Decay on Apple Fruit During Cold Storage in Pennsylvania. Plant Dis. 2014, 98, 690. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, T.; Liu, Y.; Shujian, H.; Moosa, A. First Record of Alternaria alternata Causing Postharvest Fruit Rot of Sweet Cherry (Prunus avium) in China. Plant Dis. 2020, 104, 2030. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, L.; Wu, F.; Liu, F.; Wang, Q.; Zhang, X.; Selvaraj, J.N.; Zhao, Y.; Xing, F.; Yin, W.-B.; et al. A Consensus Ochratoxin A Biosynthetic Pathway: Insights from the Genome Sequence of Aspergillus ochraceus and a Comparative Genomic Analysis. Appl. Environ. Microbiol. 2018, 84, e01009-18. [Google Scholar] [CrossRef] [PubMed]
- Tannous, J.; El Khoury, R.; Snini, S.P.; Lippi, Y.; El Khoury, A.; Atoui, A.; Lteif, R.; Oswald, I.P.; Puel, O. Sequencing, Physical Organization and Kinetic Expression of the Patulin Biosynthetic Gene Cluster from Penicillium expansum. Int. J. Food Microbiol. 2014, 189, 51–60. [Google Scholar] [CrossRef]
- Jurick, W.M., II; Peng, H.; Beard, H.S.; Garrett, W.M.; Lichtner, F.J.; Luciano-Rosario, D.; Macarisin, O.; Liu, Y.; Peter, K.A.; Gaskins, V.L.; et al. Blistering1 Modulates Penicillium expansum Virulence Via Vesicle-Mediated Protein Secretion. Mol. Cell. Proteom. 2020, 19, 344–361. [Google Scholar] [CrossRef]
- Brito, A.C.Q.; Mello, J.F.; Vieira, J.C.B.; Câmara, M.P.S.; Bezerra, J.D.P.; Souza-Motta, C.M.; Machado, A.R. First Report of Penicillium expansum Causing Postharvest Fruit Rot on Black Plum (Prunus domestica) in Brazil. Plant Dis. 2020, 104, 576. [Google Scholar] [CrossRef]
- Ballester, A.-R.; Marcet-Houben, M.; Levin, E.; Sela, N.; Selma-Lázaro, C.; Carmona, L.; Wisniewski, M.; Droby, S.; González-Candelas, L.; Gabaldón, T. Genome, Transcriptome, and Functional Analyses of Penicillium expansum Provide New Insights Into Secondary Metabolism and Pathogenicity. Mol. Plant-Microbe Interact. 2015, 28, 232–248. [Google Scholar] [CrossRef]
- Alkan, N.; Espeso, E.A.; Prusky, D. Virulence Regulation of Phytopathogenic Fungi by pH. Antioxid. Redox Signal. 2013, 19, 1012–1025. [Google Scholar] [CrossRef] [PubMed]
- Rascle, C.; Dieryckx, C.; Dupuy, J.W.; Muszkieta, L.; Souibgui, E.; Droux, M.; Bruel, C.; Girard, V.; Poussereau, N. The pH Regulator PacC: A Host-Dependent Virulence Factor in Botrytis cinerea. Environ. Microbiol. Rep. 2018, 10, 555–568. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Zhao, L.; Qi, F.; Htwe, N.M.P.S.; Li, Q.; Zhang, D.; Lin, F.; Shang-Guan, K.; Liang, Y. The Receptor-like Cytoplasmic Kinase RIPK Regulates Broad-Spectrum ROS Signaling in Multiple Layers of Plant Immune System. Mol. Plant 2021, 14, 1652–1667. [Google Scholar] [CrossRef] [PubMed]
- Barda, O.; Maor, U.; Sadhasivam, S.; Bi, Y.; Zakin, V.; Prusky, D.; Sionov, E. The pH-Responsive Transcription Factor PacC Governs Pathogenicity and Ochratoxin A Biosynthesis in Aspergillus carbonarius. Front. Microbiol. 2020, 11, 210. [Google Scholar] [CrossRef]
- Maor, U.; Sadhasivam, S.; Zakin, V.; Prusky, D.; Sionov, E. The Effect of Ambient pH Modulation on Ochratoxin A Accumulation by Aspergillus carbonarius. World Mycotoxin J. 2017, 10, 339–348. [Google Scholar] [CrossRef]
- Martínez-Bracero, M.; Alcázar, P.; Velasco-Jiménez, M.J.; Galán, C. Fungal Spores Affecting Vineyards in Montilla-Moriles Southern Spain. Eur. J. Plant Pathol. 2018, 153, 1–13. [Google Scholar] [CrossRef]
- Steel, C.C.; Greer, L.A.; Savocchia, S. Studies on Colletotrichum acutatum and Greeneria uvicola: Two Fungi Associated with Bunch Rot of Grapes in Sub-Tropical Australia. Aust. J. Grape Wine Res. 2007, 13, 23–29. [Google Scholar] [CrossRef]
- Makule, E.; Dimoso, N.; Tassou, S.A. Precooling and Cold Storage Methods for Fruits and Vegetables in Sub-Saharan Africa—A Review. Horticulturae 2022, 8, 776. [Google Scholar] [CrossRef]
- Yi, T.; Hong, Y.; Li, M.; Li, X. Examination and Characterization of Key Factors to Seasonal Epidemics of Downy Mildew in Native Grape Species Vitis davidii in Southern China with the use of Disease Warning Systems. Eur. J. Plant Pathol. 2019, 154, 389–404. [Google Scholar] [CrossRef]
- Amiri, A.; Mortazavi, S.M.H.; Ramezanian, A.; Mahmoodi Sourestani, M.; Mottaghipisheh, J.; Iriti, M.; Vitalini, S. Prevention of Decay and Maintenance of Bioactive Compounds in Strawberry by Application of UV-C and Essential Oils. J. Food Meas. Charact. 2021, 15, 5310–5317. [Google Scholar] [CrossRef]
- Pinto, J.A.V.; Schorr, M.R.W.; Thewes, F.R.; Ceconi, D.L.; Both, V.; Brackmann, A.; Fronza, D. Relative Humidity during Cold Storage on Postharvest Quality of “Niagara Rosada” Table Grapes. Ciênc. Rural 2015, 45, 386–391. [Google Scholar] [CrossRef]
- Igo, M.J.; Schaffner, D.W. Models for Factors Influencing Pathogen Survival in Low Water Activity Foods from Literature Data Are Highly Significant but Show Large Unexplained Variance. Food Microbiol. 2021, 98, 103783. [Google Scholar] [CrossRef]
- Kogkaki, E.A.; Natskoulis, P.I.; Magan, N.; Panagou, E.Z. Effect of Interaction between Aspergillus carbonarius and Non-Ochratoxigenic Grape-Associated Fungal Isolates on Growth and Ochratoxin A Production at Different Water Activities and Temperatures. Food Microbiol. 2015, 46, 521–527. [Google Scholar] [CrossRef]
- Van Kan, J.A.L. Licensed to Kill: The Lifestyle of a Necrotrophic Plant Pathogen. Trends Plant Sci. 2006, 11, 247–253. [Google Scholar] [CrossRef]
- Jiang, B.; Liu, R.; Fang, X.; Wu, W.; Han, Y.; Chen, H.; Xu, F.; Gao, H. Botrytis cinerea Infection Affects Wax Composition, Content and Gene Expression in Blueberry Fruit. Postharvest Biol. Technol. 2022, 192, 112020. [Google Scholar] [CrossRef]
- Choquer, M.; Fournier, E.; Kunz, C.; Levis, C.; Pradier, J.-M.; Simon, A.; Viaud, M. Botrytis cinerea Virulence Factors: New Insights into a Necrotrophic and Polyphageous Pathogen. FEMS Microbiol. Lett. 2007, 277, 1–10. [Google Scholar] [CrossRef]
- Armijo, G.; Schlechter, R.; Agurto, M.; Muñoz, D.; Nuñez, C.; Arce-Johnson, P. Grapevine Pathogenic Microorganisms: Understanding Infection Strategies and Host Response Scenarios. Front. Plant Sci. 2016, 7, 382. [Google Scholar] [CrossRef]
- Vilanova, L.; Vall-llaura, N.; Torres, R.; Usall, J.; Teixidó, N.; Larrigaudière, C.; Giné-Bordonaba, J. Penicillium expansum (Compatible) and Penicillium digitatum (Non-Host) Pathogen Infection Differentially alter Ethylene Biosynthesis in Apple Fruit. Plant Physiol. Biochem. 2017, 120, 132–143. [Google Scholar] [CrossRef]
- Deng, B.; Wang, W.; Deng, L.; Yao, S.; Ming, J.; Zeng, K. Comparative RNA-Seq Analysis of Citrus Fruit in Response to Infection with Three Major Postharvest Fungi. Postharvest Biol. Technol. 2018, 146, 134–146. [Google Scholar] [CrossRef]
- Lorrai, R.; Ferrari, S. Host Cell Wall Damage during Pathogen Infection: Mechanisms of Perception and Role in Plant-Pathogen Interactions. Plants 2021, 10, 399. [Google Scholar] [CrossRef]
- Xu, D.; Deng, Y.; Han, T.; Jiang, L.; Xi, P.; Wang, Q.; Jiang, Z.; Gao, L. In Vitro and in Vivo Effectiveness of Phenolic Compounds for the Control of Postharvest Gray Mold of Table Grapes. Postharvest Biol. Technol. 2018, 139, 106–114. [Google Scholar] [CrossRef]
- Kong, Q.; An, P.; Xu, Z.; Zhang, R.; Qi, J.; Ren, X. New Insights into the Alleviating Role of Melaleuca alternifolia Oil on Metabolites Pathway Disorder of Grapes Caused by Aspergillus niger, Verified by Corresponding Key Genes Expression. Food Chem. 2020, 327, 127083. [Google Scholar] [CrossRef]
- Kupfer, V.M.; Vogt, E.I.; Ziegler, T.; Vogel, R.F.; Niessen, L. Comparative Protein Profile Analysis of Wines Made from Botrytis cinerea Infected and Healthy Grapes Reveals a Novel Biomarker for Gushing in Sparkling Wine. Food Res. Int. 2017, 99, 501–509. [Google Scholar] [CrossRef]
- Crespo-Sempere, A.; Marín, S.; Sanchis, V.; Ramos, A.J. VeA and LaeA Transcriptional Factors Regulate Ochratoxin A Biosynthesis in Aspergillus carbonarius. Int. J. Food Microbiol. 2013, 166, 479–486. [Google Scholar] [CrossRef]
- Zhu, P.; Xu, L.; Zhang, C.; Toyoda, H.; Gan, S.-S. Ethylene Produced by Botrytis cinerea Can Affect Early Fungal Development and Can Be Used as a Marker for Infection during Storage of Grapes. Postharvest Biol. Technol. 2012, 66, 23–29. [Google Scholar] [CrossRef]
- Martínez-Soto, D.; Ruiz-Herrera, J. Functional Analysis of the MAPK Pathways in Fungi. Rev. Iberoam. Micol. 2017, 34, 192–202. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Zhang, X.; Liu, H.; Xu, J.-R. Mitogen-Activated Protein Kinase Signaling in Plant Pathogenic Fungi. PLoS Pathog. 2018, 14, e1006875. [Google Scholar] [CrossRef]
- Zhao, X.; Mehrabi, R.; Xu, J.-R. Mitogen-Activated Protein Kinase Pathways and Fungal Pathogenesis. Eukaryot. Cell 2007, 6, 1701–1714. [Google Scholar] [CrossRef]
- Hedbacker, K. SNF1/AMPK Pathways in Yeast. Front. Biosci. 2008, 13, 2408. [Google Scholar] [CrossRef]
- Zhang, T.; Xu, Q.; Sun, X.; Li, H. The Calcineurin-Responsive Transcription Factor Crz1 Is Required for Conidation, Full Virulence and DMI Resistance in Penicillium digitatum. Microbiol. Res. 2013, 168, 211–222. [Google Scholar] [CrossRef]
- Bushweller, J.H. Targeting Transcription Factors in Cancer—From Undruggable to Reality. Nat. Rev. Cancer 2019, 19, 611–624. [Google Scholar] [CrossRef] [PubMed]
- Tannous, J.; Barda, O.; Luciano-Rosario, D.; Prusky, D.B.; Sionov, E.; Keller, N.P. New Insight Into Pathogenicity and Secondary Metabolism of the Plant Pathogen Penicillium expansum Through Deletion of the Epigenetic Reader SntB. Front. Microbiol. 2020, 11, 610. [Google Scholar] [CrossRef] [PubMed]
- Voß, B.; Kirschhöfer, F.; Brenner-Weiß, G.; Fischer, R. Alternaria alternata Uses Two Siderophore Systems for Iron Acquisition. Sci. Rep. 2020, 10, 3587. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.; Liu, X.; Shi, T.; Li, C.; Huang, G. The Colletotrichum gloeosporioides Perilipin Homologue CAP 20 Regulates Functional Appressorial Formation and Fungal Virulence. J. Phytopathol. 2017, 166, 216–225. [Google Scholar] [CrossRef]
- Bi, F.; Ment, D.; Luria, N.; Meng, X.; Prusky, D. Mutation of AREA Affects Growth, Sporulation, Nitrogen Regulation, and Pathogenicity in Colletotrichum gloeosporioides. Fungal Genet. Biol. 2017, 99, 29–39. [Google Scholar] [CrossRef]
- Chudasama, K.S.; Monpara, J.K.; Thaker, V.S. Identification and Characterization of Pectin Lyase Gene as a Virulence Factor in Colletotrichum gloeosporioides. Physiol. Mol. Plant Pathol. 2021, 116, 101706. [Google Scholar] [CrossRef]
- Yakoby, N.; Beno-Moualem, D.; Keen, N.T.; Dinoor, A.; Pines, O.; Prusky, D. Colletotrichum gloeosporioides pelB Is an Important Virulence Factor in Avocado Fruit-Fungus Interaction. Mol. Plant-Microbe Interact. 2001, 14, 988–995. [Google Scholar] [CrossRef]
- Yang, Q.; Chen, Y.; Ma, Z. Involvement of BcVeA and BcVelB in Regulating Conidiation, Pigmentation and Virulence in Botrytis cinerea. Fungal Genet. Biol. 2013, 50, 63–71. [Google Scholar] [CrossRef]
- Archana, S.; Raguchander, T.; Prabakar, K. Detection of -Tubulin Gene from Benomyl Sensitive Isolates of Colletotrichum gloeosporioides Causing Anthracnose Disease in Mango. Afr. J. Microbiol. Res. 2018, 12, 806–814. [Google Scholar] [CrossRef]
- Zhang, H.; Mahunu, G.K.; Castoria, R.; Yang, Q.; Apaliya, M.T. Recent Developments in the Enhancement of Some Postharvest Biocontrol Agents with Unconventional Chemicals Compounds. Trends Food Sci. Technol. 2018, 78, 180–187. [Google Scholar] [CrossRef]
- Sánchez-Torres, P.; Vilanova, L.; Ballester, A.R.; López-Pérez, M.; Teixidó, N.; Viñas, I.; Usall, J.; González-Candelas, L.; Torres, R. Unravelling the Contribution of the Penicillium expansum PeSte12 Transcription Factor to Virulence during Apple Fruit Infection. Food Microbiol. 2018, 69, 123–135. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Ngea, G.L.N.; Godana, E.A.; Shi, Y.; Lanhuang, B.; Zhang, X.; Zhao, L.; Yang, Q.; Wang, S.; Zhang, H. Recent Advances in Penicillium expansum Infection Mechanisms and Current Methods in Controlling P. expansum in Postharvest Apples. Critical Rev. Food Sci. Nutr. 2021. [Google Scholar] [CrossRef] [PubMed]
- De Ramón-Carbonell, M.; Sánchez-Torres, P. The Transcription Factor PdSte12 Contributes to Penicillium digitatum Virulence during Citrus Fruit Infection. Postharvest Biol. Technol. 2017, 125, 129–139. [Google Scholar] [CrossRef]
- Wang, M.; Chen, C.; Zhu, C.; Sun, X.; Ruan, R.; Li, H. Os2 MAP Kinase-Mediated Osmostress Tolerance in Penicillium digitatum Is Associated with Its Positive Regulation on Glycerol Synthesis and Negative Regulation on Ergosterol Synthesis. Microbiol. Res. 2014, 169, 511–521. [Google Scholar] [CrossRef]
- Vilanova, L.; López-Pérez, M.; Ballester, A.-R.; Teixidó, N.; Usall, J.; Lara, I.; Viñas, I.; Torres, R.; González-Candelas, L. Differential Contribution of the Two Major Polygalacturonases from Penicillium digitatum to Virulence towards Citrus Fruit. Int. J. Food Microbiol. 2018, 282, 16–23. [Google Scholar] [CrossRef]
- Zhu, C.; Wang, W.; Wang, M.; Ruan, R.; Sun, X.; He, M.; Mao, C.; Li, H. Deletion of PdMit1, a Homolog of Yeast Csg1, Affects Growth and Ca2+ Sensitivity of the Fungus Penicillium digitatum, but Does Not Alter Virulence. Res. Microbiol. 2015, 166, 143–152. [Google Scholar] [CrossRef]
- Wang, W.; Wang, M.; Wang, J.; Zhu, C.; Chung, K.-R.; Li, H. Adenylyl Cyclase Is Required for cAMP Production, Growth, Conidial Germination, and Virulence in the Citrus Green Mold Pathogen Penicillium digitatum. Microbiol. Res. 2016, 192, 11–20. [Google Scholar] [CrossRef]
- Ruan, R.; Wang, M.; Liu, X.; Sun, X.; Chung, K.-R.; Li, H. Functional Analysis of Two Sterol Regulatory Element Binding Proteins in Penicillium digitatum. PLoS ONE 2017, 12, e0176485. [Google Scholar] [CrossRef]
- De Ramón-Carbonell, M.; Sánchez-Torres, P. Involvement of Penicillium digitatum PdSUT1 in Fungicide Sensitivity and Virulence during Citrus Fruit Infection. Microbiol. Res. 2017, 203, 57–67. [Google Scholar] [CrossRef]
- De Ramón-Carbonell, M.; López-Pérez, M.; González-Candelas, L.; Sánchez-Torres, P. PdMFS1 Transporter Contributes to Penicilliun digitatum Fungicide Resistance and Fungal Virulence during Citrus Fruit Infection. J. Fungi 2019, 5, 100. [Google Scholar] [CrossRef]
- De Ramón-Carbonell, M.; Sánchez-Torres, P. PdSlt2 Penicillium digitatum Mitogen-Activated-Protein Kinase Controls Sporulation and Virulence during Citrus Fruit Infection. Fungal Biol. 2017, 121, 1063–1074. [Google Scholar] [CrossRef]
- Bano, A.; Gupta, A.; Rai, S.; Sharma, S.; Pathak, N. Elucidation of Bioactive Potential of Two Commonly Grown North Indian Psidium guajava Viz., Lalit and Shweta against Pathogenic Foodborne and MDR Bacteria. Biointerface Res. Appl. Chem. 2021, 11, 14090–14102. [Google Scholar] [CrossRef]
- Gupta, A.; Mishra, R.; Rai, S.; Bano, A.; Pathak, N.; Fujita, M.; Kumar, M.; Hasanuzzaman, M. Mechanistic Insights of Plant Growth Promoting Bacteria Mediated Drought and Salt Stress Tolerance in Plants for Sustainable Agriculture. Int. J. Mol. Sci. 2022, 23, 3741. [Google Scholar] [CrossRef]
- Bano, A.; Gupta, A.; Rai, S.; Fatima, T.; Sharma, S.; Pathak, N. Mechanistic Role of Reactive Oxygen Species and Its Regulation via the Antioxidant System under Environmental Stress. In Plant Stress Physiology: Perspectives in Agriculture; InTech Open: London, UK, 2022. [Google Scholar] [CrossRef]
- Gupta, A.; Bano, A.; Rai, S.; Mishra, R.; Singh, M.; Sharma, S.; Pathak, N. Mechanistic Insights of Plant-Microbe Interaction towards Drought and Salinity Stress in Plants for Enhancing the Agriculture Productivity. Plant Stress 2022, 4, 100073. [Google Scholar] [CrossRef]
- Zhang, Z.-Q.; Chen, T.; Li, B.-Q.; Qin, G.-Z.; Tian, S.-P. Molecular Basis of Pathogenesis of Postharvest Pathogenic Fungi and Control Strategy in Fruits: Progress and Prospect. Mol. Hortic. 2021, 1, 2. [Google Scholar] [CrossRef]
- Ezzat, A.; Szabó, S.; Szabó, Z.; Hegedűs, A.; Berényi, D.; Holb, I.J. Temporal Patterns and Inter-Correlations among Physical and Antioxidant Attributes and Enzyme Activities of Apricot Fruit Inoculated with Monilinia laxa under Salicylic Acid and Methyl Jasmonate Treatments under Shelf-Life Conditions. J. Fungi 2021, 7, 341. [Google Scholar] [CrossRef]
- Zhang, Z.; Chen, Y.; Li, B.; Chen, T.; Tian, S. Reactive Oxygen Species: A Generalist in Regulating Development and Pathogenicity of Phytopathogenic Fungi. Comput. Struct. Biotechnol. J. 2020, 18, 3344–3349. [Google Scholar] [CrossRef]
- Bedard, K.; Lardy, B.; Krause, K. NOX Family NADPH Oxidases: Not Just in Mammals. Biochimie 2007, 89, 1107–1112. [Google Scholar] [CrossRef]
- Segmüller, N.; Kokkelink, L.; Giesbert, S.; Odinius, D.; van Kan, J.; Tudzynski, P. NADPH Oxidases Are Involved in Differentiation and Pathogenicity in Botrytis cinerea. Mol. Plant-Microbe Interact. 2008, 21, 808–819. [Google Scholar] [CrossRef]
- Li, H.; Zhang, Z.; He, C.; Qin, G.; Tian, S. Comparative Proteomics Reveals the Potential Targets of BcNoxR, a Putative Regulatory Subunit of NADPH Oxidase of Botrytis cinerea. Mol. Plant-Microbe Interact. 2016, 29, 990–1003. [Google Scholar] [CrossRef]
- Da Silva, R.R. Enzyme Technology in Food Preservation: A Promising and Sustainable Strategy for Biocontrol of Post-Harvest Fungal Pathogens. Food Chem. 2019, 277, 531–532. [Google Scholar] [CrossRef]
- Leiter, É.; Gáll, T.; Csernoch, L.; Pócsi, I. Biofungicide Utilizations of Antifungal Proteins of Filamentous Ascomycetes: Current and Foreseeable Future Developments. BioControl 2016, 62, 125–138. [Google Scholar] [CrossRef]
- Gregori, R.; Mari, M.; Bertolini, P.; Barajas, J.A.S.; Tian, J.B.; Labavitch, J.M. Reduction of Colletotrichum acutatum Infection by a Polygalacturonase Inhibitor Protein Extracted from Apple. Postharvest Biol. Technol. 2008, 48, 309–313. [Google Scholar] [CrossRef]
- Sansone, G.; Rezza, I.; Fernández, G.; Calvente, V.; Benuzzi, D.; Sanz, M.I. Inhibitors of Polygalacturonase and Laccase of Botrytis cinerea and Their Application to the Control of This Fungus. Int. Biodeterior. Biodegrad. 2011, 65, 243–247. [Google Scholar] [CrossRef]
- Ballester, A.-R.; López-Pérez, M.; de la Fuente, B.; González-Candelas, L. Functional and Pharmacological Analyses of the Role of Penicillium digitatum Proteases on Virulence. Microorganisms 2019, 7, 198. [Google Scholar] [CrossRef]
- Hernandez Montiel, L.G.; Zulueta Rodriguez, R.; Angulo, C.; Rueda Puente, E.O.; Quiñonez Aguilar, E.E.; Galicia, R. Marine Yeasts and Bacteria as Biological Control Agents against Anthracnose on Mango. J. Phytopathol. 2017, 165, 833–840. [Google Scholar] [CrossRef]
- Aiello, D.; Restuccia, C.; Stefani, E.; Vitale, A.; Cirvilleri, G. Postharvest Biocontrol Ability of Pseudomonas synxantha against Monilinia fructicola and Monilinia fructigena on Stone Fruit. Postharvest Biol. Technol. 2019, 149, 83–89. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, Y.; Fu, X.; Li, Y.; Wang, Q. Isolation and Characterization of Bacillus amyloliquefaciens PG12 for the Biological Control of Apple Ring Rot. Postharvest Biol. Technol. 2016, 115, 113–121. [Google Scholar] [CrossRef]
- Wallace, R.L.; Hirkala, D.L.; Nelson, L.M. Mechanisms of Action of Three Isolates of Pseudomonas fluorescens Active against Postharvest Grey Mold Decay of Apple during Commercial Storage. Biol. Control 2018, 117, 13–20. [Google Scholar] [CrossRef]
- Khleekorn, S.; Wongrueng, S. Evaluation of antagonistic bacteria inhibitory to Colletotrichum musae on banana. J. Agric. Technol. 2014, 10, 383–390. [Google Scholar]
- Fu, G.; Huang, S.; Ye, Y.; Wu, Y.; Cen, Z.; Lin, S. Characterization of a Bacterial Biocontrol Strain B106 and Its Efficacies on Controlling Banana Leaf Spot and Post-Harvest Anthracnose Diseases. Biol. Control 2010, 55, 1–10. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, Z.; Zhu, P.; Liu, Y.; Zhang, Z.; Mastuda, Y.; Toyoda, H.; Xu, L. Postharvest biological control of melon pathogens using Bacillus subtilis EXWB1. J. Plant Pathol. 2010, 92, 645–652. [Google Scholar]
- Kasfi, K.; Taheri, P.; Jafarpour, B.; Tarighi, S. Identification of Epiphytic Yeasts and Bacteria with Potential for Biocontrol of Grey Mold Disease on Table Grapes Caused by Botrytis cinerea. Span. J. Agric. Res. 2018, 16, e1002. [Google Scholar] [CrossRef]
- Kim, Y.S.; Balaraju, K.; Jeon, Y. Effects of Rhizobacteria Paenibacillus polymyxa APEC136 and Bacillus subtilis APEC170 on Biocontrol of Postharvest Pathogens of Apple Fruits. J. Zhejiang Univ. Sci. B 2016, 17, 931–940. [Google Scholar] [CrossRef] [PubMed]
- Calvo, H.; Marco, P.; Blanco, D.; Oria, R.; Venturini, M.E. Potential of a New Strain of Bacillus amyloliquefaciens BUZ-14 as a Biocontrol Agent of Postharvest Fruit Diseases. Food Microbiol. 2017, 63, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Wu, Y.; Li, Y.; Zhang, J.; Bi, S. Chemical Composition and Biological Activity of the Essential Oil from Rhododendronanwheiense Flowers. Chem. Nat. Compd. 2022, 58, 947–950. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, W.; Bi, Y.; Han, Y.; Zong, Y.; Prusky, D. Cuminal Inhibits Trichothecium roseum Growth by Triggering Cell Starvation: Transcriptome and Proteome Analysis. Microorganisms 2020, 8, 256. [Google Scholar] [CrossRef]
- Tripathi, P.; Dubey, N.K. Exploitation of Natural Products as an Alternative Strategy to Control Postharvest Fungal Rotting of Fruit and Vegetables. Postharvest Biol. Technol. 2004, 32, 235–245. [Google Scholar] [CrossRef]
- Sivakumar, D.; Bautista-Baños, S. A Review on the Use of Essential Oils for Postharvest Decay Control and Maintenance of Fruit Quality during Storage. Crop Prot. 2014, 64, 27–37. [Google Scholar] [CrossRef]
- Sompila, A.W.G.T.; Mabika, A.B.M.; Pambou-Tobi, N.P.G.; Gouollaly, T.; Moussounga, J.E.; N’simba, G.L.L.B.; Nguie, R.; Matos, L. Evaluation of Some Secondary Metabolites and Determination of the Antioxidant Potential of Different Extracts from the Plant of Pteridium aquilinum. Am. J. Anal. Chem. 2021, 12, 506–519. [Google Scholar] [CrossRef]
- Kanan, G.; Al-Najar, R. In Vitro and In Vivo Activity of Selected Plant Crude Extracts and Fractions Against Penicillium italicum. J. Plant Prot. Res. 2009, 49, 341–352. [Google Scholar] [CrossRef]
- Pangallo, S.; Li Destri Nicosia, M.G.; Raphael, G.; Levin, E.; Ballistreri, G.; Cacciola, S.O.; Rapisarda, P.; Droby, S.; Schena, L. Elicitation of Resistance Responses in Grapefruit and Lemon Fruits Treated with a Pomegranate Peel Extract. Plant Pathol. 2016, 66, 633–640. [Google Scholar] [CrossRef]
- Wu, T.; He, M.; Zang, X.; Zhou, Y.; Qiu, T.; Pan, S.; Xu, X. A Structure–Activity Relationship Study of Flavonoids as Inhibitors of E. coli by Membrane Interaction Effect. Biochim. Biophys. Acta (BBA) Biomembr. 2013, 1828, 2751–2756. [Google Scholar] [CrossRef] [PubMed]
- Hua, C.; Kai, K.; Wang, X.; Shi, W.; Zhang, D.; Liu, Y. Curcumin Inhibits Gray Mold Development in Kiwifruit by Targeting Mitogen-Activated Protein Kinase (MAPK) Cascades in Botrytis cinerea. Postharvest Biol. Technol. 2019, 151, 152–159. [Google Scholar] [CrossRef]
- Teshome, E.; Forsido, S.F.; Rupasinghe, H.P.V.; Olika Keyata, E. Potentials of Natural Preservatives to Enhance Food Safety and Shelf Life: A Review. Sci. World J. 2022, 2022, 9901018. [Google Scholar] [CrossRef]
- Siddaiah, C.N.; Prasanth, K.V.H.; Satyanarayana, N.R.; Mudili, V.; Gupta, V.K.; Kalagatur, N.K.; Satyavati, T.; Dai, X.-F.; Chen, J.-Y.; Mocan, A.; et al. Chitosan Nanoparticles Having Higher Degree of Acetylation Induce Resistance against Pearl Millet Downy Mildew through Nitric Oxide Generation. Sci. Rep. 2018, 8, 2485. [Google Scholar] [CrossRef]
- Zeng, K.; Deng, Y.; Ming, J.; Deng, L. Induction of Disease Resistance and ROS Metabolism in Navel Oranges by Chitosan. Sci. Hortic. 2010, 126, 223–228. [Google Scholar] [CrossRef]
- García-Rincón, J.; Vega-Pérez, J.; Guerra-Sánchez, M.G.; Hernández-Lauzardo, A.N.; Peña-Díaz, A.; Velázquez-Del Valle, M.G. Effect of Chitosan on Growth and Plasma Membrane Properties of Rhizopus stolonifer (Ehrenb.:Fr.) Vuill. Pestic. Biochem. Physiol. 2010, 97, 275–278. [Google Scholar] [CrossRef]
- Akter, J.; Jannat, R.; Hossain, M.M.; Ahmed, J.U.; Rubayet, T. Chitosan for Plant Growth Promotion and Disease Suppression against Anthracnose in Chilli. Int. J. Environ. Agric. Biotechnol. 2018, 3, 806–817. [Google Scholar] [CrossRef]
- Gupta, A.; Bano, A.; Rai, S.; Kumar, M.; Ali, J.; Sharma, S. ACC deaminase producing Plant Growth Promoting Rhizobacteria enhance salinity stress tolerance in Pisum sativum. 3 Biotech 2021, 11, 514. [Google Scholar] [CrossRef]
- Lopez-Moya, F.; Kowbel, D.; Nueda, M.J.; Palma-Guerrero, J.; Glass, N.L.; Lopez-Llorca, L.V. Neurospora crassa Transcriptomics Reveals Oxidative Stress and Plasma Membrane Homeostasis Biology Genes as Key Targets in Response to Chitosan. Mol. BioSyst. 2016, 12, 391–403. [Google Scholar] [CrossRef] [PubMed]
- Jaime, M.D.; Lopez-Llorca, L.V.; Conesa, A.; Lee, A.Y.; Proctor, M.; Heisler, L.E.; Gebbia, M.; Giaever, G.; Westwood, J.T.; Nislow, C. Identification of Yeast Genes That Confer Resistance to Chitosan Oligosaccharide (COS) Using Chemogenomics. BMC Genom. 2012, 13, 267. [Google Scholar] [CrossRef] [PubMed]
- Di Francesco, A.; Placì, N.; Scialanga, B.; Ceredi, G.; Baraldi, E. Ripe Indexes, Hot Water Treatments, and Biocontrol Agents as Synergistic Combination to Control Apple Bull’s Eye Rot. Biocontrol Sci. Technol. 2022, 32, 1016–1026. [Google Scholar] [CrossRef]
- Ngolong Ngea, G.L.; Qian, X.; Yang, Q.; Dhanasekaran, S.; Ianiri, G.; Ballester, A.R.; Zhang, X.; Castoria, R.; Zhang, H. Securing fruit production: Opportunities from the elucidation of the molecular mechanisms of postharvest fungal infections. Compr. Rev. Food Sci. Food Saf. 2021, 20, 2508–2533. [Google Scholar] [CrossRef]
- Luria, N.; Sela, N.; Yaari, M.; Feygenberg, O.; Kobiler, I.; Lers, A.; Prusky, D. De-Novo Assembly of Mango Fruit Peel Transcriptome Reveals Mechanisms of Mango Response to Hot Water Treatment. BMC Genom. 2014, 15, 957. [Google Scholar] [CrossRef]
- Jeong, M.-A.; Jeong, R.-D. Applications of Ionizing Radiation for the Control of Postharvest Diseases in Fresh Produce: Recent Advances. Plant Pathol. 2017, 67, 18–29. [Google Scholar] [CrossRef]
- Rothkamm, K.; Barnard, S.; Moquet, J.; Ellender, M.; Rana, Z.; Burdak-Rothkamm, S. DNA Damage Foci: Meaning and Significance. Environ. Mol. Mutagen. 2015, 56, 491–504. [Google Scholar] [CrossRef]
- Loconsole, D.; Santamaria, P. UV Lighting in Horticulture: A Sustainable Tool for Improving Production Quality and Food Safety. Horticulturae 2021, 7, 9. [Google Scholar] [CrossRef]
- Kaewsuksaeng, S.; Urano, Y.; Aiamla-or, S.; Shigyo, M.; Yamauchi, N. Effect of UV-B Irradiation on Chlorophyll-Degrading Enzyme Activities and Postharvest Quality in Stored Lime (Citrus latifolia Tan.) Fruit. Postharvest Biol. Technol. 2011, 61, 124–130. [Google Scholar] [CrossRef]
- Ruiz, V.E.; Cerioni, L.; Zampini, I.C.; Cuello, S.; Isla, M.I.; Hilal, M.; Rapisarda, V.A. UV-B Radiation on Lemons Enhances Antifungal Activity of Flavedo Extracts against Penicillium digitatum. LWT Food Sci. Technol. 2017, 85, 96–103. [Google Scholar] [CrossRef]
- Usall, J.; Ippolito, A.; Sisquella, M.; Neri, F. Physical Treatments to Control Postharvest Diseases of Fresh Fruits and Vegetables. Postharvest Biol. Technol. 2016, 122, 30–40. [Google Scholar] [CrossRef]
- Lastochkina, O.; Seifikalhor, M.; Aliniaeifard, S.; Baymiev, A.; Pusenkova, L.; Garipova, S.; Kulabuhova, D.; Maksimov, I. Bacillus spp.: Efficient Biotic Strategy to Control Postharvest Diseases of Fruits and Vegetables. Plants 2019, 8, 97. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Bano, A.; Pandey, N.; Rai, S.; Sharma, S.; Pathak, N. Recent Advances in Biotic and Abiotic Stresses of Solanum melongena L. In Solanum melongena: Production, Cultivation and Nutrition; Nova Science Publishers: Hauppauge, NY, USA, 2021; pp. 151–171. ISBN 978-1-68507-311-4. [Google Scholar]
- Gupta, A.; Aamina, S.; Bano, A.; Rai, S.; Bansal, P.; Sharma, S. Effect of Plant Growth Promoting Rhizobacteria (PGPR) on Plant growth under biotic stress condition. In Recent Advances and Approaches for Crop Improvement and Sustainable Agriculture; Aargon Press: New Delhi, India, 2022; pp. 1–12. [Google Scholar]





| Fruits | Phytopathogen | Mycotoxin | Structure (a) | Mycotoxin Biosynthesis Clusters | Reference |
|---|---|---|---|---|---|
| Apple, grape | Penicillium citrinum | Citrinin |  | 6 genes b: (mrl6), citS (mrl4), citE (mrl7), citD (mrl2), citC (mrl1), citB citA | [67] |
| Grape, fig, peach, pepper, tomato, apple | Aspergillus flavus | Aflatoxin B1 |  | 29 genes c: aflC, hypC, aflF, aflD, aflU, aflT, aflH, aflS, aflJ, aflE, aflA, aflB, aflR, aflM, hypE aflV, aflX, aflW, aflY, aflN, aflL, aflG, hypD, hypB, aflI, aflO, aflP, aflK, aflQ, | [68,69,70] |
| Pomegranate, tomato, cherry, peach, pear, pepper, melon, mango, apple, cucumber, citrus, kiwi, litchi, fig, persimmon | Alternaria alternata | Alternariol |  | 6 genes: moxI, pksI, doxI aohR, sdrI, omtI | [71,72,73,74] |
| Tomato, grape, mango, banana, cherry | Aspergillus niger | Ochratoxin A |  | 5 genes: otaR1 otaD, otaC, otaB, otaA | [75] |
| Apple, cherry, quince, pear, plum | Penicillium expansum | Patulin |  | 15 genes: patA, to patO | [76,77,78,79] |
| Host Fruit | Fungi | Gene Name | Disease | Gene Function or Conclusion | References |
|---|---|---|---|---|---|
| Citrus | A. alternata | Nps6 | Anthracnose | Production of siderophores, non-ribosomal peptide synthase, required for full virulence. | [112] |
| Avocado | C. gloeosporioides | CAP20, CAP22 | Anthracnose | Appressorium formation. | [113] |
| Plum; avocado | C. gloeosporioides | AreA | Anthracnose | Conidia production, regulation of nitrogen metabolism, and virulence, influence fungal development. | [114] |
| Avocado | C. gloeosporioides | Pelb | Anthracnose | Decline synthesis of pectinase. | [115,116] |
| Grape | B. cinerea | BcVelB, BcVeA | Gray mold | Response to oxidative stress, Regulate fungal development, and virulence. | [117] |
| Mango | C. gloeosporioides | TUB 1 and TUB2 | Anthracnose | Oxidative stress. | [118] |
| Grape | B. cinerea | SDR1 | Gray mold | Influence development, growth, and pathogenicity. | [119] |
| Apple | P. expansum | VeA | Blue mold | Regulatory mechanism of secondary metabolism. | [13] |
| Apple | P. expansum | PeSte12 | Blue mold | Influence asexual reproduction and virulence. | [120] |
| Apple | P. expansum | LaeA | Blue mold | Modulates virulence. | [121] |
| Citrus | P. digitatum | Ste12 | Green mold | Asexual reproduction. | [122] |
| Citrus | P. digitatum | Os2 | Green mold | Virulence functions. | [123] |
| Citrus | P. digitatum | Pg1; Pg2 | Green mold | Produce polygalacturonases. | [124] |
| Citrus | P. digitatum | Mit1 | Green mold | Contribution to mycelium growth, conidial germination, and sporulation. | [125] |
| Citrus | P. digitatum | Ac1 | Green mold | Required for conidial growth, germination, cAMP production, and virulence. | [126] |
| Citrus | P. digitatum | SreA, SreB | Green mold | Produces aerial mycelia. | [127] |
| Citrus | P. digitatum | Crz1 | Green mold | Conidiation, virulence. | [109] |
| Citrus | P. digitatum | SUT1 | Green mold | Influence fungicide sensitivity and contribute to virulence. | [128] |
| Citrus | P. digitatum | PdMFS1 | Green mold | During citrus infection, they contribute to a variety of fungicide resistance and fungal virulence. | [129] |
| Citrus | P. digitatum | Slt2 | Green mold | Control virulence and sporulation. | [130] |
| Hosts | Diseases | Bacterial | Phytopathogens | Inhibition (%) | References |
|---|---|---|---|---|---|
| Mango | Anthracnose | Stenotrophomonas rhizophila | Colletotrichum gloeosporioides | 89 | [146] |
| Peach | Brown rot | Pseudomonas synxantha | Monilinia fructicola | 70 | [147] |
| Apple | Brown rot | Bacillus amyloliquefaciens | Monilinia sp. | 75 | [148] |
| Apple | Blue mold | Pseudomonas fluorescens | Penicillium expansum | 88 | [149] |
| Banana | Anthracnose | Pantoea agglomerans | Colletotrichum musae | 94 | [150] |
| Banana | Anthracnose | Bacillus subtilis | Colletotrichum musae | 72 | [151] |
| Melon | Rot | B. subtilis | Alternaria alternata | 77 | [152] |
| Grape | Gray mold | Bacillus sp. | Botrytis cinerea | 50 | [153] |
| Apple | Anthracnose | Paenibacillus polymyxa | C. gloeosporioides | 83 | [154] |
| Apple | Blue mold | B. amyloliquefaciens | Penicillium expansum | 80 | [155] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bano, A.; Gupta, A.; Prusty, M.R.; Kumar, M. Elicitation of Fruit Fungi Infection and Its Protective Response to Improve the Postharvest Quality of Fruits. Stresses 2023, 3, 231-255. https://doi.org/10.3390/stresses3010018
Bano A, Gupta A, Prusty MR, Kumar M. Elicitation of Fruit Fungi Infection and Its Protective Response to Improve the Postharvest Quality of Fruits. Stresses. 2023; 3(1):231-255. https://doi.org/10.3390/stresses3010018
Chicago/Turabian StyleBano, Ambreen, Anmol Gupta, Manas Ranjan Prusty, and Manoj Kumar. 2023. "Elicitation of Fruit Fungi Infection and Its Protective Response to Improve the Postharvest Quality of Fruits" Stresses 3, no. 1: 231-255. https://doi.org/10.3390/stresses3010018
APA StyleBano, A., Gupta, A., Prusty, M. R., & Kumar, M. (2023). Elicitation of Fruit Fungi Infection and Its Protective Response to Improve the Postharvest Quality of Fruits. Stresses, 3(1), 231-255. https://doi.org/10.3390/stresses3010018