3. Results
GNAPHOSIDAE Banks, 1892
Civizelotes akmon n.sp.
Type Material
Holotype: male (NHMC, FC17141), Evros district, Loutros-Dadia, N 25 km after, 140 m alt, [41.0513 N 26.0743 E] grassland near the river with Paliurus spina christi, 14/05/2015–22/06/2015, leg. M. Komnenov.
Etymology
The name of this species refers to the peculiar shape of the large retrolateral median apophysis of the bulbus, resembling the shape of the second little bone of the human ear, called “akmon” (in Greek άκμων), hence, it is a noun in apposition.
Diagnostic remarks and Diagnosis
The generic placement of this new species is problematic, firstly because the female has not yet been found, and secondly, because both emboli of the single male found in this material were broken off. Therefore, it is unclear how long the undamaged embolus is and in which direction it curves, knowledge that may confirm or change the placement of the species. It is here tentatively placed in
Civizelotes Senglet, 2012, based on the generic definition by Senglet [
11] stating that in this genus:
(a) the embolar radix originates from the posterior margin of the tegulum, and at the level of the embolus it becomes a large plate of bowl, (b) there is no intercalary sclerite, (c) the terminal apophysis is reduced, and (d) the embolus is long. Suggesting that this species is most similar to
C. gracilis, the characteristic “
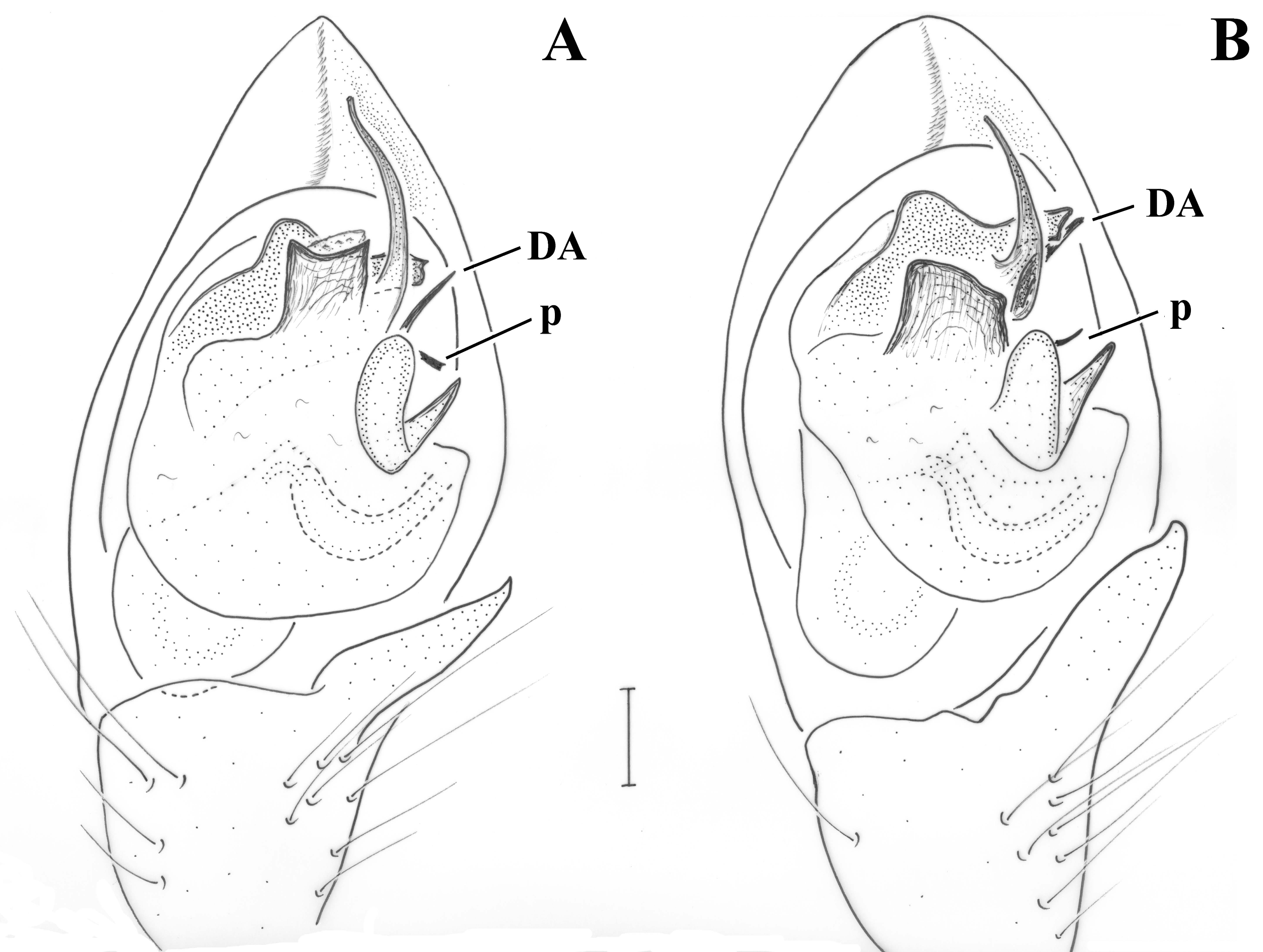
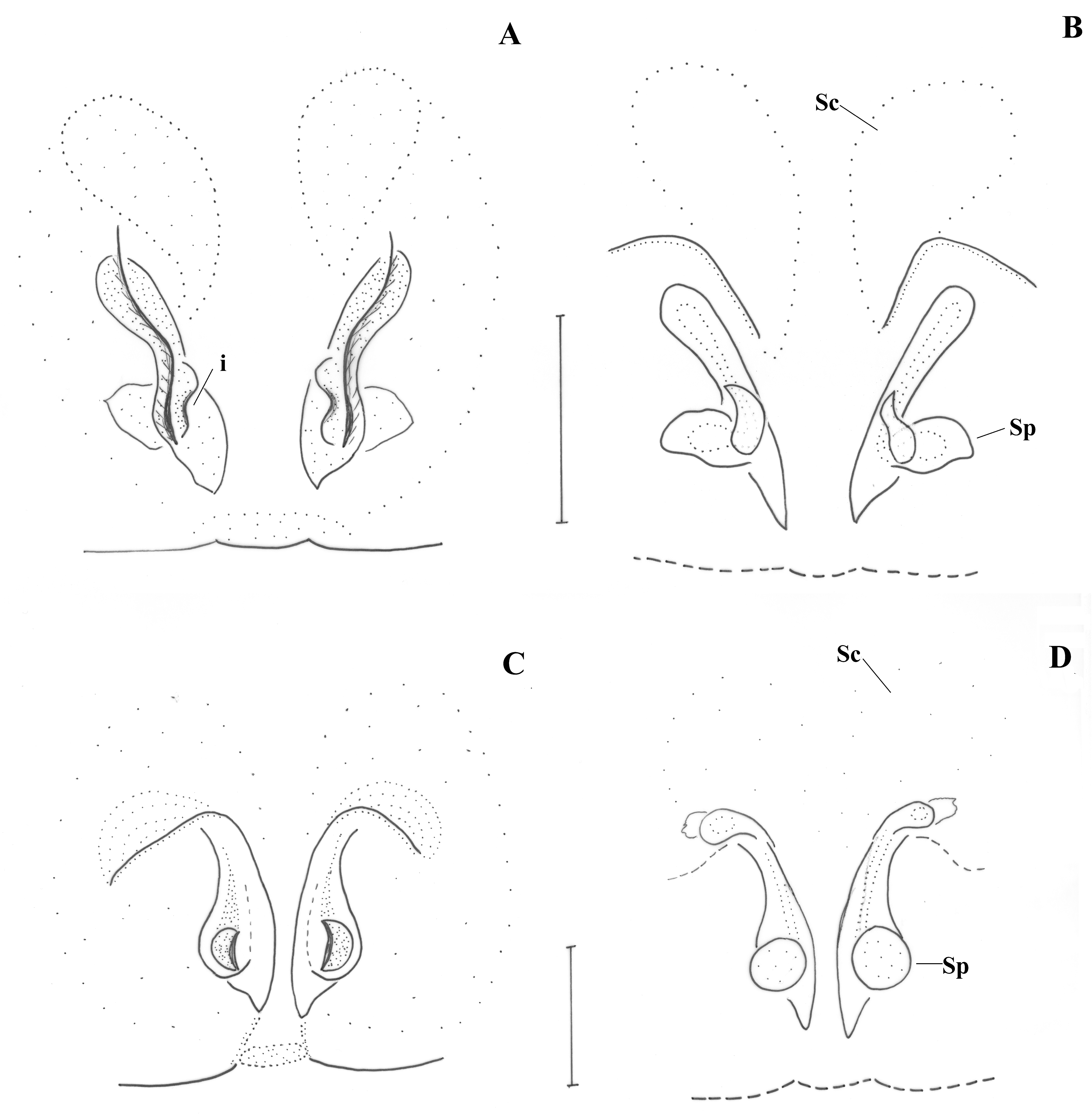
vestigial embolar base” of this species group could be added as diagnostic for the genus identification. Based on the fact that the free part of the embolus starts at ca. the dorso-central to retrolateral end of the tegulum (
Figure 2A), and on the general configuration of the bulb (e.g., tibial apophysis short, median apophysis large), it could also belong to
Heser Tuneva, 2004, but the apical part of the tegulum is definitely not “
grooved to support the embolus” [
11], p. 521 and the embolus would rather rest on the retrolateral part of the median apophysis (and not on the prolateral part—[
11], p. 521).
If the above argument is correct, the closest species to compare
Civizelotes akmon n.sp. to are
C. gracilis (Canestrini, 1868) and
C. pygmaeus (Miller, 1943) (see ([
12] (p. 191–192, figs. 232a/b, 238–239) and [
13] (p. 176–177, figs. 35–36)) resembling both in the form of the median apophysis, in the presence of a prolateral pointed tegular apophysis (
Figure 2A and
Figure 3B) and in the shape of the retrolateral tibial apophysis (
Figure 2C,D) in addition to the strong dorsal setae of the palpal tibia (
Figure 3A). From both, the new species differs by a distinct enlarged embolar radix, by the origin of the free part of the embolus at the dorso-central to retrolateral part of the tegulum, near the median apophysis, as well as by the lack of extra thorn-like apophyses at the base of the embolus (
Figure 2A). The species is further characterized by the peculiar set of stiff setae at the retro-dorsal part of the tibia, giving the impression of a second apophysis when observed in small magnification.
Description
Male (holotype). Small spiders of brown coloration. All Fe, Pa, and Ti darker than the rest of the leg segments, which are light brown. Measurements: CL 1.23, CW 1.05, AL 1.46. Eye sizes and interdistances: AME 0.050, ALE 0.065, PME 0.056, PLE 0.039; PME and AME almost touching, PME-PLE separated by ca. ½ PLE diameter, AME almost touching with ALE. PER slightly procurved, AER slightly recurved. Chelicerae with 2 denticles on RM and 4 teeth on PM. Triangular scutum covering half of the abdominal dorsal surface. Leg formula is IV > I > II > III. Leg spination: Leg I: Fe 2d; leg II: Fe 3d Ti 2v Mt 2v; leg III: Fe 5-6d Ti/Mt spinose; leg IV: Fe 3d Ti/Mt spinose. Mt III and IV with preening combs.
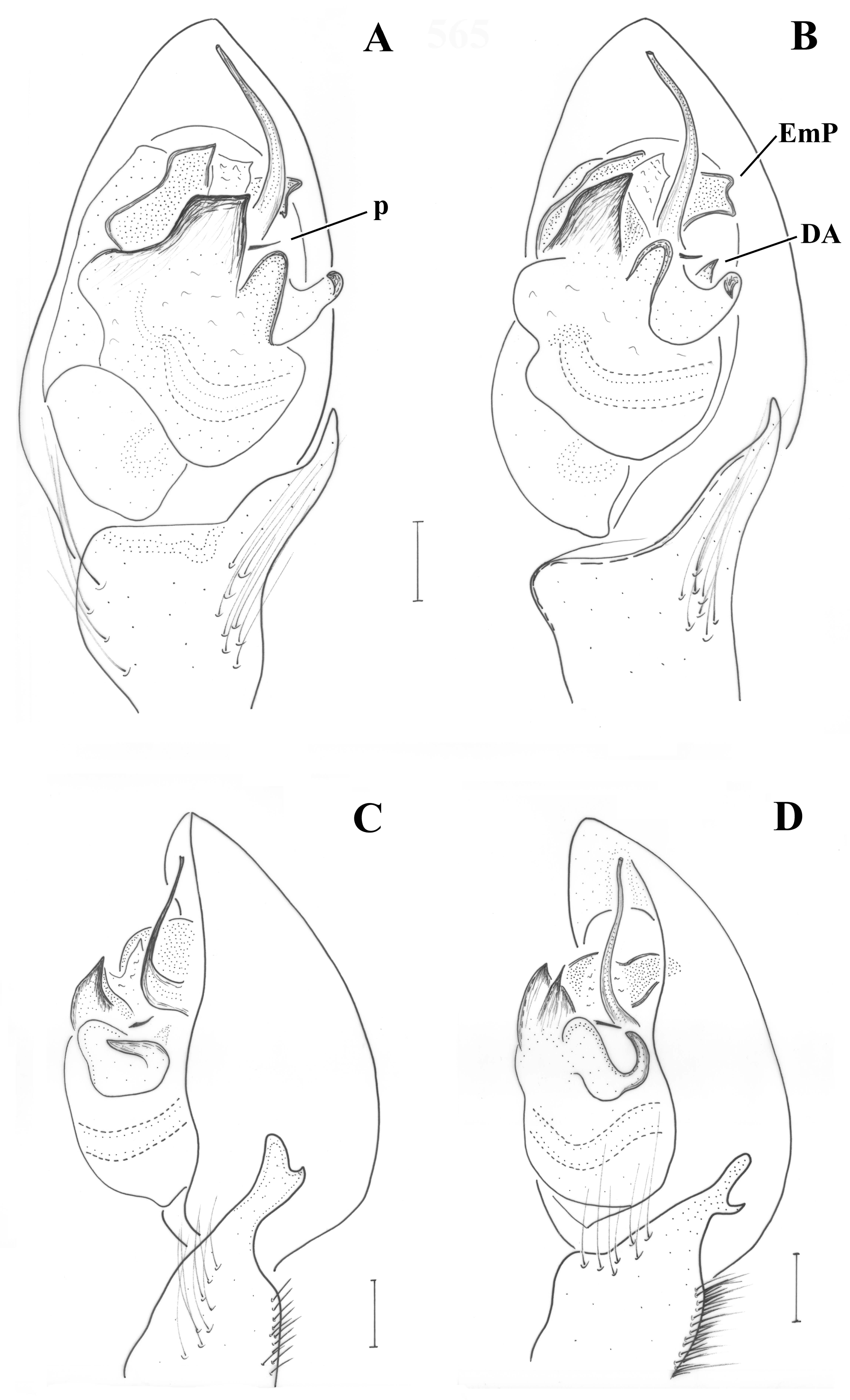
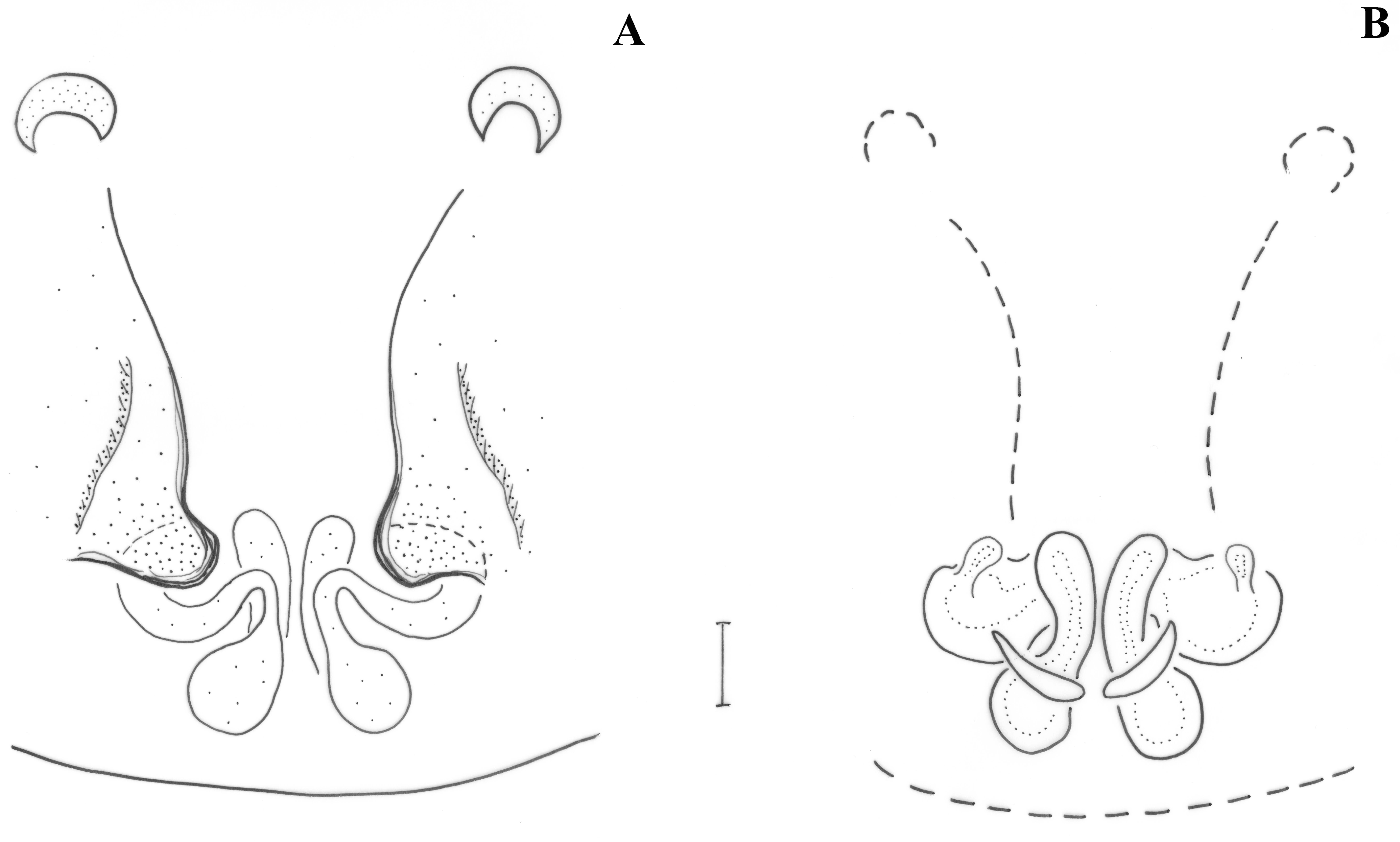
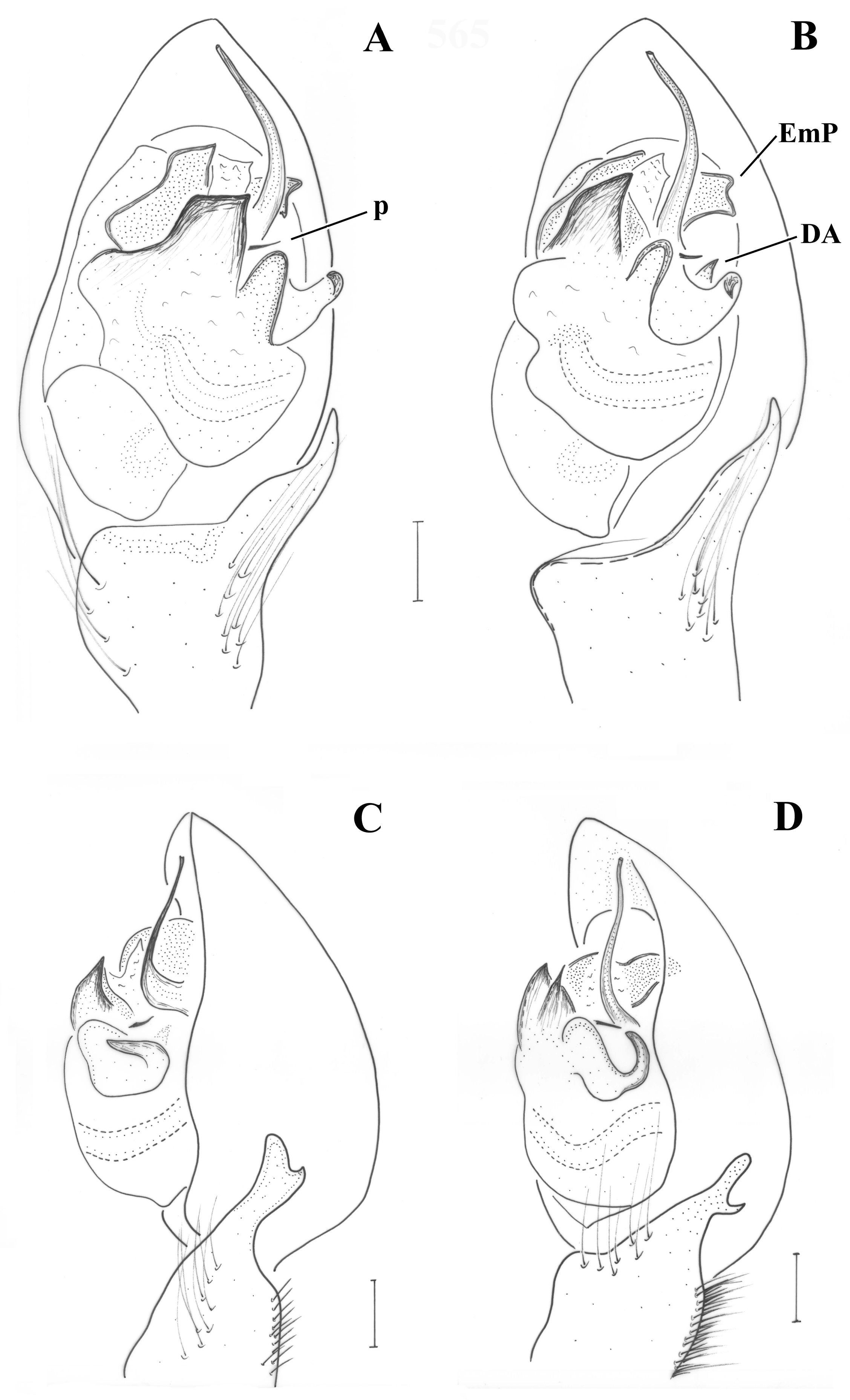
Pedipalp (
Figure 2;
Figure 3). Retrolateral tibial apophysis relatively small, with dorsally pointing tip; dorso-retrolateral stiff setae of tibia may look like a second apophysis, if not observed by large magnification. Median apophysis (MA) large with sclerotized pointed tip directed posteriorly. Terminal apophysis (TA) reduced, barely detectable, not strongly sclerotized. Prolateral edge of tegulum with pointed crest. Embolar radix (EmR) apical, largely extended; embolar base relatively small resting at prolateral side of tegulum, mostly covered by radix; embolus shape unknown, broken off in holotype, but possibly long and filiform.
Civizelotes gracilis (Canestrini, 1868)
Material
Evros: Loutros-Dadia, N 25 km after, 140 m alt (41.0513 N 26.0743 E), grassland near rhe river with Paliurus spina christi [FC: 17141, 14/05–22/06/2015], 2 ♂♂ 1 ♀; same, [FC: 17190, 22/06–21/07/2015], 1 ♂ 7 ♀♀; Loutros-Dadia, N 32 km after, 360 m alt (41.0949 N 26.0930 E), grazed grassland pasture [FC: 17144, 15/05–22/06/2015], 10 ♂♂ 6 ♀♀; Aisymi-Mega Dereio, 5 km before, 580 m alt (41.2143 N 25.9930 E), broad-leaved Quercus forest with some Juniperus at the substrate [FC: 17147, 15/05–23/06/2015], 3 ♂♂ 2 ♀♀; same [FC: 17196, 23/06–22/07/2015], 1 ♂); Aisymi-Mega Dereio, 7 km before, 666 m alt (41.1988 N 25.9728 E), open dry grassland, probably old pasture [FC: 17148, 15/05–23/06/2015], 12 ♂♂ 3 ♀♀; same [FC: 17197, 23/06–22/07/2015], 1 ♂ 3 ♀♀; Mega Dereio-Mikro Dereio, 9 km after, 166 m alt (41.2776 N 26.0671 E), grassland within Quercus/Pinus mixed forest and riverine vegetation, water in 50 m distance [FC: 17153, 16/05–23/06/2015], 3 ♂♂; Mikro Dereio Metaxades, 11.5 km after, 320 m alt (41.3509 N 26.1595 E), open area not cultivated, within agriculture lands and in close vicinity to Quercus forest [FC: 17157, 17/05–23/06/2015], 22 ♂♂ 8 ♀♀; Polia-Ladi, 1.5 km after (41.4491 N 26.2382 E), dry grassland in an open area, near a Quercus forest [FC: 17161, 17/05–23/06/2015], 7 ♂♂ 4 ♀♀; same [FC: 17257, 23/06–22/07/2015], 1 ♂—all legs M. Komnenov & K. Zografou; Evros Delta, (40.8343 N 26.0017 E), marsh meadows in Anthia beach [FC: 17185, 17/06–17/07/2015], 1 ♀; Evros Delta, before Drana (40.8416 N 26.0487 E), rushy marsh field [FC: 12200, 13/05–17/06/2015], 6 ♂♂—all leg. M. Komnenov; Florina, Prespes, 4,5 km SW of Ag. Germanos, 877 m alt (40.81439 N 21.12520 E), agricultural area, dry board by alfalfa field [FC: 17002, 24/06–29/06/2014], 2 ♂♂; [FC: 17002, 24/06-29/06/2014], 1 ♀; [FC: 17002, 24/06–29/06/2014], 1 ♀; Grevena, 1,6 km S of Perivoli, 1400 m alt (39.96530 N 21.11720 E), rocky slope with Juniperus and open coniferous shrubland at the roadside [FC: 17088, 24/06/2014], 1 ♀—all leg. C. Kropf et al.; Magnisia, Portaria to Chania, 1.5 km after (39.3767 N 23.0296 E), maquis dominated by Phlomis sp. [FC: 9281, 24/04–28/06/2014], 1 ♂, leg. D. Kaltsas; Ipeiros, Voio Mt, Kyparissi, 950 m alt (40.1431 N 21.2254 E), mature oak trees and grassland by a rural street behind a football court [FC: 11573, 24/6–16/7/2011], 1 ♀, leg. S. Zakkak.
Comparative Material Examined
C. pygmaeus: Austria, Südtirol, Castelfeder 400 m (2 ♂♂, May to June 1988—Cth, AR22790); idem (2 ♂♂, NMW 14.999)—sub C. gracilis (misidentification).
C. gracilis: Austria, Burgenland, Pandorfer Platte (3 ♂♂ 1 ♀—Cth, AR 22792, leg. Thaler, Meyer, Steinberger, 1988); Austria, Ritzing, Rabenkropf (1 ♀, June 1984- Cth, AR22791).
Comments
The taxonomic status of this species is somehow problematic due to the fine differences of the male genitalia with
C. pygmaeus (Miller, 1943). The identification of the Greek material cited above was based on the close examination of the literature ([
12] (p. 191–192, figs. 232a/b, 238–239) and [
13] (p. 176–177, figs. 35–36)), as well as of comparative material from the K. Thaler collection deposited in the natural history museum of Bern and of Wien (see comparative material). In the course of this process, it was concluded that the species assigned to
C. gracilis by [
13] is actually
C. pygmaeus. Interestingly, in their discussion, the authors also state that the details of the male genitalia found in south Tirol (as also illustrated in their Figure fig. 35) correspond to
C. pygmaeus, but in the absence of females in their material and also because of geographical reasons they took the erroneous decision of assigning it to
C. gracilis.
Our specimens fully correspond to the detailed drawings of Grimm [
12] (p. 191–192, figs. 232a/b, 238–239) with only one exception (correctly noted by Thaler & Noflatscher [
13]): the embolar base (or radix
sensu Senglet [
11]) of males has two thorn-like apophyses, the one directed to the retrolateral side really almost indistinct and visible only from certain angles.
Distribution
Europe, Russia.
Drassodes omalosis Roewer, 1928 sp. reval.
Drassodes omalosis Roewer, 1928: 100, figs. 4–6
D. omalosis: Chatzaki et al., 2002: 615, figs. 32, 33, 36
Not Drassodes unicolor (O. Pickard-Cambridge, 1872): Levy, 2004, p. 8, figs. 20–26
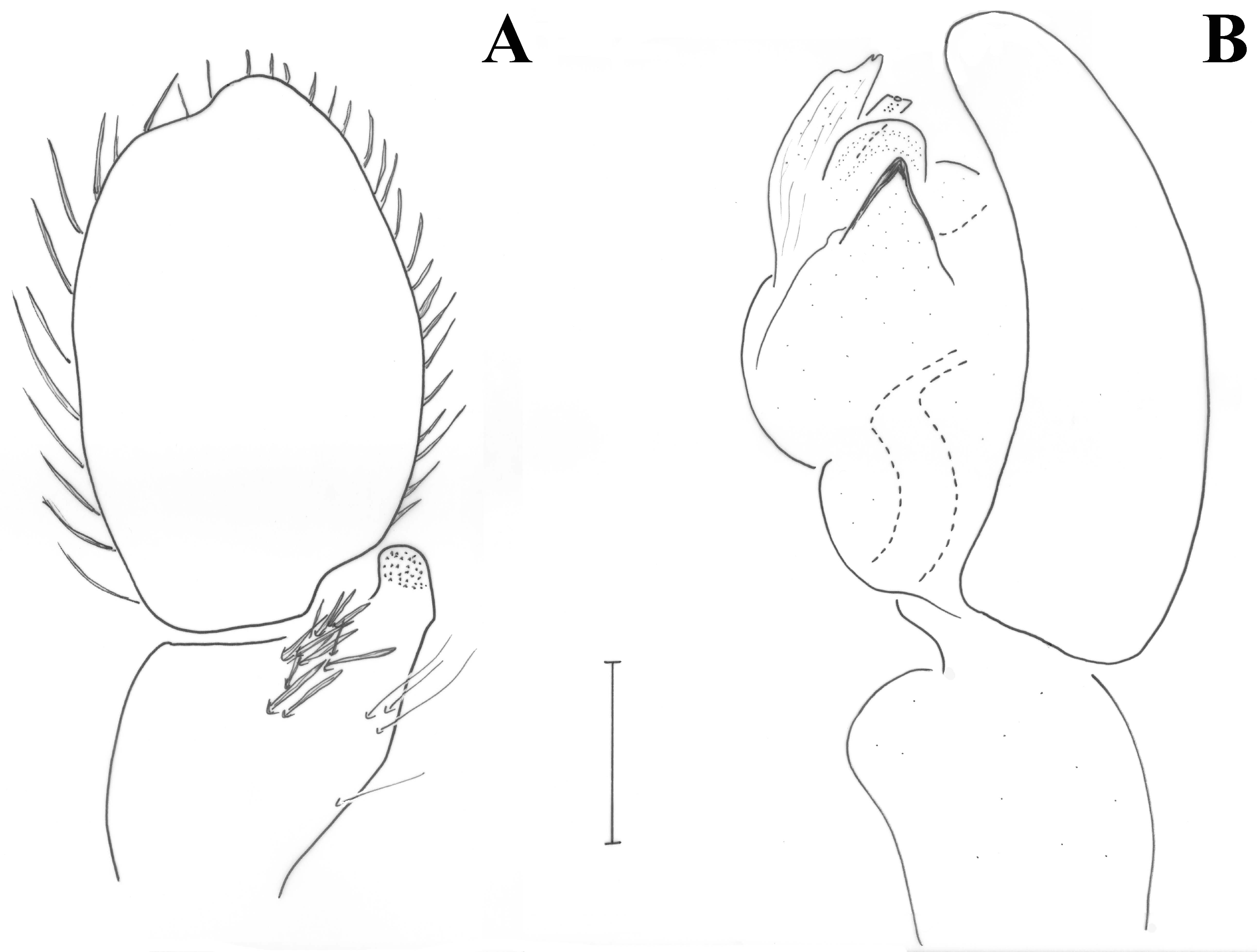
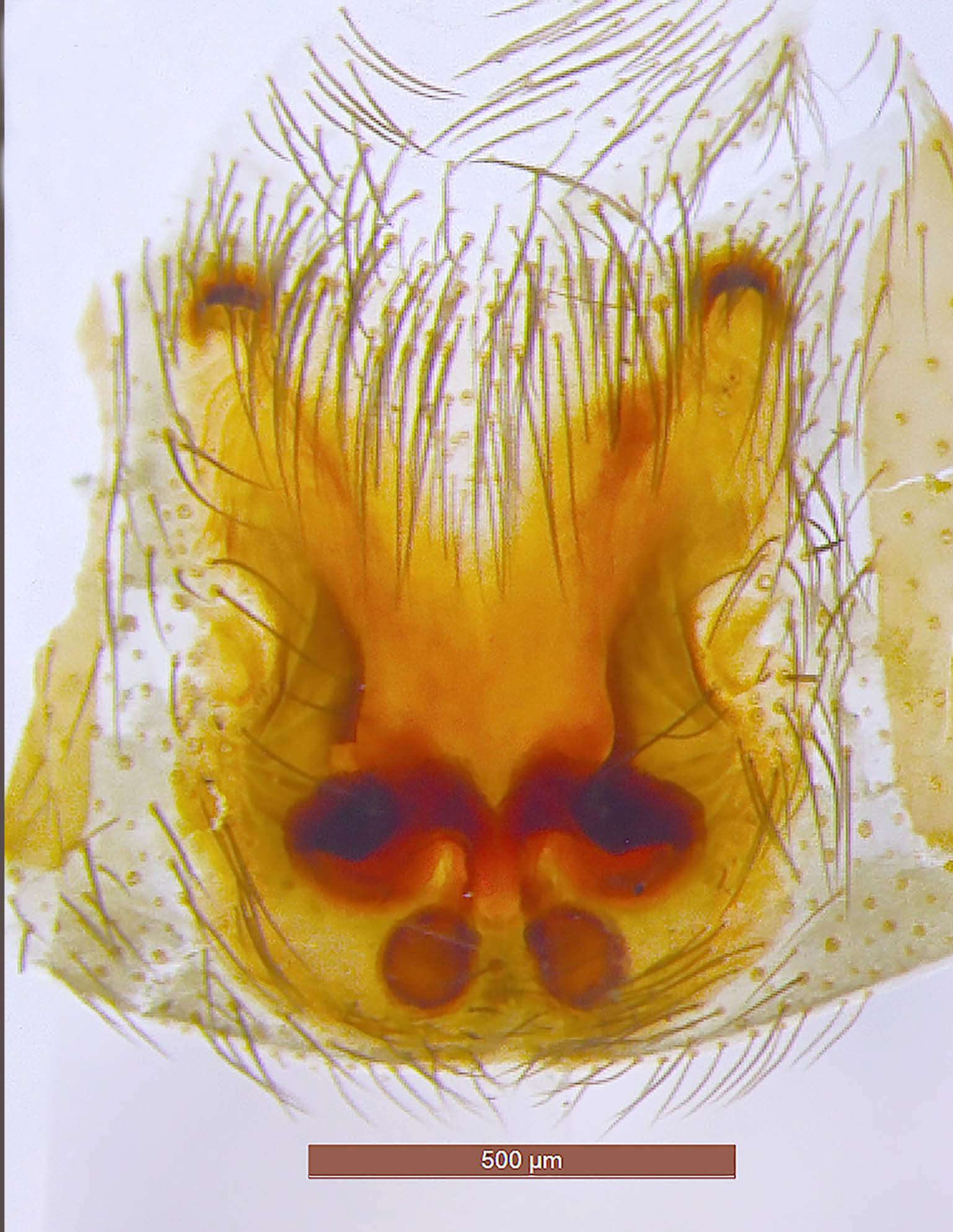
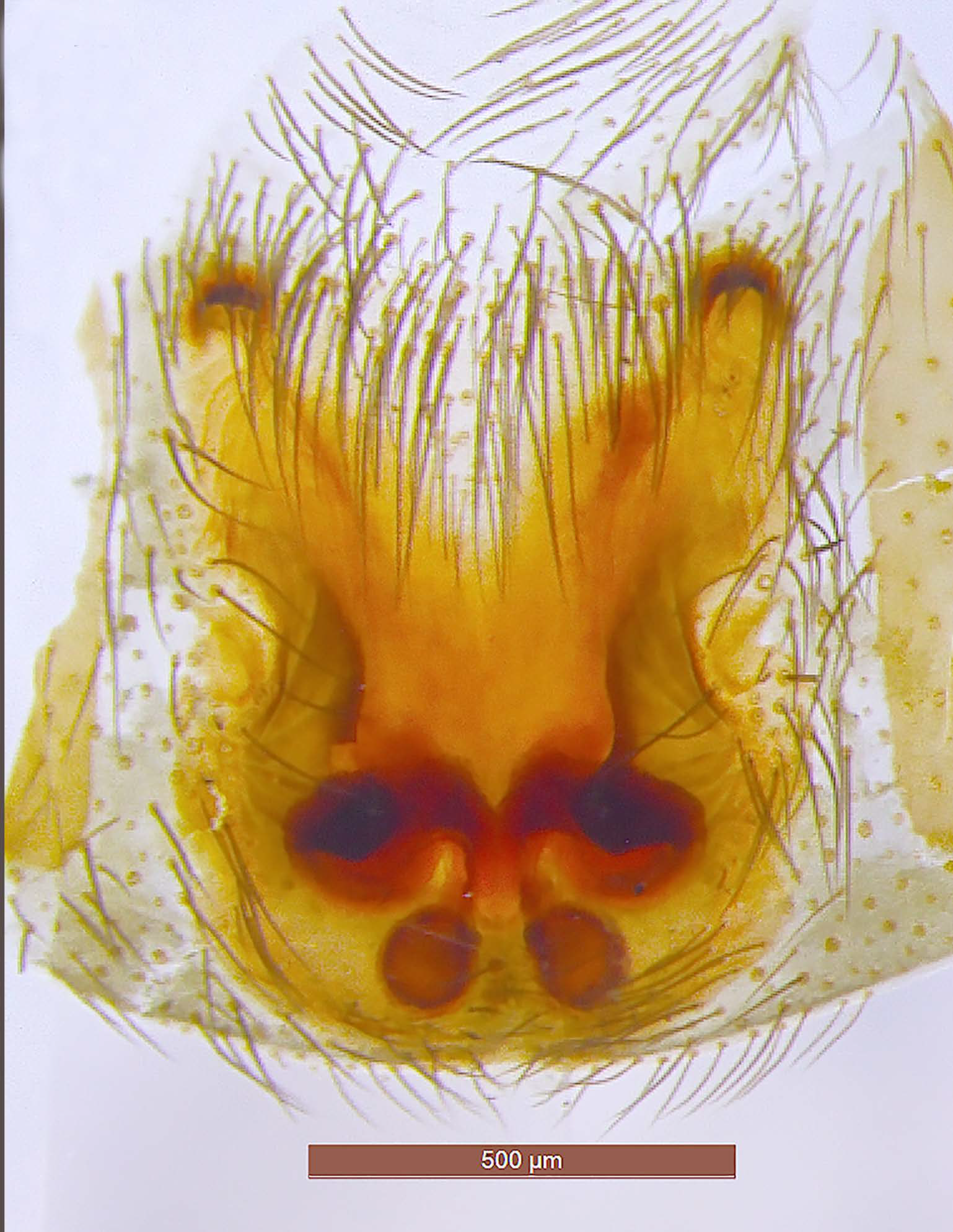
Figure 4.
Drassodes omalosis, female (FC: 8792). (A) Epigyne, (B) vulva.
Figure 4.
Drassodes omalosis, female (FC: 8792). (A) Epigyne, (B) vulva.
Figure 5.
Drassodes omalosis, epigyne (FC: 8792).
Figure 5.
Drassodes omalosis, epigyne (FC: 8792).
Material
Ioannina, Katara (39.7911 N 21.2263 E), beech forest (Fagus sp.) [FC: 8792, 16/06–08/08/2014], legs D. Kaltsas & O. Mettouris, 1 ♀.
Comparative Material Examined
Drassodes unicolor: ISRAEL: Sansan Reserve 157/123.3.1, 1 ♀, 31.03.2001, leg. Y.Mendelik; Mt. Meron, 1 ♀, 8.05.1975, leg. M. Warburg; Jerusalem, 1 ♀, 22.12.1949, leg. A. Shulov [HUJ 11982]; Kfar-Ezion, 1 ♀, 6.01.1944, leg. A. Shulov [HUJ 11990]; Mt Meron 05.07.1961, 1 ♂, leg. G. Levy [HUJ 14701]; Sansan Reserve 157/123.3.9, 11.11.2001, 1 ♂, leg. Y.Mendelik; Near Battir, 16.11.1940, 1 ♂, leg. A. Shulov [HUJ 11988].
Comments
Drassodes omalosis was described by Roewer [
14] on the basis of a couple of specimens from Crete. The type material deposited in the Senckenberg Museum of Frankfurt (SMF) was examined and illustrated by Chatzaki et al. ([
5], p. 615, figs. 32, 33, 36). Based on the new information provided in that paper, Levy ([
15]) proposed the synonymy of
D. omalosis with
D. unicolor (O. Pickard-Cambridge, 1872), but with no further justification. Although the comparison of illustrations of the two species raised doubts about the validity of this synonymy at the time, the absence of fresh material from Crete or from any other Greek territory, as well as the fact that
D. unicolor was not yet examined by the first author, did not allow any relevant detailed comparison up to the present. The record of a female in the material from N. Greece here presented raised the topic again, since it showed close resemblance to the illustrations of the type of
D. omalosis provided by Chatzaki et al. ([
5], fig. 36), but not those of
D. unicolor by Levy ([
15], figs. 25–26). The first author had the chance to examine
D. unicolor from Israel in 2019 and confirmed the accuracy of the drawings provided by Levy, and hence the differences of the two species. Here, new drawings and pictures of the female from Ioannina, West Greece, are presented. The main difference of the two species, as far as females are concerned, is in the shape of the median plate, in
D. omalosis forming a trapezium with a transverse and wider anterior frame, rather than a pot-like shape as in
D. unicolor. Furthermore, the embolus in the male of
D. omalosis seems more robust than in
D. unicolor and the membrane following the median apophysis lies near its outer part (opposite in
D. unicolor).
Therefore, the distribution of D. omalosis is expanded from Crete to the rest of Greece and that of D. unicolor is limited again in Israel and presumably the other countries reported by Levy, i.e., Libya, Egypt, and Lebanon (due to biogeographic affinities to Israel). Interestingly though, it should be noted that D. omalosis has not been found in Crete since its original description, although the island (and especially the topotypic area of the species) has been searched many times and by several collectors.
Distribution
Greece, Libya?, Egypt?, Lebanon?, NOT Israel.
Drassyllus centrohellenicus n.sp.
Type Material
Holotype: male (NHMC, FC 9277). Magnisia, Kouri [39.1918 N 22.7436 E] lowlands oak forest (Quercus pubescens, Q. ithaburensis, Q. pendunculiflora), 23/04–27/06/2014, leg. D. Kaltsas. Paratype: same—1 ♀.
Material
Same as holotype, 5 ♀♀; Korinthia, Geraneia Mt, Perachora-Iraio, 2 km after (W) (38.0335 N 22.9248 E), maquis with Olea europaea trees and thorny bushes [FC: 9268, 04/04–09/06/2014], 1 ♂ 1 ♀, leg. D. Kornilios.
Etymology
The specific name of this species refers to the geographic area in which it has been found so far.
Diagnosis
Drassyllus centrohellenicus n.sp. belongs to the same morphological group as
D. dadia Komnenov & Chatzaki, 2016,
D. dadianus n.sp.,
D. pusillus (C. L. Koch, 1833), and
D. villicus (Thorell, 1875). Males differ from the latter two species by the same elements as described in the diagnosis of
D. dadianus n.sp. From
D. dadia, and
D. dadianus n.sp. It differs by the presence of a smaller triangular scutum (rectangular and broader in the other two species), different shape of the dorsal apophysis (small and conical in
D. centrohellenicus n.sp. (
Figure 6B), long and filiform in the other two), and broader end of the embolar projection, as well as other smaller differences in the palpal configuration. Females are mostly similar to
D. dadianus n.sp. in that they both have continuous anterior margin with no central protrusion (as is the case in
D. dadia), but differ from it by the shape of the epigynal plate, being shorter (epigynal plate lenth/width = 1.15 vs. 1.5 in
D. dadianus n.sp.) (
Figure 7).
Description
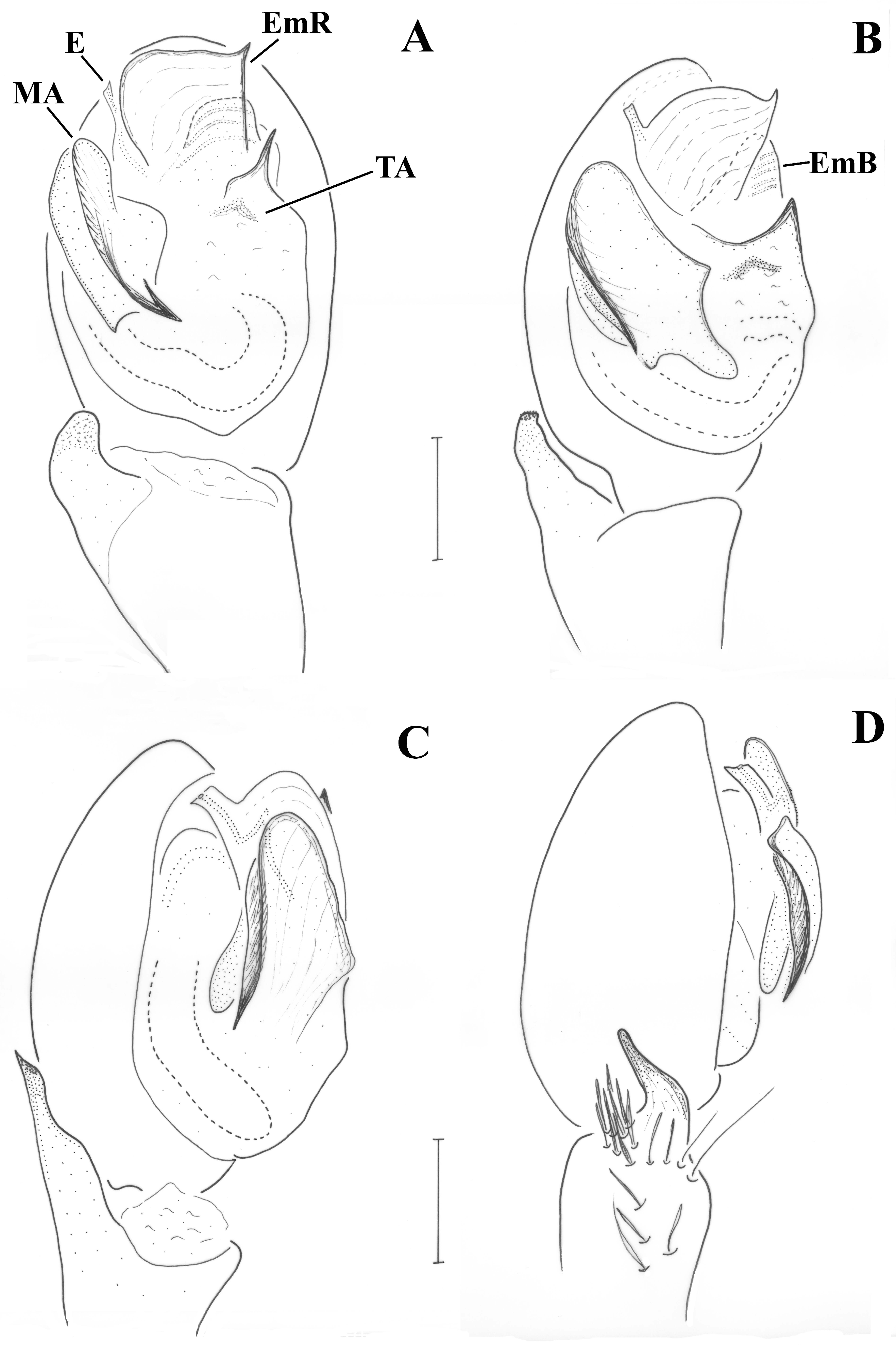
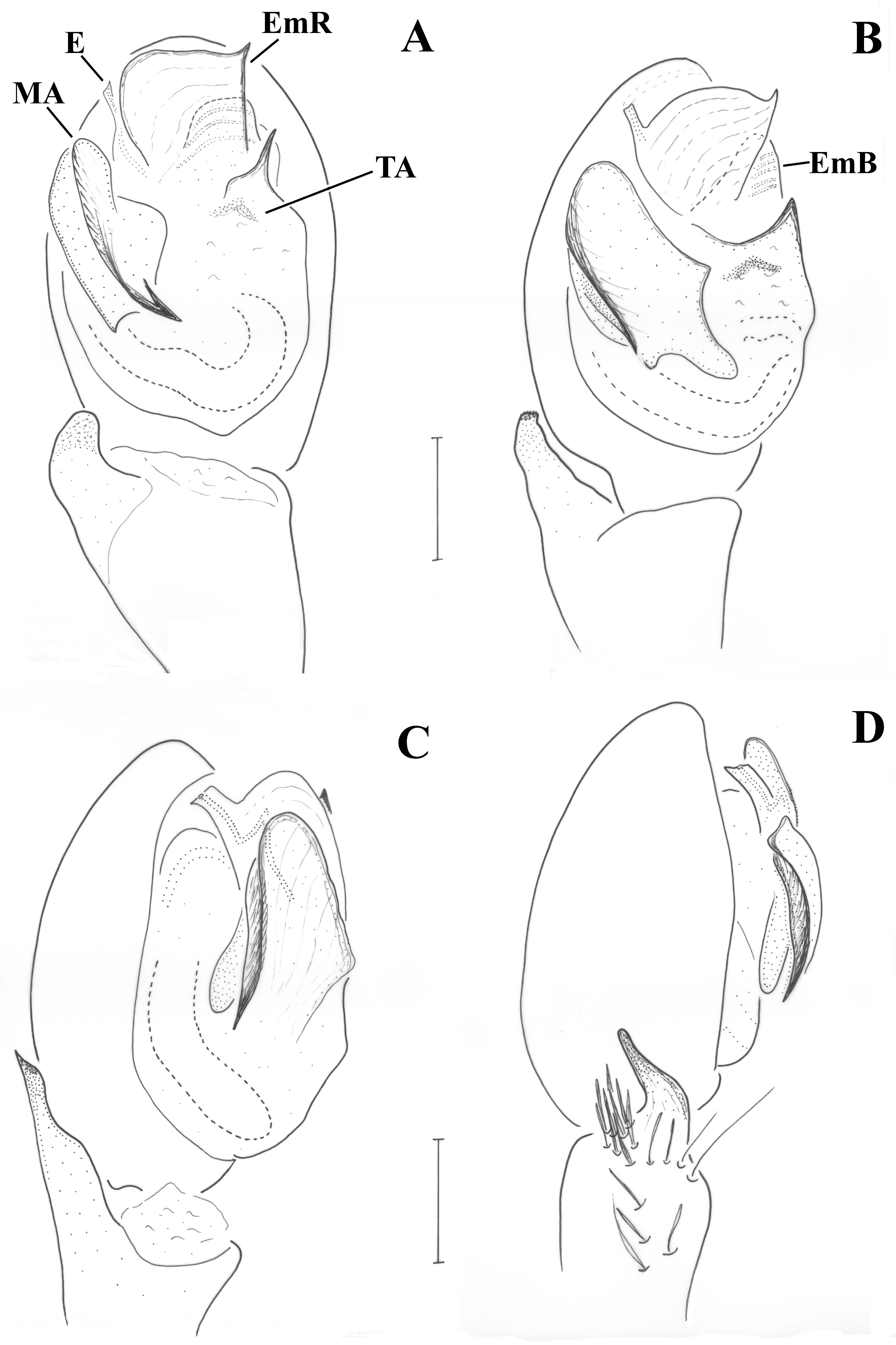
Male (holotype). CL 1.72, CW 1.43, AL 1.99. Eye sizes and interdistances: AME 0.061, ALE 0.100, PME 0.080, PLE 0.070, PME oblique, almost touching, PME-PLE separated by ca. 1/2 the diameter of PLE, AME separated by ca. their diameter, AME-ALE almost touching. Chelicerae with 0–1 tooth at PM and 2 denticles at RM. Legs orange, except femur and coxa, which are blackish brown. Leg spination: Leg I: Fe 2d, Ti 1v, Mt 2v; leg II: Fe 2d, Ti 2v, Mt 2v; leg III: Fe 5d, Ti/Mt spinose; leg IV: Fe ??, Ti/Mt spinose. All tibiae and patellae with one long seta at apical end; metatarsi I and II with additional short spinny setae in two rows. Abdomen grey, dorsally with triangular scutum covering less than 1/2 of the abdomen.
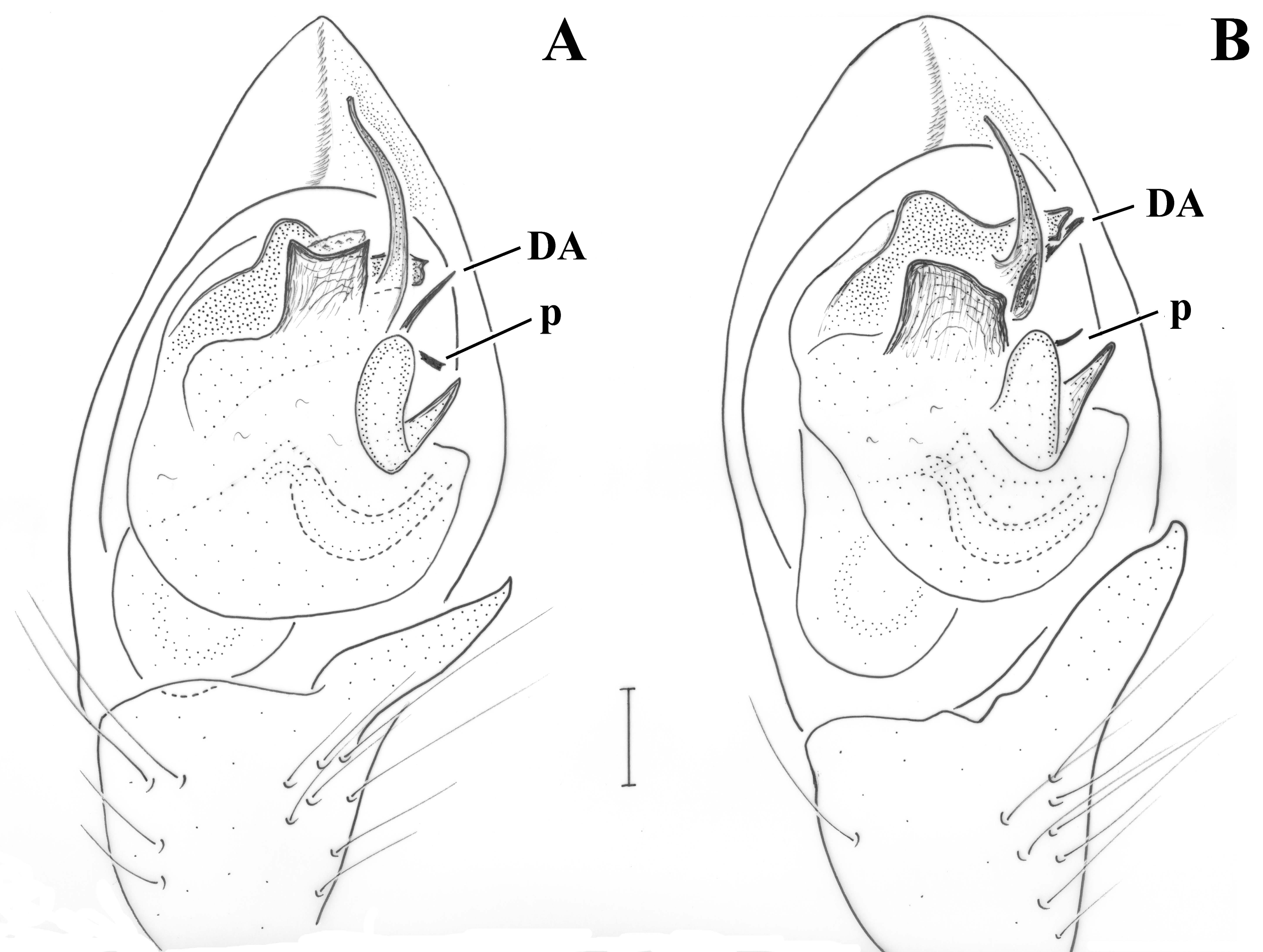
Pedipalp as in
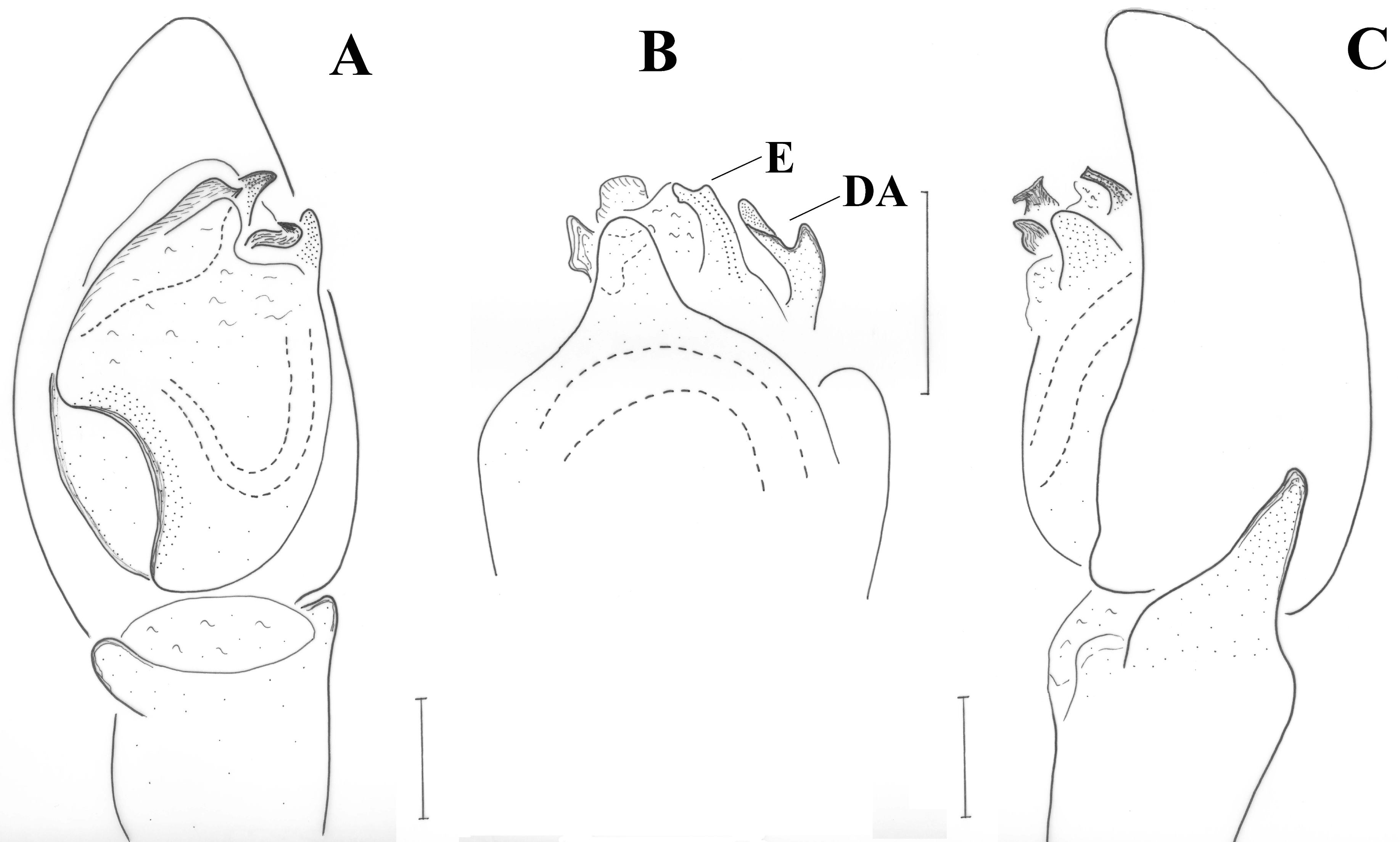
Figure 6A–C. Tibia with stiff setae dorsolaterally; retrolateral tibial apophysis with two pointed tips like the human thumb and index finger; terminal apophysis rectangular, longer than wide, with distal surface almost straight, transverse and with slightly pointed retrolateral edge; base of terminal apophysis with retrolaterally directed transverse projection (p), needle-shaped; embolar projection (EmP) transverse, sclerotized, with blunt end and forming two crests; a membrane is filling the space between the embolus and the embolar projection; embolus long, filiform, almost reaching the end of the cymbium; dorsal apophysis (DA) small, conical, hardly visible behind the median apophysis and only when the palp is turned slightly towards the retrolateral side.
Female(paratype). CL 1.71, CW 1.37, AL 2.71. Eye sizes and interdistances: AME 0.063, ALE 0.104, PME 0.092, PLE 0.068, PME oblique, almost touching, PME-PLE separated by ca. ½ of PME diameter, AME separated by ca. ½ their diameter, AME-ALE almost touching. Chelicerae with 4 teeth at PM and 2 denticles at RM. Legs orange, except femur and coxa, which are blackish brown (no female with a full set of legs were available).
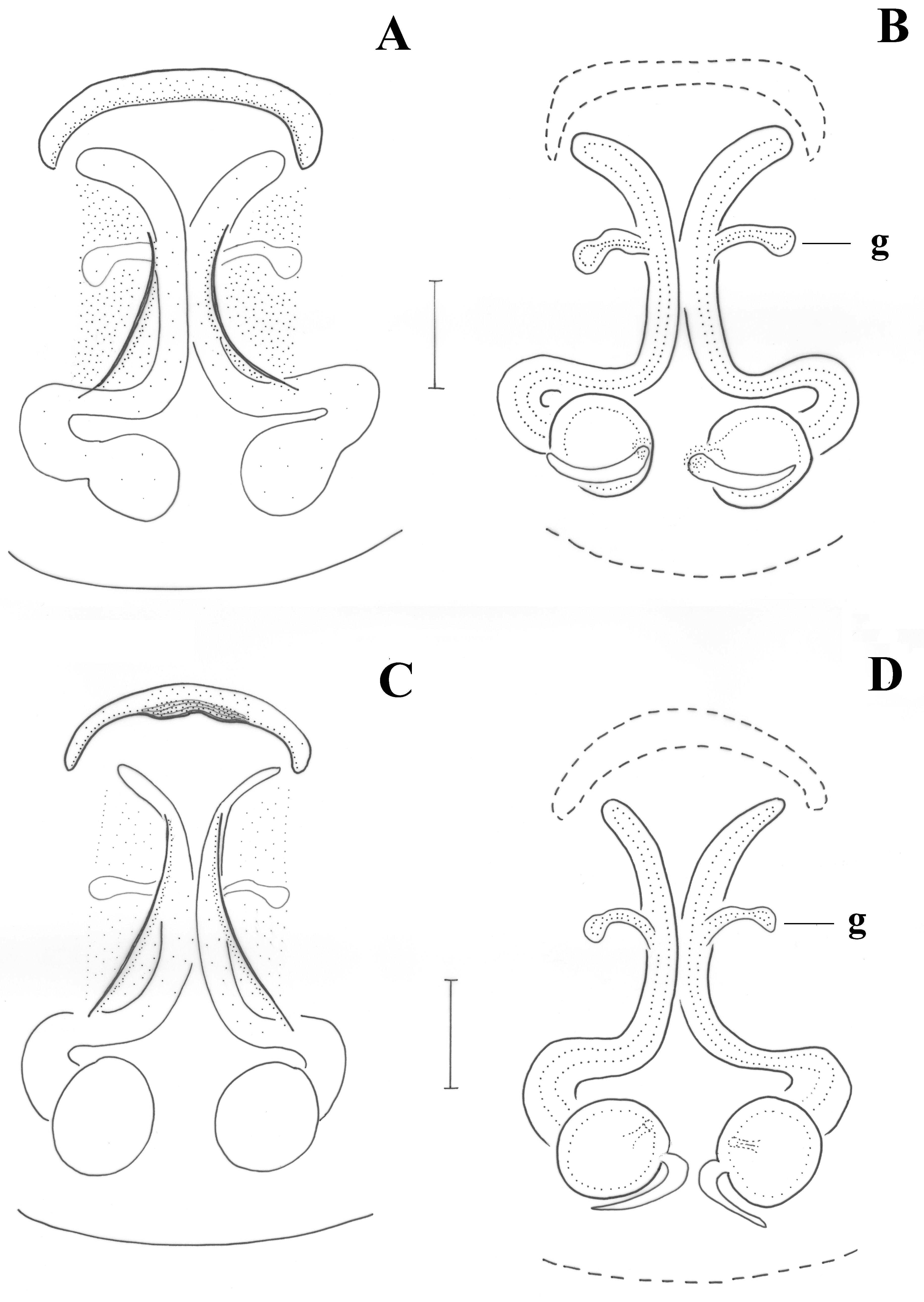
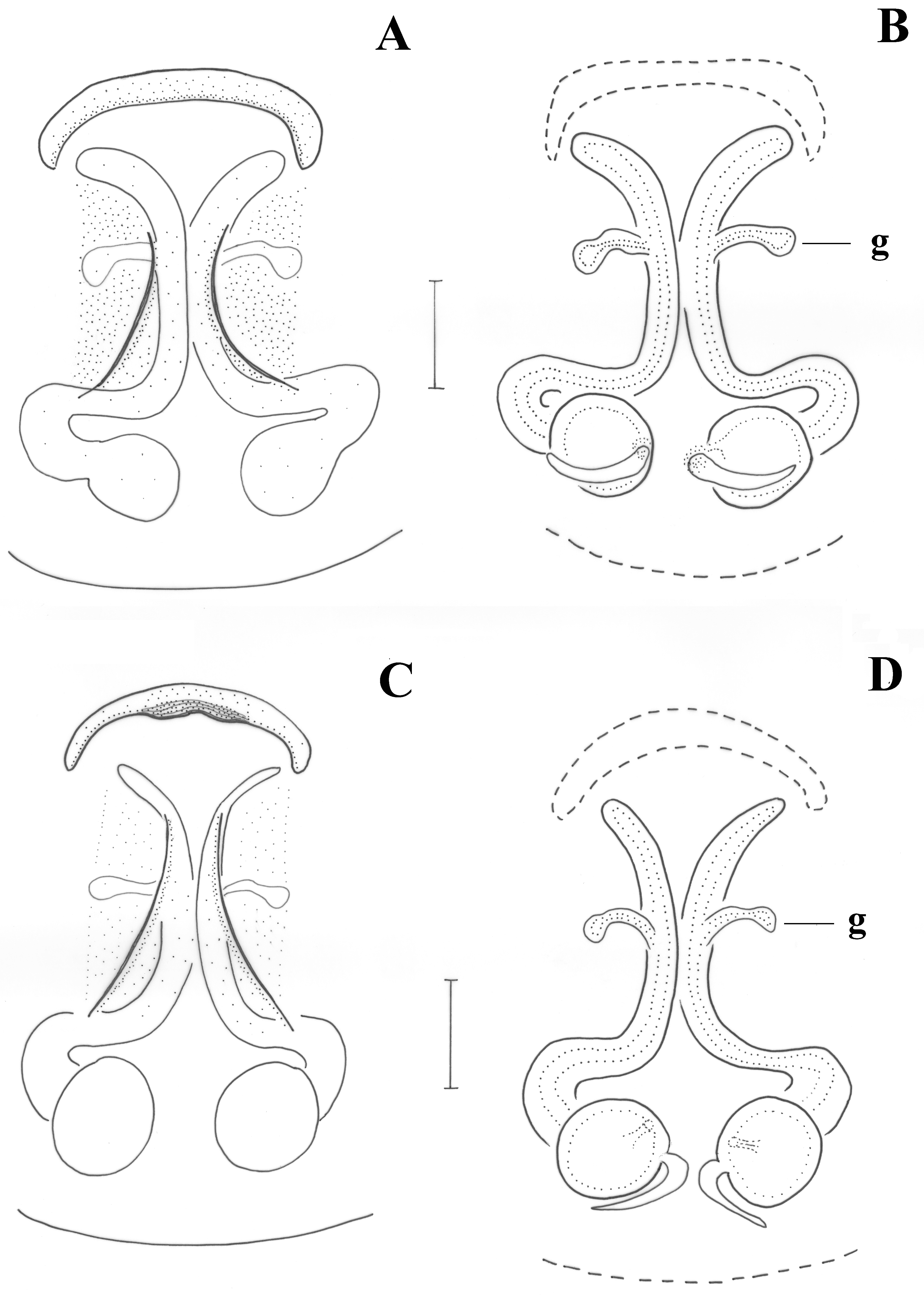
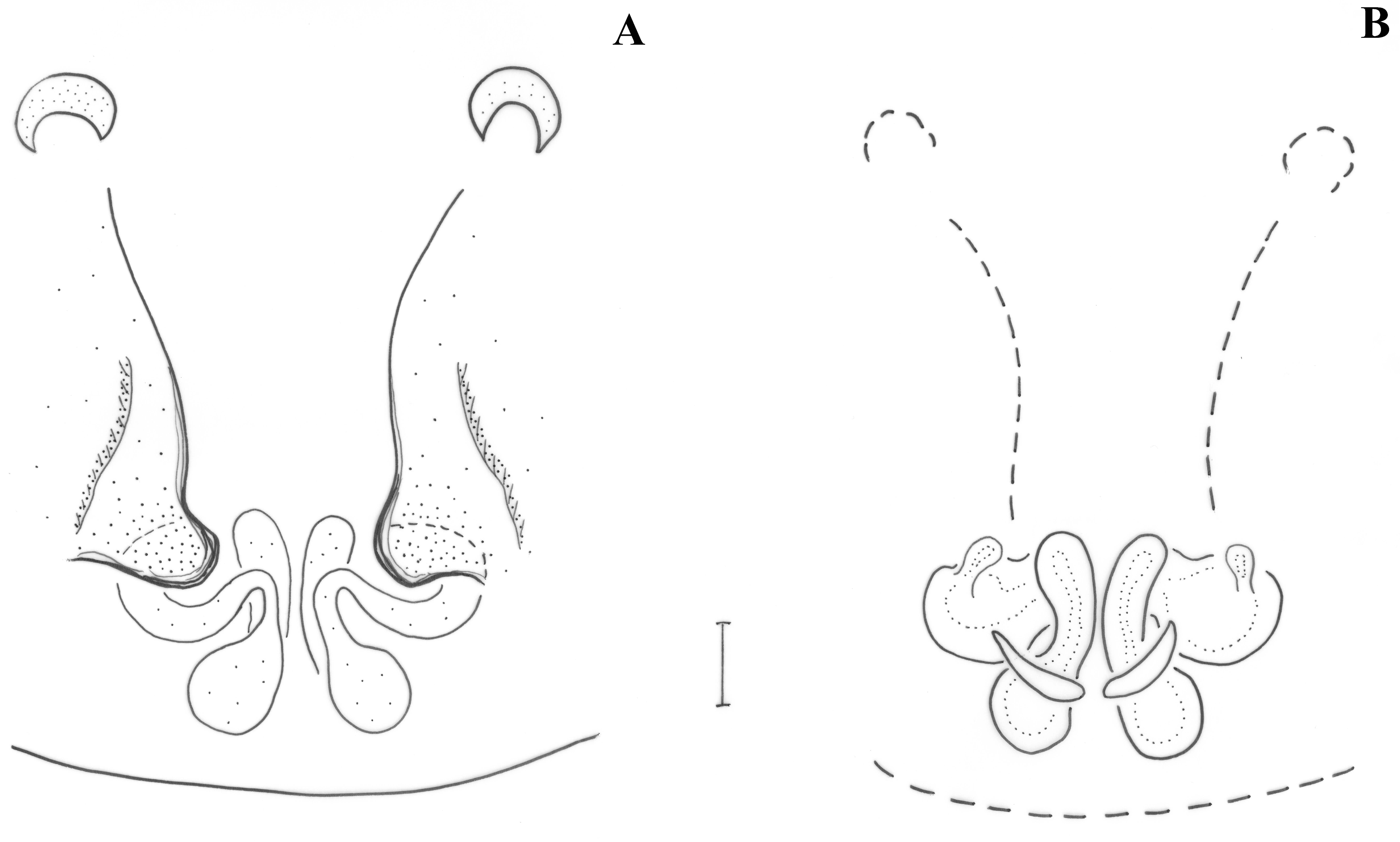
Epigyne as in
Figure 7A. Epigynal plate longer than wide; anterior margin continuous, almost straight. Vulva as in
Figure 7B. Copulatory ducts semi-circular, parallel in the middle, widening posteriorly to reach the spermathecae; lateral glands (g) above the middle of copulatory ducts; distance between spermathecae ca. half their diameter.
Drassyllus covid n.sp.
Type Material
Holotype: female (NHMC, FC17194), Evros district, Treis Vryses-Dadia, 7.5 km after, 500 m alt [41.0950 N 26.0759 E], broad-leaved forest with dense litter and high canopy cover, 22/06–21/07/2015, legs K. Zografou and M. Komnenov.
Etymology
The specific name is attributed to the difficult situation experienced globally during the period of the description due to the COVID-19 virus pandemia, hence, it is a noun in apposition.
Diagnosis
Drassyllus covid n.sp. is very similar to
D. villicus ([
12], p. 276, figs. 329–330) but differs from it by the absence of anterior epigynal margin and the size of additional spermathecal sacs and lateral glands, in
D. covid n.sp. being narrower and shorter, respectively (
Figure 9).
Description
Female (holotype). CL 1.20, CW 1.73, AL 2.90. Eye sizes and interdistances: AME 0.06, ALE 0.10, PME 0.085, PLE 0.08, PME oblique, separated by ca. 1/2 their diameter, PME-PLE separated by their diameter, distance between AME more than their diameter, AME-ALE almost touching. Chelicerae with 4 teeth on PM and 2 denticles on RM. No legs available.
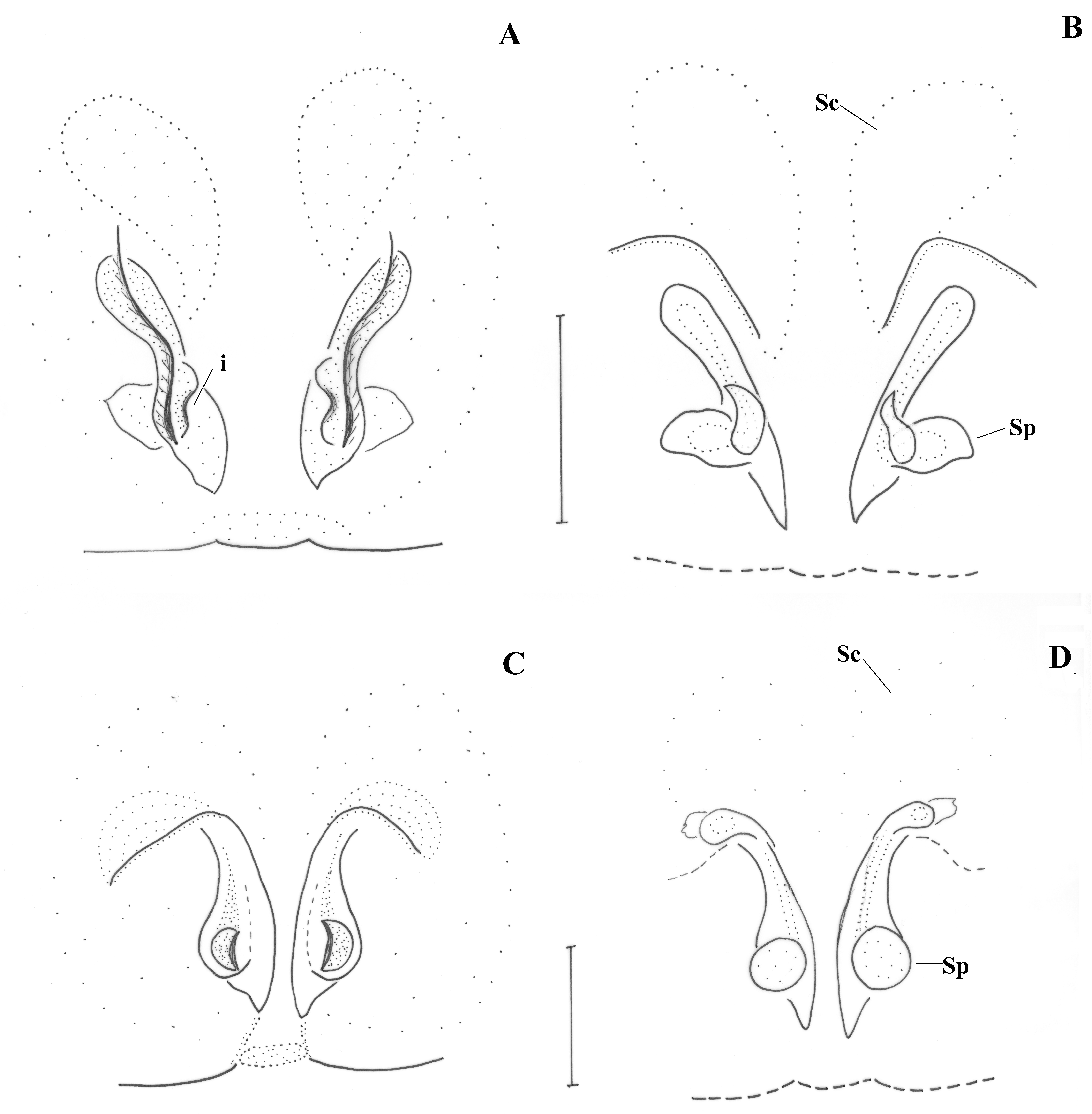
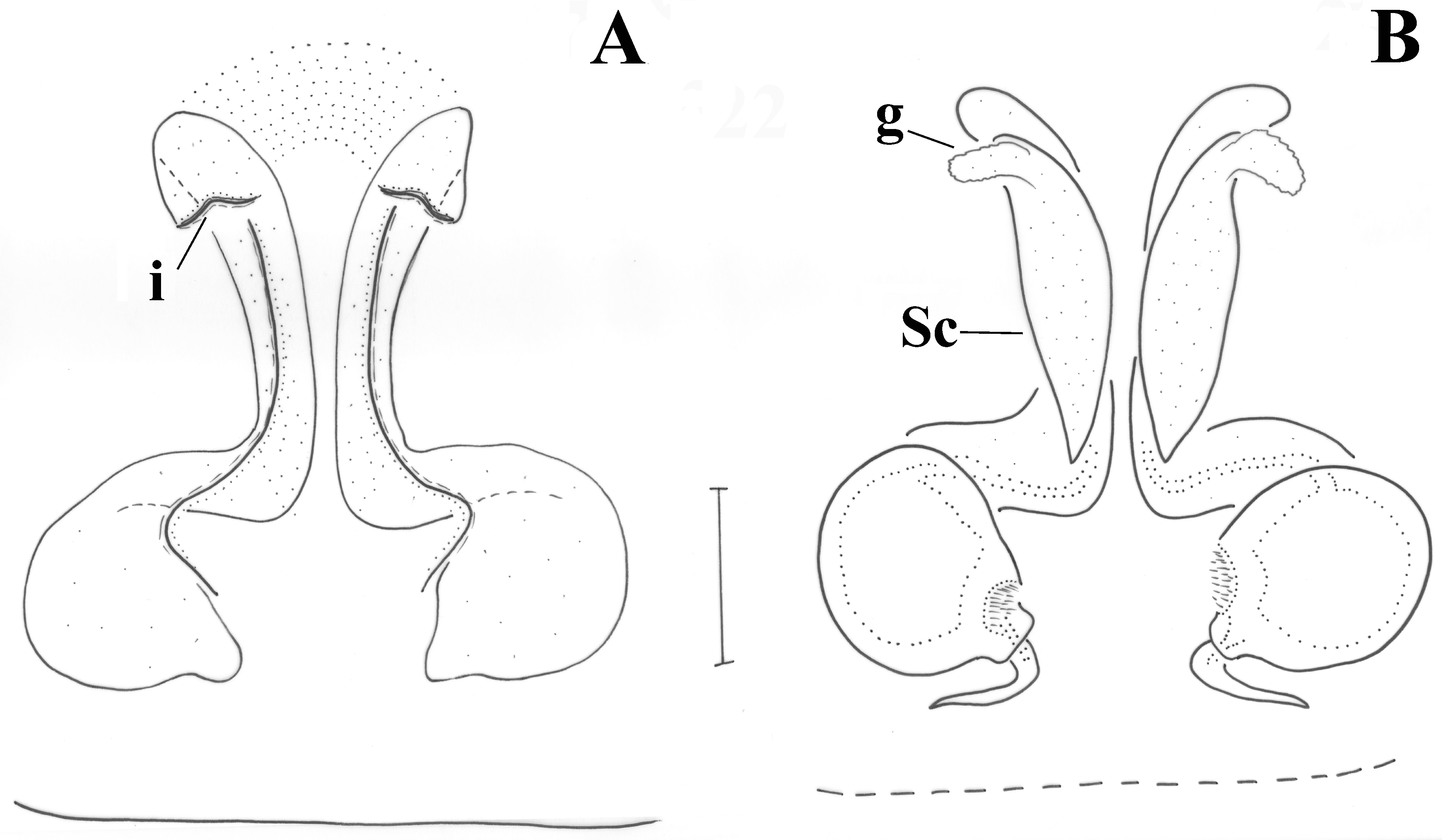
Epigyne as in
Figure 8A. Epigynal plate longer than wide; anterior margin absent; introductory orifices (i) transverse, at anterior part of epigyne; fovea narrow, widening posteriorly. Vulva as in
Figure 8B. Copulatory ducts tubular, parallel and almost touching each other in the middle, widening posteriorly to reach the spermathecae; additional spermathecal sacs (Sc) along the vertical part of the copulatory ducts, with short glands (g) anteriorly; distance between spermathecae ca. their diameter.
Drassyllus dadia Komnenov & Chatzaki, 2016
Figure 9.
Drassyllus dadianus n.sp. (holotype), and D. dadia (holotype), male pedipalps, ventral view. (A) D. dadianus n.sp., (B) D. dadia. p: transverse projection, DA: dorsal apophysis.
Figure 9.
Drassyllus dadianus n.sp. (holotype), and D. dadia (holotype), male pedipalps, ventral view. (A) D. dadianus n.sp., (B) D. dadia. p: transverse projection, DA: dorsal apophysis.
Material
Evros: Loutros-Dadia, N 32 km after, 360 m alt (41.0949 N 26.0930 E), grazed grassland pasture [FC: 17144, 15/05–22/06/2015], 2 ♀; Polia, 1km N, 126 m alt (41.4532 N 26.2291 E), young Quercus forest with high canopy cover and no signs of grazing [FC: 17159, 17/05–23/06/2015], 1 ♀; same [FC: 17256, 23/06–22/07/2015], 1 ♀—all legs M. Komnenov & K. Zografou.
Comments
In the original description of
D. dadia there is no reference to the filiform dorsal apophysis (DA) originating from the base of the tegulum and reaching the tip of the embolar projection retrolaterally (
Figure 9B). This apophysis is not evident in ventral view of the pedipalp (see [
8], p. 46, figs. 115, 120), and is usually hidden behind the embolus in ventral view, which is the reason why it was missed in the original description.
Distribution
Greece.
Drassyllus dadianus n.sp.
Drassyllus dadia Komnenov & Chatzaki, 2016: Evros district, Dadia, 1 ♂1 ♀ (misidentification)
Type Material
Holotype: male (NHMC, FC17159), Evros district, Polia, 1 km N, [41.4532 N 26.2291 E] young Quercus forest with high canopy cover and no signs of grazing, 17/05/2015–23/06/2015, leg. M. Komnenov. Paratypes: same—1 ♂ 2 ♀♀.
Material
Evros: Loutros-Dadia, N 1.5 km after (40.8969 N 26.0575 E), maquis with sparce openings [FC: 17136, 14/05–22/06/2015], 3 ♀♀; same [FC: 17186, 22/06–21/07/2015], 1 ♀; Loutros 1km N, towards Profitis Ilias (40.8899 N 26.0487 E), mixed forest with Quercus, Ostria, Juniperus and goat paths with visible grazing [FC: 17138, 14/05–22/06/2015], 2 ♂♂ 4 ♀♀; Loutros-Dadia, N 32 km after, 360 m alt (41.0949 N 26.0930 E), grazed grassland pasture [FC: 17144, 15/05–22/06/2015], 2 ♀♀; Polia, 1km N, 126 m alt (41.4532 N 26.2291 E), young Quercus forest with high canopy cover and no signs of grazing [FC: 17159, 17/05–23/06/2015], 8 ♂♂ 14 ♀♀; Polia-Ladi, 1.5 km after (41.4491 N 26.2382 E), dry grassland in an open area, near a Quercus forest [FC: 17161, 17/05–23/06/2015], 1 ♀—all legs M. Komnenov & K. Zografou; Evros Delta, Loutros spring (40.8422 N 26.0247 E), riverbank [FC: 12199, 13/05–17/06/2015], 2 ♀♀, Leg. M. Komnenov.
Comparative Material Examined
[sub D. dadia]: Evros district, Dadia: 1 ♀ (rural mosaics –FC: 12148, 41.0147N 26.1842E- 24/05/11–02/07/11, leg. K. Zografou) 1 ♂ (oak forest –FC: 12110, 41.0971N 26.1006E- 03/04/09–05/05/09, leg. K. Karakatsanis).
Etymology
The specific name refers to the close similarity of the new species to its syntopic Drassyllus dadia. It is an adjective in apposition.
Diagnosis
Drassyllus dadianus n.sp. is very similar to
D. dadia Komnenov & Chatzaki, 2016 ([
8], p. 46 figs. 114–122). Males are difficult to distinguish, but differ from
D. dadia by a longer embolus almost reaching the end of the cymbium (
Figure 9A) (in
D. dadia, it reaches less than half the distance between the end of the cymbium and alveolus,
Figure 9B) and by the shape of the terminal apophysis having sharper edges (smoother in
D. dadia). Other minor differences are found in the tibial apophysis (
Figure 6D) (in
D. dadianus n.sp. the difference in size between the two lobes is greater) and in the embolar projection (in
D. dadianus n.sp. the pointed crests are slightly sharper). The scutum of
D. dadianus n.sp. is longer, covering a bit more than half the abdominal surface, while in
D. dadia it hardly covers half of it. Females are easier to distinguish from
D. dadia by the anterior epigynal margin being continuous and without any protrusion, but instead with a small central recess (
Figure 7C) (narrow, with small central protrusion and disconnected from the introductory orifices in
D. dadia). Furthermore,
D. dadianus n.sp. males resemble
D. pusillus ([
12], p. 276, figs. 327–328) and
D. villicus ([
12], p. 276, figs. 329–330), but differ from both of them by the sharper edges of the terminal apophysis and the shape of the embolar base (transverse, with no prolateral hump in
D. villicus and more sclerotized with different retrolateral end in
D. pusillus). Females of
D. villicus are quite different from
D. dadianus n.sp. and those of
D. pusillus differ in the shape of the anterior margin and in the size and position of the lateral glands of spermathecae.
Description
Male (holotype). CL 1.97, CW 1.53. AL 2.28. Eye sizes and interdistances: AME 0.073, ALE 0.104, PME 0.091, PLE 0.079, PME oblique to almost transverse, almost touching, PME-PLE separated by less than their diameter, AME separated by ca. their diameter, AME-ALE almost touching. Chelicerae with 4 teeth on PM and 2 denticles on RM. Legs orange, except femur and coxa, which are blackish brown. Leg spination: Leg I: Fe 2d, Ti 1v, Mt 2v; leg II: Fe 2d, Ti 2v, Mt 2v; leg III: Fe 5d, Ti/Mt spinose; leg IV: Fe 4d, Ti/Mt spinose. Μetatarsi: I and II with additional short spinny setae in two rows. Abdomen grey, dorsally with large rectangular scutum covering more than 1/2 of the abdomen.
Pedipalp as in
Figure 6D and
Figure 9A. Tibia with stiff setae dorsolaterally; retrolateral tibial apophysis with two pointed tips like the human thumb and index finger; terminal apophysis rectangular, approximately as wide as long, with distal surface almost straight, transverse and with pointed edges; base of terminal apophysis with retrolaterally directed transverse projection (p), needle-shaped; embolar projection (EmP) transverse, sclerotized, with blunt end and forming two small crests; a membrane is filling the space between the embolus and the embolar projection; embolus long, filiform, almost reaching the end of the cymbium; dorsal apophysis (DA) straight and slender, filiform, reaching the embolar projection.
Female(paratype). CL 1.67, CW 1.34, AL 2.26. Eye sizes and interdistances: AME 0.062, ALE 0.088, PME 0.082, PLE 0.072, PME oblique to almost transverse, almost touching, PME-PLE separated by ca. ½ the diameter of PLE, AME separated by ca. their diameter, AME-ALE almost touching. Chelicerae with 5 teeth on PM and 3 denticles on RM. Legs orange, except femur and coxa, which are blackish brown. Leg spination: Leg I: Fe 2d, Mt 2v; leg II: Fe 2d, Ti 1v, Mt 2v; leg III: Fe 3d, Ti/Mt spinose; leg IV: Fe 3d, Ti/Mt spinose. Μetatarsi I and II with additional short spinny setae in two rows.
Epigyne as in
Figure 7C. Epigynal plate longer than wide; anterior margin continuous, almost straight, with a small central sclerotized recess, narrower than maximum epigynal width. Introductory orifices not evident but most probably at the outer rims of the anterior margins. Vulva as in
Figure 7D. Copulatory ducts semi-circular, parallel in the middle, widening posteriorly to reach the spermathecae; lateral glands (g) at the middle of copulatory ducts; distance between spermathecae ca. half of their diameter.
Comments
In the course of examining the new material from the area of Evros district, interesting findings considering the fine differences in the morphology of the
Drassyllus “cf.
dadia” specimens were detected, which led the authors to decide on this new species (see diagnosis). At the same time, the closer examination of the older material collected from this region revealed some deviations from the original description of
D. dadia [
8], which are considered here (see
D. dadia) as well as the misplacement of some individuals attributed to it. Apart from being morphologically very close, the two species are sympatric and even syntopic.
Haplodrassus caspius Ponomarev & Belosludtsev, 2008
Material
Evros: Evros Delta, Drana (40.8255 N 26.0163 E), marsh meadow [FC: 17084, 13/05–17/06/2015], 4 ♂♂; Evros Delta, water reserve between E and W part (40.7982 N 26.0718 E) [FC: 17086, 13/05–17/06/2015], 1 ♂—all leg. M. Komnenov.
Comments
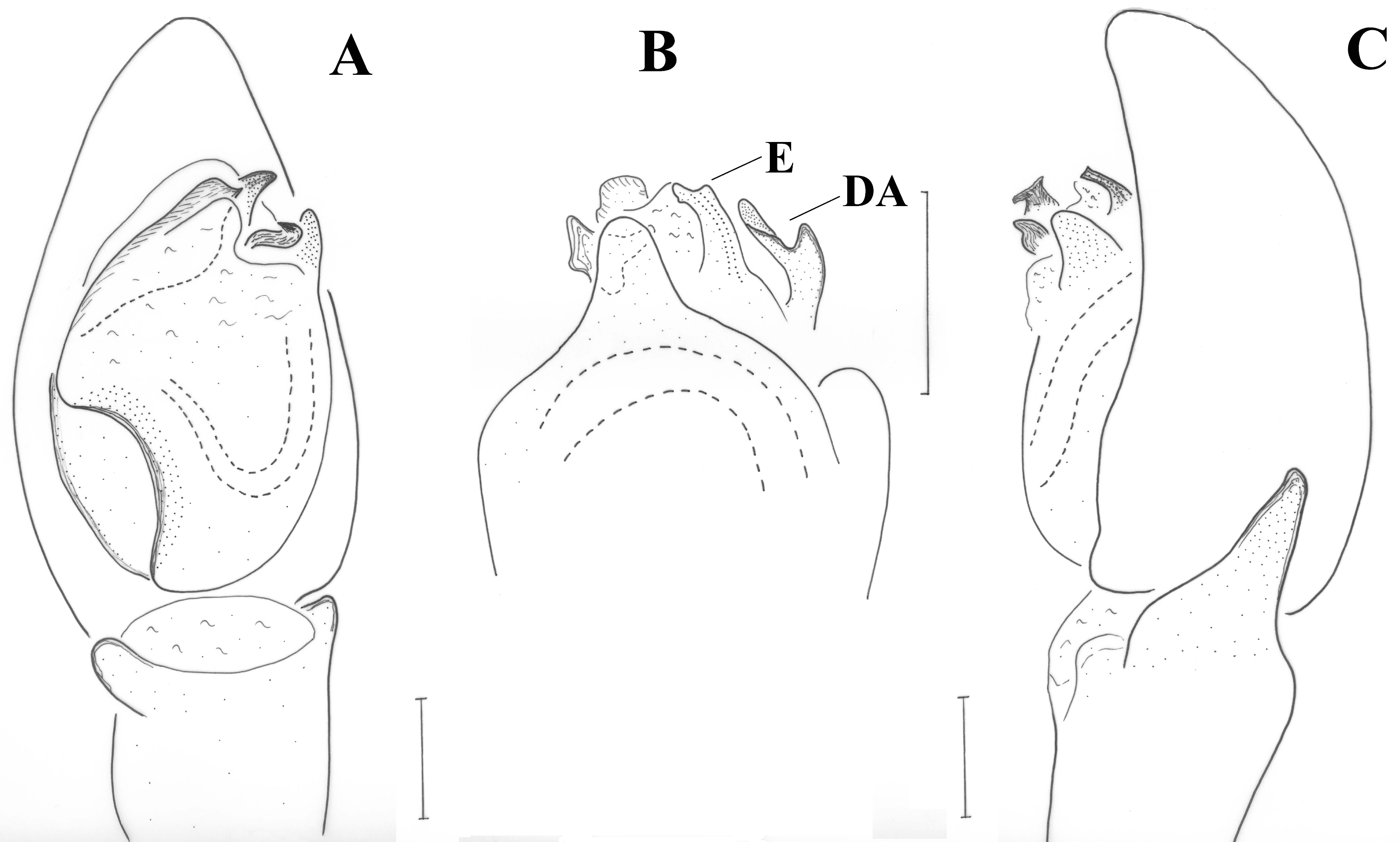
Following the re-description by Kovblyuk et al. [
16], the specimens conform to
Haplodrassus caspius. According to the authors, an interesting diagnostic character is the shape of the embolus, described as “hook-like”. Apart from this strange form not stated before for any gnaphosid species, the close examination of the male palp at any angle or side did not result in such shape of the embolus. In dorsal view, however, the embolus has a tapering end with two lobes (see
Figure 10B), which is perhaps what the authors misinterpreted as hook-like. This examination revealed an additional diagnostic character: a dorsal bifid apophysis (DA) at the prolateral side of the embolus, (
Figure 10B).
Haplodrassus caspius is a new record for Greece.
Distribution
Russia, Kazakhstan, Azerbaijan, Greece.
Phaeocedus similaris n.sp.
Type Material
Holotype: male (NHMC, FC17152), Evros district, Loutros-Pefka, 2 km after, [40.8835 N 26.0379 E] maquis with thorny substrate and some Pinus, Juniperus, Ostria, Quercus individuals, 16/05/2015–22/06/2015, leg. M. Komnenov.
Comparative Material Examined
P. hebraeus Levy, 1999: Holotype male (HUJ 11948, Poriya, 20.III.1940, leg. A. Shulov).
Etymology
The specific name refers to the close similarity of the two congeneric species, namely P. hebraeus and P. vankeeri Chatzaki, 2019. It is an adjective in apposition.
Diagnosis
Males of this species resemble
P. hebraeus and
P. vankeeri by having a spermophore with vertical coils, retrolateral tibial apophysis diagonally directed and with a single pointed tip, and by the number and shape of the white bands in the opisthosoma (elements that distinguish them from
P. braccatus (L. Koch, 1866)). From both
P. hebraeus and
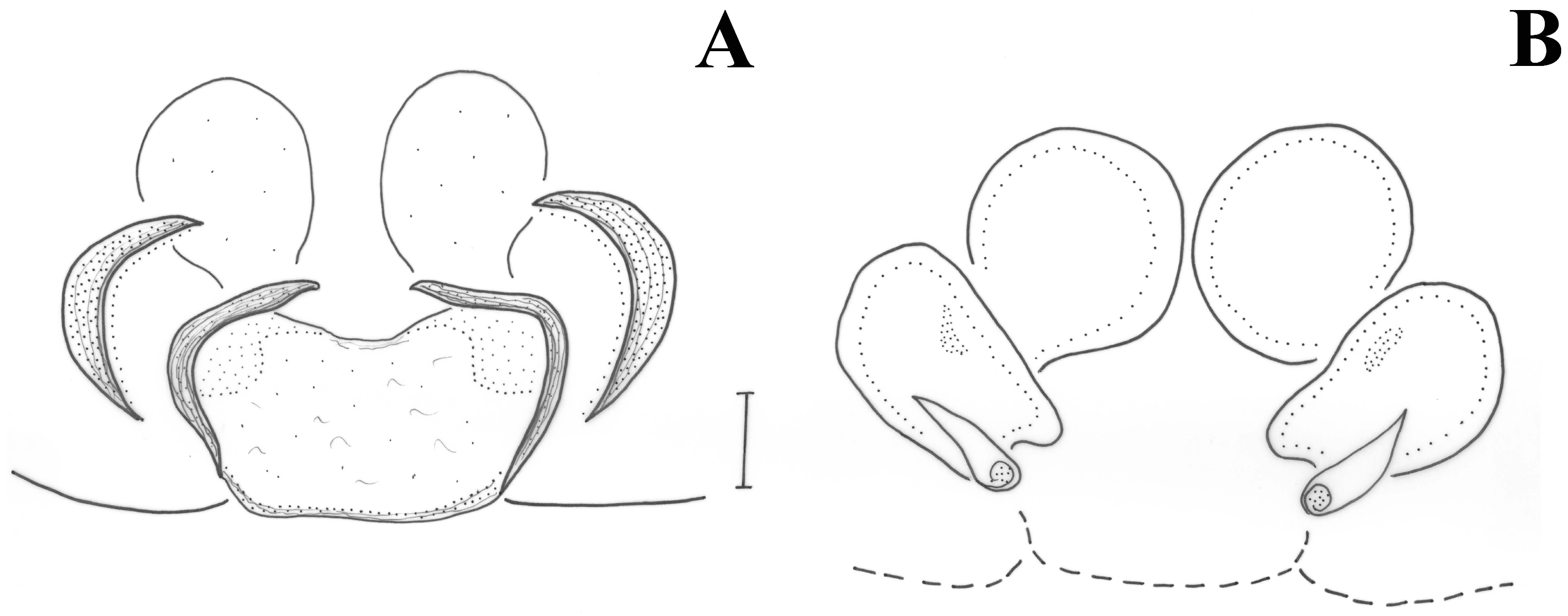
P. vankeeri, the new species differs by the shape and size of the abdominal scutum (in
P. hebraeus it is long and narrow, covering more than ½ of the abdominal length and ca. 1/3 of the abdominal width, in
P. vankeeri it is long and wide covering ca. ½ of the abdominal length and almost all abdominal width (
Figure 11B) and in
P. similaris n.sp. it is short and relatively wide, covering less than ½ of the abdominal length and ca. 1/2 of the abdominal width (
Figure 11A)). The conductor of
P. similaris n.sp. is more similar to that of
P. hebraeus with a relatively enlarged membranous fold (
Figure 12A) at the apical part when compared to
P. vankeeri (
Figure 12B). In the initial examination and draft illustration of the single specimen available (holotype) in 2017, the embolus of
P. similaris n.sp. presented a small filiform free part (drawn in dotted lines in
Figure 13A,B,D), absent in both other species, but also absent from the specimen at the time that this paper is being prepared. This leads to the logical suspicion that all three species have a small free part in their emboli.
Description
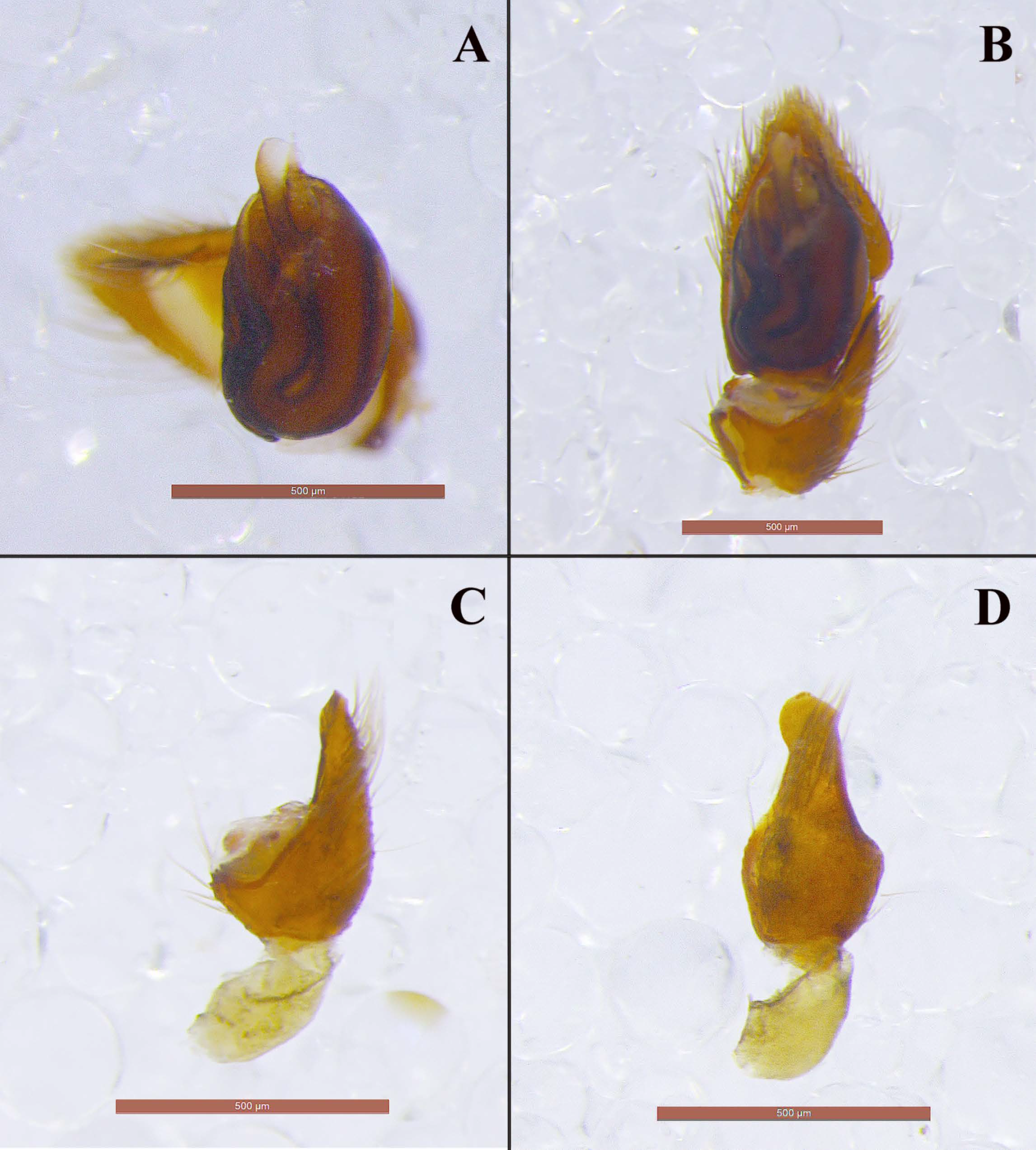
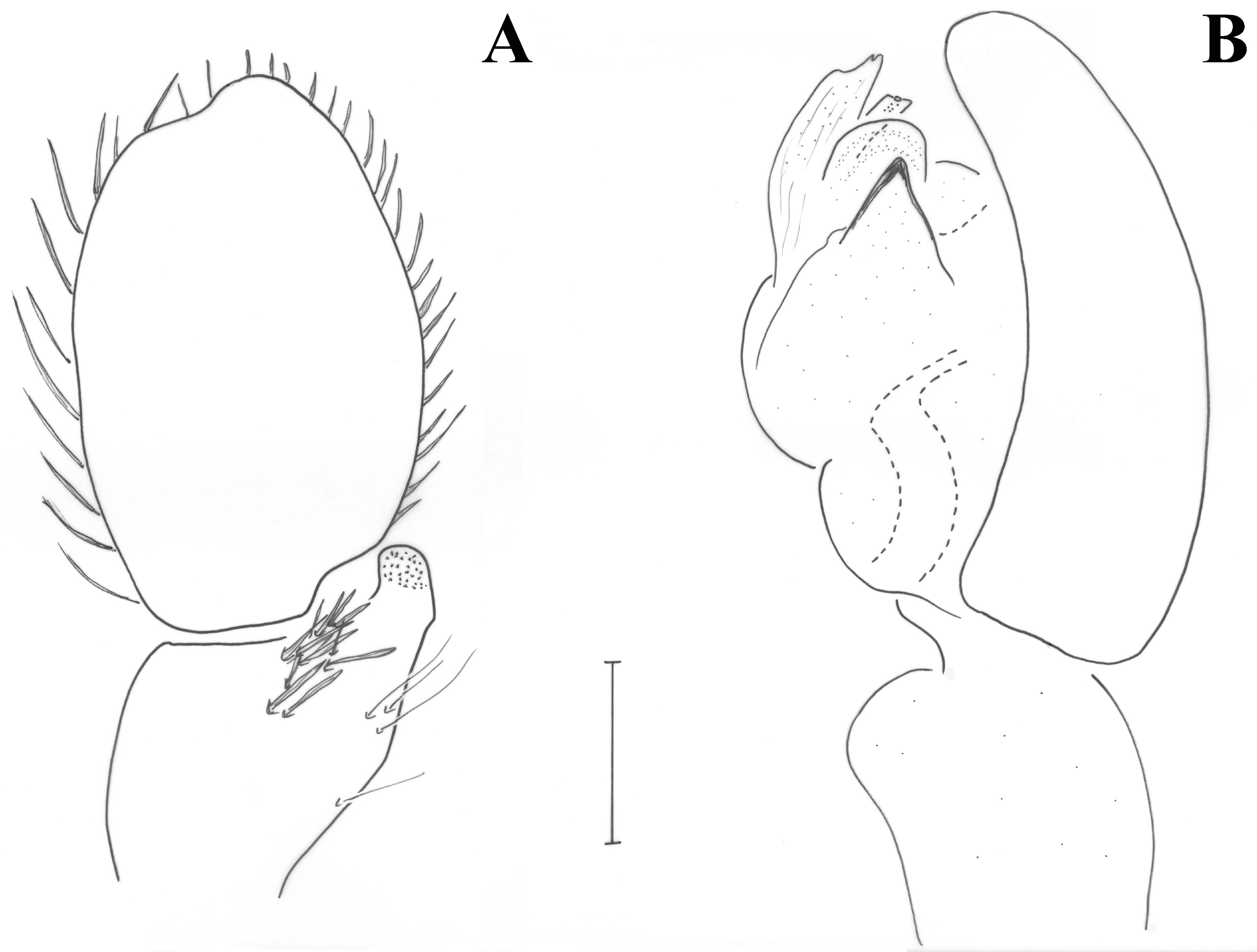
Male (holotype). Medium sized spiders of dark brown color (
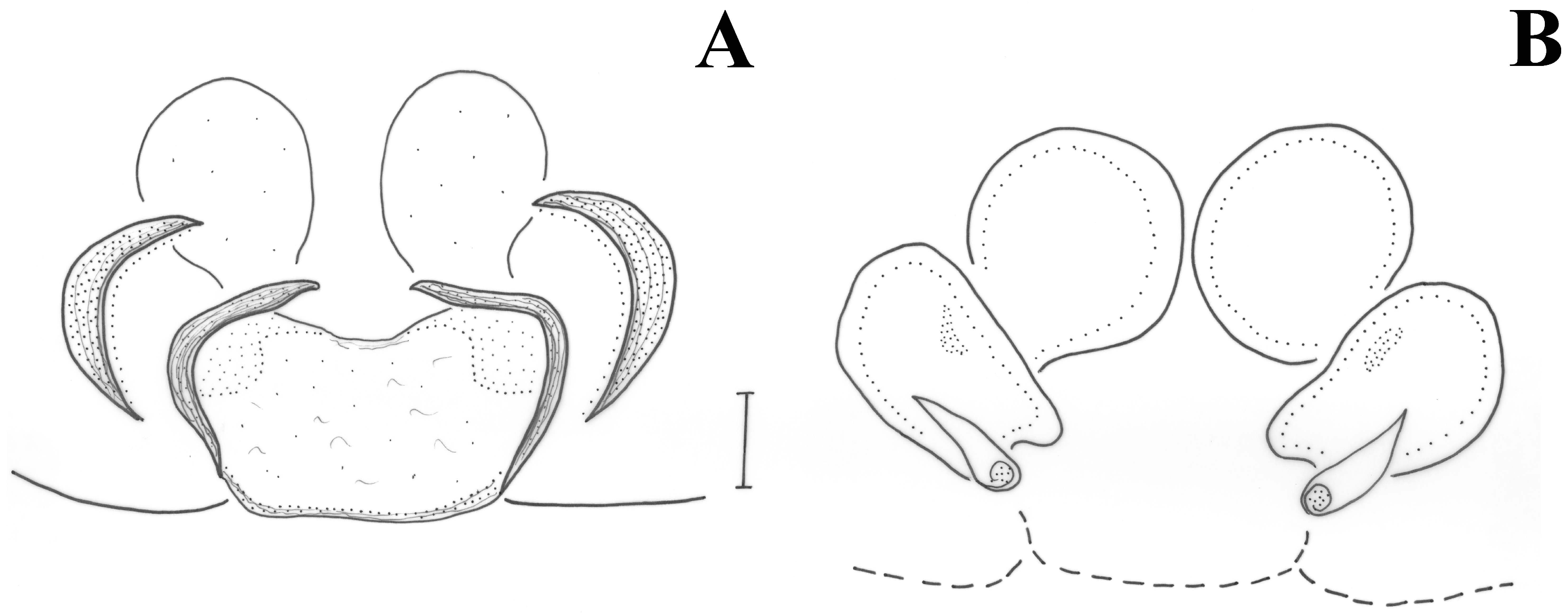
Figure 11A). Opisthosoma with two median white stripes and two at its anterior end. Legs uniformly brown. Measurements: CL 2.09, CW 1.47, AL 2.68. Eye sizes and interdistances: AME 0.069, PME 0.061, PLE 0.077, ALE 0.121, PME-PME 0.104, PME-PLE 0.082, AME-AME 0.042, AME-ALE 0.03 (almost touching). AME black, PME white, lateral eyes with black surroundings. PER straight, AER slightly recurved. Prosoma long with slightly widening thoracic region, fovea about 1/4 of abdominal length. Chelicerae toothless but with crest with flat upper surface. Scutum medium-sized, rectangular, covering less than half of the abdominal dorsal length and ca. half its width (at the base). ALS long, cylindrical, separated by more than their diameter. Leg formula is IV > I > II > III. Legs spination: Leg I: Fe 4d, Ti 2v, Mt 2v; leg II: Fe 3d, Ti 2v, Mt 2v; leg III: Fe 7d, Pa 1rl, Ti/Mt spinose; leg IV: Fe 4d, Ti/Mt spinose. Ta not densely scopulated.
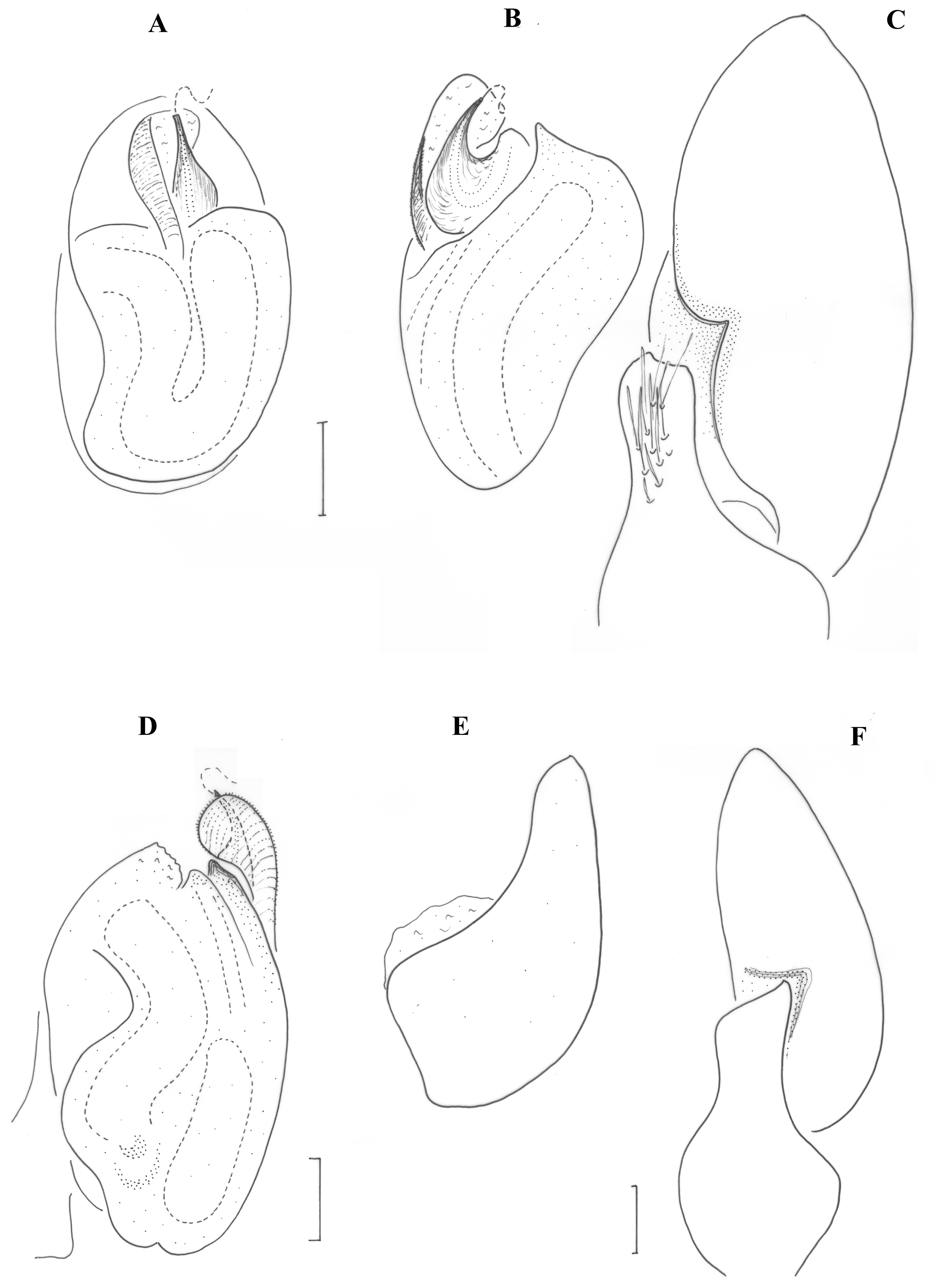
Pedipalp as in
Figure 12A,C,D and
Figure 13. Large retrolateral tibial apophysis diagonally directed and accommodated into cymbium depression, with round ventral edge and tapering dorsal tip, taking various shapes in different angles (see
Figure 12C,D and
Figure 13C,E,F). Large spermophore U-shaped. Embolus with broad round base hidden behind conductor in ventral view, tapering at its end, and terminating in a fine filiform free part. Conductor with stout vertical part and enlarged membranous fold encircling the embolus.
Zelotes histio n.sp.
Type Material
Holotype: female (NHMC, FC17161), Evros district, Polia-Ladi, 1.5 km after, [41.4491 N 26.2382 E] dry grassland in an open area, near a Quercus forest, 17/05/2015–23/06/2015, leg. M. Komnenov & K. Zografou.
Other Material
Penultimate form: Evros: Evros Delta, water reserve between E and W part (40.7982 N 26.0718 E) [FC: 17086, 13/05-17/06/2015], 1 ♀, leg. M. Komnenov; Aisymi-Mega Dereio, 5 km before, 580 m alt (41.2143 N 25.9930 E), broad-leaved Quercus forest with some Juniperus at the substrate [FC: 17147, 15/05-23/06/2015], 1 ♀; Mega Dereio-Mikro Dereio, 9 km after, 166 m alt (41.2776 N 26.0671 E), grassland within Quercus / Pinus mixed forest and riverine vegetation, water in 50 m distance [FC: 17153, 16/05-23/06/2015], 2 ♀♀—all legs M. Komnenov & K. Zografou.
Fully developed form (like type): Evros, Loutros-Dadia, N 1.5 km after (40.8969 N 26.0575 E), maquis with sparse openings [FC: 17136, 14/05-22/06/2015], 2 ♀♀; Polia-Ladi, 1.5 km after (41.4491 N 26.2382 E), dry grassland in an open area, near a Quercus forest [FC: 17161, 17/05–23/06/2015], 1 ♀, all legs M. Komnenov & K. Zografou.
Etymology
The specific name refers to the peculiar shape of the epigynum resembling a sail (ιστίο [histio] in Greek). It is a noun in apposition.
Diagnosis
The female of this species is distinguished from any other
Zelotes by the epigynal sclerotizations with two ventral S-shaped lines converging posteriorly, two dorsal lines like inverted “L” lines, and by the very small spermathecae of the vulva shaped as a droplet. The examination of the specimens attributed to this species reveals two very closely related but different forms in terms of both habitus (
Figure 14A,B) and epigynal basic structures (
Figure 15A–D). Of them, one form (
Figure 14B and
Figure 15C,D) seems to belong to not fully mature specimens (smaller individuals with lighter color), still, with some sclerotizations on their epigyne, making it possible to be dissected. Hence we present both forms here in order to better illustrate the forms in which the species may be found in the field.
Description
Female (holotype). Medium-sized spiders of dark brown color (
Figure 14). Legs uniformly brown, except for tarsi which are of lighter color (in the penultimate form tarsi and metatarsi are yellow,
Figure 14B). Measurements (in parenthesis metrics for penultimate specimen from Site 28a): CL 2.92 (2.08), CW 2.27 (1.74), AL 3.37 (3.27). Eye sizes and interdistances (Site 28a): AME 0.069 (0.054), PME 0.090 (0.086), PLE 0.083 (0.076), ALE 0.105 (0.094), PME almost touching (same), PME-PLE separated by ca. the PLE diameter—0.080 (0.054), AME separated by ca. their diameter −0.054 (same), AME-ALE almost touching (same). PER & AER almost straight (slightly procurved to almost straight). Prosoma long with widening thoracic region, fovea about 1/4 of abdominal length. Chelicerae with 3 teeth +1 smaller on PM (4 teeth) and 1 tooth + 1denticle on RM (2 denticles). ALS long, cylindrical, separated by more than their diameter. Leg formula is IV > I > II > III. Legs spination (in parenthesis metrics for variation 1): Leg I: Fe 3d (2d), Mt 2v (2 or 3v); leg II: Fe 3d (2d), (Ti 2v), Mt 2v (same); leg III: Fe 5d (same), Pa 1rl (same), Ti/Mt spinose (same); leg IV: Fe 5d (3d), Pa 1rl (-), Ti/Mt spinose (same).
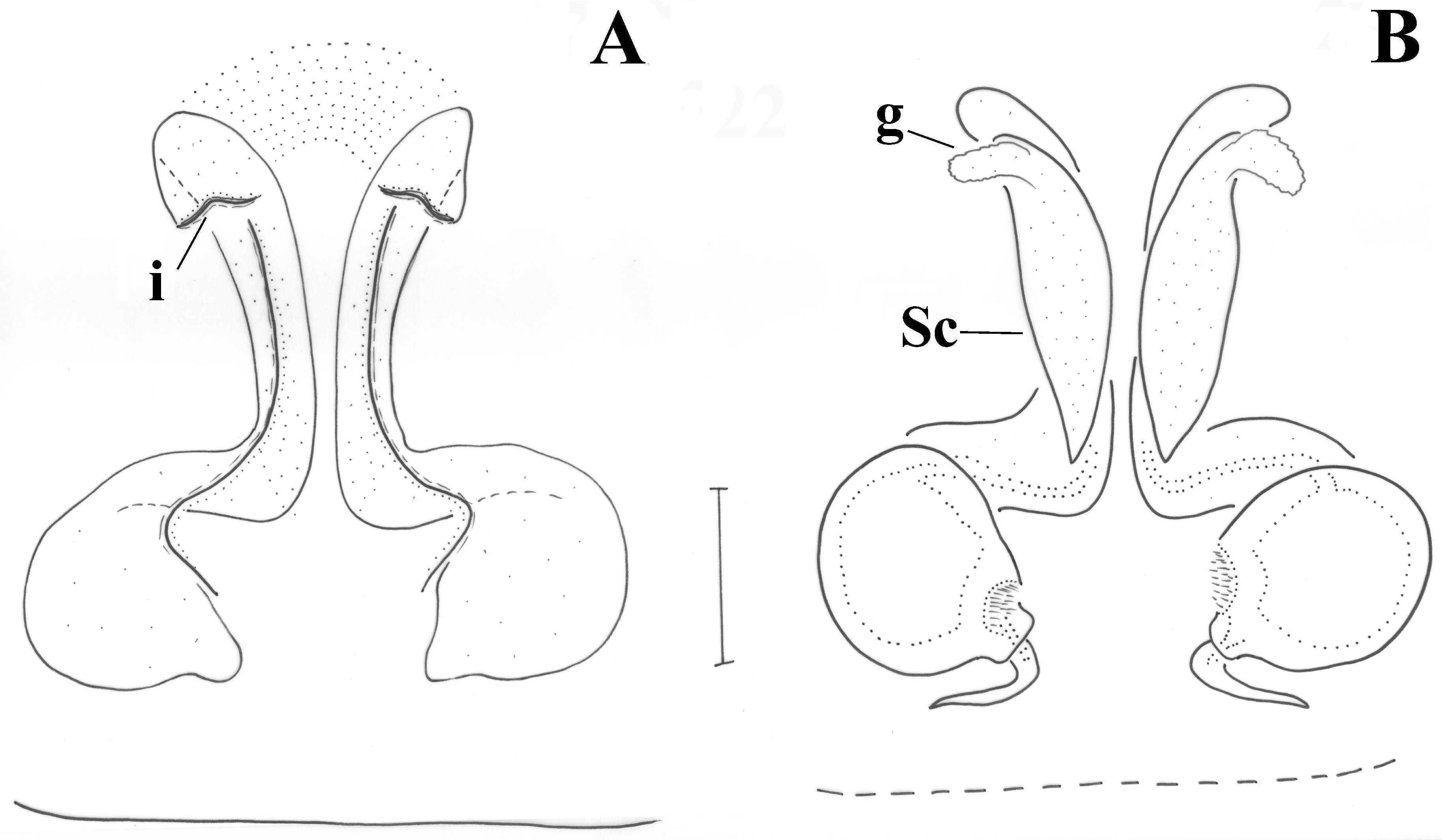
Epigyne as in
Figure 15A,C. Lateral margins like two slightly curved and diverging lines; no anterior margin present. In the penultimate form, there appear two lines like inverted L’s (
Figure 15C), which in the fully developed holotype are only evident viewed from dorsally (
Figure 15B). Introductory openings towards posterior end. Vulva as in
Figure 15B,D. Spermathecae (Sp) of peculiar shape like transverse droplets; copulatory ducts small, straight. Two additional sacs (Sc) appear just above the L-shaped sclerotizations. In the penultimate form, these sacs, as well as the transverse spermathecae are hardly formed (they appear rather globular); no fertilization ducts are evident in them (
Figure 15D).
Zelotes olympi (Kulczyński, 1903)
Material
Florina, Prespes, 1.9 km NW of Pisoderi (40.79085 N 21.23345 E), beech forest edges, ruderal strip, dry and wet meadows [FC: 17048, 24/06/2014], 1 ♀, leg. C. Kropf et al.
Comments
Z. olympi was initially described from Turkey (Mt. Olympus Bithynicus in Asia Minor), an area which was misinterpreted by Bosmans & Chatzaki [
17] as belonging to the Greek mountain Olympus in Thessalia. Chatzaki [
18] commented on this mistake and further examined material of
Z. olympi (the holotype and a couple of specimens from Bulgaria) as well as the holotype of the very similar
Z. talpinus (L. Koch, 1872). She concluded that the first species does not occur in Greece, and the latter most probably does not occur in Bulgaria. Hence, the two species were correspondingly excluded from the checklists of Greece and Bulgaria, and therefore
Z. talpinus was annulated from the lists of the Balkan Peninsula.
A single female found in the north-west of Greece (Prespes Lake) re-establishes
Z. olympi in the Greek arachnofauna, since the configuration of its genitalia seems to conform to the diagnostic characters of this species.
Sensu stricto, this female would be an intermediate variant of
Z. olympi and
Z. talpinus, because the main point to distinguish these two species is the level of curvature of the anterior part of the lateral margins ([
19], figs. 3: 1–3), in
Z. olympi being almost straight and parallel and in
Z. talpinus being S-shaped. In our specimen, the lateral margins are not S-shaped in their anterior part, but are not parallel either. In addition, the median plate is wider with respect to the epigynal width when compared to
Z. olympi (see
Figure 17). In fact, the width of the median plate formed by the lateral margins is 1/2 its length in
Z. olympi, as long as wide in
Z. talpinus and 2/3 its length in our specimen. In the absence of a matching male, in which differences between the two species are clearer, this identification is tentative, although our female is mostly similar to
Z. olympi. It is possible that the two species belong to a species complex presenting a gradient of populations with small differences and lots of variants.
Zelotes olympi is a new record for Greece.
Distribution
Bulgaria, Turkey, Ukraine, Greece.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}