
Monocytes in Neonatal Bacterial Sepsis: Think Tank or Workhorse?

{kind=link}
{kind=link}
Abstract
1. Introduction
2. Monocytes and Their Role in Processes of Bacterial Sepsis and Inflammation
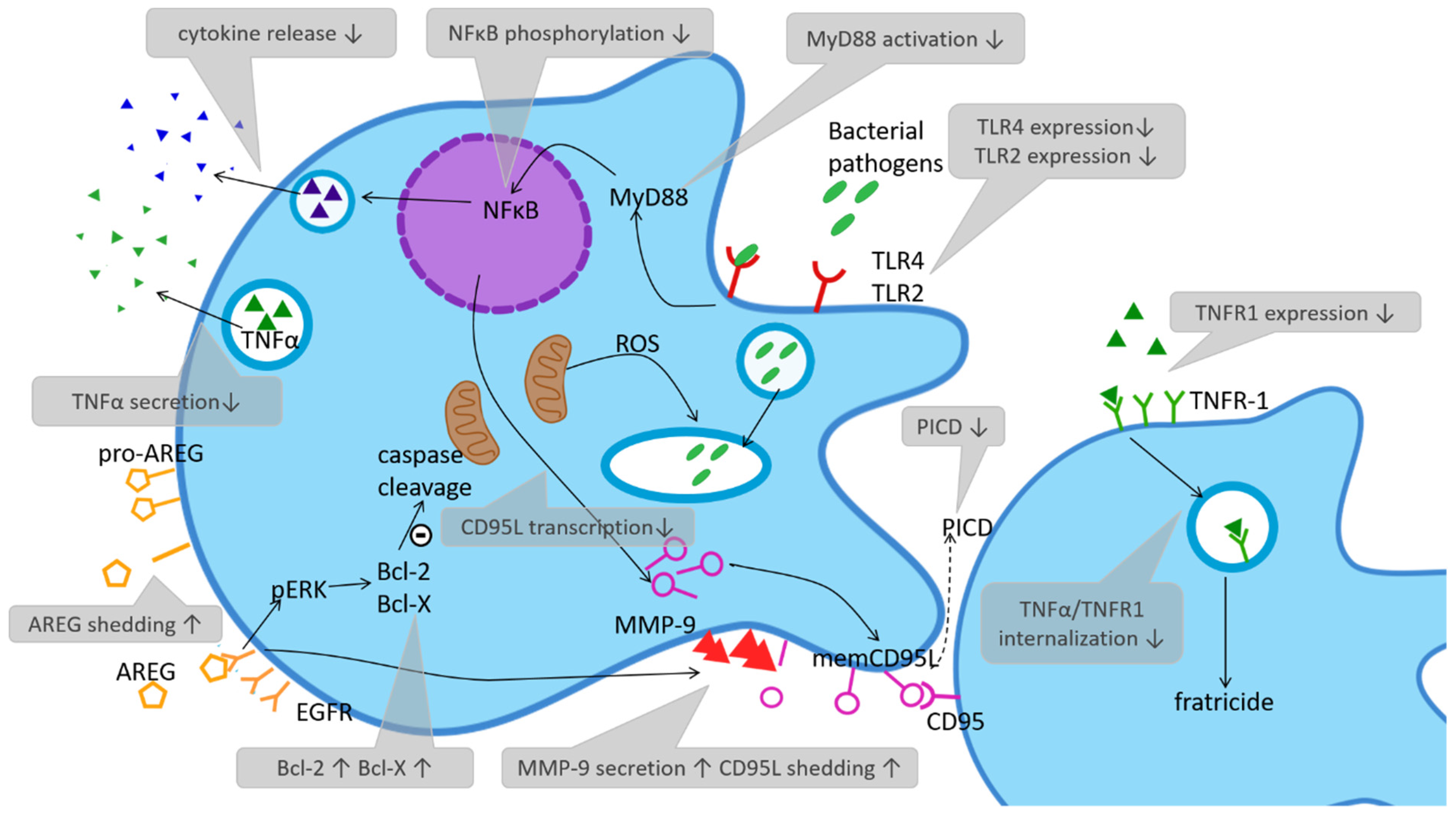
2.1. Detection of Pathogens
2.2. Phagocytosis and Killing
2.3. Phagocytose Induced Cell Death (PICD)—Regulating the Immune Response via Depletion
2.4. Monocytes in Organ Destruction
2.5. Functions of Monocyte Subsets
2.6. Cytokine Production and Responsiveness
2.7. Modulating Effector Cell Functions of Monocytes in Neonatal Sepsis
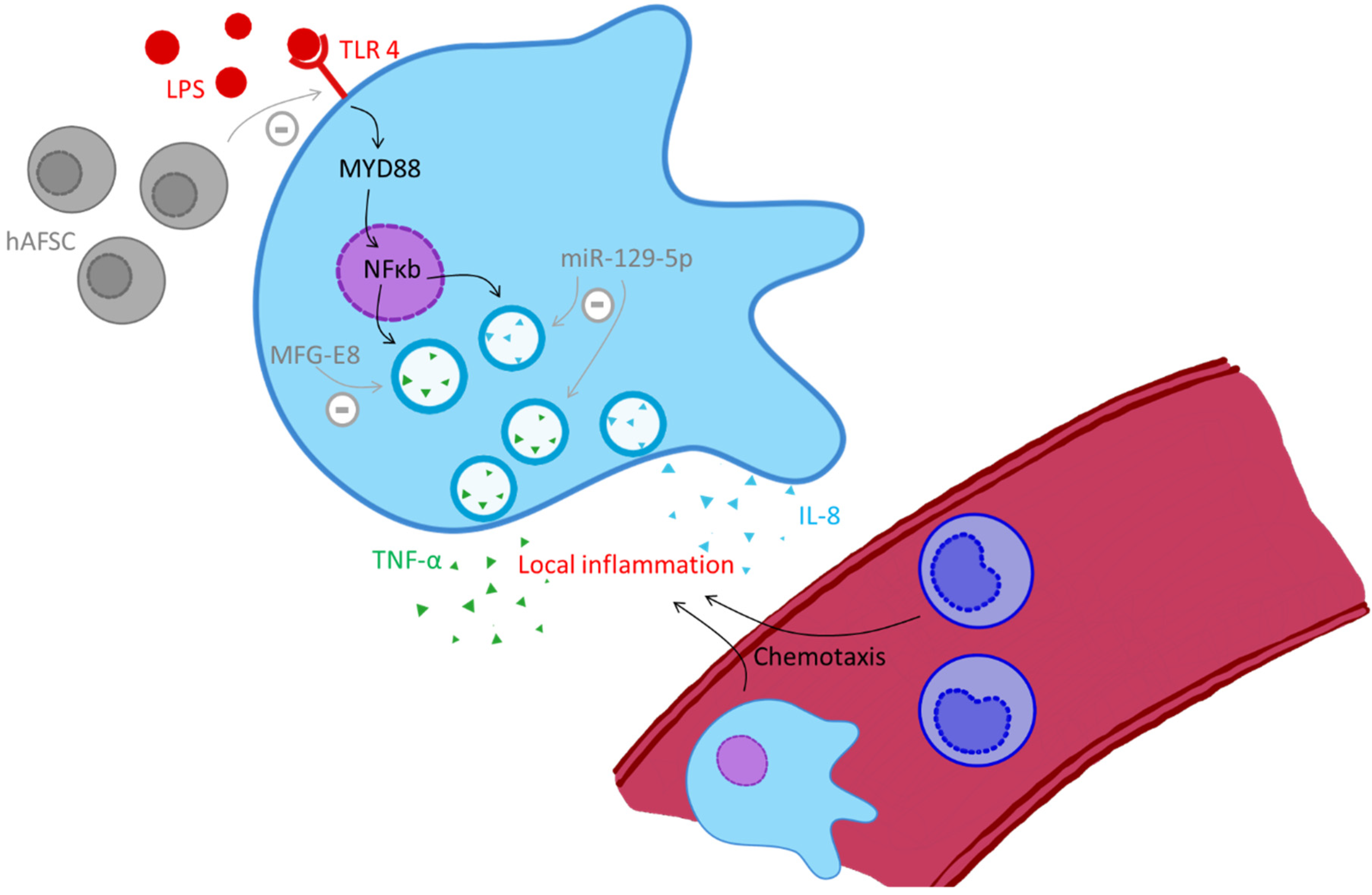
2.8. S100-Alarmins in Neonatal Sepsis
2.9. Immunometabolism of Neonatal Monocytes
3. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, L.; Oza, S.; Hogan, D.; Chu, Y.; Perin, J.; Zhu, J.; Lawn, J.E.; Cousens, S.; Mathers, C.; Black, R.E. Global, regional, and national causes of under-5 mortality in 2000–2015: An updated systematic analysis with implications for the Sustainable Development Goals. Lancet 2016, 388, 3027–3035. [Google Scholar] [CrossRef]
- Fleischmann-Struzek, C.; Goldfarb, D.M.; Schlattmann, P.; Schlapbach, L.J.; Reinhart, K.; Kissoon, N. The global burden of paediatric and neonatal sepsis: A systematic review. Lancet Respir Med. 2018, 6, 223–230. [Google Scholar] [CrossRef]
- Jong, d.E.; Strunk, T.; Burgner, D.; Lavoie, P.M.; Currie, A. The phenotype and function of preterm infant monocytes: Implications for susceptibility to infection. J. Leukoc. Biol. 2017, 102, 645–656. [Google Scholar] [CrossRef] [PubMed]
- Strunk, T.; Hibbert, J.; Doherty, D.; Nathan, E.; Simmer, K.; Richmond, P.; Currie, A.; Burgner, D. Impaired Cytokine Responses to Live Staphylococcus epidermidis in Preterm Infants Precede Gram-positive, Late-onset Sepsis. Clin. Infect. Dis. 2021, 72, 27. [Google Scholar] [CrossRef]
- Bone, R.C.; Balk, R.A.; Cerra, F.B.; Dellinger, R.P.; Fein, A.M.; Knaus, W.A.; Schein, R.M.; Sibbald, W.J. Definitions for sepsis and organ failure and guidelines for the use of innovative therapies in sepsis. The ACCP/SCCM Consensus Conference Committee. American College of Chest Physicians/Society of Critical Care Medicine. Chest 1992, 101, 1644–1655. [Google Scholar] [CrossRef]
- Cai, S.; Thompson, D.K.; Anderson, P.J.; Yang, J.Y. Short- and Long-Term Neurodevelopmental Outcomes of Very Preterm Infants with Neonatal Sepsis: A Systematic Review and Meta-Analysis. Children 2019, 6, 131. [Google Scholar] [CrossRef] [PubMed]
- Jung, E.; Romero, R.; Yeo, L.; Diaz-Primera, R.; Marin-Concha, J.; Para, R.; Lopez, A.M.; Pacora, N. Gomez-Lopez, P.; Yoon, B.H.; et al. The fetal inflammatory response syndrome: The origins of a concept, pathophysiology, diagnosis, and obstetrical implications. Semin Fetal Neonatal Med. 2020, 25, 101146. [Google Scholar] [CrossRef]
- Hotchkiss, R.S.; Coopersmith, C.M.; McDunn, J.W.; Ferguson, T.A. The sepsis seesaw: Tilting toward immunosuppression. Nat. Med. 2009, 15, 496–497. [Google Scholar] [CrossRef]
- Janeway, C.A. Approaching the asymptote? Evolution and revolution in immunology. Cold Spring Harb. Symp. Quant Biol. 1989, 54, 1–13. [Google Scholar] [CrossRef]
- Underhill, D.M.; Ozinsky, A. Toll-like receptors: Key mediators of microbe detection. Curr. Opin. Immunol. 2002, 14, 103–110. [Google Scholar] [CrossRef]
- Viemann, D.; Dubbel, G.; Schleifenbaum, S.; Harms, E.; Sorg, C.; Roth, J. Expression of toll-like receptors in neonatal sepsis. Pediatr Res. 2005, 58, 654–659. [Google Scholar] [CrossRef]
- Sadeghi, K.; Berger, A.; Langgartner, M.; Prusa, A.; Hayde, M.; Herkner, K.; Pollak, A.; Spittler, A.; Förster-Waldl, E. Immaturity of infection control in preterm and term newborns is associated with impaired toll-like receptor signaling. J. Infect. Dis. 2007, 195, 296–302. [Google Scholar] [CrossRef] [PubMed]
- Yua, S.; Zhanga, W.; Yua, J.; Feng, S.; Guob, T.; Lua, H. Inhibiting effect of Radix Hedysari Polysaccharide (HPS) on endotoxin-induced uveitis in rats. Immunopharmacology 2014, 21, 361–368. [Google Scholar] [CrossRef] [PubMed]
- Terashima-Hasegawa, M.; Ashino, T.; Kawazoe, Y.; Shiba, T.; Manabe, A.; Numazawa, S. Inorganic polyphosphate protects against lipopolysaccharide-induced lethality and tissue injury through regulation of macrophage recruitment. Biochem. Pharmacol. 2019, 159, 96–105. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, K.; Ichiyama, T.; Isumi, H.; Nakata, M.; Sase, M.; Furukawa, S. NF-kappaB activation in peripheral blood mononuclear cells in neonatal asphyxia. Clin. Exp. Immunol. 2004, 132, 261–264. [Google Scholar] [CrossRef] [PubMed]
- Wisgrill, L.; Groschopf, A.; Herndl, E.; Sadeghi, K.; Spittler, A.; Berger, A.; Forster-Waldl, E. Reduced TNF-alpha response in preterm neonates is associated with impaired nonclassic monocyte function. J. Leukoc. Biol. 2016, 100, 607–612. [Google Scholar] [CrossRef]
- Hegge, I.; Niepel, F.; Lange, A.; Vogelgesang, A.; Heckmann, M.; Ruhnau, J. Functional analysis of granulocyte and monocyte subpopulations in neonates. Mol. Cell Pediatr. 2019, 1, 5. [Google Scholar] [CrossRef]
- Dreschers, S.; Saupp, P.; Hornef, M.; Prehn, A.; Platen, C.; Morschhauser, J.; Orlikowsky, T.W. Reduced PICD in Monocytes Mounts Altered Neonate Immune Response to Candida albicans. PLoS ONE. 2016, 11, e0166648. [Google Scholar]
- Sun, J.; Katz, S.; Dutta, B.; Wang, Z.; Fraser, I.D. Genome-wide siRNA screen of genes regulating the LPS-induced TNF-α response in human macrophages. Sci. Data. 2017, 4, 170007. [Google Scholar] [CrossRef] [PubMed]
- Inohara, N.; Chamaillard, M.; McDonald, C.; Nuñez, G. NOD-LRR proteins: Role in host-microbial interactions and inflammatory disease. Annu. Rev. Biochem. 2005, 74, 355–383. [Google Scholar] [CrossRef] [PubMed]
- Franchi, L.; Warner, N.; Viani, K.; Nuñez, G. Function of Nod-like receptors in microbial recognition and host defense. Immunol. Rev. 2009, 227, 106–128. [Google Scholar] [CrossRef]
- Chen, S.; Zuo, Y.; Huang, L.; Sherchan, P.; Zhang, J.; Yu, Z.; Peng, J.; Zhang, J.; Zhao, L.; Doycheva, D.; et al. The MC 4 receptor agonist RO27–3225 inhibits NLRP1-dependent neuronal pyroptosis via the ASK1/JNK/p38 MAPK pathway in a mouse model of intracerebral haemorrhage. Br. J. Pharmacol. 2019, 176, 1341–1356. [Google Scholar] [CrossRef]
- Silveira-Lessa, A.L.; Quinello, C.; Lima, L.; Redondo, A.C.C.; Ceccon, M.; Carneiro-Sampaio, M.; Palmeira, P. TLR expression, phagocytosis and oxidative burst in healthy and septic newborns in response to Gram-negative and Gram-positive rods. Hum. Immunol. 2016, 77, 972–980. [Google Scholar] [CrossRef] [PubMed]
- Gille, C.; Leiber, A.; Mundle, J.; Spring, B.; Abele, H.; Spellerberg, B.; Hartmann, H.; Poets, F.C.; Orlikowsky, T.W. Phagocytosis and postphagocytic reaction of cord blood and adult blood monocyte after infection with green fluorescent protein-labeled Escherichia coli and group B Streptococci. Cytometry. B Clin. Cytom. 2009, 76, 271–284. [Google Scholar] [CrossRef] [PubMed]
- Dreschers, S.; Gille, C.; Haas, M.; Seubert, F.; Platen, C.; Orlikowsky, T.W. Reduced internalization of TNF-a/TNFR1 down-regulates caspase dependent phagocytosis induced cell death (PICD) in neonatal monocytes. PLoS ONE 2017, 12, e0182415. [Google Scholar] [CrossRef] [PubMed]
- Zychlinsky, A.; Prevost, M.C.; Sansonetti, P.J. Shigella flexneri induces apoptosis in infected macrophages. Nature 1992, 358, 167–169. [Google Scholar] [CrossRef]
- DeLeo, F.R. Modulation of phagocyte apoptosis by bacterial pathogens. Apoptosis 2004, 9, 399–413. [Google Scholar] [CrossRef]
- Giamarellos-Bourboulis, E.J.; Routsi, C.; Plachouras, D.; Markaki, V.; Raftogiannis, M.; Zervakis, D.; Koussoulas, V.; Orfanos, S.; Kotanidou, A.; Armaganidis, A. Early apoptosis of blood monocytes in the septic host: Is it a mechanism of protection in the event of septic shock? Crit. Care 2006, 10, R76. [Google Scholar] [CrossRef]
- Gille, C.; Leiber, A.; Spring, B.; Kempf, V.A.; Loeffler, J.; Poets, C.F.; Orlikowsky, T.W. Diminished phagocytosis-induced cell death (PICD) in neonatal monocytes upon infection with Escherichia coli. Pediatr. Res. 2008, 63, 33–38. [Google Scholar] [CrossRef]
- Gille, C.; Dreschers, S.; Leiber, A.; Lepiorz, F.; Krusch, M.; Grosse-Opphoff, J.; Spring, B.; Haas, M.; Urschitz, M.; Poets, C.F.; et al. The CD95/CD95L pathway is involved in phagocytosis-induced cell death of monocytes and may account for sustained inflammation in neonates. Pediatr. Res. 2013, 73, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Dreschers, S.; Platen, C.; Ludwig, A.; Gille, C.; Köstlin, N.; Orlikowsky, T.W. Metalloproteinases TACE and MMP-9 Differentially Regulate Death Factors on Adult and Neonatal Monocytes After Infection with Escherichia coli. Int. J. Mol. Sci. 2019, 20, 1399. [Google Scholar] [CrossRef]
- Leiber, A.; Graf, B.; Spring, B.; Rudner, J.; Köstlin, N.; Orlikowsky, T.W.; Poets, C.F.; Gille, C. Neonatal monocytes express antiapoptotic pattern of Bcl-2 proteins and show diminished apoptosis upon infection with Escherichia coli. Pediatr. Res. 2014, 76, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Platen, C.; Dreschers, S.; Reiss, K.L.; Wappler, J.; Orlikowsky, T.W. Amphiregulin Regulates Phagocytosis-Induced Cell Death in Monocytes via EGFR and Matrix Metalloproteinases. Mediat. Inflamm. 2018, 2018, 4310419. [Google Scholar] [CrossRef] [PubMed]
- Platen, C.; Dreschers, S.; Wappler, J.; Ludwig, A.; Düsterhöft, S.; Reiss, L.K.; Orlikowsky, T.W. Amphiregulin Regulates Phagocytosis-Induced Cell Death in Monocytes via EGFR and the Bcl-2 Protein Family. Mediat. Inflamm. 2019, 2019, 1603131. [Google Scholar] [CrossRef]
- Dreschers, S.; Gille, C.; Haas, M.; Grosse-Ophoff, J.; Schneider, M.; Leiber, A.; Bühring, H.-J.; Orlikowsky, T.W. Infection–induced Bystander-Apoptosis of Monocytes Is TNF-alpha-mediated. PLoS ONE. 2013, 8, e53589. [Google Scholar] [CrossRef] [PubMed]
- Parameswaran, N.; Patial, S. Tumor necrosis factor-alpha signaling in macrophages. Crit. Rev. Eukaryot. Gene. Expr. 2010, 20, 87–103. [Google Scholar] [CrossRef]
- Humberg, A.; Fortmann, I.; Siller, B.; Kopp, M.V.; Herting, E.; Gopel, W.; Hartel, C.; German Neonatal Network, German Center for Lung Research and Priming Immunity at the Beginning of Life (PRIMAL) Consortium. Preterm birth and sustained inflammation: Consequences for the neonate. Semin. Immunopathol. 2020, 4, 451–468. [Google Scholar] [CrossRef]
- Ito, S.; Tanaka, Y.; Oshino, R.; Okado, S.; Hori, M.; Isobe, K.I. GADD34 suppresses lipopolysaccharide-induced sepsis and tissue injury through the regulation of macrophage activation. Cell Death Dis. 2016, 7, e2219. [Google Scholar] [CrossRef]
- Li, M.; Huang, X.; Zhuo, Q.; Zhang, J.; Ju, X. Clinical significance of miR-129–5p in patients with neonatal sepsis and its regulatory role in the lipopolysaccharide-induced inflammatory response. Bosn. J. Basic Med. Sci. 2020. [Google Scholar] [CrossRef]
- Carbone, F.; Montecucco, F.; Sahebkar, A. Current and emerging treatments for neonatal sepsis. Expert Opin. Pharm. 2020, 21, 549–556. [Google Scholar] [CrossRef]
- Aziz, M.; Jacob, A.; Matsuda, A.; Wu, R.; Zhou, M.; Dong, W.; Yang, W.L.; Wang, P. Pre-treatment of recombinant mouse MFG-E8 downregulates LPS-induced TNF-alpha production in macrophages via STAT3-mediated SOCS3 activation. PLoS ONE 2011, 6, e27685. [Google Scholar] [CrossRef]
- Shen, H.; Lei, Y.; He, X.; Liu, D.; He, Z. Role of lactadherin in intestinal barrier integrity in experimental neonatal necrotizing enterocolitis. J. Cell Biochem. 2019, 2, 19509–19517. [Google Scholar] [CrossRef]
- Hansen, L.W.; Yang, W.L.; Bolognese, A.C.; Jacob, A.; Chen, T.; Prince, J.M.; Nicastro, J.M.; Coppa, G.F.; Wang, P. Treatment with milk fat globule epidermal growth factor-factor 8 (MFG-E8) reduces inflammation and lung injury in neonatal sepsis. Surgery 2017, 162, 349–357. [Google Scholar] [CrossRef]
- Hansen, L.W.; Khader, A.; Yang, W.L.; Jacob, A.; Chen, T.; Nicastro, J.M.; Coppa, G.F.; Prince, J.M.; Wang, P. Deficiency in milk fat globule-epidermal growth factor-factor 8 exacerbates organ injury and mortality in neonatal sepsis. J. Pediatric Surg. 2017, 52, 1520–1527. [Google Scholar] [CrossRef]
- Asaro, J.A.; Khan, Z.; Brewer, M.; Klose, K.; Pesce, C.; Schanler, R.J.; Codipilly, C.N. Relationship Between Milk Fat Globule-Epidermal Growth Factor 8 and Intestinal Cytokines in Infants Born Preterm. J. Pediatr. 2021, 230, 71–75. [Google Scholar] [CrossRef] [PubMed]
- Martinez, F.O.; Sica, A.; Mantovani, A.; Locati, M. Macrophage activation and polarization. Front. Biosci. 2008, 13, 453–461. [Google Scholar] [CrossRef]
- Orlikowsky, T.; Wang, Z.Q.; Dudhane, A.; Horowitz, H.; Conti, B.; Hoffmann, M.K. Two distinct pathways of human macrophage differentiation are mediated by interferon-gamma and interleukin-10. Mmunology 1997, 191, 104–108. [Google Scholar]
- Gille, C.; Spring, B.; Tewes, L.J.; Loffler, J.; Dannecker, G.E.; Hoffmann, M.K.; Eichner, M.; Poets, C.F.; Orlikowsky, T.W. Diminished response to interleukin-10 and reduced antibody-dependent cellular cytotoxicity of cord blood monocyte-derived macrophages. Pediatr. Res. 2006, 60, 152–157. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Dreschers, S.; Ohl, K.; Schulte, N.; Tenbrock, K.; Orlikowsky, T.W. Impaired functional capacity of polarised neonatal macrophages. Sci. Rep. 2020, 10, 624. [Google Scholar] [CrossRef]
- Passlick, B.; Flieger, D.; Ziegler-Heitbrock, H.W. Identification and characterization of a novel monocyte subpopulation in human peripheral blood. Blood 1089, 74, 2527–2534. [Google Scholar] [CrossRef]
- Wong, K.L.; Yeap, W.H.; Yi Tai, J.J.; Ong, S.W.; Dang, T.M.; Wong, S.C. The three human monocyte subsets: Implications for health and disease. Immunol. Res. 2011, 53, 41–57. [Google Scholar] [CrossRef]
- Ozanska, A.; Szymczak, D.; Rybka, J. Pattern of human monocyte subpopulations in health and disease. Scand. J. Immunol. 2020, 92, e12883. [Google Scholar] [CrossRef] [PubMed]
- da Mota, N.V.F.; Brunialti, M.K.C.; Santos, S.S.; Machado, F.R.; Assuncao, M.; Azevedo, L.C.P.; Salomao, R. Immunophenotyping of Monocytes During Human Sepsis Shows Impairment in Antigen Presentation: A Shift Toward Nonclassical Differentiation and Upregulation of FCgammaRi-Receptor. Shock 2018, 50, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Greco, M.; Mazzei, A.; Palumbo, C.; Verri, T.; Lobreglio, G. Flow Cytometric Analysis of Monocytes Polarization and Reprogramming from Inflammatory to Immunosuppressive Phase During Sepsis. EJIFCC 2019, 30, 371–384. [Google Scholar]
- Nguyen, M.; Leuridan, E.; Zhang, T.; De Wit, D.; Willems, F.; Van Damme, P.; Goldman, M.; Goriely, S. Acquisition of adult-like TLR4 and TLR9 responses during the first year of life. PLoS ONE 2010, 5, e10407. [Google Scholar] [CrossRef] [PubMed]
- Damasceno, D.; Teodosio, C.; van den Bossche, V.B.L.; Perez-Andres, M.; Arriba, S.; Munoz-Bellvis, L.; Romero, A.; Blanco, J.F.; Remesal, A.; Puig, N.; et al. Distribution of subsets of blood monocytic cells throughout life. J. Allergy. Clin. Immunol. 2019, 144, 320–323. [Google Scholar] [CrossRef]
- Murphy, F.J.; Reen, D.J. Differential expression of function-related antigens on newborn and adult monocyte subpopulations. Immunology 1996, 89, 587–591. [Google Scholar] [CrossRef]
- Skrzeczynska-Moncznik, J.; Bzowska, M.; Loseke, S.; Grage-Griebenow, E.; Zembala, M.; Pryjma, J. Peripheral blood CD14high CD16+ monocytes are main producers of IL-10. Scand. J. Immunol. 2008, 67, 152–159. [Google Scholar] [CrossRef]
- Brook, B.; Harbeson, D.; Ben-Othman, R.; Viemann, D.; Kollmann, T.R. Newborn susceptibility to infection vs. disease depends on complex in vivo interactions of host and pathogen. Semin. Immunopathol. 2017, 36, 615–625. [Google Scholar] [CrossRef]
- Sugitharini, V.; Prema, A.; Berla Thangam, E. Inflammatory mediators of systemic inflammation in neonatal sepsis. Inflamm. Res. 2013, 62, 1025–1034. [Google Scholar] [CrossRef]
- Ye, Q.; Du, L.Z.; Shao, W.X.; Shang, S.Q. Utility of cytokines to predict neonatal sepsis. Pediatr. Res. 2017, 81, 616–621. [Google Scholar] [CrossRef] [PubMed]
- Khaertynov, K.S.; Boichuk, S.V.; Khaiboullina, S.F.; Anokhin, V.A.; Andreeva, A.A.; Lombardi, V.C.; Satrutdinov, M.A.; Agafonova, E.A.; Rizvanov, A.A. Comparative Assessment of Cytokine Pattern in Early and Late Onset of Neonatal Sepsis. J. Immunol. Res. 2017, 2017, 8601063. [Google Scholar] [CrossRef] [PubMed]
- Leal, Y.A.; Alvarez-Nemegyei, J.; Lavadores-May, A.I.; Giron-Carrillo, J.L.; Cedillo-Rivera, R.; Velazquez, J.R. Cytokine profile as diagnostic and prognostic factor in neonatal sepsis. J. Matern. Fetal. Neonatal. Med. 2019, 32, 2830–2836. [Google Scholar] [CrossRef] [PubMed]
- Ebenebe, C.U.; Hesse, F.; Blohm, M.E.R.; Jung, R.; Kunzmann, S.; Singer, D. Diagnostic accuracy of interleukin-6 for early-onset sepsis in preterm neonates. J. Matern. Fetal. Neonatal. Med. 2021, 34, 253–258. [Google Scholar] [CrossRef]
- Rodrıguez-Gaspar, M.; Santolaria, F.; Lopez, A.J.; Gonzalez-Reimers, E.; Milena, A.; Rodrıguez-Rodrıguez, E.; Gomez-Sirvent, J.L. Prognostic value of cytokines in SIRS general medical patients. Cytokine 2001, 15, 232–236. [Google Scholar] [CrossRef]
- Chen, X.F.; Wu, J.; Zhang, Y.D.; Zhang, C.X.; Chen, X.T.; Sun, J.H.; Chen, T.X.; Sun, J.H. Role of Zc3h12a in enhanced IL-6 production by newborn mononuclear cells in response to lipopolysaccharide. Pediatr. Neonatol. 2018, 59, 288–295. [Google Scholar] [CrossRef]
- Chen, X.F.; Wu, J.; Zhang, Y.D.; Zhang, C.X.; Chen, X.T.; Zhao, W.; Chen, T.X. Role of SOCS3 in enhanced acute-phase protein genes by neonatal macrophages in response to IL-6. J. Microbiol. Immunol. Infect. 2021, 54, 206–212. [Google Scholar] [CrossRef]
- Lingnau, M.; Höflich, C.; Volk, H.-D.; Sabat, R.; Döcke, W.-D. Interleukin-10 enhances the CD14-dependent phagocytosis of bacteria and apoptotic cells by human monocytes. Human Immunol. 2007, 68, 730–738. [Google Scholar] [CrossRef] [PubMed]
- Mazer, M.; Unsinger, J.; Drewry, A.; Walton, A.; Osborne, D.; Blood, T.; Hotchkiss, R.; Remy, K.E. IL-10 Has Differential Effects on the Innate and Adaptive Immune Systems of Septic Patients. J. Immunol. 2019, 203, 2088–2099. [Google Scholar] [CrossRef]
- Wu, H.P.; Chu, C.M.; Kao, K.C.; Huang, S.H.; Chuang, D.Y. High Interleukin-10 Expression in Type 2 T Helper Cells in Septic Patients. Immunol. Investig. 2017, 46, 385–394. [Google Scholar] [CrossRef]
- Fabri, A.; Kandara, K.; Coudereau, R.; Gossez, M.; Abraham, P.; Monard, C.; Cour, M.; Rimmele, T.; Argaud, L.; Monneret, G.; et al. Characterization of Circulating IL-10-Producing Cells in Septic Shock Patients: A Proof of Concept Study. Front. Immunl. 2020, 11, 615009. [Google Scholar] [CrossRef]
- Davidson, D.; Miskolci, V.; Clark, D.C.; Dolmaian, G.; Vancurova, I. Interleukin-10 production after pro-inflammatory stimulation of neutrophils and monocytic cells of the newborn. Neonatology 2007, 92, 127–133. [Google Scholar] [CrossRef]
- Roger, T.; Schneider, A.; Weier, M.; Sweep, F.C.; le Roy, D.; Bernhagen, J.; Calandra, T.; Giannoni, E. High expression levels of macrophage migration inhibitory factor sustain the innate immune responses of neonates. Proc. Natl. Acad. Sci. USA 2016, 113, E997–E1005. [Google Scholar] [CrossRef]
- Lawrence, S.M.; Ruoss, J.L.; Wynn, J.L. IL-17 in neonatal health and disease. Am. J. Reprod. Immunol. 2018, 79, e12800. [Google Scholar] [CrossRef]
- Cherwinski, H.M.; Schumacher, J.H.; Brown, K.D.; Mosmann, T.R. Two types of mouse helper T cell clone. III. Further differences in lymphokine synthesis between Th1 and Th2 clones revealed by RNA hybridization, functionally monospecific bioassays, and monoclonal antibodies. J. Exp. Med. 1987, 166, 1229–1244. [Google Scholar] [CrossRef] [PubMed]
- Maródi, L.; Goda, K.; Palicz, A.; Szabó, G. Cytokine receptor signalling in neonatal macrophages: Defective STAT-1 phosphorylation in response to stimulation with IFN-gamma. Clin. Exp. Immunol. 2001, 26, 456–460. [Google Scholar] [CrossRef] [PubMed]
- Marodi, L. Neonatal innate immunity to infectious agents. Infect. Immun. 2006, 4, 1999–2006. [Google Scholar] [CrossRef]
- Marodi, L. Down-regulation of Th1 responses in human neonates. Clin. Exp. Immunol. 2002, 128, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Sung, N.; Gilman-Sachs, A.; Kwak-Kim, J. T Helper (Th) Cell Profiles in Pregnancy and Recurrent Pregnancy Losses: Th1/Th2/Th9/Th17/Th22/Tfh Cells. Front. Immunol. 2020, 11, 2025. [Google Scholar] [CrossRef] [PubMed]
- Gajewski, T.F.; Fitch, F.W. Anti-proliferative effect of IFN-gamma in immune regulation. I. IFN-gamma inhibits the proliferation of Th2 but not Th1 murine helper T lymphocyte clones. J. Immunol. 1988, 140, 4245–4252. [Google Scholar]
- Fiorentino, D.F.; Zlotnik, A.; Vieira, P.; Mosmann, T.R.; Howard, M.; Moore, K.W.; O’Garra, A. IL-10 acts on the antigen-presenting cell to inhibit cytokine production by Th1 cells. J. Immunol. 1991, 146, 3444–3451. [Google Scholar] [PubMed]
- Coakley, J.D.; Breen, E.P.; Moreno-Olivera, A.; Al-Harbi, A.I.; Melo, A.M.; O’Connell, B.; McManus, R.; Doherty, D.G.; Ryan, T. Dysregulated T helper type 1 (Th1) and Th17 responses in elderly hospitalised patients with infection and sepsis. PLoS ONE 2019, 14, e0224276. [Google Scholar] [CrossRef] [PubMed]
- Manzoli, T.F.; Troster, E.J.; Ferreira-Ferranti, J.; Mirtes-Sales, M. Prolonged suppression of monocytic human leukocyte antigen–DR expression correlates with mortality in pediatric septic patients in a pediatric tertiary Intensive Care Unit. J. Crit. Care 2016, 33, 84–89. [Google Scholar] [CrossRef] [PubMed]
- Boeddha, N.P.; Kerklaan, D.; Dunbar, A.; van Puffelen, E.; Nagtzaam, N.M.A.; Vanhorebeek, I.; Van den Berghe, G.; Hazelzet, J.A.; Joosten, K.F.; Verbruggen, S.C.; et al. HLA-DR Expression on Monocyte Subsets in Critically Ill Children. Pediatr. Infect. Dis. J. 2018, 37, 1034–1040. [Google Scholar] [CrossRef] [PubMed]
- Genel, F.; Atlihan, F.; Ozsu, F.; Ozbek, E. Monocyte HLA-DR expression as predictor of poor outcome in neonates with late onset neonatal sepsis. J Infect. 2010, 60, 224–228. [Google Scholar] [CrossRef]
- Schefold, J.C.; Porz, L.; Uebe, B.; Poehlmann, H.; von Haehling, S.; Jung, A.; Unterwalder, N.; Meisel, C. Diminished HLA-DR expression on monocyte and dendritic cell subsets indicating impairment of cellular immunity in pre-term neonates: A prospective observational analysis. J. Perinat. Med. 2015, 43, 609–618. [Google Scholar] [CrossRef]
- Juskewitch, J.E.; Abraham, R.S.; League, S.C.; Jenkins, S.M.; Smith, C.Y.; Enders, F.T.; Grebe, S.K.; Carey, W.A.; Huskins, W.C. Monocyte HLA-DR expression and neutrophil CD64 expression as biomarkers of infection in critically ill neonates and infants. Pediatr. Res. 2015, 78, 683–690. [Google Scholar] [CrossRef]
- Pradhan, R.; Jain, P.; Paria, A.; Saha, A.; Sahoo, J.; Sen, A.; Mukherjee, S.; Som, T.; Hazra, A.; Warner, N.; et al. Ratio of neutrophilic CD64 and monocytic HLA-DR: A novel parameter in diagnosis and prognostication of neonatal sepsis. Cytom. B Clin Cytom. 2016, 90, 295–302. [Google Scholar] [CrossRef]
- Venet, F.; Pachot, A.; Debard, A.I.; Bohe, J.; Bienvenu, J.; Lepape, A.; Monneret, G. Increased percentage of CD4+CD25+ regulatory T cells during septic shock is due to the decrease of CD4+CD25- lymphocytes. Crit. Care Med. 2004, 32, 2329–2331. [Google Scholar] [CrossRef]
- Nascimento, D.C.; Melo, P.H.; Piñeros, A.R.; Ferreira, R.G.; Colón, D.F.; Donate, P.B.; Castanheira, F.V.; Gozzi, A.; Czaikoski, P.G.; Niedbala, W.; et al. IL-33 contributes to sepsis-induced long-term immunosuppression by expanding the regulatory T cell population. Nat. Commun. 2017, 8, 14919. [Google Scholar] [CrossRef]
- Wolk, K.; Docke, W.; von Baehr, V.; Volk, H.; Sabat, R. Comparison of monocyte functions after LPS- or IL-10-induced reorientation: Importance in clinical immunoparalysis. Pathobiology 1999, 67, 253–256. [Google Scholar] [CrossRef]
- Hikita, N.; Cho, Y.; Tachibana, D.; Hamazaki, T.; Koyama, M.; Tokuhara, D. Cell surface antigens of neonatal monocytes are selectively impaired in basal expression, but hyperresponsive to lipopolysaccharide and zymosan. J. Reprod. Immunol. 2019, 136, 102614. [Google Scholar] [CrossRef]
- Zahran, A.M.; Saad, K.; Abdel-Raheem, Y.F.; Elsayh, K.I.; El-Houfey, A.A.; Aboul-Khair, M.D.; Alblihed, M.A. Characterization of Regulatory T Cells in Preterm and Term Infants. Arch. Immunol. Ther. Exp. 2019, 67, 49–54. [Google Scholar] [CrossRef]
- Lee, J.G.; Jaeger, K.E.; Seki, Y.; Lim, Y.W.; Cunha, C.; Vuchkovska, A.; Nelson, A.J.; Nikolai, A.; Kim, D.; Nishimura, M.; et al. Human CD36hi monocytes induce Foxp3+ CD25+ T cells with regulatory functions from CD4 and CD8 subsets. Immunology 2021, 163, 293–309. [Google Scholar] [CrossRef]
- Pang, Y.; Du, X.; Xu, X.; Wang, M.; Li, Z. Monocyte activation and inflammation can exacerbate Treg/Th17 imbalance in infants with neonatal necrotizing enterocolitis. Int. Immunopharmacol. 2018, 59, 354–360. [Google Scholar] [CrossRef] [PubMed]
- Heinemann, A.S.; Pirr, S.; Fehlhaber, B.; Mellinger, L.; Burgmann, J.; Busse, M.; Ginzel, M.; Friesenhagen, J.; von Kockritz-Blickwede, M.; Ulas, T.; et al. In neonates S100A8/S100A9 alarmins prevent the expansion of a specific inflammatory monocyte population promoting septic shock. FASEB J. 2017, 31, 1153–1164. [Google Scholar] [CrossRef] [PubMed]
- Ehrchen, J.M.; Sunderkötter, C.; Foell, D.; Vogl, T.; Roth, J. The endogenous Toll-like receptor 4 agonist S100A8/S100A9 (calprotectin) as innate amplifier of infection, autoimmunity, and cancer. J. Leukoc. Biol. 2009, 86, 557–566. [Google Scholar] [CrossRef] [PubMed]
- Pruenster, M.; Vogl, T.; Roth, J.; Sperandio, M. S100A8/A9: From basic science to clinical application. Pharmacol. Ther. 2016, 167, 120–131. [Google Scholar] [CrossRef] [PubMed]
- Austermann, J.; Friesenhagen, J.; Fassl, S.K.; Petersen, B.; Ortkras, T.; Burgmann, J.; Barczyk-Kahlert, M.; Faist, E.; Zedler, S.; Pirr, S.; et al. Alarmins MRP8 and MRP14 induce stress tolerance in phagocytes under sterile inflammatory conditions. Cell Rep. 2014, 9, 2112–2123. [Google Scholar] [CrossRef] [PubMed]
- Vogl, T.; Tenbrock, K.; Ludwig, S.; Leukert, N.; Ehrhardt, C.; van Zoelen, M.A.; Nacken, W.; Foell, D.; van der Poll, C.; Sorg, C.; et al. Mrp8 and Mrp14 are endogenous activators of Toll-like receptor 4, promoting lethal, endotoxin-induced shock. N. Nat. Med. 2007, 13, 1042–1049. [Google Scholar] [CrossRef]
- Ulas, T.; Pirr, S.; Fehlhaber, B.; Bickes, M.S.; Loof, T.G.; Vogl, T.; Mellinger, L.; Heinemann, A.S.; Burgmann, J.; Schöning, J.; et al. S100-alarmin-induced innate immune programming protects newborn infants from sepsis. Nat. Immunol. 2017, 18, 622–632. [Google Scholar] [CrossRef]
- Pirr, S.; Richter, M.; Fehlhaber, B.; Pagel, J.; Hartel, C.; Roth, J.; Vogl, T.; Viemann, D. High Amounts of S100-Alarmins Confer Antimicrobial Activity on Human Breast Milk Targeting Pathogens Relevant in Neonatal Sepsis. Front. Immunol. 2017, 8, 1822. [Google Scholar] [CrossRef]
- Van Lent, P.L.; Grevers, L.C.; Blom, A.B.; Arntz, O.J.; van de Loo, F.A.; van den Kraan, P.; Abdollahi-Roodsaz, S.; Srikrishna, G.; Freeze, H.; Sloetjes, A.; et al. Stimulation of chondrocyte-mediated cartilage destruction by S100A8 in experimental murine arthritis. Arthritis Rheum. 2008, 58, 3776–3787. [Google Scholar] [CrossRef]
- Golubinskaya, V.; Puttonen, H.; Fyhr, I.M.; Rydbeck, H.; Hellstrom, A.; Jacobsson, B.; Nilsson, H.; Mallard, C.; Savman, K. Expression of S100A Alarmins in Cord Blood Monocytes Is Highly Associated with Chorioamnionitis and Fetal Inflammation in Preterm Infants. Front. Immunol. 2020, 11, 1194. [Google Scholar] [CrossRef]
- Viemann, D. S100-Alarmins Are Essential Pilots of Postnatal Innate Immune Adaptation. Front. Immunol. 2020, 11, 688. [Google Scholar] [CrossRef] [PubMed]
- Willers, M.; Ulas, T.; Vollger, L.; Vogl, T.; Heinemann, A.S.; Pirr, S.; Pagel, J.; Fehlhaber, B.; Halle, O.; Schoning, J.; et al. S100A8 and S100A9 Are Important for Postnatal Development of Gut Microbiota and Immune System in Mice and Infants. Gastroenterology 2020, 159, 2130–2145. [Google Scholar] [CrossRef] [PubMed]
- Dreschers, S.; Ohl, K.; Lehrke, M.; Möllmann, J.; Denecke, B.; Costa, I.; Vogl, T.; Viemann, D.; Roth, J.; Orlikowsky, T.; et al. Impaired cellular energy metabolism in cord blood macrophages contributes to abortive response toward inflammatory threats. Nat. Commun. 2019, 10, 1685. [Google Scholar] [CrossRef] [PubMed]
- Diskin, C.; Palsson-McDermott, E.M. Metabolic Modulation in Macrophage Effector Function. Front. Immunol. 2018, 9, 270. [Google Scholar] [CrossRef]
- Freemerman, A.J.; Johnson, A.R.; Sacks, G.N.; Milner, J.J.; Kirk, E.L.; Troester, M.A.; Macintyre, A.N.; Goraksha-Hicks, P.; Rathmell, J.C.; Makowski, L. Metabolic reprogramming of macrophages: Glucose transporter 1 (GLUT1)-mediated glucose metabolism drives a proinflammatory phenotype. J. Biol. Chem. 2014, 289, 7884–7896. [Google Scholar] [CrossRef]
- Millet, P.; Vachharajani, V.; McPhail, L.; Yoza, B.; McCall, C.E. GAPDH Binding to TNF-alpha mRNA Contributes to Posttranscriptional Repression in Monocytes: A Novel Mechanism of Communication between Inflammation and Metabolism. J. Immunol. 2016, 196, 6. [Google Scholar] [CrossRef]
- Warburg, O.; Wind, F.; Negelein, E. The Metabolism of Tumors in the Body. J. Gen. Physiol. 1927, 8, 519–530. [Google Scholar] [CrossRef]
- Huang, S.C.; Smith, A.M.; Everts, B.; Colonna, M.; Pearce, E.L.; Schilling, J.D.; Pearce, E.J. Metabolic Reprogramming Mediated by the mTORC2-IRF4 Signaling Axis Is Essential for Macrophage Alternative Activation. Immunity 2016, 45, 817–830. [Google Scholar] [CrossRef]
- Kan, B.; Michalski, C.; Fu, H.; Au, H.H.T.; Lee, K.; Marchant, E.A.; Cheng, M.F.; Anderson-Baucum, E.; Aharoni-Simon, M.; Tilley, P.; et al. Cellular metabolism constrains innate immune responses in early human ontogeny. Nat. Commun. 2018, 16, 4822. [Google Scholar] [CrossRef]
- Maoldomhnaigh, C.Ó.; Cox, D.J.; Phelan, J.J.; Malone, F.D.; Keane, J.; Basdeo, S.A. The Warburg Effect Occurs Rapidly in Stimulated Human Adult but Not Umbilical Cord Blood Derived Macrophages. Front. Immunol. 2021, 12, 657261. [Google Scholar] [CrossRef]
- Dreschers, S.; Ohl, K.; Mollmann, J.; Tenbrock, K.; Orlikowsky, T.W. Leucine Reconstitutes Phagocytosis-Induced Cell Death in E. coli-Infected Neonatal Monocytes-Effects on Energy Metabolism and mTOR Signaling. Int. J. Mol. Sci. 2021, 22, 4271. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.C.; Scicluna, B.P.; Arts, R.J.W.; Gresnigt, M.S.; Lachmandas, E.; Giamarellos-Bourboulis, E.J.; Kox, M.; Manjeri, G.R.; Wagenaars, J.A.L.; Cremer, O.L.; et al. Broad defects in the energy metabolism of leukocytes underlie immunoparalysis in se. Nat. Immunol. 2016, 17, 406–413. [Google Scholar] [CrossRef] [PubMed]
- Harbeson, D.; Francis, F.; Bao, W.; Amenyogbe, N.A.; Kollmann, T. Energy Demands of Early Life Drive a Disease Tolerant Phenotype and Dictate Outcome in Neonatal Bacterial Sepsis. Front. Immunol. 2018, 9, 1918. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.L.; Dickinson, P.; Forster, T.; Craigon, M.; Ross, A.; Khondoker, M.R.; France, R.; Ivens, A.; Lynn, D.J.; Orme, J.; et al. Identification of a human neonatal immune-metabolic network associated with bacterial infection. Nat. Commun. 2014, 14, 4649. [Google Scholar] [CrossRef]
- Conti, M.G.; Angelidou, A.; Diray-Arce, J.; Smolen, K.K.; Lasky-Su, J.; De Curtis, M.; Levy, O. Immunometabolic approaches to prevent, detect, and treat neonatal sepsis. Pediatr. Res. 2020, 87, 399–405. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Doughty, C.; Oppermann, L.; Hartmann, N.-U.; Dreschers, S.; Gille, C.; Orlikowsky, T. Monocytes in Neonatal Bacterial Sepsis: Think Tank or Workhorse? BioChem 2022, 2, 27-42. https://doi.org/10.3390/biochem2010003
Doughty C, Oppermann L, Hartmann N-U, Dreschers S, Gille C, Orlikowsky T. Monocytes in Neonatal Bacterial Sepsis: Think Tank or Workhorse? BioChem. 2022; 2(1):27-42. https://doi.org/10.3390/biochem2010003
Chicago/Turabian StyleDoughty, Caitlin, Louise Oppermann, Niels-Ulrik Hartmann, Stephan Dreschers, Christian Gille, and Thorsten Orlikowsky. 2022. "Monocytes in Neonatal Bacterial Sepsis: Think Tank or Workhorse?" BioChem 2, no. 1: 27-42. https://doi.org/10.3390/biochem2010003
APA StyleDoughty, C., Oppermann, L., Hartmann, N.-U., Dreschers, S., Gille, C., & Orlikowsky, T. (2022). Monocytes in Neonatal Bacterial Sepsis: Think Tank or Workhorse? BioChem, 2(1), 27-42. https://doi.org/10.3390/biochem2010003