
Evaluation of 5-Aminolevulenic Acid Plus Ferrous Ion for Its Potential to Improve Symptoms of Bovine Babesiosis


 ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Results
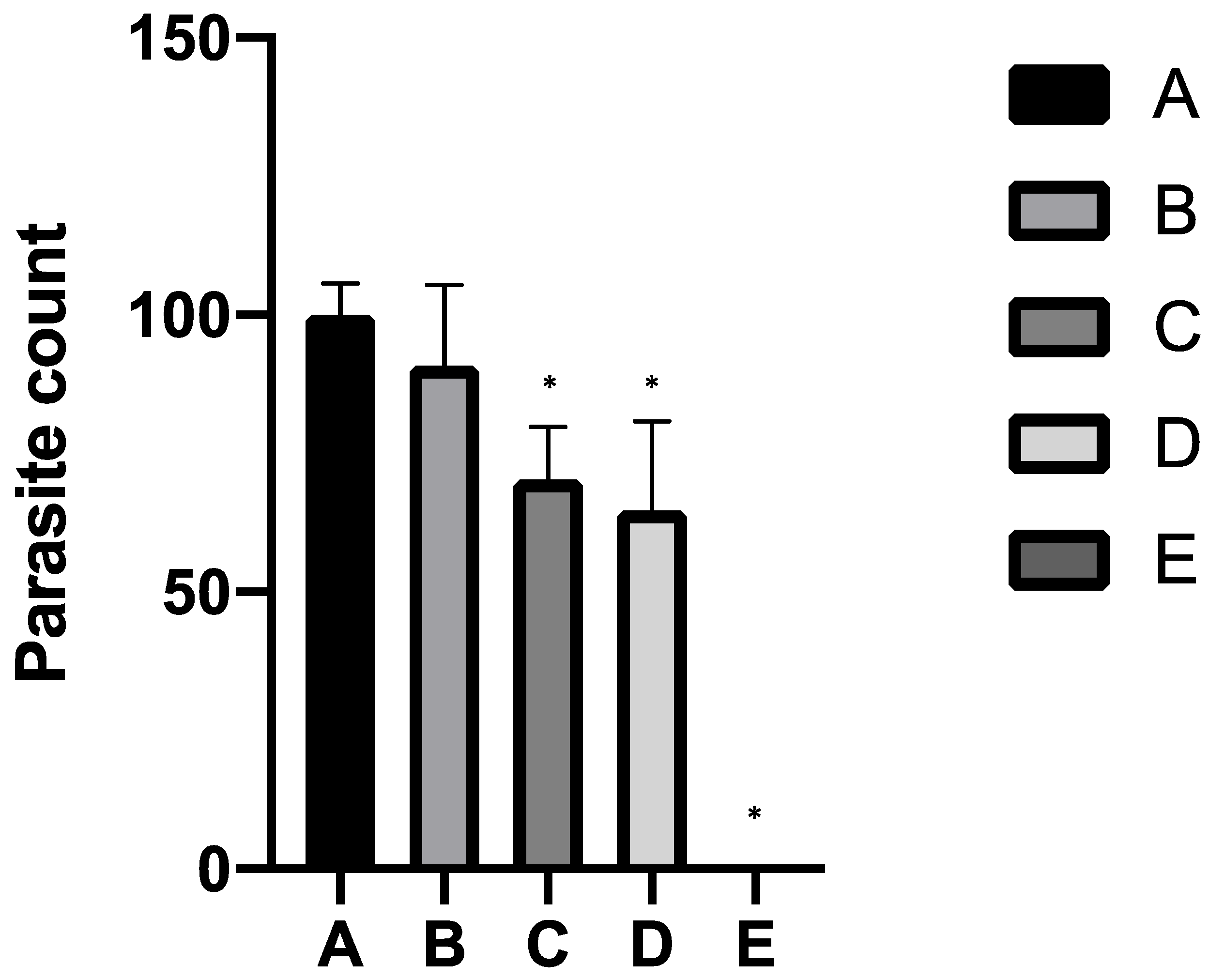
2.1. In Vitro Study
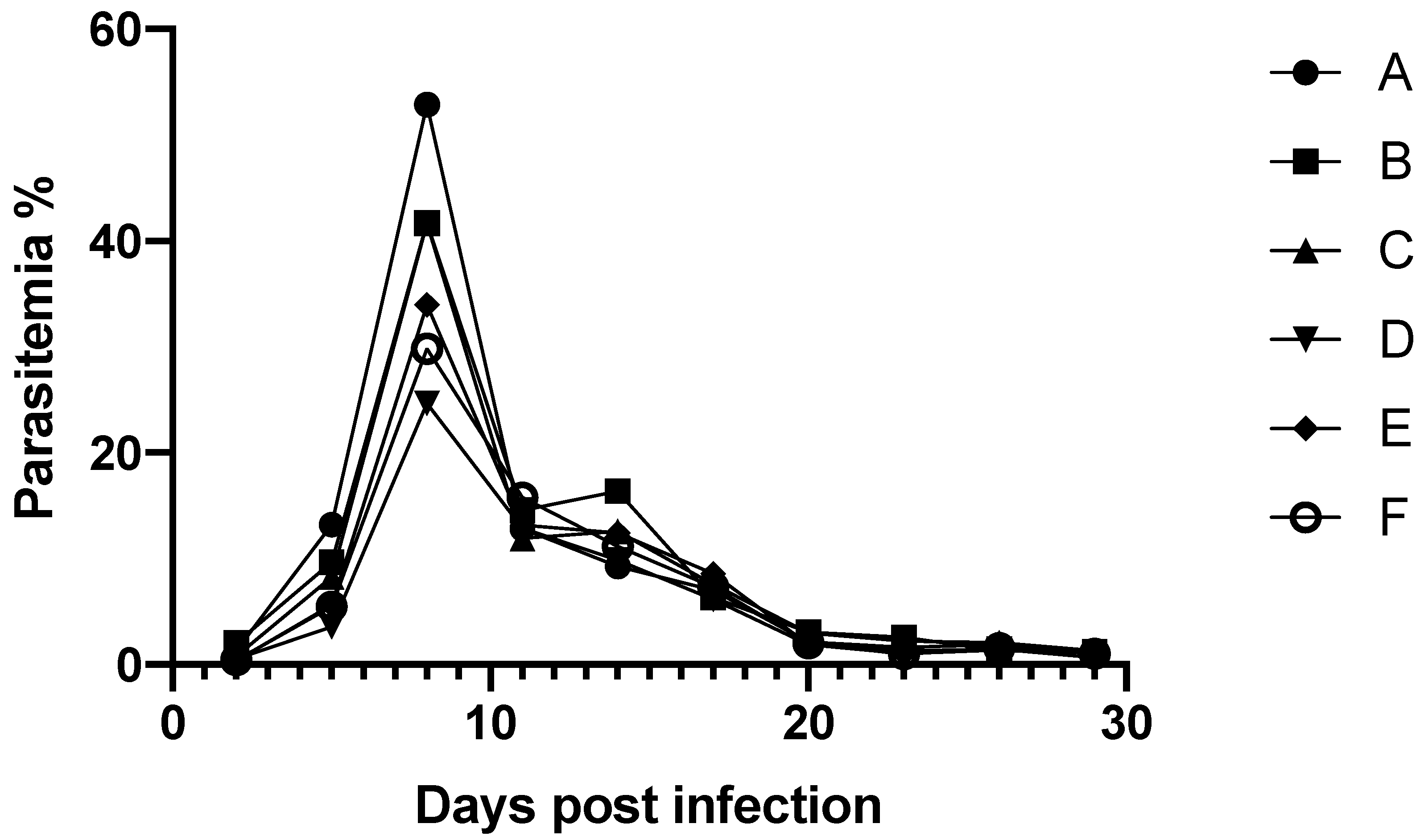
2.2. In Vivo Study
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Parasite Culture and Growth Inhibition Assay
4.3. Animals
4.4. Parasite Infection and Animal Experimentations
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Florin-Christensen, M.; Suarez, C.E.; Rodriguez, A.E.; Flores, D.A.; Schnittger, L. Vaccines against bovine babesiosis: Where we are now and possible roads ahead. Parasitology 2014, 141, 1563–1592. [Google Scholar] [CrossRef] [PubMed]
- Gray, J.; Zintl, A.; Hildebrandt, A.; Hunfeld, K.-P.; Weiss, L. Zoonotic babesiosis: Overview of the disease and novel aspects of pathogen identity. Ticks Tick Borne Dis. 2010, 1, 3–10. [Google Scholar] [CrossRef]
- Guswanto, A.; Nugraha, A.B.; Tuvshintulga, B.; Tayebwa, D.S.; Rizk, M.A.; Batiha, G.E.; Gantuya, S.; Sivakumar, T.; Yokoyama, N.; Igarashi, I. 17-DMAG inhibits the multiplication of several Babesia species and Theileria equi on in vitro cultures, and Babesia microti in mice. Int. J. Parasitol. Drugs Drug Resist. 2018, 8, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Goo, Y.K.; Terkawi, M.A.; Jia, H.; Aboge, G.O.; Ooka, H.; Nelson, B.; Kim, S.; Sunaga, F.; Namikawa, K.; Igarashi, I.; et al. Artesunate, a potential drug for treatment of Babesia infection. Parasitol. Int. 2010, 59, 481–486. [Google Scholar] [CrossRef] [PubMed]
- Hakimi, H.; Asada, M.; Kawazu, S. Recent advances in molecular genetics tools for Babesia. Vet. Sci. 2021, 8, 222. [Google Scholar] [CrossRef]
- Garg, A.; Stein, A.; Zhao, W.; Dwivedi, A.; Frutos, R.; Cornillot, E.; Ben Mamoun, C. Sequence and annotation of the apicoplast genome of the human pathogen Babesia microti. PLoS ONE 2014, 9, e107939. [Google Scholar] [CrossRef] [PubMed]
- Sheiner, L.; Vaidya, A.B.; McFadden, G.I. The metabolic roles of the endosymbiotic organelles of Toxoplasma and Plasmodium spp. Curr. Opin. Microbiol. 2013, 16, 452–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez, M.H.; Rodriguez, B.L.; Shintani, T.T. Use of the dietary supplement 5-aminiolevulinic acid (5-ALA) and its relationship with glucose levels and hemoglobin A1C among individuals with prediabetes. Clin. Transl. Sci. 2012, 5, 314–320. [Google Scholar]
- Suzuki, S.; Hikosaka, K.; Balogun, E.O.; Komatsuya, K.; Niikura, M.; Kobayashi, F.; Takahashi, K.; Tanaka, T.; Nakajima, M.; Kita, K. In vivo curative and protective potential of orally administered 5-aminolevulinic acid plus ferrous ion against malaria. Antimicrob. Agents Chemother. 2015, 59, 6960–6967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komatsuya, K.; Hata, M.; Balogun, E.; Hikosaka, K.; Suzuki, S.; Takahashi, K.; Tanaka, T.; Nakajima, M.; Ogura, S.; Sato, S.; et al. Synergy of ferrous ion on 5-aminolevulinic acid-mediated growth inhibition of Plasmodium falciparum. J. Biochem. 2013, 154, 501–504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brayton, K.A.; Lau, A.O.; Herndon, D.R.; Hannick, L.; Kappmeyer, L.S.; Berens, S.J.; Bidwell, S.L.; Brown, W.C.; Crabtree, J.; Fadrosh, D.; et al. Genome sequence of Babesia bovis and comparative analysis of apicomplexan hemoprotozoa. PLoS Pathog. 2007, 3, 1401–1413. [Google Scholar] [CrossRef] [PubMed]
- Fleige, T.; Limenitakis, J.; Soldati-Favre, D. Apicoplast: Keep it or leave it. Microbes Infect. 2010, 12, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Takano, T.; Satoh, K.; Doki, T. Possible antiviral activity of 5-aminolevulinic acid in feline infectious peritonitis virus (feline coronavirus) infection. Front. Vet. Sci. 2021, 8, 647189. [Google Scholar] [CrossRef] [PubMed]
- Hendawy, A.O.; Shirai, M.; Takeya, H.; Sugimura, S.; Miyanari, S.; Taniguchi, S.; Sato, K. Effects of 5-aminolevulinic acid supplementation on milk production, iron status, and immune response of dairy cows. J. Dairy Sci. 2019, 102, 11009–11015. [Google Scholar] [CrossRef] [PubMed]
- Bork, S.; Okamura, M.; Matsuo, T.; Kumar, S.; Yokoyama, N.; Igarashi, I. Host serum modifies the drug susceptibility of Babesia bovis in vitro. Parasitology 2005, 130, 489–492. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Group | Dose (mg/kg) of * | Parameters | |||||
|---|---|---|---|---|---|---|---|
| ALA | SFC | PE | BW | RBC | HGB | HCT | |
| A ** | |||||||
| B | 10 | 8 dpi | 11, 17, 20, 23, 26, and 29 dpi | ||||
| C | 100 | 8 dpi | |||||
| D | 4 | 0.4 | 5 and 8 dpi | 8 dpi | 8 dpi | 8 dpi | 8 dpi |
| E | 20 | 2 | 8 dpi | 8 dpi | |||
| F | 100 | 10 | 8 dpi | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elata, A.; Suganuma, K.; Mossaad, E.; Zafar, I.; Galon, E.M.; Yoshimoto, K.; Tanaka, T.; Xuenan, X.; Kawazu, S.-I. Evaluation of 5-Aminolevulenic Acid Plus Ferrous Ion for Its Potential to Improve Symptoms of Bovine Babesiosis. Parasitologia 2021, 1, 218-224. https://doi.org/10.3390/parasitologia1040023
Elata A, Suganuma K, Mossaad E, Zafar I, Galon EM, Yoshimoto K, Tanaka T, Xuenan X, Kawazu S-I. Evaluation of 5-Aminolevulenic Acid Plus Ferrous Ion for Its Potential to Improve Symptoms of Bovine Babesiosis. Parasitologia. 2021; 1(4):218-224. https://doi.org/10.3390/parasitologia1040023
Chicago/Turabian StyleElata, Afraa, Keisuke Suganuma, Ehab Mossaad, Iqra Zafar, Eloiza May Galon, Keiichiro Yoshimoto, Tohru Tanaka, Xuan Xuenan, and Shin-Ichiro Kawazu. 2021. "Evaluation of 5-Aminolevulenic Acid Plus Ferrous Ion for Its Potential to Improve Symptoms of Bovine Babesiosis" Parasitologia 1, no. 4: 218-224. https://doi.org/10.3390/parasitologia1040023
APA StyleElata, A., Suganuma, K., Mossaad, E., Zafar, I., Galon, E. M., Yoshimoto, K., Tanaka, T., Xuenan, X., & Kawazu, S.-I. (2021). Evaluation of 5-Aminolevulenic Acid Plus Ferrous Ion for Its Potential to Improve Symptoms of Bovine Babesiosis. Parasitologia, 1(4), 218-224. https://doi.org/10.3390/parasitologia1040023