Effect of Gender and Anthropometrics on the Kinematics of the Lunge in Young Foil Fencers (Lunge Kinematics in Young Fencers)



Abstract
:1. Introduction
2. Materials and Methods
3. Anthropometry
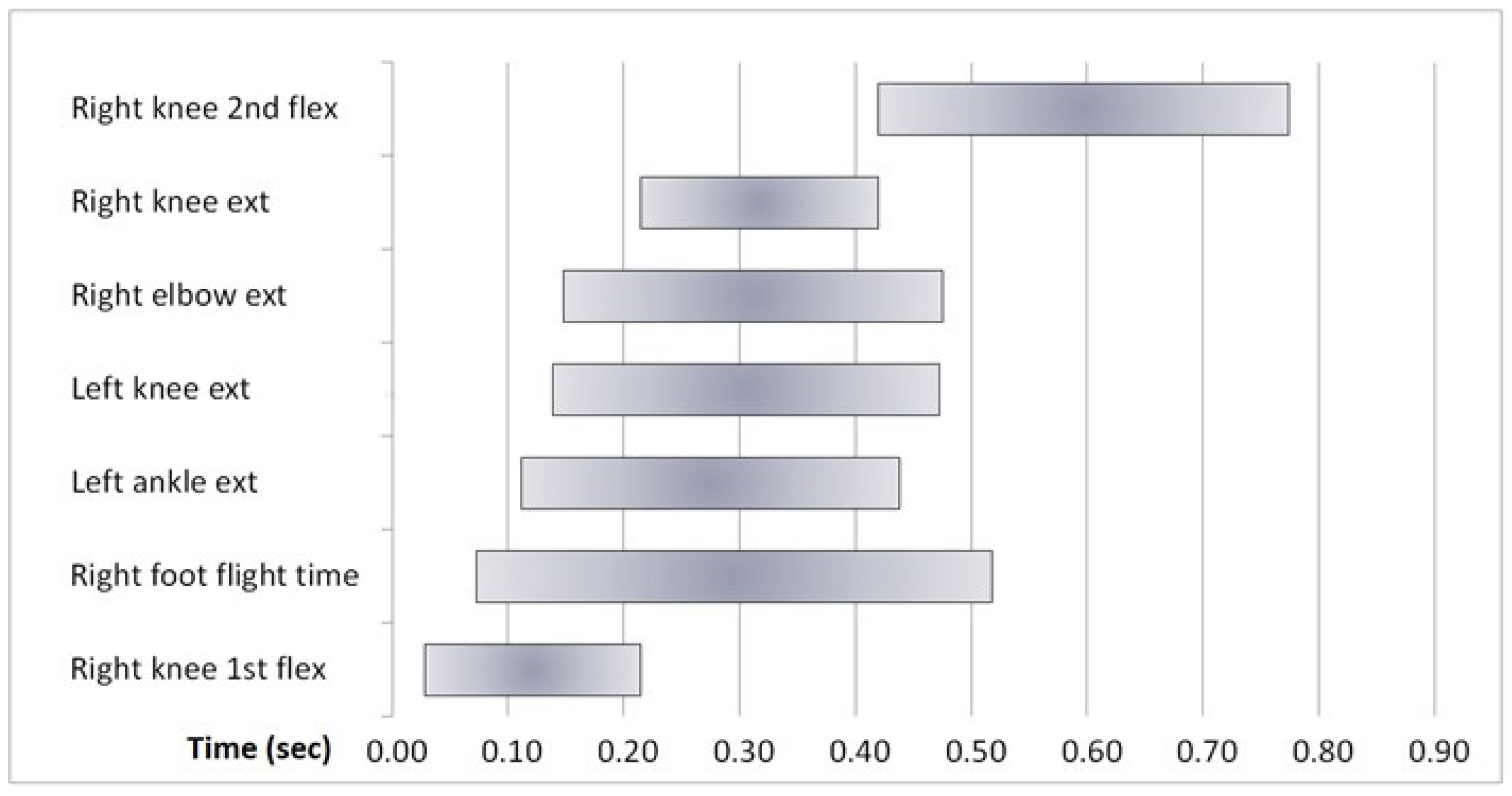
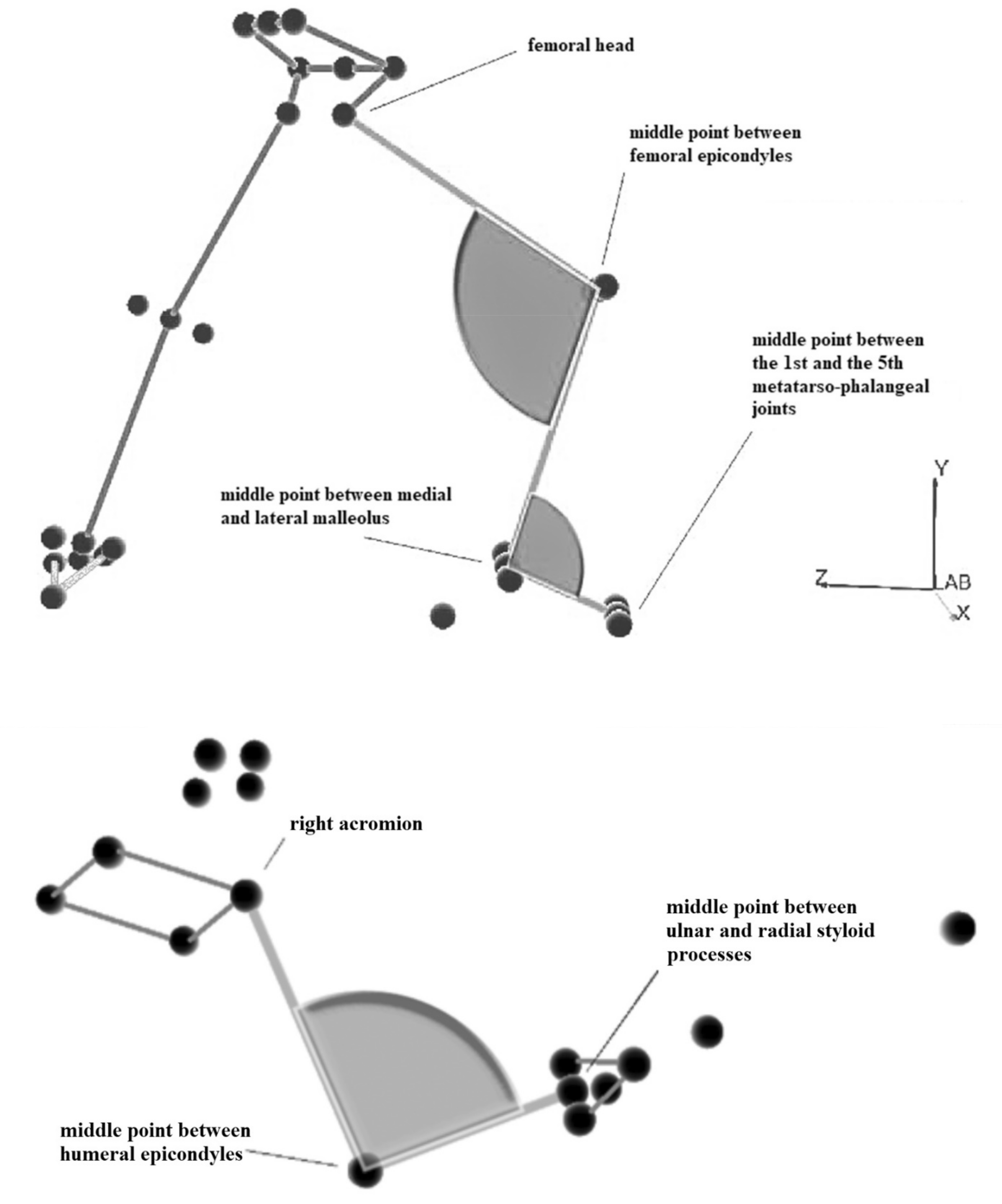
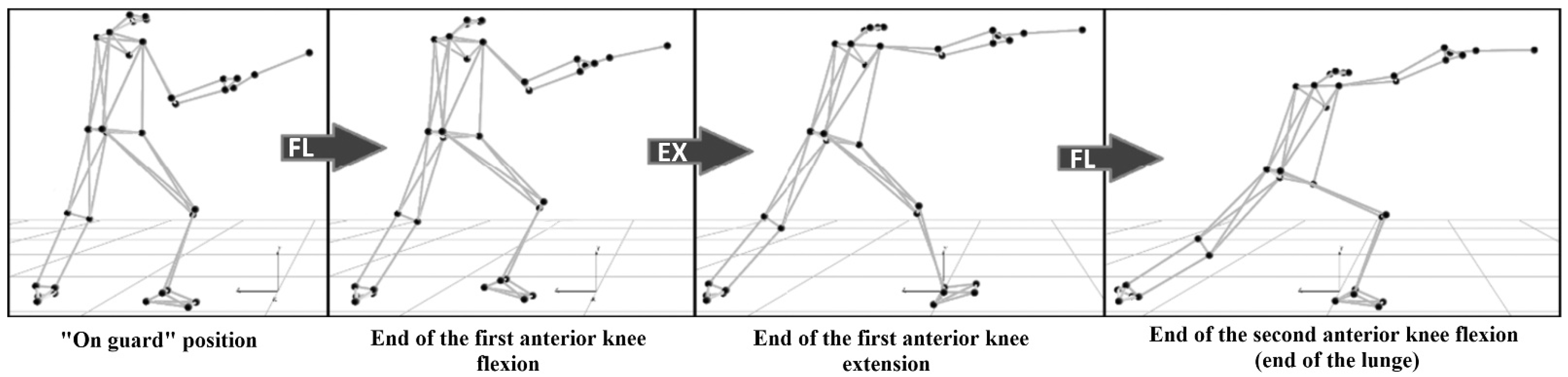
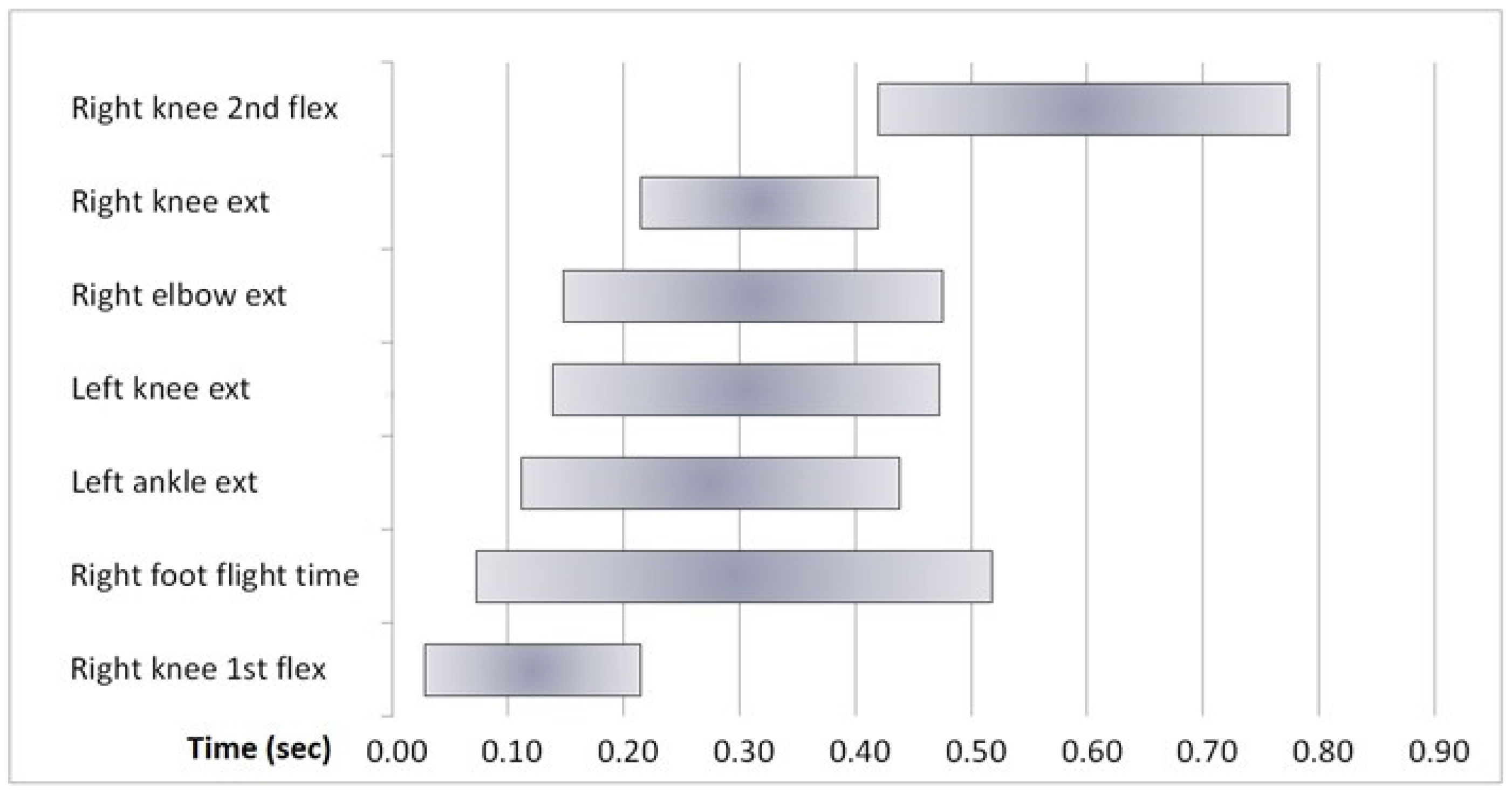
4. Kinematics
5. Statistics
6. Results
7. Discussion
7.1. Anthropometry
7.2. Kinematics
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Aquili, A.; Tancredi, V.; Triossi, T.; De Sanctis, D.; Padua, E.; D’Arcangelo, G.; Melchiorri, G. Performance analysis in saber. J. Strength Cond. Res. 2013, 7, 624–630. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.W.L.; Wong, D.W.; Wang, Y.; Ren, S.; Yan, F.; Zhang, M. Biomechanics of fencing sport: A scoping review. PLoS ONE 2017, 2, e0171578. [Google Scholar] [CrossRef] [PubMed]
- Hassan, S.E.A.; Klauck, J. Kinematics of lower and upper extremities motions during the fencing lunge: Results and training implications. In Proceedings of the 16th International Symposium on Biomechanics in Sports Conference, Konstanz, Germany, 21–25 July 1998; Volume 1, pp. 170–173. [Google Scholar]
- Gholipour, M.; Tabriz, A.; Farahmand, F. Kinematics analysis of lunge fencing using stereophotogrametry. World J. Sport Sci. 2008, 1, 32–37. [Google Scholar]
- Frère, J.; Gopfert, B.; Nuesch, C.; Huber, C.; Fischer, M.; Wirz, D.; Friederich, N.F. Kinematical and EMG-classifications of a fencing attack. Int. J. Sport Med. 2011, 32, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Morris, N.; Farnsworth, M.; Robertson, D.G.E. Kinetic analyses of two fencing attacks and lunge and fleche. Port. J. Sport Sci. 2001, 11, 343–346. [Google Scholar]
- Mullo, F.; Mullineaux, D.; Irwin, G. Use of the kinematic chain in the fencing attacking lunge. In Proceedings of the 33 International Conference on Biomechanics in Sports, Poitiers, France, 29 June–3 July 2015. [Google Scholar]
- Bottoms, L.; Greenhalgh, A.; Sinclair, J. Kinematic determinants of weapon velocity during the fencing lunge in experienced épée fencers. Acta Bioeng. Biomech. 2013, 15, 109–113. [Google Scholar] [PubMed]
- Guan, Y.; Guo, L.; Wu, N.; Zhang, L.; Warburton, D.E.R. Biomechanical insights into the determinants of speed in the fencing lunge. Eur. J. Sports Sci. 2018, 18, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Sinclair, J.; Bottoms, L. Gender differences in the kinetics and lower extremity kinematics of the fencing lunge. Int. J. Perform. Anal. Sport 2013, 13, 440–451. [Google Scholar] [CrossRef]
- Marfell-Jones, M.; Olds, T.; Stewart, A.; Lindsay-Carter, L.E. ISAK Manual, International Standards for Anthropometric Assessment; International Society for the Advancement of Kinanthropometry: Lower Hutt, New Zealand, 2011. [Google Scholar]
- Martine, T.; Claessens, A.L.; Vlietinck, R.; Marchal, G.; Beunen, G. Accuracy of anthropometric estimation of muscle cross-sectional area of the arm in males. Am. J. Hum. Biol. 1997, 9, 73–86. [Google Scholar] [CrossRef]
- Knapik, J.J.; Staab, J.S.; Harman, E.A. Validity of an anthropometric estimate of thigh muscle cross-sectional area. Med. Sci. Sports Exerc. 1996, 28, 1523–1530. [Google Scholar] [CrossRef] [PubMed]
- McGinley, J.L.; Baker, R.; Wolfe, R.; Morris, M.E. The reliability of three-dimensional kinematic gait measurements: A systematic review. Gait Posture 2009, 29, 360–369. [Google Scholar] [CrossRef] [PubMed]
- Oxford Metrics. Plug-in Gait Reference Guide; Oxford Metrics: Oxford, UK, 2016. [Google Scholar]
- Wu, G.; Siegler, S.; Allard, P.; Kirtley, C.; Leardini, A.; Rosenbaum, D.; Whittle, M.; D’Lima, D.D.; Cristofolini, M.; Witte, H. ISB recommendation on definitions of joint coordinate system of various joints for the reporting of human joint motion—Part I: Ankle, hip, and spine. J. Biomech. 2002, 35, 543–548. [Google Scholar] [CrossRef]
- Wu, G.; Van der Helm, F.C.; Veeger, H.E.; Makhsous, M.; Van Roy, P.; Anglin, C.; Nagels, J.; Karduna, A.R.; McQuade, K.; Wang, X.; et al. ISB recommendation on definitions of joint coordinate systems of various joints for the reporting of human joint motion--Part II: Shoulder, elbow, wrist and hand. J. Biomech. 2005, 38, 981–992. [Google Scholar] [CrossRef] [PubMed]
- Austin, P.C.; Steyerberg, E.W. The number of subjects per variable required in linear regression analyses. J. Clin. Epidemiol. 2015, 68, 627–636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cacciari, E.; Milani, S.; Balsamo, A.; Spada, E.; Bona, G.; Cavallo, L.; Cerutti, F.; Gargantini, L.; Greggio, N.; Tonini, G. Italian cross-sectional growth charts for height, weight and BMI (2-20y). J. Endocrinol. Investig. 2006, 29, 581–593. [Google Scholar] [CrossRef] [PubMed]
- Turner, A.; Bishop, C.; Chavda, S.; Edwards, M.; Brazier, J.; Kilduff, L.P. Physical characteristics underpinning lunging and change of direction speed in fencing. J. Strength Cond. Res. 2016, 30, 2235–2241. [Google Scholar] [CrossRef] [PubMed]
- Tsolakis, C.; Bogdanis, G.C.; Vagenas, G. Anthropometric profile and limb asymmetries in young male and female fencers. J. Hum. Mov. Stud. 2006, 50, 201–215. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age (Years) | Height (cm) | Weight (kg) | |
|---|---|---|---|
| Mean ± SD | Mean ± SD | Mean ± SD | |
| Males (n. 8) | 13.6 ± 3.1 | 162.5 ± 18.5 | 53.1 ± 15.3 |
| Female (n. 7) | 11.7 ± 2 | 152.8 ± 10.7 | 46.8 ± 7.2 |
| M (N = 7) | F (N = 8) | |||
|---|---|---|---|---|
| M | ±SD | M | ±SD | |
| BMI | 19.7 | 3.2 | 20.1 | 1.5 |
| Fat mass (kg) | 44.8 | 12.9 | 35.6 | 6.9 |
| Fat mass % | 84.9 | 6.3 | 75.8 | 5.2 |
| Fat Free mass (kg) | 8.3 | 4.4 | 11.2 | 2.3 |
| Fat Free mass % | 0.0 | 6.3 | 24.2 | 0.0 |
| Upper limb length (cm) | 69.8 | 9.1 | 65.6 | 5.5 |
| Lower limb length (cm) | 81.5 | 10.3 | 77.1 | 5.3 |
| Trunk length (cm) | 81.0 | 8.3 | 75.7 | 6.3 |
| Circumf. upper arm (cm) | 23.5 | 3.7 | 23.0 | 2.1 |
| Biceps skinfold (cm) | 0.8 | 0.5 | 1.1 | 0.3 |
| Tricep skinfold (cm) | 1.2 | 0.7 | 1.6 | 0.3 |
| CSA upper arm (cm2) | 30.2 | 8.7 | 24.1 | 4.6 |
| M (N = 7) | F (N = 8) | |||||||
|---|---|---|---|---|---|---|---|---|
| Right | Left | Right | Left | |||||
| M | ±SD | M | ±SD | M | ±SD | M | ±SD | |
| Thigh length (cm) | 39.2 | 4.9 | 39.2 | 4.8 | 37.6 | 3.1 | 37.7 | 3.1 |
| Bi-epicondylar breadth (cm) | 8.5 | 0.7 | 0.0 | 0.7 | 8.2 | 0.6 | 8.1 | 0.0 |
| Thigh circumference (cm) | 49.9 | 7.2 | 48.5 | 6.8 | 51.9 | 3.3 | 51.3 | 3.3 |
| Thigh Skinfold (cm) | 1.5 | 0.6 | 1.5 | 0.6 | 2.7 | 0.4 | 2.7 | 0.4 |
| CSA thigh (cm2) | 131.9 | 32.6 | 0.0 | 32.6 | 118.3 | 18.3 | 118.3 | 0.0 |
| M (N = 7) | F (N = 8) | |||||
|---|---|---|---|---|---|---|
| M | ±DS | M | ±DS | Effect Size | p | |
| FM% | 15.1 | 6.2 | 24.2 | 5.1 | 1.6 | 0.011 |
| FFM% | 84.8 | 6.2 | 75.7 | 5.1 | 1.6 | 0.011 |
| Right thigh skinfold (cm) | 1.54 | 0.6 | 2.6 | 0.4 | 2.1 | 0.003 |
| Left thigh skinfold (cm) | 1.53 | 0.6 | 2.6 | 0.4 | 2.2 | 0.002 |
| B | SE | β | p | |
|---|---|---|---|---|
| Vm ASISm | 0.516 | 0.091 | 0.000 | |
| 0.009 | 0.002 | 0.733 | 0.002 | |
| Lunge distance | 0.531 | 0.129 | 0.001 | |
| 0.015 | 0.003 | 0.797 | 0.000 |
| Angles (deg) | Mean | ±SD |
|---|---|---|
| On guard | ||
| Elbow (deg) | 96 | 16 |
| Forward knee (deg) | 120 | 10 |
| Rear knee (deg) | 121 | 10 |
| Forward ankle (deg) | 98 | 6 |
| Rear ankle (deg) | 83 | 5 |
| During lunge | ||
| Elbow extension angle (deg) | 151 | 5 |
| End of forward knee at 1st flexion (deg) | 103 | 11 |
| End of forward knee extension (deg) | 151 | 5 |
| At final lunge position | ||
| Elbow (deg) | 150 | 5 |
| Forward knee (deg) | 90 | 9 |
| Rear knee(deg) | 170 | 4 |
| Rear ankle (deg) | 103 | 6 |
| Rear ankle (deg) | 116 | 5 |
| Range of motion | ||
| Elbow extension (deg) | 55 | 17 |
| Forward knee 1st flexion (deg) | 17 | 7 |
| Forward knee in extension (deg) | 47 | 11 |
| Forward knee 2nd flexion (deg) | 59 | 8 |
| Rear knee extension (deg) | 48 | 11 |
| Rear ankle plantar flexion (deg) | 32 | 6 |
| Linear velocities and lunge distance and duration | ||
| Mean velocity of ASISm (m/s) | 0.9 | 0.1 |
| Mean velocity of 3met (m/s) | 1.3 | 0.2 |
| Peal Velocity ASISm (m/s) | 1.7 | 0.2 |
| Peak velocity 3 met (m/s) | 2.4 | 0.3 |
| Lunge distance (m) | 1.1 | 0.2 |
| Lunge duration (s) | 0.7 | 0.1 |
| Angular Velocities | ||
| Elbow extension (deg/s) | 180.4 | 56.3 |
| Forward knee 1st flexion (deg) | 87.8 | 20.4 |
| Forward knee extension (deg) | 242.1 | 61.3 |
| Forward knee 2nd flexion (deg) | 179.5 | 26.8 |
| Rear knee extension (deg) | 160.9 | 46.9 |
| Rear ankle plantar flexion (deg) | 108.6 | 26.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mocco, A.; Wang, J.; Cicchella, A. Effect of Gender and Anthropometrics on the Kinematics of the Lunge in Young Foil Fencers (Lunge Kinematics in Young Fencers). Biomechanics 2021, 1, 167-177. https://doi.org/10.3390/biomechanics1020014
Mocco A, Wang J, Cicchella A. Effect of Gender and Anthropometrics on the Kinematics of the Lunge in Young Foil Fencers (Lunge Kinematics in Young Fencers). Biomechanics. 2021; 1(2):167-177. https://doi.org/10.3390/biomechanics1020014
Chicago/Turabian StyleMocco, Alessio, Jun Wang, and Antonio Cicchella. 2021. "Effect of Gender and Anthropometrics on the Kinematics of the Lunge in Young Foil Fencers (Lunge Kinematics in Young Fencers)" Biomechanics 1, no. 2: 167-177. https://doi.org/10.3390/biomechanics1020014
APA StyleMocco, A., Wang, J., & Cicchella, A. (2021). Effect of Gender and Anthropometrics on the Kinematics of the Lunge in Young Foil Fencers (Lunge Kinematics in Young Fencers). Biomechanics, 1(2), 167-177. https://doi.org/10.3390/biomechanics1020014