Abstract
Full-length cDNAs encoding macrophage inhibitory factor-2 (MIF-2) were cloned from Teladorsagia circumcincta (TcMIF-2) and Haemonchus contortus (HcMIF-2). TcMIF-2 and HcMIF-2 cDNA (342 bp) encoded proteins of 114 amino acids, each of which was present as a single band of about 16 kDa on SDS-PAGE. Multiple alignments of the protein sequences showed homology of 84% between TcMIF-2 and HcMIF-2, 54–76% with MIF-2s of seven nematodes, but low homology with other MIF sequences. The predicted three-dimensional structures revealed an overall structural homology of TcMIF-2 and HcMIF-2, highly conserved binding and catalytic sites and minor differences in the tautomerase binding site residues in other nematode MIF-2 homologues. A phylogenetic tree was constructed using helminth and mammalian MIF-1 and MIF-2 sequences. Soluble C-terminal MIF-2 proteins were cloned in arabinose inducible promotor AY2.4, expressed in Escherichia coli strain AY2.4 and purified. Recombinant TcMIF-2 and HcMIF-2 had similar enzyme activities in a standard tautomerase assay. Recombinant HcMIF-2 activity was approximately halved by storage at 4 °C, −20 °C or −70 °C. Antibodies in serum and saliva from field-immune, but not nematode-naïve, sheep recognised recombinant HcMIF-2 and TcMIF-2 in enzyme-linked immunosorbent assays. Recognition of the recombinant proteins by antibodies generated by exposure of sheep to the native enzyme indicates similar antigenicity of the two proteins.
1. Introduction
The abomasal nematode parasites Haemonchus contortus and Teladorsagia circumcincta cause significant loss of production and ill-health in sheep and goats. Widespread resistance to anthelmintics has necessitated developing alternative parasite control strategies, such as vaccination, which involves identification of antigens or combinations of antigens, appropriate adjuvants and vaccination protocols. Amongst the most promising antigens are multifunctional proteins, which allow the host immune response to disrupt worm physiology at more than point.
Macrophage migration inhibitory factors (MIFs), theta class glutathione transferases (GSTs) with oxido-reductase and tautomerase activities [1], are ligands of the CD74 cell receptor [2]. MIF was originally identified as a T-cell pro-inflammatory cytokine [3,4], but has subsequently been shown to be expressed in other tissues, where it has tissue-specific actions and generally promotes cell survival and inhibits senescence [5]. In humans, MIF antagonises the effects of aging and ameliorates age-related diseases [6,7]. Many of these properties are shared with non-mammalian MIF proteins [8], including those in free-living and parasitic nematodes [9,10,11,12,13,14,15,16], although some biochemical differences were apparent between nematode and mammalian MIF proteins in substrate affinity [17] and sensitivity to inhibitors [12] of the tautomerase activity. Importantly, MIF proteins released by parasitic nematodes were able to modify the host immune response and promote parasite survival [16,18,19,20].
In contrast to the single mif gene in mammals, gene duplication in nematode lineages has resulted in multiple genes. There are four mif genes in Caenorhabditis elegans [21], encoding proteins which are 15–30% identical to each other. MIF-1 and MIF-2 proteins have been identified in parasitic nematodes, with MIF-1 being more similar to mammalian MIF [9]. H. contortus MIF-1 and MIF-2 are both involved in larval development; RNAi using either mif-1 or mif-2 dsRNA resulted in strong developmental delay phenotypes: the majority after mif-1 knockout were arrested/killed at L1 and only 23% reached the L3 stage, whereas for MIF-2, 59% of worms became L3 and the others were mainly arrested at the L2 stage (unpublished data). In contrast, mif-1 knockout in C. elegans does not confer an obvious deleterious phenotype, whereas both mif-2 RNAi and a deletion mutant are embryonic lethal [21]. Interestingly, Nisbet et al. [11] did not find either their recombinant or purified native TcMIF-1 influenced sheep monocyte migration. These inconsistent observations suggest that both MIF-1 and MIF-2 could be candidate vaccine antigens and that the superiority of either one in a multi-antigen vaccine against abomasal nematodes remains to be assessed.
In the present study, cDNAs encoding T. circumcincta (TcMIF-2) and H. contortus MIF-2 (HcMIF-2) were cloned, expressed in Escherichia coli and the recombinant proteins purified. Tautomerase activity of both recombinant enzymes was confirmed. Enzyme-linked immunosorbent assays (ELISAs) were performed to determine if the recombinant proteins were recognised by antibodies in saliva and serum from immune sheep previously exposed to a wide range nematode parasites in the field.
2. Materials and Methods
All chemicals were purchased from the Sigma Chemical Co. (St. Louis, MO, USA) unless stated otherwise.
2.1. Parasites
Pure cultures of T. circumcincta and H. contortus were maintained in the laboratory by regular passage through sheep. Adult worms were recovered from the abomasa of infected sheep, as described previously [22]. Briefly, abomasal contents were mixed 2:1 with 3% agar and the solidified agar blocks incubated at 37 °C in a saline bath. Clumps of parasites were collected from the saline soon after emergence and frozen in Microcentrifuge tubes (Salt Lake City, UT, USA) at −80 °C for molecular biology procedures.
2.2. RNA Isolation and Synthesis of cDNA
Adult T. circumcincta or H. contortus (50–100 µL packed volume) in 1 mL Trizol (Life Technologies, Carlsbad, CA, USA) were ground to a fine powder in a mortar under liquid N2 and RNA was extracted according to the manufacturer’s instructions. The quality and concentration of the RNA was assessed, and first strand cDNA synthesised from 1 µg using the iScript Select cDNA Synthesis Kit (Bio-Rad, Auckland, New Zealand) and a 1:1 mixture of Oligo (dT)20 and random primers.
The full-length H. contortus MIF-2 sequence was amplified from cDNA in a PCR containing the oligonucleotide primers Hcmif-2FLF (5′-CCTATGGTAAGAGTTGC-3′) and Hcmif-2FLR (5′-CGACAGTTGCAGCTGCCAC-3′). These primers were developed from the publically available H. contortus MIF-2 EST sequence (HC07649). Initially, the primers Hcmif-2 FLF and Hcmif-2 FLR were designed and used to amplify the H. contortus MIF-2 coding region from H. contortus adult cDNA using the Expand High Fidelity PCR system according to the manufacturer’s instructions (ThermoFisher Scientific, Waltham, USA). The resulting amplification product was cloned into pCR4-TOPO using the TOPO TA cloning kit for sequencing (ThermoFisher Scientific, Waltham, USA) and sequenced to confirm identity. The restriction enzyme sites NdeI and NotI were added to the H. contortus MIF-2 coding region using the oligonucleotide primers Hcmif-2-NdeIF and Hcmif-2-NotIR, the pCR4 clone as template and the Expand High Fidelity PCR system. The product was digested with NdeI and NotI before cloning into the similarly digested expression vector AY2.4 [23] to generate a construct which enabled the arabinose-inducible production of a recombinant C-terminal His- and E-tagged H. contortus MIF-2 fusion protein in E. coli.
A partial T. circumcincta MIF-2 sequence TDC15440 (AgResearch’s Internal database), 3′ RACE primers (TcmifF-2_RACE-F1 (5′-CACTGACATATTGGCGGAGTCCATG-3′) and Tcmif-2_RACE-F2 (5′-GTTTACGCTGGCCAGCGCATTATG-3′) were used to obtain the 3′ of the gene. Subsequently, the full-length T. circumcincta MIF-2 sequence was amplified from cDNA in a PCR containing the oligonucleotide primers Tcmif-2-NdeIF (5′-ATCGCATATGCCGATGGTGAGAGTTGC-3′) and Tcmif-2-NotIR (5′-GATCGCGGCCGCGACGGTTGCAGCGGCCAC-3′). The full-length gene was cloned, using standard protocols, into the expression vector AY2.4 using the restriction enzymes Ndel and Notl (engineered into the forward and reverse primers, underlined in primer sequences) to allow the production of C-terminal His-tagged recombinant protein. The expression clone was sequenced to confirm identity.
The Muscle alignment option in Geneious Prime (Biomatters Ltd., Auckland, New Zealand) with the Blosum 62 similarity matrix was used to determine 100% similarity of TcMIF-2 to H. contortus and other MIFs. The active and binding sites labelled were identified in the structural modelling (see Section 2.3). A phylogenetic tree of TcMIF-2 and HcMIF-2 and their helminth homologues was constructed, using the genetic distance model of Jukes-Canter by the neighbour joining method in Geneious Prime (Auckland, New Zealand).
2.3. Protein Modelling and Structural Analysis of TcMIF-2 and HcMIF-2
The Position-Specific Iterative Basic Local Alignment Search Tool (PSI-BLAST) [24] was used to compare the TcMIF-2 and HcMIF-2 protein sequences with deposited structures in the Protein Data Bank (PDB). The structural models of TcMIF-2 and HcMIF-2 were constructed by submitting the amino acid sequences obtained to the I-TASSER server [25]. For TcMIF-2, only one model was obtained, and for HcMIF-2 a model was selected from the best of five possible models generated. The structural model with highest C-score was further validated using Procheck [26] and ProSA-web [27]. The substrate binding domains were identified, and active site residues deduced using PyMol [28].
2.4. Expression of Recombinant TcMIF-2 and HcMIF-2 in E. coli
E. coli strain BL21 (DE3) transformed with E. coli TcMIF-2 or HcMIF-2 in AY2.4 to allow the production of N-terminal His tagged recombinant proteins under the control of the heat-inducible tandem λPR and PL promoters [29]. The culture was grown in 10 mL Luria Broth (LB) supplemented with 100 µg/mL ampicillin for 16 h at 37 °C and 250 rpm. The culture was diluted 20-fold in LB with 100 µg/mL ampicillin and grown to OD600nm of 0.6–0.8 at 37 °C and 250 rpm. L-arabinose was added to a final concentration of 0.2% and the culture grown for an additional 4 h at 30 °C and 250 rpm. The pellet was weighted and resuspended in 10 mL per gram of pelleted bacteria of equilibration buffer (20 mM sodium bi-phosphate, 0.5 M NaCl, 20 mM Imidazole, pH 7.4). Protease inhibitors were added to the suspension, which was then passed through, the chamber of a MP110 Microfluidizer® (Microfluidics, Westwood, CA, USA) seven times consecutively under ice, at 20,000 psi to ensure the complete lysis of E. coli as recommended by the manufacturer. The crude lysate was centrifuged at 15,000× g for 20 min at 4 °C to remove all cell debris, and the supernatant was collected and filtered through a 0.22 µm filter.
2.5. Purification
Purified recombinant poly-histidine protein was obtained by Fast Protein Liquid Chromatography (FPLC) under native conditions using a HispTap HP column (GE Healthcare, Chicago, IL, USA), and the BioLogic DuoFlow chromatography system (Bio-Rad, Auckland, New Zealand). Sodium biphosphate buffer was used as an equilibration buffer, sodium biphosphate containing 20 mM imidazole as the wash buffer and sodium biphosphate containing 500 mM imidazole as elution buffer. Following the elution, the protein was dialysed overnight, and the concentration was determined by the Nanodrop 1000TM (Thermo Scientific, Waltham, MA, USA) using the A280 nm assay using the extinction coefficient 4470 M−1cm−1 and the molecular weight 16 KDa.
2.6. Gel Electrophoresis
SDS-PAGE was performed using NuPAGE Novex 4–12% Bis-Tris gels according to the instructions of the manufacturer (Invitrogen, Waltham, MA, USA). Gels were stained with SimplyBlueTM SafeStain (Invitrogen Waltham, MA, USA). A Western blot was also performed on the protein using a monoclonal anti-polyhistidine-peroxidase antibody. Blots were incubated overnight in 1:2000 antibody in buffer (4% skim milk powder in Tris-buffered saline and 0.1% Tween-20) at room temperature and developed to detect His-tagged recombinant protein.
2.7. HcMIF-2 and TcMIF-2 Assays
The tautomerase activities of recombinant HcMIF-2 and TcMIF-2 were determined from the tautomerisation of the coloured dopachrome methyl ester to its colourless enol form [30]. Dopochrome methyl ester was prepared by mixing 4 mM L-3,4-dihydroxylphenylalanine methyl ester with 8 mM sodium periodate in potassium phosphate buffer (25 mM potassium phosphate pH 6.0, 0.5 mM EDTA) for 5 min at room temperature before placing on ice for 20 min. The decrease in absorbance at 475 nm was monitored at room temperature over a 5 min period in a cuvette (1 mL final volume) containing 20 µg recombinant enzyme in assay buffer containing dopochrome methyl ester (final OD475nm 0.7–0.9). Background readings were taken before the addition of recombinant enzyme.
The effect on tautomerase activity of varying the concentration of recombinant HcMIF-2 in the assay from 2–20 µg/mL of protein was determined. The effect of storage of recombinant HcMIF-2 at 4 °C, −20 °C and −70 °C for 16 h on enzyme activity was determined and the activities of two separate batches of FPLC-purified recombinant HcMIF-2 were compared.
2.8. ELISA
Pooled serum and saliva samples collected from parasite-naive and parasite-exposed sheep were tested by ELISA for the presence of antibodies that react with recombinant TcMIF-2 or HcMIF-2. Serum and saliva samples were collected from 18 male 6–7 months-old Romney lambs previously exposed to multiple species of parasite, including H. contortus and T. circumcincta. These lambs had developed immunity against T. circumcincta infection. 5 µg/mL TcMIF-2 or HcMIF-2 were immobilised onto ELISA plates (Maxisorp, Thermo Scientific), free binding sites were blocked with Superblock (Thermo Scientific), followed by incubation for 2 h at room temperature with serial dilutions of serum (200- to 6400-fold) or saliva (20- to 160-fold) in ELISA buffer. Bound serum immunoglobulins were then detected by incubation for 2 h at 37 °C with 1:4000 diluted rabbit anti-sheep IgG-HRP and colour development with 3,3′,5,5′-tetramethylbenzidine (TMB). Salivary IgA was similarly detected with rabbit anti-sheep IgA-HRP.
3. Results
3.1. TcMIF-2 and HcMIF-2 Gene Sequences
The 342 bp full-length T. circumcincta and H. contortus cDNA sequences, amplified from adult T. circumcincta and H. contortus cDNA, respectively, have been deposited in Genbank. The predicted proteins consisted of 114 amino acids (Figure 1).

Figure 1.
Multiple protein sequence alignment of T. circumcincta MIF-2, H. contortus MIF-2, Heligmosomoides polygyrus MIF-2 (GI: VDO97074), Ancylostoma caninum MIF-2 (GI: ABU68338), Parastrongyloides trichosuri MIF-2 (GI: BM513256), Caenorhabditis elegans MIF-2 (GI: NP_001256387.1), Caenorhabditis briggsae MIF-2 (GI: CAP39673.2), Brugia malayi MIF-2 (GI: AAF91074), Onchocerca volvulus MIF-2 (GI: AAK66564), Oesophagostomum dentatum MIF-2 (GI: KHJ75996.1), Ancylostoma ceylanicum MIF-2 (GI: ABO31935), Ascaris suum MIF-2 (GI: ADY48840), Caenorhabditis elegans MIF-1 (GI: NP_499536.1), Caenorhabditis elegans MIF-3 (GI: NP_492069), Caenorhabditis elegans MIF-4 (GI: NP_500968.1), Mus musculus MIF (GI: P34884-1), Gallus gallus MIF (GI: NP_001292020), Ovis aries MIF (GI: NP_001072123), Bos tautus MIF (GI: NP_001028780) and Homo sapiens MIF (GI: CAG46452). TcMIF-2 and HcMIF-2 catalytic and active residues are marked by an asterisk (*). The % homology of each sequence with TeciMIF-2 is shown at the end of the alignment.
Multiple alignments of the protein sequences of TcMIF-2 and HcMIF-2 with homologues from Heligmosomoides polygyrus, Ancylostoma caninum, Parastrongyloides trichosuri, C. elegans, Caenorhabditis briggsae, Brugia malayi, Onchocerca volvulus, Oesophagostomum dentatum, Ancylostoma ceylanicum and Ascaris suum, and MIFs from C. elegans, Mus musculus, Gallus gallus, Ovis aries, Bos taurus and Homo sapiens, are shown in Figure 1. Residues involved in tautomerase activity in TcMIF-2 and HcMIF-2, marked by an asterisk, were largely conserved in the MIF-2s of several nematode species. There was 84% homology between TcMIF-2 and HcMIF-2 and 54–76% homology between MIF-2s of seven nematodes, but there was low homology with the other MIF sequences in the multiple alignment.
A search using PSI-BLAST against the Protein Data Bank (PDB) showed that the amino-acid sequences of TcMIF-2 and HcMIF-2 resulted in the assignment of a putative function to two of the top blast hits annotated as hypothetical protein (locus tag EYC32381 to EYC32384 of A. ceylanicum) and protein of unknown function (locus tag VDO97074 of H. polygyrus) with 65–75% and 80% identity, respectively.
In another alignment, the amino acid sequence of HcMIF-2 corresponded to the product of the gene designated as HCON_NP_LOC13552 located on chromosome 5 (GenBank accession number CP035803, BioProject accession number PRJNA517503) from the H. contortus NZ_Hco_NP genome version 1.0 [31].
The phylogenetic tree using multiple helminth and mammalian MIF sequences (Figure 2) revealed that the TcMIF-2 was most closely related to the MIF-2 of the nematode H. contortus and less like other MIFs (MIF-1, MIF-3, MIF-4) from helminths and mammals. The phylogenetic tree also revealed that MIF-1 and MIF-2 are two distinct families and clearly separated from each other.

Figure 2.
Phylogenetic tree of T. circumcincta MIF-2, H. contortus MIF-2, Caenorhabditis elegans MIF-2 (GI: NP_001256387.1), Caenorhabditis elegans MIF-3 (GI: NP_492069), Caenorhabditis elegans MIF-4 (GI: NP_500968.1), Caenorhabditis briggsae MIF-2 (GI: CAP39673.2), Heligmosomoides polygyrus MIF-2 (GI: VDO97074), Parastrongyloides trichosuri MIF-2 (GI: BM513256), Ancylostoma ceylanicum MIF-2 (GI: ABO31935), Onchocerca volvulus MIF-2 (GI: AAK66564), Brugia malayi MIF-2 (GI: AAF91074), Ancylostoma caninum MIF (GI: ABU68338), Ascaris suum MIF-2 (GI: ADY48840), Oesophagostomum dentatum MIF-2 (GI: KHJ75996.1), Mus musculus MIF (GI: P34884-1), Gallus gallus MIF (GI: NP_001292020), Ovis aries MIF (GI: NP_001072123) Homo sapiens MIF (GI: CAG46452), Ostertagia ostertagi MIF-1 (GI: AIE56464.1), T. circumcincta MIF-1 (GI: PIO59269.1), H. contortus MIF-1, Haemonchus placei MIF-1 (GI: VDO39061.1), A. ceylanicum MIF-1 (GI: ACC54555.1), Caenorhabditis elegans MIF-1 (GI: NP_499536.1), Trichuris trichiura MIF (GI: CAB46355), Trichinella spiralis MIF-1.1 (GI: XP_003378412.1), Trichinella spiralis MIF-1.2 (GI: CAB46354), Trichinella pseudospiralis MIF (GI: AAL12630), O. volvulus MIF-1 (GI: AAK66563), B. malayi MIF-1 (GI: U88035.1) and A. suum MIF-1 (GI: BAD24819) using the genetic distance model of Jukes-Canter by the neighbour joining method in Geneious Prime.
3.2. MIF-2 Structure
The predicted 3D structures of TcMIF-2 and HcMIF-2 and binding and catalytic sites over a wide range of ligands are shown in Figure 3. The protein structures of both TcMIF-2 and HcMIF-2 best fit superimposed structural models corresponding to the monomer of A. ceylanicum MIF-2 (Link to the structure 2OS5) [12]. For TcMIF-2, only one possible model was produced, with a C-score of 1.37, a TM value of 0.90 ± 0.06, a RMSD (root-mean-square deviation) of 1.7 ± 1.5 Å, with normalised z-scores less than 4.13. From the best five models obtained for HcMIF-2, the selected model had a C-score of 0.65, a TM value of 0.63 ± 0.14, a RMSD of 6.0 ± 3.7 Å, and normalised z-scores less than 2.51. For TcMIF-2, the catalytic and active site residues that fell within 4 Å of the substrate were Pro-2, Met-3, Lys-33, Pro-34, Glu-63, Ser-64, Ile-65, Leu-102, Val-107 and Val-114 (Figure 3F), whereas the corresponding residues in HcMIF-2 were Pro-2, Met-3, Lys-33, Arg-37, Glu-63, Ser-64, Ile-65, Leu-102, Val-107 and Val-114 (Figure 3G).

Figure 3.
The predicted tertiary structures of the TcMIF-2 and HcMIF-2 monomers. Location of the C- and N-terminus in the predicted tertiary structure of TcMIF-2 (A) and HcMIF-2 (B). (C) Superposition of the predicted tertiary structure of TcMIF-2 from T. circumcincta (blue) and HcMIF-2 from H. contortus (orange). Location of the active site (pink) within TcMIF-2 (D) and HcMIF-2 (E). (F) The active site of TcMIF-2 (blue) within 4 Å of the superimposed 2OS5 (yellow) with polar bonds also shown in magenta. (G) The active site of HcMIF-2 (green) within 4 Å of the superimposed 2OS5 (yellow) with polar bonds also shown in magenta.
3.3. Recombinant Protein Expression
Maximal production of functional recombinant MIF-2 was obtained in the E. coli strain BL21 (DE3) when expression was induced with 0.2% L-arabinose for 4 h at 30 °C. The purified C-terminal His-tagged recombinant TcMIF-2 and HcMIF-2 proteins appeared as single bands of about 16 kDa on SDS-PAGE (Figure 4). The presence of each of the His-tagged recombinant proteins was confirmed by Western blotting.

Figure 4.
SDS-PAGE of recombinant TcMIF-2 and HcMIF-2. Lane 1- Seeblue Plus2 Pre-stained standard (Invitrogen); lane 2- Unbound material post-passage on column; lane 3- Wash; lane 4- TcMIF-2; lane 5- HcMIF-2.
3.4. Enzyme Assays
The tautomerase activities of HcMIF-2 and TcMIF-2 were similar at 20 nmoles min−1mg−1 protein, using 20 µg/mL enzyme in the assay. HcMIF-2 activity was directly proportional to the amount of protein added over the range of 2–20 µg/mL (Figure 5A). The two batches of this purified protein had similar tautomerase activities. Storage at 4 °C, −20 °C or −70 °C reduced activity by approximately 50% (Figure 5B).
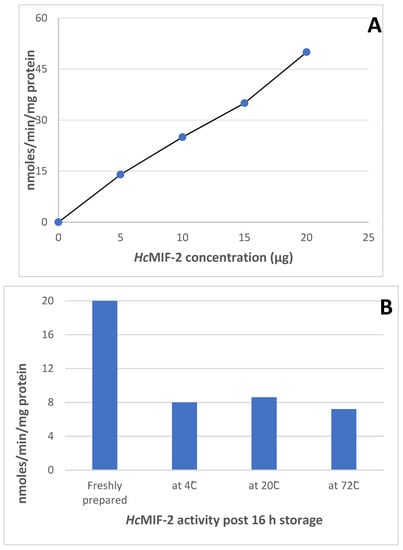
Figure 5.
The effect of using increasing concentrations of recombinant HcMIF-2 (A) and storage at different temperatures (B) on the protein activity (mean ± SD, n = 2). The activity was calculated by tautomerisation of the coloured dopachrome methyl ester to its colourless enol form, measured spectrophotometrically at 475 nm.
3.5. Host Recognition
Both TcMIF-2 and HcMIF-2 were recognised in an ELISA by antibodies in both serum and saliva collected from sheep exposed to nematodes in the field (Figure 6). No antibody was detected when serum or saliva samples from parasite-naïve animals was used.

Figure 6.
Recognition of TcMIF-2 and HcMIF-2 by serially diluted immune saliva (IgA) (A) and serum (IgG) (B) but not by parasite-naïve controls.
4. Discussion
This study showed the close structural and functional relationships between recombinant MIF-2 enzymes of the abomasal parasitic nematodes T. circumcincta and H. contortus. There was also similarity to homologues from several other nematodes. Both proteins were enzymatically typical of MIF-2 characterised from several helminths and were recognised by antibodies in both serum and saliva from field-immune sheep, but not nematode-naïve animals.
T. circumcincta and H. contortus MIF-2, designated TcMIF-2 and HcMIF-2, respectively, were shown to be 16 kDa proteins encoded by 342 bp full length cDNA sequences. The amino acid sequence of HcMIF-2 corresponded to the product of the gene designated as HCON_NP_LOC13552, located on chromosome 5 [31], the representative chromosome-level genome of the anthelmintic-susceptible H. contortus field strain isolated from pasture-grazed New Zealand sheep.
The two predicted 114 amino acid proteins had 84% homology (Figure 1), were closely related in a phylogenetic tree (Figure 2) and the 3D structures had almost identical active sites (Figure 1 and Figure 3). There was 54–76% homology of TcMIF-2 with MIF-2s of seven nematodes, but it was low with other MIF sequences, including the MIF-2 of the filarial nematodes B. malayi and O. volvulus. The MIF-2 proteins belonged to a different protein family from MIF-1 of the two nematodes; the MIF-1s were closely related to each other in the phylogenetic tree (Figure 2).
Approximately half of all proteins coded by genes in helminth genomes are currently of unknown function [32]. As a result of the search using PSI-BLAST against the Protein Data Bank (PDB) in this study, a putative function could be assigned to two of the top blast hits currently annotated as hypothetical protein (locus tag EYC32381 to EYC32384 of A. ceylanicum) and protein of unknown function (locus tag VDO97074 of H. polygyrus), with 65–75% and 80% identity, respectively.
The predicted TcMIF-2 and HcMIF-2 3-dimensional structures and binding and catalytic sites have been determined over a wide range of ligands and shown to be highly conserved (Figure 4). This was also true of many other nematode MIF-2 homologues (Figure 1), with only minor differences in the tautomerase binding site residues.
Given these almost identical active sites, it is unsurprising that the two recombinant enzymes also had quantitatively similar tautomerase activity in the standard assay in which coloured dopachrome methyl ester is changed to its colourless enol form [30]. Given the close similarity of the active sites, it is not surprising that the two recombinant enzymes had similar tautomerase activity in the standard assay in which coloured dopachrome methyl ester is changed to its colourless enol form [30]. Both recombinant TcMIF-2 and HcMIF-2 had similar activities of 20 nmoles min−1 mg−1 protein, using 20 µg/mL enzyme in the assay. Storage at 4 °C, −20 °C or −70 °C approximately halved tautomerase activity (Figure 5B). It appears that the position of proline at position 2 is cruciasl for tautomerase activity studies [15]. Both recombinant TcMIF-2 and HcMIF-2 have proline at position 2, but the measurable enzymatic activity was many-fold lower when compared with other parasites MIF-2 activity [15,16]. The reason might be the location of the parasite within the host and the % homology of recombinant TcMIF-2 and HcMIF-2 with other MIF-2 proteins. Human filarial parasite W. bancrofti was manyfold active, and this might be because of its location within the human lymphatic system. The tautomerase activity of O. volvulus might be higher than the recombinant proteins in this study because of significant differences in the amino acid homology, and O. volvulus shares only 29% similarity.
Antibodies in both serum and saliva from field-immune, but not nematode-naïve, sheep-recognised recombinant TcMIF-2 and HcMIF-2 in an ELISA (Figure 6). The recognition of both recombinant proteins by antibodies generated by exposure of sheep in the field to native MIF-2s indicates cross-antigenicity of the native and recombinant MIF-2s. This strongly indicates using MIF-2 as a vaccine candidate to control the parasites of ruminants. In addition, the quantitative similarity of recombinant TcMIF-2 and HcMIF-2 in the ELISA suggests that vaccines developed against antigens derived from one of the abomasal parasitic nematodes may also be protective against the other common parasite of sheep.
Author Contributions
Conceptualization, S.U., W.G. and J.S.K.; methodology, C.B. and S.B.C.; formal analysis, S.U., N.P. and J.S.K.; writing—S.U., J.S.K., C.B. and H.V.S.; supervision, W.G.; funding acquisition, W.G. and S.U. All authors have read and agreed to the published version of the manuscript.
Funding
The financial support of AGMARDT (Grant No. P14003) and Merck is gratefully acknowledged.
Institutional Review Board Statement
Use of experimental animals for culturing and harvesting adult worms for RNA extraction has been approved by the AgResearch Grasslands Animal Ethics Committee (protocol #13052).
Informed Consent Statement
Not applicable.
Data Availability Statement
The raw data can be made available on request.
Acknowledgments
The authors would like to thank Marian Price-Carter and Luis Carvalho for critically reviewing the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Blocki, F.A.; Ellis, L.B.M.; Wackett, L.P. MIF protein are theta-class glutathione S-transferase homologs. Protein Sci. 1993, 2, 2095–2102. [Google Scholar] [CrossRef] [PubMed]
- Leng, L.; Metz, C.N.; Fang, Y.; Xu, J.; Donnelly, S.; Baugh, J.; Delohery, T.; Chen, Y.; Mitchell, R.A.; Bucala, R. MIF signal transduction initiated by binding to CD74. J. Exp. Med. 2003, 197, 1467–1476. [Google Scholar] [CrossRef] [PubMed]
- Bloom, B.R.; Bennett, B. Mechanism of a reaction in vitro associated with delayed-type hypersensitivity. Science 1966, 153, 80–82. [Google Scholar] [CrossRef] [PubMed]
- David, J.R. Delayed hypersensitivity in vitro: Its mediation by cell-free substances formed by lymphoid cell-antigen interaction. Proc. Natl. Acad. Sci. USA 1966, 56, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Salminen, A.; Kaarniranta, K. Control of p53 and NF-κB signaling by WIP1 and MIF: Role in cellular senescence and organismal aging. Cell. Signal. 2011, 23, 747–752. [Google Scholar] [CrossRef]
- Sauler, M.; Bucala, R.; Lee, P.J. Role of macrophage migration inhibitory factor in age-related lung disease. Amer. J. Physiol. 2015, 309, L1–L10. [Google Scholar] [CrossRef]
- Xu, X.; Pang, J.; Chen, Y.; Bucala, R.; Zhang, Y.; Ren, J. Macrophage migration inhibitory factor (MIF) deficiency exacerbates aging-induced cardiac remodeling and dysfunction despite improved inflammation: Role of autophagy regulation. Sci. Rep. 2016, 6, 22488. [Google Scholar] [CrossRef]
- Sparkes, A.; De Baetselier, P.; Roelants, K.; De Trez, C.; Magez, S.; Van Ginderachter, J.A.; Raes, G.; Bucalaf, R.; Stijlemans, B. The non-mammalian MIF superfamily. Immunobiology 2017, 222, 473–482. [Google Scholar] [CrossRef]
- Pastrana, D.V.; Raghavan, N.; Fitzgerald, P.; Eisinger, S.W.; Metz, C.; Bucala, R.; Schleimer, R.P.; Bickel, C.; Scott, A.L. Filarial nematode parasites secrete a homologue of the human cytokine macrophage migration inhibitory factor. Infect. Immunit. 1998, 66, 5955–5963. [Google Scholar] [CrossRef]
- Vermeire, J.J.; Cho, Y.; Lolis, E.; Bucala, R.; Cappello, M. Orthologs of macrophage migration inhibitory factor from parasitic nematodes. Trend. Parasitol. 2008, 24, 355–363. [Google Scholar] [CrossRef]
- Nisbet, A.J.; Bell, N.E.; McNeilly, T.N.; Knox, D.P.; Maizels, R.M.; Meikle, L.I.; Wildblood, L.A.; Matthews, J.B. A macrophage migration inhibitory factor-like tautomerase from Teladorsagia circumcincta (Nematoda: Strongylida). Parasite Immunol. 2010, 32, 503–511. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.; Jones, B.F.; Vermeire, J.J.; Leng, L.; DiFedele, L.; Harrison, L.M.; Xiong, H.; Kwong, Y.K.; Chen, Y.; Bucala, R.; et al. Structural and functional characterisation of a secreted hookworm Macrophage Migration Inhibitory Factor (MIF) that interacts with the human MIF receptor CD74. J. Biol. Chem. 2007, 282, 23447–23456. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.; Hoti, S.L.; Meena, R.L.; Vasuki, V.; Sankari, T.; Kaliraj, P. Molecular and functional characterisation of macrophage migration inhibitory factor (MIF) homolog of human from lymphatic filarial parasite Wuchereria bancrofti. Parasitol. Res. 2012, 111, 2035–2047. [Google Scholar] [CrossRef] [PubMed]
- Younis, A.E.; Soblik, H.; Ajonina-Ekoti, I.; Erttmann, K.D.; Luersen, K.; Liebau, E.; Brattig, N.W. Characterisation of a secreted macrophage migration inhibitory factor homologue of the parasitic nematode Strongyloides acting at the parasite-host cell interface. Microbes Infect. 2012, 14, 279–289. [Google Scholar] [CrossRef] [PubMed]
- Ajonina-Ekoti, I.; Kurosinski, M.A.; Younis, A.E.; Ndjonka, D.; Tanyi, M.K.; Achukwi, M.; Eisenbarth, A.; Ajonina, C.; Lüersen, K.; Breloer, M.; et al. Comparative analysis of macrophage migration inhibitory factors (MIFs) from the parasitic nematode Onchocerca volvulus and the free-living nematode Caenorhabditis elegans. Parasitol. Res. 2013, 11, 3335–3346. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, A.; Sharma, R.; Hoti, S.L. Identification and biochemical characterisation of macrophage migration inhibitory factor-2 (MIF-2) homologue of human lymphatic filarial parasite, Wuchereria bancrofti. Act. Tropica 2015, 142, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Zang, X.; Taylor, P.; Wang, J.M.; Meyer, D.J.; Scott, A.L.; Walkinshaw, M.D.; Maizels, R.M. Homologues of Human Macrophage Migration Inhibitory Factor from a Parasitic Nematode. J. Biol. Chem. 2002, 277, 44261–44267. [Google Scholar] [CrossRef]
- Maizels, R.M.; Balic, A.; Gomez-Escobar, N.; Nair, M.; Taylor, M.D.; Allen, J.A. Helminth parasites—Masters of regulation. Immunol. Rev. 2004, 201, 89–116. [Google Scholar] [CrossRef]
- Matoušová, P.; Vokrál, I.; Lamka, J.; Skálová, L. The role of xenobiotic metabolising enzymes in anthelmintic deactivation and resistance in helminths. Trend. Parasitol. 2016, 32, 481–491. [Google Scholar] [CrossRef]
- Wang, Y.; Lu, M.; Wang, S.; Ehsan, M.; Yan, R.; Song, X.; Xu, L.; Li, X. Characterisation of a secreted macrophage migration inhibitory factor homologue of the parasitic nematode Haemonchus contortus acting at the parasite-host cell interface. Oncotarget 2017, 8, 40052–40064. [Google Scholar] [CrossRef]
- Marson, A.L.; Tarr, E.K.; Scott, A.L. Macrophage migration inhibitory factor (mif) transcription is significantly elevated in Caenorhabditis elegans dauer larvae. Gene 2001, 278, 53–62. [Google Scholar] [CrossRef]
- Umair, S.; Ria, C.; Knight, J.S.; Bland, R.J.; Simpson, H.V. Sarcosine metabolism in Haemonchus contortus and Teladorsagia circumcincta. Exp. Parasitol. 2013, 134, 1–6. [Google Scholar] [CrossRef]
- Knight, J.S.; Broadwell, A.H.; Grant, W.N.; Shoemaker, C.B. A strategy for shuffling numerous Bacillus thuringiensis crystal protein domains. J. Econ. Entomol. 2004, 9, 1805–1813. [Google Scholar] [CrossRef]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef]
- Yang, J.; Yan, R.; Roy, A.; Xu, D.; Poisson, J.; Zhang, Y. The I-TASSER Suite: Protein structure and function prediction. Nat. Method. 2015, 12, 7–8. [Google Scholar] [CrossRef]
- Laskowski, R.A.; Rullmann, J.A.C.; MacArthur, M.W.; Kaptein, R.; Thornton, J.M. AQUA and PROCHECK-NMR: Programs for checking the quality of protein structures solved by NMR. J. Biomol. NMR 2004, 8, 477–486. [Google Scholar] [CrossRef]
- Wiederstein, M.; Sippl, M.J. ProSA-web: Interactive web service for the recognition of errors in three-dimensional structures of proteins. Nucleic Acid. Res. 2007, 35, W407–W410. [Google Scholar] [CrossRef]
- Schrodinger, L.L.C. The PyMOL Molecular Graphics System. Available online: https://pymol.org/2/ (accessed on 28 March 2022).
- Love, C.A.; Lilley, P.E.; Dixon, N.E. Stable high-copy-number bacteriophage lambda promoter vectors for overproduction of proteins in Escherichia coli. Gene 1996, 176, 49–53. [Google Scholar] [CrossRef]
- Swope, M.; Sun, H.-W.; Blake, P.R.; Lolis, E. Direct link between cytokine activity and a catalytic site for macrophage migration inhibitory factor. EMBO J. 1998, 17, 3534–3541. [Google Scholar] [CrossRef]
- Palevich, N.; Maclean, P.H.; Baten, A.; Scott, R.W.; Leathwick, D.M. The Genome Sequence of the Anthelmintic-Susceptible New Zealand Haemonchus contortus. Genome Biol. Evol. 2019, 11, 1965–1970. [Google Scholar] [CrossRef]
- Palevich, N.; Britton, C.; Kamenetzky, L.; Mitreva, M.; de Moraes Mourão, M.; Slatko, B.E. Tackling hypotheticals in helminth genomes. Trends Parasitol. 2018, 34, 179–183. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).