Abstract
(1) Background: Scavenger activity can pose significant challenges in forensic investigations. When examining damage on a victim, forensic investigators must first determine whether the damage resulted from human actions or animal scavenging. Accordingly, knowledge of the scavenger populations inhabiting a specific region and their scavenging behaviors is essential for accurately reconstructing the environmental context of the remains and interpreting associated taphonomic evidence in forensic investigations. (2) Methods: This study investigates the scavenging behavior of bobcats (Lynx rufus), a species native to North America, under controlled experimental conditions. The authors placed the carcasses of seven adult white-tailed deer in an outdoor field in Murfreesboro, Tennessee, and systematically documented the scavenging activities. (3) Results: Bobcats were observed to primarily scavenge soft tissue from the torso, focusing on musculature while generally avoiding bony regions. When unsatisfied with the torso, they shifted attention to the upper limbs, with minimal interest shown in the head or neck. Bobcats also exhibited a tendency to feed in a single area for extended periods, leading to concentrated damage in specific locations on the remains. (4) Conclusions: The findings demonstrate that bobcats can significantly influence the postmortem conditions of remains, producing characteristic patterns of modification. Recognizing these patterns is essential for forensic investigators and examiners when interpreting scavenger-related alterations in forensic contexts.
1. Introduction
Scavengers play a significant role in forensic contexts, particularly in outdoor crime scenes where remains are vulnerable to animal interference. Scavenging activity can obscure or mimic perimortem trauma, complicating the interpretation of injuries [1,2,3]. When examining a body with trauma, forensic investigators must therefore distinguish between damage caused by human actions and that resulting from scavenger activity. This assessment requires knowledge of the local scavenger populations and their feeding behaviors, which is essential for accurately reconstructing the depositional context and interpreting associated taphonomic evidence [1,4,5].
This study aims to investigate the scavenging behavior of bobcats (Lynx rufus), a native North American species, under controlled conditions. Bobcats are well known for their proficient hunting abilities and predominantly prey on birds, rodents, white-tailed deer, rabbits, and hares, while occasionally feeding on raccoons, opossums, skunks, and foxes [6,7,8]. As crepuscular predators, they exhibit peak activity during dawn and dusk, hunting primarily by stealthily approaching their prey and attacking the neck to sever the spinal cord of their prey. Notably, bobcats scent-mark areas where they cache larger prey, likely as a means to deter other scavengers while they are away, returning later to feed more [9].
Although bobcats are widely recognized as proficient hunters, recent studies have documented their engagement in scavenging behavior. Reported instances include the scavenging of human cadavers [9,10] and ruffed grouse carcasses [11]. Specifically, Rippley and colleagues [9] captured bobcat scavenging activities on a human cadaver using a motion-activated camera, reporting that their limb-targeted scavenging pattern and covering behavior with leaf debris are unique observations of bobcat scavenging. Additionally, analysis of the stomach contents of a road-killed bobcat revealed the remains of a grey squirrel containing blowfly eggs, indicating the squirrel was already dead when consumed [12]. While younger bobcats may scavenge more frequently due to their underdeveloped hunting skills [9], observations of scavenging among adult bobcats are increasing. Although bobcats are known to feed on roadkill and wounded animals, they demonstrate a strong preference for fresh meat and typically cease feeding once decomposition advances [9,13]. The scavenger community in Middle Tennessee has been characterized by species such as vultures (Cathartes and Coragyps genera), coyotes (Canis latrans), opossums (Didelphis virginiana), and raccoons (Procyon lotor); however, recent studies have also documented scavenging activity by bobcats in this region [14].
The purpose of this study is to investigate the scavenging behavior of bobcats in the Middle Tennessee area from a forensic perspective, using seven white-tailed deer carcasses as proxies for human remains. Specifically, this study analyzes the initiation and duration of scavenging, seasonal and diurnal variations in scavenging frequency, feeding preferences for specific body regions, and the modification patterns observed in soft and hard tissues. The findings aim to improve forensic investigators’ ability to differentiate bobcat-induced markings and alterations from human-inflicted trauma, thereby enhancing the accuracy of crime scene reconstructions. Ultimately, the study will contribute to a broader understanding of taphonomic alterations involving bobcats, providing valuable insights for the forensic community as a whole.
2. Materials and Methods
Seven adult white-tailed deer (Odocoileus virginianus) roadkill specimens were used in this study. The carcasses were salvaged between January 2022 and April 2023 under scientific permits issued by the Tennessee Wildlife Resources Agency. All specimens were retrieved from roadways within 24 h postmortem and transported to the research facility. Although all carcasses were in the fresh stage of decomposition with comparable postmortem intervals, their preservation conditions varied, particularly with respect to trauma. These differences were influenced by the severity of the perimortem collisions and any postmortem scavenger activity prior to collection. Notably, one carcass (Specimen #6) was recovered without a head, presumably decapitated post-collision by someone seeking antlers. Table 1 outlines the initial condition of each specimen at the time of salvage.

Table 1.
Collection dates and initial visible conditions of specimens.
Each specimen was wrapped in a tarp and transported directly to a designated plot within the MTSU Outdoor Forensic Facility (MOFF), a 12-acre outdoor decomposition research facility managed by the MTSU Forensic Science Program. The MOFF is located in the rural northwest region of Murfreesboro, Tennessee, at an elevation of approximately 186 m above sea level. The facility primarily consists of flat grasslands interspersed with various broadleaf tree species, including oak, hickory, maple, and beech. Each plot measured 238.8 cm in length and 170.2 cm in width, with a uniform fence height of 172.7 cm and an apex height of 221.0 cm at the center (Figure 1). Plots were spaced 5 to 10 m apart. The lower perimeter of each enclosure was constructed from flexible chicken wire, 76.2 cm tall. Although the fencing was not designed to allow access, evidence indicated that some animals were able to dig beneath it to reach the carcasses. Scavenger activity and carcass taphonomy were monitored using motion-activated trail cameras (Vikeri, model# A1) positioned at three locations around each plot. One camera was mounted directly above the specimens, secured to the apex of the fencing, while a second camera was placed diagonally at a lower corner of the enclosure (Figure 1). A third camera, positioned approximately 1 m outside the plot, captured activity occurring in the surrounding area that might not have been recorded by the other cameras. Cameras were programmed to capture bursts of 10 photographs upon detecting motion, with a 5-s delay between bursts, continuing until the motion source exited the frame. Except for the two specimens removed by coyotes midway through the experiment (see Results section), all other specimens remained within the plots until complete skeletonization, which occurred over approximately 10 months. It is noteworthy that multiple specimens were often present at the MOFF simultaneously; however, due to variations in placement dates, their decomposition stages differed. The decomposition stage of each specimen was classified according to the five-stage system: fresh, bloat, active decay, advanced decay, and dry/skeleton [15].

Figure 1.
Design of a plot used to encase a specimen. Red circles indicate the positions of the motion-activated trail cameras.
Additionally, a weather monitoring device (AcuRite®, model# 01528MCB) installed at the site recorded environmental data, including temperature, humidity, rainfall, and wind speed, at 12 min intervals. The recorded weather data were synchronized with the timestamps of bobcat appearances captured by the cameras. Camera memory cards were collected weekly, and all images were transferred to a laptop for review. The date and time of each bobcat appearance were logged into an Excel spreadsheet alongside the corresponding weather data.
Bobcat behavior was categorized into five types, following Rippley and colleagues [9]: feeding, covering/scent-marking, resting, appearing with no activity, and unknown. Notably, the ‘feeding’ category was assigned only when photographs clearly depicted the removal or tearing of the specimen’s soft tissues. Each behavior was further classified as occurring during peak hours (04:00–10:00 and 18:00–24:00) or non-peak hours (0:00–03:59 and 10:01–17:59), as proposed by Buie and colleagues [16], to validate these timeframes. Upon completion of data collection, a descriptive analysis was conducted to determine the frequency of each behavioral category, with results organized into a table based on the defined timeframes.
3. Results
According to the Köppen–Geiger climate classification system [17], Murfreesboro, Tennessee, is situated within a humid subtropical climate zone (Cfa). Summarized weather data at the MOFF during the study period is presented in Table S1.
3.1. Descriptive Observations of Bobcat Activities to Each Specimen
3.1.1. Specimen #1
A total of eleven bobcat occurrences were documented, with feeding behavior observed in four instances. Bobcats engaged in scavenging activity while the specimen remained in the fresh stage of decomposition, primarily consuming the abdominal region and hindlimb. Notably, two instances involved simultaneous feeding by two bobcats (Figure 2a). The final recorded bobcat appearance occurred on 6 February 2022, at which point the specimen had progressed to the active decay stage.

Figure 2.
Representative images of bobcat scavenging behaviors observed during the study. (a) Primary bobcat feeding on the central abdominal region of Specimen #1, while a secondary bobcat rests and observes (red circle); (b) bobcat scent-marking vegetation in the corner of Specimen #3’s enclosure (red circle); (c) bobcat covering Specimen #5 to conceal it from other potential scavengers; (d) bobcat dragging Specimen #6 towards itself, while a disturbed opossum observes cautiously from a distance (red circle).
3.1.2. Specimen #2
No bobcat appearances were recorded for this specimen. However, scavenging activity by other animals was observed, indicating that the specimen did not display deterrent factors that would inhibit bobcat access. Approximately one month after placement, Specimen #2 was removed from the enclosure by coyotes while in the advanced decay stage of decomposition, resulting in the termination of the research for this specimen.
3.1.3. Specimen #3
A total of fourteen bobcat appearances were recorded for this specimen, with feeding behavior observed during the majority of encounters, along with single instances of investigative behavior and scent-marking. The first bobcat appearance occurred four days after placement, when the specimen was in the bloat stage with all musculature still visually red and fresh. Initial feeding activity targeted the upper hindlimb, followed by the forelimbs and torso. Over the subsequent three days, eleven additional feedings were recorded, each lasting approximately an hour or less and primarily focused on the torso. Scent-marking behavior was also observed during this period (Figure 2b). The final feeding occurred one week later, when the specimen had progressed to the active decay stage. Notably, feeding occurred consistently over a concentrated three-day period, with bobcat appearances recorded across various times of day.
3.1.4. Specimen #4
A total of seven bobcat appearances were recorded, but the bobcats showed no interest in this specimen and simply inspected the fence before leaving. When the bobcats appeared for the last time, the specimen was in the advanced decay state of decomposition.
3.1.5. Specimen #5
A total of seventeen bobcat appearances were recorded for this specimen. Of these, eight involved feeding, three involved covering behavior, three showed no specific activity, and three involved resting. The first appearance occurred one week after placement, during which the bobcat fed on the left forelimb for approximately 15 min before covering the carcass to conceal it from other scavengers (Figure 2c). Scavenging activity continued over the following three weeks, during which the specimen progressed to the advanced decay stage of decomposition.
3.1.6. Specimen #6
The specimen was first visited by a bobcat on the day of placement and recorded a total of twenty-six appearances, the highest number among all specimens in this study. During the initial appearance, the bobcat inspected the carcass, exhibited scent-marking behavior, and fed on the lower abdominal region for approximately two hours. Feeding activity continued for about one month, culminating when the specimen reached the advanced decay stage of decomposition. Notably, one observation captured an incidental encounter between the bobcat and an opossum; the two species did not scavenge simultaneously, as the opossum vacated the area upon the bobcat’s approach (Figure 2d). Approximately half of the bobcat appearances involved feeding, while four instances showed no discernible activity.
3.1.7. Specimen #7
The specimen was not visited by bobcats, likely due to rapid decomposition caused by vulture activity. Vulture scavenging began approximately one week after placement, significantly accelerating skeletonization and disturbing the remains, thereby reducing opportunities for bobcat access.
To summarize, excluding the two specimens that did not receive any bobcat visits, bobcats were observed visiting individual carcasses between seven and twenty-six times. The initial bobcat visit occurred as early as the first day post-placement (Specimen #6) and continued for up to 27 days (Specimens #5 and #6). At the time of the final recorded bobcat appearance, the decomposition stage of each specimen ranged from active decay to advanced decay. The frequency, onset, and duration of bobcat visits, along with the decomposition stages at the time of the first and last visits, are presented in Table 2.
Table 2.
Onset, duration, and decomposition stages associated with bobcat scavenging activity per specimen.
3.2. Bobcats’ Scavenging Behaviors
3.2.1. Timing and Duration of Bobcat Appearances
The research findings indicate that bobcats are frequent scavengers in the Middle Tennessee region. Over the course of the research, bobcats visited five out of seven specimens, with a total of 71 appearances recorded by the cameras (Table 2). As shown in Table 2, bobcats appeared within 1 to 8 days after specimen placement, except for Specimen #4, where the first appearance occurred 41 days post-placement. Once a carcass was detected, bobcats continued to revisit the site for periods ranging from 21 to 46 days, except in the case of Specimen #3, where bobcat appearance ceased 4 days after initial scavenging. Scavenging typically commenced when specimens were in the fresh or bloat stages of decomposition, with the exception of Specimen #4, which was first visited during the active decay stage. Bobcat activity generally ceased when the specimens had reached either the active decay or advanced decay stage. Regarding scavenging behavior categorized as ‘feeding’, all bobcats started feeding on the specimens when they were in the fresh or bloat stages and continued while visually fresh meat was available, regardless of the overall decomposition stage. Additionally, no noticeable differences were observed in the onset or duration of scavenging between specimens that exhibited visible trauma and those without trauma at the time of placement.
In terms of monthly frequency, bobcat appearances were most frequent in January, accounting for 31 out of the total 71 recorded appearances (43.7%). February followed with 26 occurrences (36.6%), and November recorded 14 occurrences (19.7%). The absence of bobcat activity during other months may be attributed to the lack of fresh meaty elements and viscera on the carcasses, as specimens exposed in March and from June through August had largely decomposed to dry tissue and bone. However, it is more plausible that bobcats increase their reliance on scavenging during cold months, when food resources are relatively scarce and competition with invertebrate scavengers is reduced [18]. Indeed, no bobcat scavenging activity was recorded in April, May, September, and October, despite the availability of substantial fresh tissue on specimens during these months.
Bobcat activity was observed across a range of temperatures ranging between −9.4 °C and 25 °C. The highest frequency of appearances occurred when temperatures ranged between 5 °C and 10 °C, accounting for 17 appearances (23.9%), followed by 16 appearances (22.5%) recorded at temperatures between 10 °C and 15 °C (Figure 3).
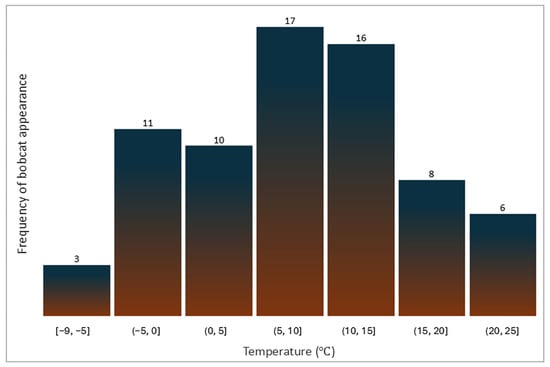
Figure 3.
Distribution of temperature ranges at which bobcat appearances were observed.
Analysis of visit duration supports the idea that bobcats primarily engage in scavenging for brief periods. Of the 71 recorded appearances, 35 visits (49%) lasted less than 10 min. The next most frequent duration, ranging from 30 min to 1 h, was observed 16 times (23%). Visits lasting between 1 and 5 h occurred in 11 instances (15%). The least common duration, ranging from 10 to 30 min, was recorded 9 times (13%) (Figure 4).
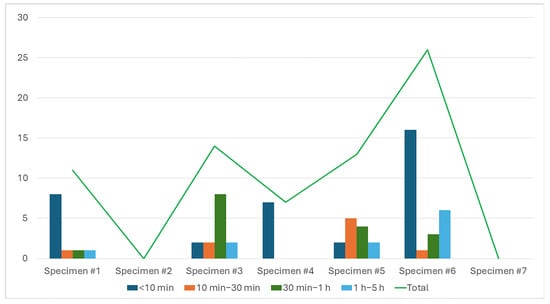
Figure 4.
Duration of bobcat visit on each specimen. Bar height represents the frequency distribution of visit durations, while the green line indicates the total number of visits per specimen.
Analysis of bobcat appearances by time of day indicates a clear preference for nocturnal scavenging. The highest frequency of appearances occurred between 20:00 and 21:00, with 8 recorded sightings. The next most common time periods were 18:00–19:00 and 22:00–23:00, each with 7 sightings. Outside these hours, bobcat appearances were sporadic but consistently occurred during nocturnal or crepuscular periods. With the exception of 2 outliers recorded at 11:00–12:00 and 15:00–16:00, all appearances occurred between 17:00 and 09:00. In total, 49 of the 71 appearances (69%) occurred during the peak hours defined by Buie and colleagues [16] (04:00–10:00 and 18:00–24:00). However, based on the observed data, revised peak hours were established to better reflect bobcat activity patterns in this study. The revised peak hours, defined as 17:00–07:00, encompassed 64 of the 71 bobcat appearances (90.1%). Only 7 appearances occurred outside this revised timeframe, supporting the validity of the adjusted structure.
3.2.2. Frequency of Distinct Scavenging Behaviors
During the 71 recorded bobcat visits, a total of 78 distinct behaviors were documented (Table 3). The number of observed behaviors exceeds the total number of appearances due to instances involving multiple bobcats or a single bobcat engaging in multiple actions during a single visit. Feeding behavior was the most frequently observed, accounting for 38 instances (48.7%). In 27 instances (34.6%), bobcats were present but did not physically interact with the specimens, instead appearing to inspect the carcasses or patrol the perimeter. Covering behavior or scent-marking within the enclosure was documented 5 times (6.4%), and resting behavior was observed on 5 occasions (6.4%). In 3 instances (3.8%), the bobcats’ activities could not be clearly determined from the camera footage and were categorized as ‘Unknown’.
Table 3.
Frequency of distinct bobcat scavenging behaviors.
3.2.3. Feeding Pattern
To assess whether bobcats exhibited preferences for specific body regions, their feeding behavior was analyzed based on activity observed in 5 regions of the specimens: head, neck, torso, forelimbs, and hindlimbs. The analysis revealed that a total of 38 feeding events involved the consumption of 61 body regions, indicating that bobcats frequently fed on multiple body regions during a single appearance.
Bobcats demonstrated a strong preference for the torso, which accounted for 32 out of the 61 recorded feedings (52.5%) (Table 4). This preference is expected, as the torso contains a substantial amount of soft tissue and musculature, making it more accessible and easier to consume compared to the bony extremities such as the head or distal limbs. Additionally, the torso likely serves as a consistent food source, particularly when time is limited, as it is more likely to contain unconsumed edible tissue [5,19]. It is noteworthy that in 21 of the 32 torso feedings (65.6%), additional body region(s) were also targeted during the same event. The anatomical centrality of the torso may facilitate transitions from adjacent regions, contributing to its higher frequency of consumption. The forelimbs were the second most frequently scavenged body region, with 14 recorded feedings (23.0%), followed by the hindlimbs with 9 feedings (14.8%). The neck was targeted less frequently, with only 6 feeding events (9.8%). No instances of feeding in the head region were documented throughout the study. Overall, the duration of feeding events for each body region was proportional to its relative frequency of occurrence (Table 4).
Table 4.
Frequency and duration of bobcat feedings per body region.
In their feeding behavior, bobcats typically initiated scavenging by clawing and licking the upper portion of the forelimb or hindlimb near the torso. They subsequently used their teeth to puncture a small hole in the skin. Beginning at this initial opening, the bobcats continued to lick and bite, often shaking their heads to gradually enlarge the hole, extending toward both the limbs and the torso. Their carnassial teeth were frequently employed to cut and tear the skin and underlying flesh. While standing on their hind legs for support, bobcats gripped the specimen’s flesh with their mouths and pulled it towards their bodies to tear off pieces of meat. However, this pulling behavior was not forceful enough to result in significant repositioning of the carcass. The process of creating the initial opening typically took approximately 10 min, after which the bobcats proceeded to widen it more rapidly (Figure 5).

Figure 5.
Sequential captures of bobcat feeding behavior on the forelimb and torso of Specimen #3. The bobcat approached the specimen (a), initially targeted the upper portion of the forelimb (b,c) and subsequently expanded the feeding area to include the torso (d).
Bobcat scavenging consistently resulted in extensive damage to the soft tissues of the specimens. Most muscles on the affected body regions were consumed, with some tendons and ligaments left behind. Scratch marks, oriented in random directions, were identifiable on the fur, primarily concentrated on the torso. The overall shape of the tissue damage was rounded, with relatively clean and smooth margins rather than ragged edges (Figure 6). In some cases, articular cartilage exhibited faint tooth marks, attributed to the bobcats’ repetitive chewing motions used to remove attached musculature. However, once the specimens were fully skeletonized and the cartilage was removed, no identifiable tooth marks or other modifications were observed on the bones.
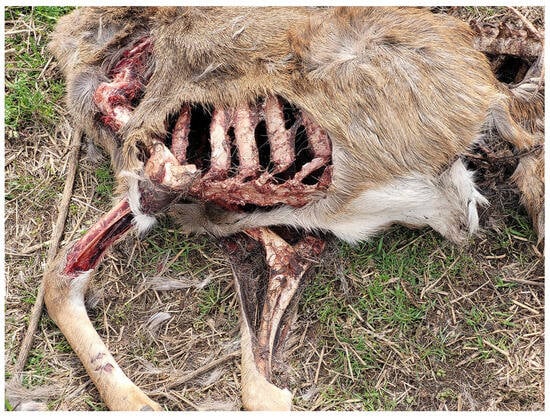
Figure 6.
Soft tissue damage resulting from bobcat scavenging. Note that the scavenging typically produces rounded, extensive areas of damage with clean, smooth margins.
4. Discussion
This study aimed to observe bobcat scavenging behaviors from a forensic perspective using white-tailed deer carcasses. As reported by Fain and Jeong [14], vultures, coyotes, opossums, and raccoons were also observed scavenging in this area. However, direct interactions between these species and bobcats were not recorded, with the exception of a single incident involving Specimen #6 (Figure 2d). Given that vultures are primarily diurnal, their activity likely did not overlap with that of bobcats, which are predominantly crepuscular or nocturnal. Although other scavengers exhibited temporal overlap in activity, encounters between bobcats and these species within the same plot were rarely observed. This may suggest that, when bobcats were actively scavenging, other animals either remained beyond the detection range of the cameras, possibly detecting the bobcat’s presence and choosing to wait, or actively avoided the vicinity.
The use of animal models allowed the study to circumvent various practical and legal constraints associated with the use of human remains, while securing a relatively large sample size (n = 7). However, the use of deer carcasses raises questions regarding the direct applicability of the findings to human remains. Although deer and humans have comparable overall body weights, there are notable anatomical and physiological differences, including quadrupedal locomotion, herbivorous diet, and the presence of thicker skin and fur in deer. These differences may influence decomposition patterns and, consequently, bobcat scavenging behavior, as bobcats tend to prefer fresh-appearing, red-colored muscle tissues [9]. To the authors’ knowledge, no direct comparative studies between deer and human decomposition exist. However, findings by Dautartas and colleagues [20] demonstrate interspecies differences in decomposition patterns, suggesting that deer may exhibit distinct decomposition trajectories compared to humans. Factors contributing to these differences include variations in body composition, diet, and the diversity of microbial communities, with humans reported to be more prone to scavenging and exhibiting quicker mummification [20]. Further support comes from Mashaly and colleagues [21], who observed that different insect species are attracted to carcasses of different species, reinforcing the likelihood of species-specific decomposition patterns. Nonetheless, Dautartas and colleagues [20] also highlight body weight as a significant factor influencing decomposition rates and patterns. Given the similar body mass of deer and humans, it is plausible that certain aspects of their decomposition processes may be comparable. Additionally, considering that both species are muscle-rich mammals and that bobcats exhibit a clear preference for muscle tissue, the use of deer carcasses as proxies for human remains may be considered a reasonable approach within the context of this study. Despite this justification, the reliance on deer carcasses remains a limitation, underscoring the need for future research utilizing multiple human specimens to validate and expand upon these findings.
All five categories of bobcat scavenging behaviors previously documented (feeding, covering/scent-marking, resting, appearance with no activity, and unknown) [9] were also observed in this study. Of the 78 recorded behaviors, 38 involved feeding, indicating that bobcats chose to feed on the specimens in 48.7% of their appearances. Interestingly, in 34.6% of observations, bobcats were present near the carcasses without engaging in any direct interaction. This result may have been influenced by the behavior observed at Specimen #4, where all 7 bobcat appearances involved complete disregard for the carcass. Similarly, the absence of any bobcat appearances at Specimens #2 and #7 is intriguing, particularly given their well-preserved condition at the time of placement. With a sample size of 7, this study effectively documented a diverse range of bobcat scavenging behaviors.
Bobcats are typically solitary animals, as noted in previous research [22,23], and this tendency toward solitary behavior appears to extend to their scavenging patterns. However, an exception was recorded in this study. On the night of 16 January 2023, two bobcats were observed scavenging simultaneously at Specimen #1 (Figure 2a). There are two plausible explanations for this joint scavenging event. One possibility is that the two individuals were a mother and her offspring; bobcat mothers generally keep their kittens with them through their first winter before they become independent [12,24,25,26]. In this scenario, the smaller bobcat, observed resting while the larger individual fed, may have been the previous year’s offspring. Alternatively, the two bobcats could have been a mating pair, as their breeding season typically begins in winter [24,27]. They may have remained together during this period, seeking sustenance either before or after mating activities. Unfortunately, the camera angle did not allow for identification of the bobcats’ sex, preventing conclusive determination of their relationship. Nevertheless, this rare observation provides valuable insight into the occasionally more social behaviors exhibited by this typically solitary species.
When considering all types of documented behaviors, bobcats typically stayed near the carcasses for 10 min or less per visit, observed in 49% of total appearances (Figure 4). However, it is important to note that this duration includes brief pass-by instances and does not necessarily reflect the length of direct scavenging activities. Indeed, the most common duration for actual scavenging behaviors, categorized as feeding, covering/scent-marking, and resting ranged between 30 min and an hour.
When feeding, bobcats predominantly targeted the torso, which was recorded in 32 out of the 38 total feeding observations. This preference is likely attributed to the high concentration of musculature in the central body area of the carcasses. The bobcats focused primarily on fresh-appearing muscle tissues, rarely consuming limbs distal to the knees or regions above the neck. This selective feeding behavior aligns with previous observations indicating that bobcats favor fresh meat over more decomposed carrion when scavenging [9,12,28]. Such specialized foraging strategies, coupled with their acute sense of smell, confer adaptive advantages, enabling bobcats to avoid highly decomposed remains, where nutritional value and palatability are diminished [22,29,30]. By concentrating on areas with the greatest muscle mass and minimal decay, the bobcats in this study efficiently extracted valuable nutrients and energy. In this context, the lack of feeding on Specimen #4 may be explained by the advanced decomposition and loss of fresh tissue during the 41-day period before the bobcat’s initial appearance.
This study also revealed deviations from previously established patterns concerning bobcat activity hours. A significant portion of the bobcat appearances recorded (22 of 71, 30.1%) occurred outside the peak hours of 04:00–10:00 and 18:00–24:00 proposed by Buie et al. [16]. Similarly, Rippley and colleagues [9] report that only 56.82% of bobcat activities occurred within those timeframes in their study involving a human cadaver in southeastern Texas. These findings suggest that previously established peak hours may not universally apply and that regional variations likely influence bobcat scavenging patterns. The newly identified peak hours of 17:00–07:00 proposed in this study encompassed 64 of the 71 appearances (90.1%) and may more accurately reflect bobcat scavenging behaviors in the southeastern United States. Further research is necessary to test the applicability of these revised peak hours in other regions and across larger sample sizes.
All affected specimens exhibited soft tissue damage patterns consistent with previous research, including long, narrow claw marks in random directions and extensive cut defects with smooth margins [9] (Figure 6). This damage pattern likely resulted from the repetitive actions of licking, pulling, and tearing, as illustrated in Figure 5. However, unlike Rippley et al.’s findings [9], no evidence of bobcat-induced damage was observed on the hard tissues of the specimens. Although some articular cartilage displayed faint tooth marks, no definitive bone modifications were identified once the remains reached the dry/skeletonization stage of decomposition. This discrepancy may reflect anatomical differences and variations in bone strength between deer and human remains. Additional research will be necessary to assess the frequency and extent of hard tissue damage resulting from bobcat scavenging.
Several limitations should be acknowledged in this study. Resource constraints limited the sample size to seven specimens over a 15-month period, potentially restricting the generalizability of the findings. Extending the research duration, particularly into the summer season, and increasing the sample size could provide additional insights, particularly regarding the influence of seasonality on bobcat scavenging behavior. Improved identification tools for distinguishing individual bobcats would facilitate more detailed behavioral analysis. Introducing a control specimen, protected from vertebrate scavengers, would allow for more precise monitoring of decomposition processes without scavenger interference. Additionally, the premature removal of two specimens by coyotes may have introduced bias into the dataset, necessitating careful interpretation of the results. Lastly, as noted earlier, the use of animal models instead of human models may limit the direct applicability of the findings to forensic contexts involving human remains.
Despite these limitations, this study provides valuable insights into the scavenging behavior of bobcats in the middle Tennessee region, a topic that remains understudied. The findings contribute to a broader understanding of local scavenger ecology and its potential implications for forensic investigations. Further research is warranted to build upon these observations and refine our understanding of bobcat scavenging dynamics across varying ecological and forensic settings.
5. Conclusions
The findings of this study suggest that bobcats may influence forensic investigations, underscoring the need for forensic investigators and examiners to be aware of and consider their scavenging patterns during case evaluations. A clear understanding of bobcat scavenging behaviors can help prevent errors in investigating postmortem events. Observations revealed that bobcats preferentially scavenged near the torso, focusing primarily on the musculature while avoiding bony regions. If unsatisfied with feeding on the torso, they typically transitioned to the adjacent sections, with minimal interest shown in the head and neck regions. Bobcats also exhibited a tendency to feed in a localized area for extended periods, potentially resulting in concentrated damage on the remains. Their feeding behavior consisted of shorter bursts, often revisiting the same region until the available tissue was depleted. Notably, scavenging activity ceased after one to two months of carcass placement, suggesting that prolonged or extensive damage observed beyond this timeframe is likely attributable to other factors. Overall, this study highlights the critical role of understanding bobcat scavenging behavior in improving the accuracy of forensic analyses and supporting the integrity of criminal investigations.
Supplementary Materials
The following Supporting Information can be downloaded at: https://www.mdpi.com/article/10.3390/forensicsci5020024/s1, Table S1: Weather condition at MTSU Outdoor Forensic Facility (MOFF) during research period.
Author Contributions
Conceptualization, Y.J. and E.J.W.; methodology, Y.J. and A.G.; software, Y.J.; formal analysis, Y.J. and A.G.; investigation, Y.J. and E.J.W.; data curation, Y.J.; writing—original draft preparation, Y.J. and A.G.; writing—review and editing, E.J.W.; visualization, E.J.W.; supervision, Y.J. and E.J.W.; project administration, Y.J. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Ethical review and approval were waived for this study, as specimen acquisition was conducted under scientific permits (ID: 1850; Type: Terrestrial) issued by the Tennessee Wildlife Resources Agency, which authorized the use of animal remains for research purposes.
Informed Consent Statement
Not applicable.
Data Availability Statement
Inquiries can be directed to the corresponding author.
Acknowledgments
Authors are sincerely grateful to Ddoksoon Lee and anonymous reviewers for reviewing the manuscript and providing valuable comments.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Ubelaker, D.H. Taphonomic applications in forensic anthropology. In Forensic Taphonomy: The Postmortem Fate of Human Remains, 1st ed.; Sorg, M.H., Haglund, W.D., Eds.; CRC Press: Boca Raton, FL, USA, 1997; pp. 77–90. [Google Scholar]
- Indra, L.; Lösch, S.; Errickson, D.; Finaughty, D. Forensic experiments on animal scavenging: A systematic literature review on what we have and what we need. Forensic Sci. Int. 2023, 353, 111862. [Google Scholar] [CrossRef] [PubMed]
- Indra, L.; Errickson, D.; Young, A.; Lösch, S. Uncovering forensic taphonomic agents: Animal scavenging in the European context. Biology 2022, 11, 601. [Google Scholar] [CrossRef] [PubMed]
- Dirkmaat, D.C.; Cabo, L.L. Forensic archaeology and forensic taphonomy: Basic considerations on how to properly process and interpret the outdoor forensic scene. Acad. Forensic Pathol. 2016, 6, 439–454. [Google Scholar] [CrossRef] [PubMed]
- Ubelaker, D.H.; DeGaglia, C.M. The impact of scavenging: Perspective from casework in forensic anthropology. Forensic Sci. Res. 2020, 5, 32–37. [Google Scholar] [CrossRef]
- Kitchener, A.C.; Breitenmoser-Würsten, C.; Eizirik, E.; Gentry, A.; Werdelin, L.; Wilting, A.; Yamaguchi, N.; Abramov, A.V.; Christiansen, P.; Driscoll, C.; et al. A revised taxonomy of the Felidae: The final report of the Cat Classification Task Force of the IUCN Cat Specialist Group. Cat News 2017, 11, 80. Available online: https://repository.si.edu/handle/10088/32616 (accessed on 1 April 2025).
- Larivière, S.; Walton, L. Lynx rufus. Mamm. Species 1997, 563, 1–8. [Google Scholar] [CrossRef]
- Tewes, M.E.; Mock, J.M.; Young, J.H. Bobcat predation on quail, birds, and mesomammals. Natl. Quail Symp. Proc. 2002, 5, 9. [Google Scholar]
- Rippley, A.; Larison, N.C.; Moss, K.E.; Kelly, J.D.; Bytheway, J.A. Scavenging behavior of Lynx rufus on human remains during the winter months of Southeast Texas. J. Forensic Sci. 2012, 57, 699–705. [Google Scholar] [CrossRef]
- Moss, K.E. The Effects of Avian and Terrestrial Scavenger Activity on Human Remains and Decomposition in Southeast Texas During an 18 Month Study. Master’s Thesis, University of Houston, Houston, TX, USA, 2012. [Google Scholar]
- Bumann, G.B.; Stauffer, D.F. Scavenging of ruffed grouse in the Appalachians: Influences and implications. Wildl. Soc. Bull. 2002, 30, 853–860. Available online: https://www.jstor.org/stable/3784239 (accessed on 1 April 2025).
- Platt, S.G.; Salmon, G.T.; Miller, S.M.; Rainwater, T.R. Scavenging by a bobcat, Lynx rufus. Can. Field Nat. 2010, 124, 265–267. [Google Scholar] [CrossRef][Green Version]
- King, K.A.; Lord, W.D.; Ketchum, H.R.; O’Brien, R.C. Facultative scavenging and carrion guild participation by Lynx rufus in the presence of young. Southwest Nat. 2015, 60, 381–385. [Google Scholar] [CrossRef]
- Fain, M.; Jeong, Y. Surveying scavengers and their taphonomic effects in Murfreesboro, Tennessee. In Proceedings of the American Academy of Forensic Sciences, Orlando, FL, USA, 17 February 2023. [Google Scholar]
- Christensen, A.M.; Passalacqua, N.V.; Bartelink, E.J. Forensic Anthropology: Current Methods and Practice, 1st ed.; Cognella: San Diego, CA, USA, 2024; pp. 123–150. [Google Scholar]
- Buie, D.E.; Fendley, T.T.; McNab, H. Fall and winter home ranges of adult bobcats on the Savannah River Plant, South Carolina. In Proceedings of the Bobcat Research Conference: Current Research on Biology and Management of Lynx rufus, Reston, VA, USA, 16–18 October 1979. [Google Scholar]
- Peel, M.; Finlayson, B.; McMahon, T. Updated world map of the Köppen–Geiger climate classification. Hydrol. Earth Syst. Sci. 2007, 11, 1633–1644. [Google Scholar] [CrossRef]
- Steadman, D.W.; Dautartas, A.; Kenyhercz, M.W.; Jantz, L.M.; Murdorff, A.; Vidoli, G.M. Differential scavenging among pig, rabbit, and human subjects. J. Forensic Sci. 2018, 63, 1684–1691. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Opo, R.; Trujillano, A.; Margalida, A. Behavioral coexistence and feeding efficiency drive niche partitioning in European avian scavengers. Behav. Ecol. 2016, 27, 1041–1052. [Google Scholar] [CrossRef]
- Dautartas, A.; Kenyhercz, M.W.; Vidoli, G.M.; Jantz, L.M.; Mundorff, A.; Steadman, D.W. Differential decomposition among pig, rabbit, and human remains. J. Forensic Sci. 2018, 63, 1673–1683. [Google Scholar] [CrossRef]
- Mashaly, A.M.; Al-Ajmi, R.A.; Rady, A.; Al-Musawi, Z.; Farrukh, A. Species richness of scavenger insects on different carcass types. Trop. Biomed. 2019, 36, 630–639. [Google Scholar]
- Anderson, E.M.; Lovallo, M.J. Bobcat and Lynx Wild Mammals of North America: Biology, Management, and Economics, 1st ed.; Johns Hopkins Press: Baltimore, MD, USA, 2003; pp. 758–786. [Google Scholar]
- Bixler, A.; Gittleman, J.L. Variation in home range and use of habitat in the striped skunk (Memphitis mephitis). J. Zool. 2000, 251, 525–533. [Google Scholar] [CrossRef]
- McNitt, D.C.; Alonso, R.; Cherry, M.; Fies, M.L.; Kelly, M.J. Sex-specific effects of reproductive season on bobcat space use, movement, and resource selection in the Appalachian Mountains of Virginia. PLoS ONE 2020, 15, e0225355. [Google Scholar] [CrossRef]
- Winegarner, C.E.; Winegarner, M.S. Reproductive history of a bobcat. J. Mammal. 1982, 63, 680–681. [Google Scholar] [CrossRef]
- Kitchings, J.T.; Story, J.D. Movements and dispersal of bobcats in east Tennessee. J. Wildl. Manag. 1984, 48, 957–961. [Google Scholar] [CrossRef]
- Sandell, M. The mating tactics and spacing patterns of solitary carnivores. In Carnivore Behavior, Ecology, and Evolution, 1st ed.; Gittleman, J.L., Ed.; Cornell University Press: Ithaca, NY, USA, 1989; pp. 164–182. [Google Scholar]
- Witmer, G.W.; DeCalesta, D.S. Resource use by unexploited sympatric bobcats and coyotes in Oregon. Can. J. Zool. 1986, 64, 2333–2338. [Google Scholar] [CrossRef]
- Börger, L.; Dalziel, B.D.; Fryxell, J.M. Are there general mechanisms of animal home range behaviour? A review and prospects for future research: Home range modelling. Ecol. Lett. 2008, 11, 637–650. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.H. The comparison of usage and availability measurements for evaluating resource preference. Ecology 1980, 61, 65–71. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).