
Urban Environmental Predictors of Group Size in Cliff Swallows (Petrochelidon pyrrhonota): A Test Using Community-Science Data

Simple Summary
Abstract
1. Introduction
2. Methods
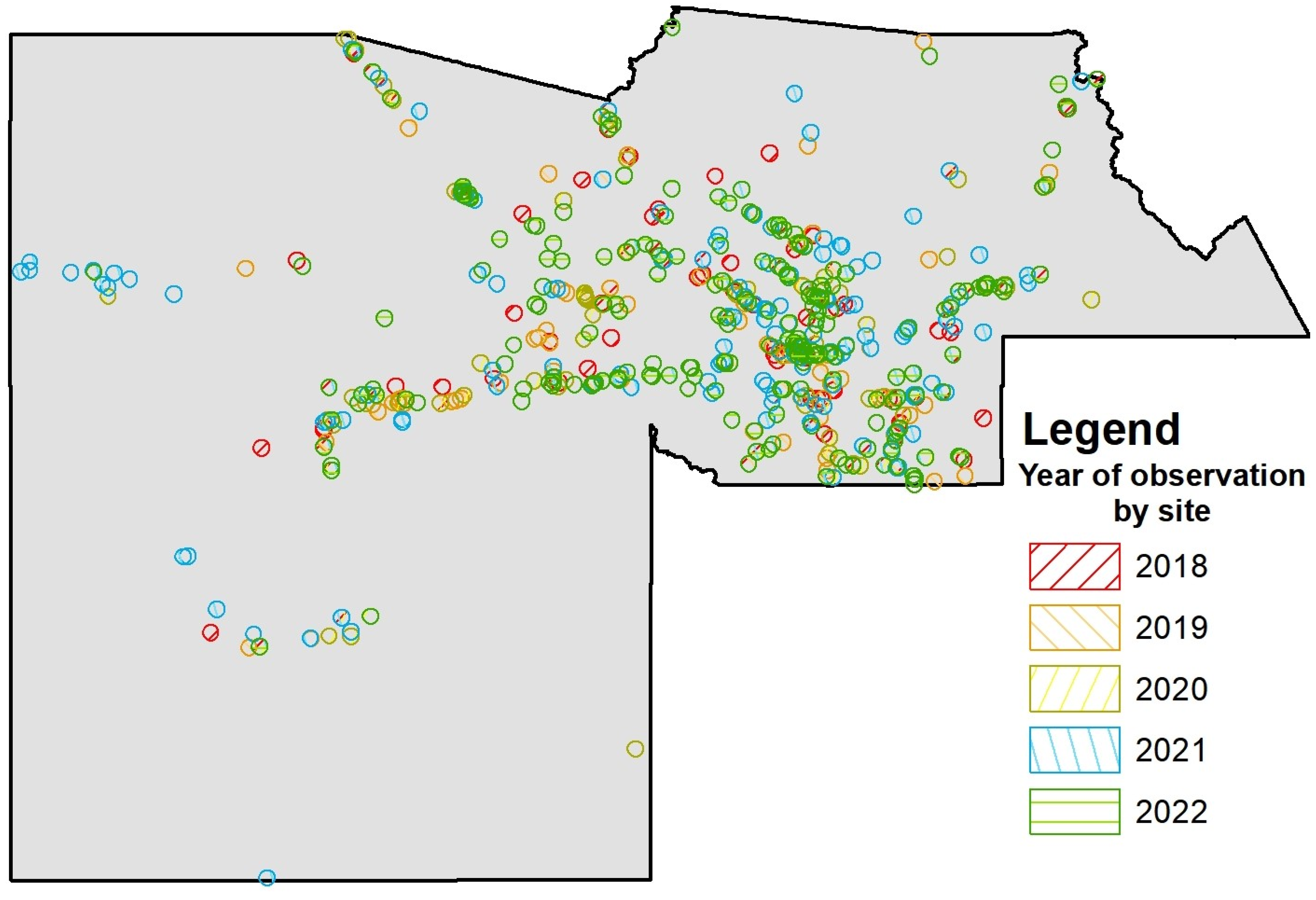
2.1. Acquisition of Swallow Group-Size Dataset
2.2. Urban and Environmental Parameters
2.3. Data Reduction and Variable Selection
2.4. Statistics
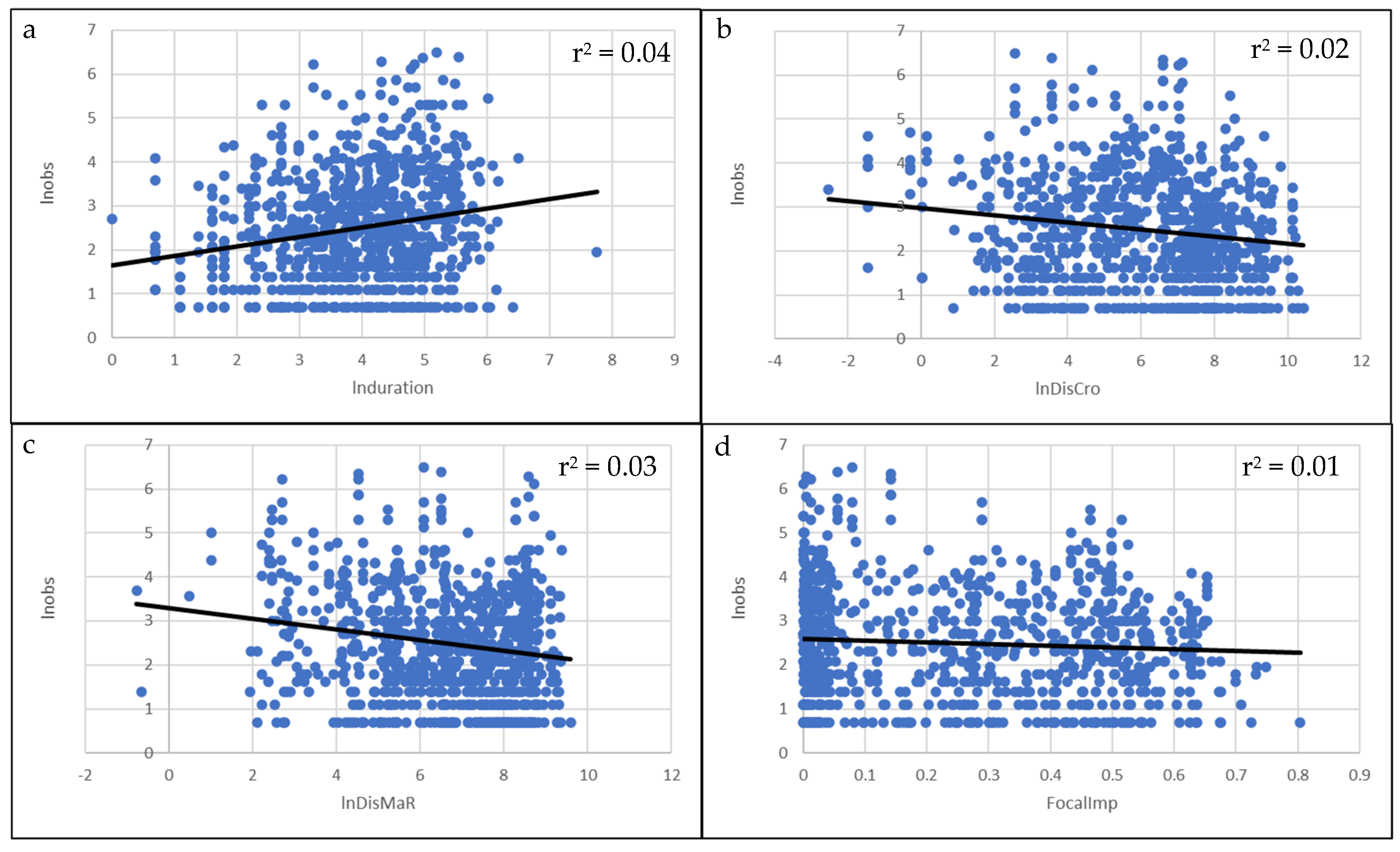
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- United Nations Department of Economic and Social Affairs Population Division. World Urbanization Prospects: The 2018 Revision (ST/ESA/SER.A/420); United Nations: New York, NY, USA, 2019; Available online: https://population.un.org/wup/assets/WUP2018-Report.pdf (accessed on 16 February 2025).
- Collins, M.K.; Magle, S.B.; Gallo, T. Global trends in urban wildlife ecology and conservation. Biol. Cons. 2021, 261, 109236. [Google Scholar] [CrossRef]
- Marzluff, J.M.; Bowman, R.; Donnelly, R. (Eds.) Avian Ecology and Conservation in an Urbanizing World; Springer Science and Business Media: Berlin/Heidelberg, Germany, 2012. [Google Scholar] [CrossRef]
- Hensley, C.B.; Trisos, C.H.; Warren, P.S.; MacFarland, J.; Blumenshine, S.; Reece, J.; Katti, M. Effects of urbanization on native bird species in three southwestern US cities. Front. Ecol. Evol. 2019, 7, 00071. [Google Scholar] [CrossRef]
- Marín-Gómez, O.H.; MacGregor-Fors, I. A global synthesis of the impacts of urbanization on bird dawn choruses. Ibis 2021, 163, 1133–1154. [Google Scholar] [CrossRef]
- Tsurim, I.; Kotler, B.P.; Gilad, A.; Elazary, S.; Abramsky, Z. Foraging behavior of an urban bird species: Molt gaps, distance to shelter, and predation risk. Ecology 2010, 91, 233–241. [Google Scholar] [CrossRef]
- Reynolds, S.J.; Galbraith, J.A.; Smith, J.A.; Jones, D.N. Garden bird feeding: Insights and prospects from a north-south comparison of this global urban phenomenon. Front. Ecol. Evol. 2017, 5, 24. [Google Scholar] [CrossRef]
- Capilla-Lasheras, P.; Thompson, M.J.; Sánchez-Tójar, A.; Haddou, Y.; Branston, C.J.; Réale, D.; Charmentier, A.; Dominoni, D.M. A global meta-analysis reveals higher variation in breeding phenology in urban birds than in their non-urban neighbours. Ecol. Lett. 2022, 25, 2552–2570. [Google Scholar] [CrossRef]
- Van Doren, B.M.; Horton, K.G.; Dokter, A.M.; Klinck, H.; Elbin, S.B.; Farnsworth, A. High-intensity urban light installation dramatically alters nocturnal bird migration. Proc. Natl. Acad. Sci. USA 2017, 114, 11175–11180. [Google Scholar] [CrossRef]
- Jokimäki, J.; Suhonen, J.; Jokimäki-Kaisanlahti, M.L.; Carbó-Ramírez, P. Effects of urbanization on breeding birds in European towns: Impacts of species traits. Urban Ecosyst. 2016, 19, 1565–1577. [Google Scholar] [CrossRef]
- Zanette, L.R.S.; Martins, R.P.; Ribeiro, S.P. Effects of urbanization on Neotropical wasp and bee assemblages in a Brazilian metropolis. Landsc. Urban Plann. 2005, 71, 105–121. [Google Scholar] [CrossRef]
- Hernandez, J.L.; Frankie, G.W.; Thorp, R.W. Ecology of urban bees: A review of current knowledge and directions for future study. Cities Environ. 2009, 2, 3. [Google Scholar] [CrossRef]
- Samuelson, A.E.; Gill, R.J.; Brown, M.J.; Leadbeater, E. Lower bumblebee colony reproductive success in agricultural compared with urban environments. Proc. R. Soc. B 2018, 285, 20180807. [Google Scholar] [CrossRef] [PubMed]
- Coulson, J. Colonial breeding in seabirds. In Biology of Marine Birds; Schreiber, E.A., Burger, J., Eds.; CRC Press: Boca Raton, FL, USA, 2002; pp. 87–113. [Google Scholar]
- Jovani, R.; Lascelles, B.; Garamszegi, L.Z.; Mavor, R.; Thaxter, C.B.; Oro, D. Colony size and foraging range in seabirds. Oikos 2015, 125, 968–974. [Google Scholar] [CrossRef]
- Krebs, J.R. Colonial nesting and social feeding as strategies for exploiting food resources in the great blue heron (Ardea herodias). Behaviour 1974, 51, 99–134. [Google Scholar] [CrossRef]
- Brown, C.R.; Brown, M.B.; Hannebaum, S.L.; Wagnon, G.S.; Pletcher, O.M.; Page, C.E.; West, A.C.; O’Brien, V.A. Social foraging and the associated benefits of group-living in cliff swallows decrease over 40 years. Ecol. Monogr. 2024, 94, e1602. [Google Scholar] [CrossRef]
- Melle, M.E.; Mbole, V.; Nasako, P.N.; Esong, E.L.; Bekarikoh, E.N.; Agborta, O.S. The destruction of feeding and nesting behaviors of weaver-birds in urban crop-farms, Buea Municipality, Southwest Region, Cameroon. Int. J. Agricult. Sci. 2020, 1, 1–15. [Google Scholar]
- Picman, J.; Pribil, S.; Isabelle, A. Antipredation value of colonial nesting in yellow-headed blackbirds. Auk 2002, 119, 461–472. [Google Scholar] [CrossRef]
- Negro, J.J.; Prenda, J.; Ferrero, J.J.; Rodriguez, A.; Reig-Ferrer, A. A timeline for the urbanization of wild birds: The case of the lesser kestrel. Quat. Sci. Rev. 2020, 249, 106638. [Google Scholar] [CrossRef]
- Solaro, C.; Sarasola, J.H. Breeding performance is explained for coloniality and phenology but not for urbanization in a generalist raptor bird. Urban Ecosyst. 2023, 26, 743–753. [Google Scholar] [CrossRef]
- Ma, Q.; He, C.; Wu, J. Behind the rapid expansion of urban impervious surfaces in China: Major influencing factors revealed by a hierarchical multiscale analysis. Land Use Policy 2016, 59, 434–445. [Google Scholar] [CrossRef]
- Narango, D.L.; Tallamy, D.W.; Marra, P.P. Native plants improve breeding and foraging habitat for an insectivorous bird. Biol. Cons. 2017, 213, 42–50. [Google Scholar] [CrossRef]
- Luck, G.W.; Smallbone, L.T.; Sheffield, K.J. Environmental and socio-economic factors related to urban bird communities. Austral Ecol. 2013, 38, 111–120. [Google Scholar] [CrossRef]
- Banville, M.J.; Bateman, H.L.; Earl, S.R.; Warren, P.S. Decadal declines in bird abundance and diversity in urban riparian zones. Landsc. Urban Plann. 2017, 159, 48–61. [Google Scholar] [CrossRef]
- Souza, F.L.; Valente-Neto, F.; Severo-Neto, F.; Bueno, B.; Ochoa-Quintero, J.M.; Laps, R.R.; Bolzan, F.; de Oliveira Roque, F. Impervious surface and heterogeneity are opposite drivers to maintain bird richness in a Cerrado city. Landsc. Urban Plann. 2019, 192, 103643. [Google Scholar] [CrossRef]
- Stewart, I.D. A systematic review and scientific critique of methodology in modern urban heat island literature. Int. J. Climatol. 2011, 31, 200–217. [Google Scholar] [CrossRef]
- McDonald, R.I.; Green, P.; Balk, D.; Fekete, B.M.; Revenga, C.; Todd, M.; Montgomery, M. Urban growth, climate change, and freshwater availability. Proc. Natl. Acad. Sci. USA 2011, 108, 6312–6317. [Google Scholar] [CrossRef]
- Fenoglio, M.S.; Calviño, A.; González, E.; Salvo, A.; Videla, M. Urbanisation drivers and underlying mechanisms of terrestrial insect diversity loss in cities. Ecol. Entomol. 2021, 46, 757–771. [Google Scholar] [CrossRef]
- Brown, C.R.; Stutchbury, B.J.; Walsh, P.D. Choice of colony size in birds. Trends Ecol. Evol. 1990, 5, 398–403. [Google Scholar] [CrossRef]
- Evans, K.L.; Chamberlain, D.E.; Hatchwell, B.J.; Gregory, R.D.; Gaston, K.J. What makes an urban bird? Glob. Change Biol. 2011, 17, 32–44. [Google Scholar] [CrossRef]
- Emlen, J.T. Social behavior in nesting cliff swallows. Condor 1952, 54, 177–199. [Google Scholar] [CrossRef]
- Brown, C.R.; Brown, M.B. Coloniality in the Cliff Swallow: The Effect of Group Size on Social Behavior; University of Chicago Press: Chicago, IL, USA, 1996. [Google Scholar]
- Hannebaum, S.L.; Wagnon, G.S.; Brown, C.R. Variation in neophobia among cliff swallows at different colonies. PLoS ONE 2019, 14, e0226886. [Google Scholar] [CrossRef]
- Brown, C.R. Cliff swallow colonies as information centers. Science 1986, 234, 83–85. [Google Scholar] [CrossRef] [PubMed]
- Brown, C.R. How cliff swallows choose where to live. Phi Kappa Phi Forum 2010, 90, 4–7. [Google Scholar]
- Brown, C.R.; Brown, M.B.; Pyle, P.; Patten, M.A. Cliff swallow (Petrochelidon pyrrhonota), version 1.0. In Birds of the World; Rodewald, P.G., Ed.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar] [CrossRef]
- Gorenzel, W.P.; Salmon, T.P. The cliff swallow—Biology and control. Proc. 10th Vert. Pest Conf. 1982, 17, 179–185. [Google Scholar]
- Conklin, J.S.; Delwiche, M.J.; Gorenzel, W.P.; Coates, R.W. Deterring cliff-swallow nesting on highway structures using bioacoustics and surface modifications. Human-Wildl. Confl. 2009, 3, 93–102. [Google Scholar] [CrossRef]
- Delwiche, M.J.; Coates, R.W.; Gorenzel, W.P.; Salmon, T.P. Improved methods for deterring cliff swallow nesting on highway structures. Human-Wildl. Interact. 2010, 4, 293–303. [Google Scholar] [CrossRef]
- U.S. Census Bureau. Metropolitan and Micropolitan Statistical Areas Population Totals and Components of Change: 2020–2021. 2023. Available online: https://www.census.gov/data/tables/time-series/demo/popest/2020s-total-metro-and-micro-statistical-areas.html (accessed on 16 February 2025).
- Johnston, A.; Hochachka, W.M.; Strimas-Mackey, M.E.; Ruiz Gutierrez, V.; Robinson, O.J.; Miller, E.T.; Auer, T.; Kelling, S.T.; Fink, D. Analytical guidelines to increase the value of community science data: An example using eBird data to estimate species distributions. Divers. Distrib. 2021, 27, 1265–1277. [Google Scholar] [CrossRef]
- Sullivan, B.L.; Aycrigg, J.L.; Barry, J.H.; Bonney, R.E.; Bruns, N.; Cooper, C.B.; Damoulas, T.; Dhondt, A.A.; Dietterich, T.; Farnsworth, A.; et al. The eBird enterprise: An integrated approach to development and application of citizen science. Biol. Cons. 2014, 169, 31–40. [Google Scholar] [CrossRef]
- McNair, D.B. Range expansion and population trends of cliff swallows in northeastern South Carolina and southeastern North Carolina, especially at colony sites along the Great Pee Dee River corridor. Southeast. Nat. 2024, 23, 13–28. [Google Scholar] [CrossRef]
- Robinson, O.J.; Ruiz-Gutierrez, V.; Reynolds, M.D.; Golet, G.H.; Strimas-Mackey, M.; Fink, D. Integrating citizen science data with expert surveys increases accuracy and spatial extent of species distribution models. Divers. Distribut. 2020, 26, 976–986. [Google Scholar] [CrossRef]
- Torres-Cristiani, L.; Machkour-M’Rabet, S.; Calmé, S.; Weissenberger, H.; Escalona-Segura, G. Assessment of the American flamingo distribution, trends, and important breeding areas. PLoS ONE 2020, 15, e0244117. [Google Scholar] [CrossRef]
- Adde, A.; Amat, C.C.I.; Mazerolle, M.J.; Darveau, M.; Cumming, S.G.; O’Hara, R.B. Integrated modeling of waterfowl distribution in western Canada using aerial survey and citizen science (eBird) data. Ecosphere 2021, 12, e03790. [Google Scholar] [CrossRef]
- Brown, C.R. Enhanced foraging efficiency through information centers: A benefit of coloniality in cliff swallows. Ecology 1988, 69, 602–613. [Google Scholar] [CrossRef]
- Brown, C.R.; Brown, M.B.; Roche, E.A. Spatial and temporal unpredictability of colony size in cliff swallows across 30 years. Ecol. Monogr. 2013, 83, 511–530. [Google Scholar] [CrossRef]
- ArcMap. Regression Analysis Basics-ArcMap. Available online: https://desktop.arcgis.com/en/arcmap/latest/tools/spatial-statistics-toolbox/regression-analysis-basics.htm (accessed on 16 February 2025).
- Schell, C.J.; Dyson, K.; Fuentes, T.L.; Des Roches, S.; Harris, N.C.; Miller, D.S.; Woelfle-Erskine, C.A.; Lambert, M.R. The ecological and evolutionary consequences of systemic racism in urban environments. Science 2020, 369, eaay4497. [Google Scholar] [CrossRef]
- Brown, C.R.; Sas, C.M.; Brown, M.B. Colony choice in cliff swallows: Effects of heterogeneity in foraging habitat. Auk 2002, 119, 446–460. [Google Scholar] [CrossRef]
- Bozdogan, H. Model selection and Akaike’s information criterion (AIC): The general theory and its analytical extensions. Psychometrika 1987, 52, 345–370. [Google Scholar] [CrossRef]
- Chace, J.F.; Walsh, J.J. Urban effects on native avifauna: A review. Landsc. Urban Plann. 2006, 74, 46–69. [Google Scholar] [CrossRef]
- Belton, L.E.; Cameron, E.Z.; Dalerum, F. Social networks of spotted hyaenas in areas of contrasting human activity and infrastructure. Anim. Behav. 2018, 135, 13–23. [Google Scholar] [CrossRef]
- Maurer, A.S.; Cove, M.V.; Siegal, O.M.; Lashley, M.A. Urbanization affects the behavior of a predator-free ungulate in protected lands. Landsc. Urban Plann. 2022, 222, 104391. [Google Scholar] [CrossRef]
- Brown, C.R.; Hannebaum, S.L. Birds of a feather flock together: Extent of long-term consistency of colony-size choice in cliff swallows. Front. Ecol. Evol. 2022, 10, 860407. [Google Scholar] [CrossRef]
- Alam, A. Arizona Farmers Look for Ways to Save Water; Cronkite News—Arizona PBS. Available online: https://cronkitenews.azpbs.org/2018/02/09/arizona-farmers-look-for-ways-to-save-water/ (accessed on 16 February 2025).
- Larson, E.K.; Perrings, C. The value of water-related amenities in an arid city: The case of the Phoenix metropolitan area. Landsc. Urban Plann. 2013, 109, 45–55. [Google Scholar] [CrossRef]
- Morabito, M.; Crisci, A.; Guerri, G.; Messeri, A.; Congedo, L.; Munafò, M. Surface urban heat islands in Italian metropolitan cities: Tree cover and impervious surface influences. Sci. Total Environ. 2021, 751, 142334. [Google Scholar] [CrossRef] [PubMed]
- Sekertekin, A.; Zadbagher, E. Simulation of future land surface temperature distribution and evaluating surface urban heat island based on impervious surface area. Ecol. Ind. 2021, 122, 107230. [Google Scholar] [CrossRef]
- Arnold, C.L., Jr.; Gibbons, C.J. Impervious surface coverage: The emergence of a key environmental indicator. J. Am. Plann. Assoc. 1996, 62, 243–258. [Google Scholar] [CrossRef]
- Chithra, S.V.; Nair, M.H.; Amarnath, A.; Anjana, N.S. Impacts of impervious surfaces on the environment. Int. J. Eng. Sci. Invent. 2015, 4, 27–31. [Google Scholar]
- Hochachka, W.M.; Alonso, H.; Gutiérrez-Expósito, C.; Miller, E.; Johnston, A. Regional variation in the impacts of the COVID-19 pandemic on the quantity and quality of data collected by the project eBird. Biol. Cons. 2021, 254, 108974. [Google Scholar] [CrossRef]
- Hochachka, W.M.; Fink, D.; Hutchinson, R.A.; Sheldon, D.; Wong, W.K.; Kelling, S. Data-intensive science applied to broad-scale citizen science. Trends Ecol. Evol. 2012, 27, 130–137. [Google Scholar] [CrossRef]
- Homer, C.; Dewitz, J.; Jin, S.; Xian, G.; Costello, C.; Danielson, P.; Gass, L.; Funk, M.; Wickham, J.; Stehman, S.; et al. Conterminous United States land cover change patterns 2001–2016 from the 2016 national land cover database. ISPRS J. Photogr. Remote Sens. 2020, 162, 184–199. [Google Scholar] [CrossRef]
- Jin, S.; Dewitz, J.; Li, C.; Sorenson, D.; Zhu, Z.; Shogib, M.R.I.; Danielson, P.; Granneman, B.; Costello, C.; Case, A.; et al. National Land Cover Database 2019: A Comprehensive Strategy for Creating the 1986–2019 Forest Disturbance Product. J. Remote Sens. 2023, 3, 0021. [Google Scholar] [CrossRef]
- Wickham, J.; Stehman, S.V.; Sorenson, D.G.; Gass, L.; Dewitz, J.A. Thematic accuracy assessment of the NLCD 2016 land cover for the conterminous United States. Remote Sens. Environ. 2021, 257, 112357. [Google Scholar] [CrossRef]
- Poulsen, M.N.; Hulland, K.R.S.; Gulas, C.A.; Pham, H.; Dalgish, S.L.; Wilkinson, R.K.; Winch, P.J. Growing an urban oasis: A qualitative study of the perceived benefits of community gardening in Baltimore, Maryland. Cult. Agric. Food Environ. 2014, 36, 69–82. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Source | File Type | Spatial Resolution | When Data Were Collected | Associated Variables |
|---|---|---|---|---|
| PRISM Climate Data (Oregon State) | Raster | 4 km | Continuous | Temperature, precipitation |
| Census TIGER/Line Geodatabase | Vector | - | 2022 | Distance to primary/secondary road |
| Census Demographic Data | Vector | - | 2020 | Population density, minority percentage |
| OpenStreetMap Water Data | Raster | 3 s | 2021 | Distance to major river/canal/large lake |
| National Land Cover Database | Raster | 30 m | 2019 | Distance to open developed/low developed/medium developed/high developed/barren/forest/grassland/pasture/crops/woody wetlands/emergent herbaceous wetlands |
| Impervious Surface Data | Raster | 30 m | 2019 | Percent impervious surface |
| Variable | Coefficient | t-Statistic | p-Value | Adjusted R2 | AICc |
|---|---|---|---|---|---|
| lnduration | 0.22 | 6.25 | 0.00 | 0.10 | 2888.98 |
| Temp | −0.02 | −1.86 | 0.06 | ||
| lnDisMaR | −0.12 | −5.46 | 0.00 | ||
| PPT | −0.00 | −0.76 | 0.45 | ||
| lnDisCro | −0.10 | −5.99 | 0.00 | ||
| percmin | 0.20 | 1.08 | 0.28 | ||
| FocalForest | −1.50 | −1.46 | 0.14 | ||
| FocalImp | −0.32 | −1.77 | 0.08 |
| Variable | Coefficient | t-Statistic | p-Value | Adjusted R2 | AICc |
|---|---|---|---|---|---|
| lnduration | 0.21 | 6.13 | 0.00 | 0.10 | 2885.40 |
| lnDisMaR | −0.12 | −5.65 | 0.00 | ||
| lnDisCro | −0.11 | −6.32 | 0.00 | ||
| FocalImp | −0.32 | −1.79 | 0.07 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rueda, C.; McGraw, K.J. Urban Environmental Predictors of Group Size in Cliff Swallows (Petrochelidon pyrrhonota): A Test Using Community-Science Data. Birds 2025, 6, 17. https://doi.org/10.3390/birds6020017
Rueda C, McGraw KJ. Urban Environmental Predictors of Group Size in Cliff Swallows (Petrochelidon pyrrhonota): A Test Using Community-Science Data. Birds. 2025; 6(2):17. https://doi.org/10.3390/birds6020017
Chicago/Turabian StyleRueda, Cassie, and Kevin J. McGraw. 2025. "Urban Environmental Predictors of Group Size in Cliff Swallows (Petrochelidon pyrrhonota): A Test Using Community-Science Data" Birds 6, no. 2: 17. https://doi.org/10.3390/birds6020017
APA StyleRueda, C., & McGraw, K. J. (2025). Urban Environmental Predictors of Group Size in Cliff Swallows (Petrochelidon pyrrhonota): A Test Using Community-Science Data. Birds, 6(2), 17. https://doi.org/10.3390/birds6020017