1. Introduction
Environmental enrichment is a key component of professionally managed zoo care programs, developed to encourage species-typical behaviors, reduce stress, and mitigate abnormal repetitive behaviors [
1,
2]. Effective enrichment strategies include opportunities for problem-solving, exploration, social interaction, and species-appropriate feeding, all of which support behavioral expression and improve animal wellbeing [
3,
4]. Among these approaches, olfactory enrichment is particularly impactful, as many animal species rely heavily on scent cues for navigation, communication, and environmental engagement [
5]. Scent-based stimuli can promote investigatory and exploratory behaviors and may also affect social interactions. However, their effectiveness varies across species and contexts [
6,
7]. Despite these known benefits, enrichment programs often yield inconsistent results due to species-specific behavioral needs, challenges in monitoring behavioral outcomes, and limitations in staff time or resources [
1,
8]. These outcomes highlight the need for enrichment strategies that are biologically meaningful and grounded in an understanding of each species’ natural history and behavioral ecology.
One such underrepresented species is the binturong, which has received limited attention in enrichment research despite its unique sensory and behavioral profiles. The binturong (
Arctictis binturong), an arboreal member of the family Viverridae native to Southeast Asia, is known for its prehensile tail and elusive nature [
9]. In the wild, binturongs are primarily frugivorous with occasional opportunistic carnivory, with activity patterns that tend to peak at dusk and continue through the night [
10]. In captivity, they spend most of their time resting, engaging in less locomotion and exploration than their wild counterparts, likely due to enclosure design and environmental complexity [
11]. Given the species’ reliance on scent-marking for communication and territoriality, olfactory stimuli may offer a promising means of increasing behavioral activity and engagement with their environment [
12]. However, artificial scent stimuli could potentially interfere with natural scent-marking behaviors by masking or altering intraspecific chemical cues [
13]. Further research is necessary to determine whether specific scents enhance engagement without disrupting essential behavioral patterns.
Previous studies have shown that olfactory enrichment can enhance behavioral diversity in zoo-housed species by increasing locomotion, foraging, and exploratory behaviors, while in some cases, reducing stress-related behaviors [
13]. Lavender oil has been associated with reductions in stress-related behaviors and cortisol concentrations in zoo-housed carnivores, including Asiatic wolves, jungle cats, and hyenas [
14]; however, its effects may be short-lived and appear to vary by species. In contrast, earlier studies in primates yielded equivocal results, as lavender was tested as part of a multi-oil blend, and its specific effects could not be isolated [
15]. These mixed findings underscore the need for species-specific and ecologically relevant scent enrichment strategies. Meanwhile, food-based scents and enrichment items have successfully stimulated foraging behaviors in omnivores and carnivores [
8,
16]. In this context, lavender oil was selected for its widespread use in zoo enrichment and to represent a non-food olfactory stimulus alongside two food-based enrichment types (thawed fish and hard-boiled egg).
To help address this gap, the present exploratory study examined enrichment-related behavior in two zoo-housed juvenile male binturongs. We evaluated the behavioral responses of two binturongs at the Minnesota Zoo to three distinct stimuli: lavender oil (an olfactory-only enrichment), thawed fish, and hard-boiled egg (both olfactory and dietary enrichments). We assessed how these stimuli influenced locomotion, foraging, resting, and visibility to zoo visitors. These behavioral measures were selected because they are commonly used indicators of animal welfare in zoo environments. Locomotion and foraging reflect activity levels and the expression of species-typical behaviors, while resting may indicate arousal or relaxation states. Visibility to visitors was included as a proxy for enclosure use and voluntary environmental engagement, which may reflect increased stimulation or reduced avoidance behaviors under enriched conditions. Based on prior findings in other taxa, we predicted that food-based enrichment would likely elicit greater foraging activity than the non-food olfactory stimulus, and that lavender oil would reduce locomotion and increase resting behavior in the binturongs.
2. Materials and Methods
All procedures were reviewed and approved by the University of Minnesota Institutional Animal Care and Use Committee (IACUC#2402-41764A) and were conducted in accordance with the U.S. Animal Welfare Act.
2.1. Animals and Housing
This study was conducted at the Minnesota Zoo, an AZA-accredited facility located in Apple Valley, Minnesota. The subjects were two juvenile male binturongs (
Arctictis binturong), named Felix and Garrett, aged one and two years, weighing 17.1 kg and 18.38 kg, respectively. They were housed in a 409.21 ft
2 enclosure within the Tropics Trail exhibit, designed to resemble their natural habitat. The enclosure featured large climbing branches, two hammocks for resting, a multi-level rock structure for elevation and movement, and a cave-like tunnel that leads to a holding area that allows the binturongs to retreat from public view (
Figure 1). The space was illuminated by natural sunlight through a glass ceiling, enhancing the enclosure’s naturalistic environment.
The binturongs had ad libitum access to food and water. Their diet included seasonal fruits, vegetables, and a rotating selection of protein sources, which included mice, chicken, rabbit, quail, and rat (see full diet in
Appendix A). The animals had free movement between the public-facing enclosure and the off-exhibit holding area throughout the study period.
2.2. Experimental Design
This study assessed the behavioral effects of three enrichment stimuli:
Lavender oil (olfactory enrichment): A few drops were applied to the designated branches.
Thawed fish (olfactory and dietary enrichment): Small portions were placed on the same enclosure features.
Hard-boiled egg (olfactory and dietary enrichment): Egg pieces were similarly positioned on the same enclosure features.
These stimuli were selected based on their practicality, sensory contrast, and biological plausibility for the species. Hard-boiled egg and thawed fish were already part of the binturongs’ routine rotational diet and provided both olfactory and nutritional stimulation (
Table A1). Although these items are not core components of wild binturong diets, binturongs are opportunistic omnivores that are known to consume a wide variety of plant and animal matter, including small vertebrates, eggs, and carrion when available [
10]. These food items were selected to represent protein-based, scent-rich enrichments that reflect the species’ ecological flexibility. Although familiar, the food items were offered outside of standard feeding times and placed in novel locations within the enclosure to encourage foraging and exploratory behaviors. Lavender oil, although not biologically relevant, was included as a widely used olfactory enrichment in zoos and served as a non-food comparative stimulus. The term “naturalistic appearance” refers to the fact that these items could be discreetly placed on exhibit elements without introducing brightly colored or artificial objects, thereby maintaining the visual aesthetics of the habitat.
Each stimulus was introduced individually, with the day and time of day varying, but was always placed on the same elevated log located in the central portion of the enclosure to ensure visibility and equal accessibility for both animals and behavioral observers. This consistent placement allowed for standardized delivery across sessions and facilitated reliable observation from the public viewing area. Although varying the location of enrichment could potentially promote more locomotion, we prioritized a fixed location to control for spatial access and isolate behavioral responses to the enrichment type. Enrichment items were applied by a trained zookeeper directly onto designated enclosure features, and data collection began only after the zookeeper exited the enclosure. A minimum two-hour observation period followed each enrichment application.
Each enrichment type was presented twice, for a total of six enrichment sessions, with conditions administered in randomized order. Each enrichment stimulus was presented independently, and no two enrichment types were applied during the same session. Control sessions, defined as observations with no enrichment, were conducted prior to enrichment implementation and were collected between 31 January and 29 February 2024. These control sessions served as the comparison condition for evaluating the enrichment effects. All enrichment sessions were conducted over an eight-day period from 5 April to 12 April 2024, using a repeated-measures design in which both binturongs were exposed to all enrichment conditions. While this allowed for within-subject comparisons, the small sample size limited the ability to apply formal repeated-measures statistical models.
2.3. Data Collection and Behavioral Measurements
Behavioral observations were conducted using scan sampling at one-minute intervals for sessions that were intended to last 20 min each, following each enrichment application. The observers documented behaviors using ZooMonitor software (Version 2.5, Lincoln Park Zoo, Chicago, IL, USA, 2024), categorizing them according to the definitions outlined in
Table 1. The observers recorded data while standing in the designated public viewing area directly in front of the exhibit, where zoo visitors typically observe the binturongs. A total of four trained observers participated in the study, with only one observer collecting data per session. Although the sessions were planned to last 20 min, minor deviations in duration occurred due to practical constraints (e.g., animal visibility, observer timing, etc.), and exact session durations were recorded and used in the analyses. Each enrichment type was observed for approximately 40 min (two sessions). Although each enrichment condition was presented in two separate sessions, behavior was recorded independently for each binturong, resulting in four individual-level observations per enrichment type. Twelve control sessions (nominally 20 min each) were collected prior to enrichment implementation, totaling approximately 240 min of control data. All control data were included in the descriptive analyses, and a randomized subset of four control sessions (approximately 80 min) was used in the Friedman test to balance the session numbers across the conditions.
The observations took place between 9:00 AM and 3:00 PM and were categorized by the time of day: morning (9:00–11:00 AM), midday (11:00 AM–1:00 PM), and afternoon (1:00–3:00 PM).
2.4. Observer Training and Reliability
To promote consistency and minimize observer bias, all observers completed standardized training on the ZooMonitor program, including detailed instruction on behavioral definitions and scoring procedures prior to data collection. The observers practiced data recording before the study and participated in structured group discussions to reconcile discrepancies. Consistency in scoring was reinforced through ongoing comparisons of recorded observations.
Formal inter-observer reliability testing (e.g., Cohen’s kappa) was not feasible due to logistical constraints with animal access and zoo scheduling. This is recognized as a limitation; however, efforts were made to support alignment in behavioral scoring across observers.
To assess potential discrepancies in observer scoring, we conducted a post hoc evaluation of observer effects. Descriptive comparisons and ANOVA models revealed that one observer (Observer R) consistently recorded higher levels of locomotion behavior compared to the other observers. In response, all sessions conducted by Observer R were removed, and we attempted to reanalyze the data using the remaining dataset. However, this subset lacked a sufficient number of sessions across all enrichment types to support valid repeated-measures analysis. As a result, the full dataset was retained for final analyses, and this variation is noted as a constraint in interpreting the locomotion data.
2.5. Statistical Analysis
All statistical analyses were conducted using SAS (Version 9.4, SAS Institute Inc., Cary, NC, USA). For each observation session, behavioral data were calculated as the percentage of total session time spent performing each behavior: (behavior duration ÷ session duration) × 100. This “session-level behavior” value was used as the primary outcome for all analyses.
Data normality was assessed using the Shapiro–Wilk test. As most behavioral variables were not normally distributed, we prioritized descriptive statistics and non-parametric testing.
Descriptive statistics (e.g., mean, standard deviation, median, minimum, and maximum) were calculated for each behavior under each enrichment condition. A Friedman test was used to compare the session-level behavioral outcomes across enrichment types. To maintain balance, this analysis included four sessions per enrichment condition (two per animal) and four randomly selected control sessions. We focused on four primary behaviors, such as locomotion, foraging, inactive, and resting, which accounted for the majority of the activity budget. Rare behaviors (e.g., scent-marking, drinking, etc.) were excluded due to insufficient frequency for meaningful comparison.
Exploratory generalized linear models (GLMs) were constructed to visualize the enrichment effects on foraging frequency while adjusting for unbalanced session numbers. Models used a Poisson distribution with a log link function, with a negative binomial distribution applied when overdispersion was detected. The dependent variable was the number of foraging events per session, with the enrichment condition as a fixed effect. Session duration (natural log) was included as an offset to adjust for variations in the observation length. These GLMs were used for exploratory visualization and not for formal inferential testing.
A two-way ANOVA was conducted to examine the effects of enrichment condition and individual identity on session-level behavior. Despite some behavioral variables not meeting strict normality assumptions, ANOVA was applied based on its robustness to moderate violations when the residuals were approximately symmetrical and the sample sizes were similar [
17]. The dependent variable was the percentage of time spent in each behavior per session. Fixed effects included the enrichment type and individual identity, with their interaction tested to explore individual response differences. Post hoc comparisons were conducted using Tukey’s Honestly Significant Difference (HSD) test to control for multiple comparisons. Residual plots confirmed approximate symmetry, supporting ANOVA’s validity in this context.
These ANOVA models were conducted independently of the Friedman tests. The Friedman tests evaluated within-subject behavioral differences using a balanced subset of sessions, while ANOVA enabled exploration of individual differences across the full dataset.
Although this study used a repeated-measures design, where both binturongs were exposed to each enrichment condition, formal repeated-measures statistical models (e.g., linear mixed-effects models) were not feasible due to the small sample size (n = 2) and limited number of balanced sessions per condition. Instead, the Friedman test was used to compare the within-subject behavioral responses across enrichment conditions while partially accounting for repeated observations. However, this approach does not allow for individual-by-condition interaction modeling. Future studies should incorporate more subjects and replicate sessions to support the use of linear mixed models or generalized estimating equations, which would strengthen statistical inferences and better account for repeated-measures structure.
3. Results
We evaluated the behavioral responses of two juvenile male binturongs across four treatment conditions: a control condition (no enrichment) and three enrichment conditions. The enrichment types represented different sensory modalities. Hard-boiled egg and thawed fish served as both dietary and olfactory enrichment stimuli, while lavender oil functioned solely as an olfactory enrichment. The primary behaviors assessed were foraging, inactive (solitary), resting, and locomotion, with descriptive statistics summarized by condition (
Table 2).
To supplement the descriptive comparisons, a Friedman test was conducted using four randomly selected control sessions and four sessions from each enrichment condition. No significant differences were detected for foraging (p = 0.30), inactive (solitary) behavior (p = 0.60), or locomotion (p = 0.60). Resting behavior approached significance (p = 0.10), indicating a potential trend that may warrant further investigation. Given the absence of statistically significant effects, descriptive statistics were used as the primary method for interpreting behavioral outcomes. Observed behavioral patterns are summarized in detail below.
3.1. Effect of Enrichment on Binturong Behavior
The descriptive results indicated that foraging behavior was highest in the hard-boiled egg condition, with lower levels were observed in the control, lavender oil, and thawed fish treatments. Locomotion appeared relatively consistent across the conditions, though greater variability was observed in the lavender oil condition. Resting behavior demonstrated a trend toward increased frequency in the lavender oil condition (p = 0.10), suggesting a potential calming effect. No other behaviors showed notable differences across conditions.
3.1.1. Foraging Behavior
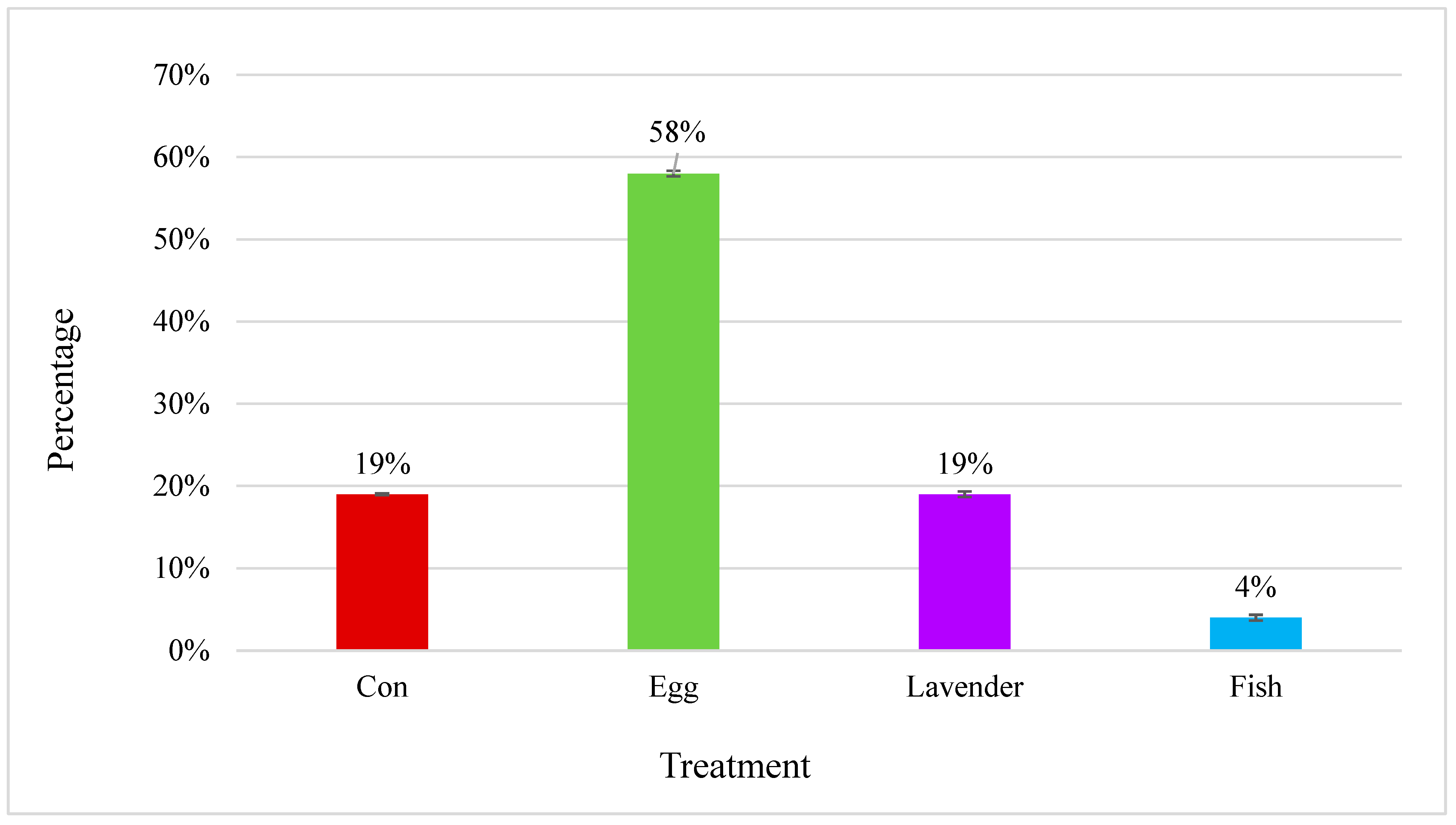
The session-level descriptive data indicated that foraging behavior was highest in the hard-boiled egg condition (mean = 7.8%), followed by the control (2.0%), lavender oil (1.8%), and thawed fish (0.5%). A Friedman test did not detect statistically significant differences across the enrichment conditions (p = 0.30).
To supplement the descriptive findings, an exploratory generalized linear model (GLM) was used to evaluate foraging behavior across all enrichment conditions while adjusting for differences in session duration. The model included all available sessions, including the overrepresented control condition, and incorporated an offset for session length to account for variations in the observation time. This approach provided estimates of the predicted foraging rates (i.e., events per minute) for each enrichment type (
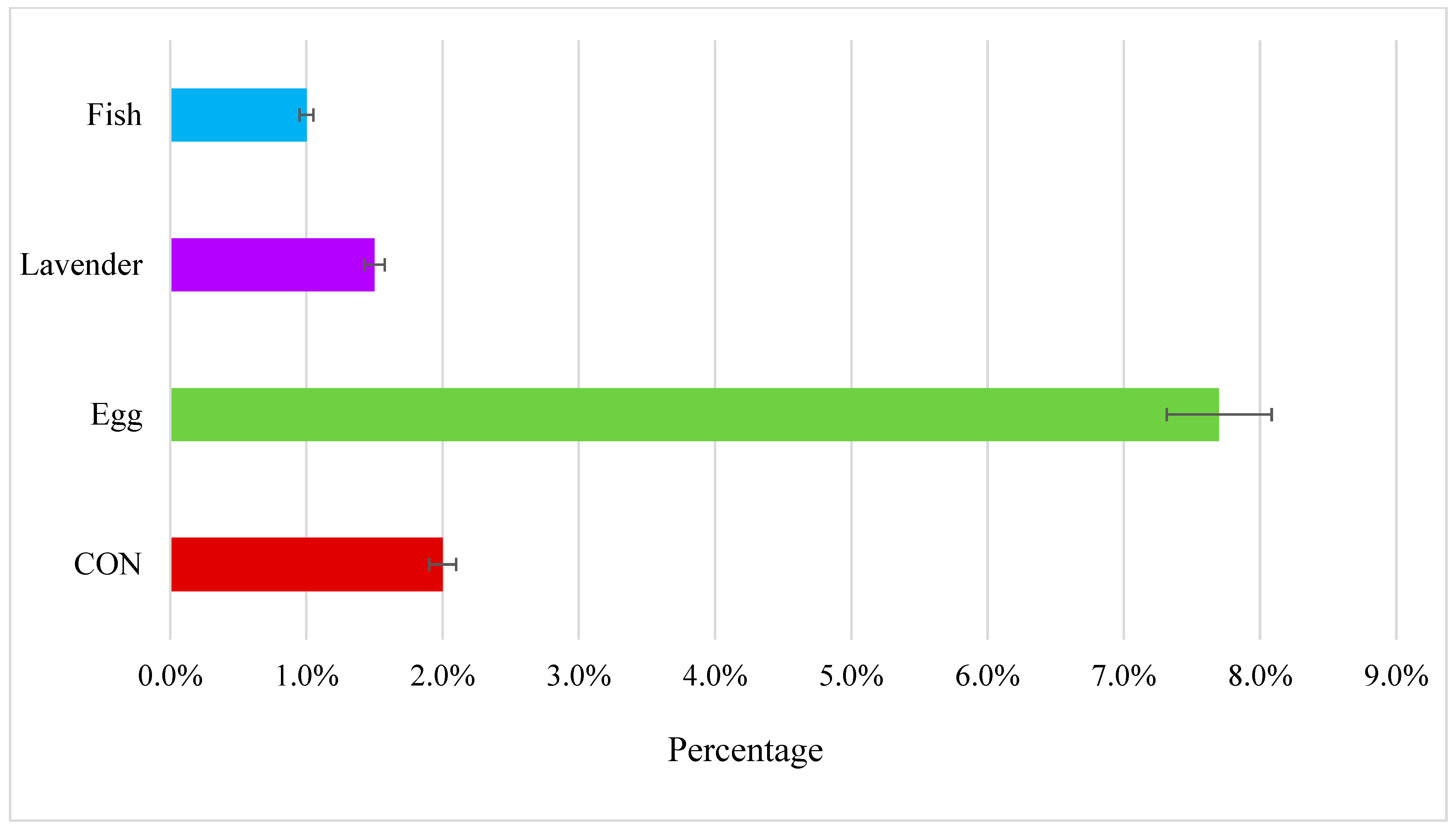
Figure 2). Additionally,
Figure 3 presents the raw observed proportion of foraging events per minute by enrichment condition.
3.1.2. Resting and Locomotion
The session-level data indicated that resting behavior was highest in the lavender oil condition (mean = 4.5%), followed by the hard-boiled egg and control conditions (both means = 1.5%), and was lowest in the thawed fish condition (mean = 0.0%). A Friedman test indicated a trend-level difference in resting behavior across the enrichment types (p = 0.10).
Locomotion appeared relatively consistent across the conditions, with the mean values ranging from 5.3% in the thawed fish condition to 10.8% in the lavender oil condition. The Friedman test did not detect significant differences in locomotion across treatments (p = 0.60).
3.2. Individual and Observer Effects
3.2.1. Influence of Individual Identity
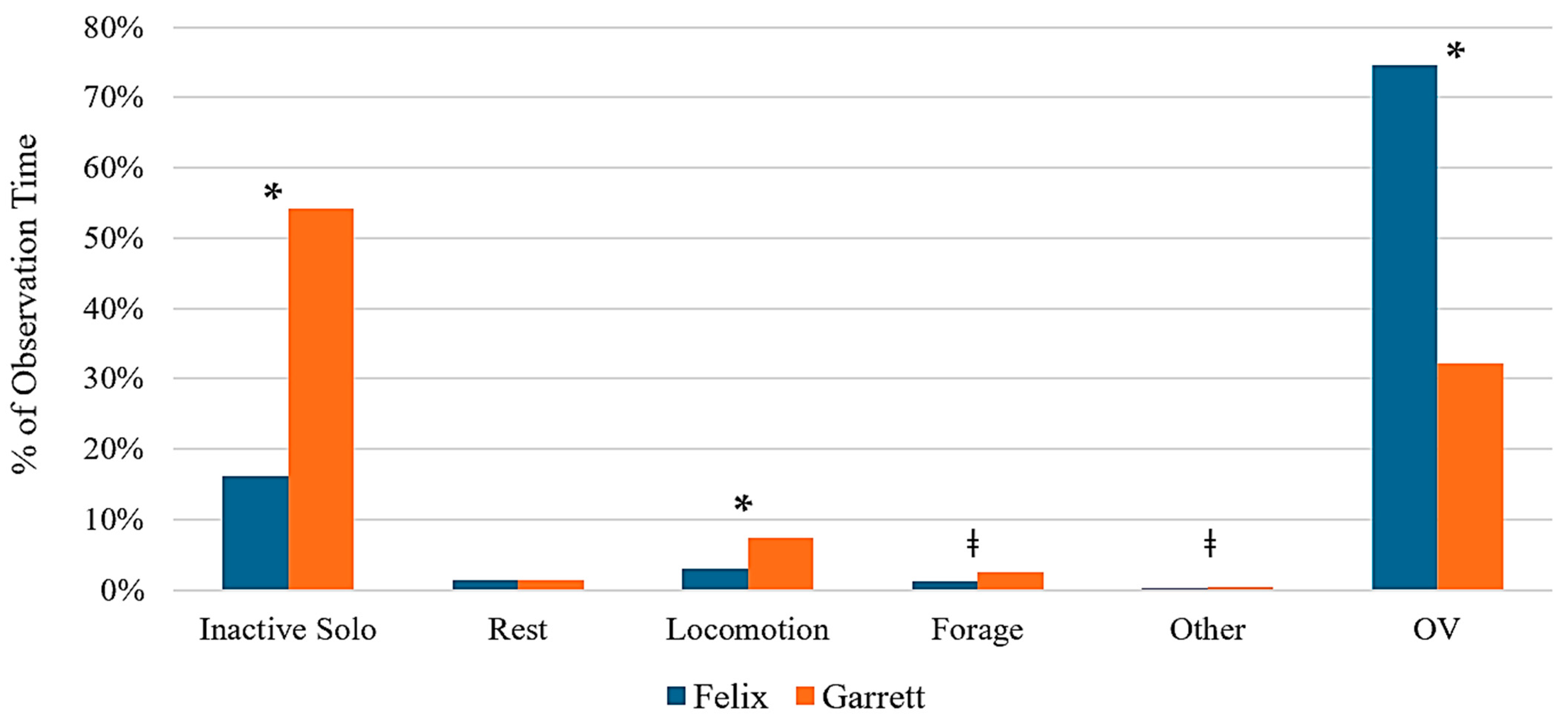
A two-way ANOVA revealed significant differences between individuals in inactive (solitary) behavior (
p < 0.001), locomotion (
p = 0.006), and time spent out of view (
p < 0.001). Trend-level effects were also observed for foraging behavior (
p = 0.08) and other behaviors (
p = 0.08), suggesting some variability in behavioral expression between individual binturongs (
Figure 4). No significant interaction effects were detected between individual identity and enrichment type (
p > 0.05).
3.2.2. Observer and Session Duration Effects
Observer identity significantly influenced the recordings of locomotion behavior (p < 0.001), while no significant differences were observed for the other behaviors. Session duration was positively correlated with locomotion (R2 = 0.21, p < 0.001), inactive (solitary) behavior (R2 = 0.13, p = 0.004), and time spent out of view (R2 = 0.18, p = 0.0008).
To address concerns about observer consistency, all data collected by Observer R were removed, and a reanalysis of the behavioral outcomes was attempted. While general descriptive patterns remained similar, removal of these sessions resulted in an insufficient number of observations for statistical testing, particularly for the hard-boiled egg and lavender oil enrichment conditions. Given the resulting reduction in sample size and the need to preserve within-subject comparisons, all statistical analyses were conducted using the full dataset. However, the locomotion results should be interpreted with caution due to potential observer-related biases.
3.3. Time of Day Effects on Behavior
Although each enrichment type was presented only twice, the session schedule was balanced across the morning, midday, and afternoon time periods. Accordingly, the time-of-day effects were evaluated across all enrichment sessions combined rather than within individual enrichment types.
A one-way ANOVA revealed a trend-level effect of the time of day on inactive (solitary) behavior (F(2,3) = 2.63,
p = 0.08) and a significant effect on locomotion (F(2,3) = 3.95,
p = 0.02;
Table 3). Post hoc comparisons indicated that inactive (solitary) behavior was significantly higher during the afternoon compared to midday (T(3) = 2.66,
p = 0.02), while no significant differences were found between the morning and other time periods (
p > 0.05). For locomotion, although the overall ANOVA was significant, no pairwise comparisons reached statistical significance. A trend-level increase in locomotion was observed in the afternoon compared to midday (T(3) = 2.43,
p = 0.06).
No significant effects of time of day were observed for foraging behavior (F(2,3) = 0.95, p = 0.43) or time spent out of view (F(2,3) = 1.21, p = 0.33). These analyses were based on the enrichment sessions only and did not include data from the control condition.
Because control data were not included in this analysis, it remains unclear whether the observed time-of-day effects reflect enrichment exposure or the binturongs’ inherent daily activity rhythms. These findings should be interpreted cautiously and considered exploratory due to the small sample size and limited session replication.
Inactive (solitary) behavior was highest during the afternoon, while locomotion showed a non-significant increase during the afternoon compared to midday (
Figure 5). These temporal trends were observed during the enrichment sessions only and should be interpreted cautiously due to the exclusion of control data.
4. Discussion
In this exploratory study, we evaluated the behavioral responses of two juvenile male binturongs to three enrichment stimuli. While no statistically significant differences were found across the enrichment types for foraging, inactive, resting, or locomotion behaviors, the descriptive trends suggest differential engagement. Foraging behavior appeared highest during the egg enrichment sessions, while resting was more common during the lavender oil sessions. Although these differences did not reach statistical significance, the observed trends offer valuable direction for future hypothesis-driven research with larger sample sizes and increased replication.
While no behavioral outcome reached statistical significance, this study was designed as an exploratory assessment and was not statistically powered to detect small or moderate effects. The small sample size and limited number of sessions per condition constrained our ability to draw definitive conclusions. However, in zoo-based research involving rare or underrepresented species like the binturong, descriptive data remain highly valuable. These data provide foundational insights into species-typical responses and inform future hypothesis-driven studies. Descriptive trends, such as the apparent increase in foraging with egg enrichment or increased resting with lavender oil, offer direction for more robust follow-up studies. Future research should incorporate power analyses or simulations during study planning to ensure adequate sample sizes for statistical inference.
Observer-related factors also warrant consideration when interpreting the results. Although formal inter-observer reliability testing was not feasible, post hoc statistical analyses indicated that observer identity significantly influenced locomotion data. No effects were found for other behaviors. One observer appeared to record higher locomotion levels than others, introducing potential scoring variations. Due to the limited number of sessions available after removing this observer’s data, the full dataset was retained to maintain sample balance and statistical power. However, this limitation highlights the need for formal reliability assessments in future behavioral studies and supports the use of overlapping observation sessions to strengthen scoring consistency.
These behavioral outcomes align with known patterns in binturong ecology and sensory reliance. As opportunistic omnivores, binturongs consume a variety of fruits, eggs, and small prey, and are known to rely heavily on olfactory cues for foraging and communication [
10,
12,
13]. The greater behavioral engagement observed in response to food-based enrichment, particularly egg, is consistent with their ecological tendencies and may reflect both gustatory and olfactory motivations. In contrast, lavender oil (a non-biologically relevant scent) elicited less consistent activity, though increased resting during lavender sessions may indicate a mild calming effect.
Encouraging activity during daytime hours is particularly important in managed care settings, as binturongs are naturally crepuscular or nocturnal and often exhibit low visibility during zoo visitor hours. Furthermore, enrichment that promotes locomotion and foraging may be beneficial for physical health, as gastrointestinal and musculoskeletal conditions are common in zoo-housed binturongs [
18]. Although preliminary, these findings support the inclusion of ecologically relevant, scent-rich food items in enrichment programs and offer a foundation for future studies exploring the welfare impacts of sensory enrichment in underrepresented species.
The two food-based enrichment stimuli, thawed fish and hard-boiled egg, appeared to elicit greater behavioral engagement than the non-food lavender oil. The descriptive trends indicated increased foraging and locomotion associated with these items, though differences were not statistically significant. These outcomes are consistent with the animals’ natural foraging tendencies, as food-based enrichment provides both olfactory stimulation and a nutritional reward [
8,
16]. However, because these foods were already part of the binturongs’ regular diet, their novelty may have been diminished. Presenting familiar items in novel contexts can still be enriching, but future studies may benefit from incorporating unfamiliar or seasonal foods to promote more robust behavioral responses.
These results align with enrichment research in other carnivorous and omnivorous species. Powell (1995) [
16] reported that food-based enrichment increased exploratory and foraging behaviors in captive African lions [
16], while Young (2013) [
8] found that scent-based stimuli tied to biologically relevant food sources sustained greater engagement than neutral scents [
8]. Collectively, these findings emphasize the importance of aligning enrichment strategies with species-specific ecology and natural foraging behaviors to promote welfare and behavioral diversity.
Although enrichment studies on binturongs remain limited, the patterns observed here are consistent with their ecology as scent-driven, opportunistic foragers. In the wild, binturongs rely heavily on olfaction and consume a broad diet that includes fruit, eggs, small prey, and carrion [
10,
11,
12]. The modest engagement seen with fish and egg enrichment likely reflects the ecological relevance of these strong-scented, high-protein stimuli. In contrast, lavender oil, despite its widespread use in zoo enrichment, elicited limited behavioral change, likely due to its lack of biological relevance for this Southeast Asian species. These results highlight the importance of taxon-specific scent cues when designing olfactory enrichment programs.
Inactivity is a persistent concern in zoo-housed binturongs, which tend to spend a substantial portion of the day resting [
11]. Their crepuscular activity pattern and arboreal habits require enrichment strategies that account for both temporal and spatial behaviors. In this study, food-based enrichment modestly increased activity levels, suggesting its potential for mitigating inactivity. Incorporating these types of stimuli into daily routines could support broader welfare goals by promoting daytime engagement, increasing movement, and encouraging species-typical foraging behaviors.
This study is limited by several key design constraints. The sample consisted of only two juvenile males housed at a single facility, restricting generalizability across sexes, age classes, or institutions. This study lacked concurrent control sessions during the enrichment phase, limiting our ability to attribute behavioral changes exclusively to the enrichment stimuli. The observation windows were brief and may not capture sustained effects. Additionally, only one non-food olfactory stimulus was tested, and external contextual variables such as ambient noise and visitor presence were not controlled.
Furthermore, this study employed a repeated-measures design in which both animals were exposed to each enrichment condition. While this approach helps reduce individual-level variability and allows for within-subject comparison, the small sample size and imbalanced number of sessions across conditions precluded the use of more robust repeated-measures statistical models (e.g., mixed-effects ANOVA). Instead, we relied on non-parametric tests and exploratory models to describe general behavioral patterns. Future studies using a larger number of individuals and more balanced replication would allow for formal modeling of repeated measures and could increase the reliability and generalizability of the enrichment effects.
Finally, although the enrichment types used in this study were selected for practicality, they were not chosen based on ecological specificity to binturongs. The dietary items matched existing husbandry routines, and lavender oil was selected for its frequent use in enrichment studies rather than its biological relevance. These limitations underscore the need for more intentional, species-informed enrichment design. Future studies should explore the effects of ecologically appropriate stimuli, such as tropical fruits, prey-scented objects, or conspecific odors. Multi-institutional collaborations, long-term observations, and the inclusion of physiological welfare metrics, such as cortisol or activity tracking, will strengthen future evaluations and inform evidence-based enrichment practices for binturongs and other underrepresented species.



 ,
, 


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}