
Examining Activity, Courtship, and Social Behaviour in Zoo-Housed Wreathed Hornbills: Potential Insights into Pair Compatibility



Abstract
1. Introduction
2. Materials and Methods
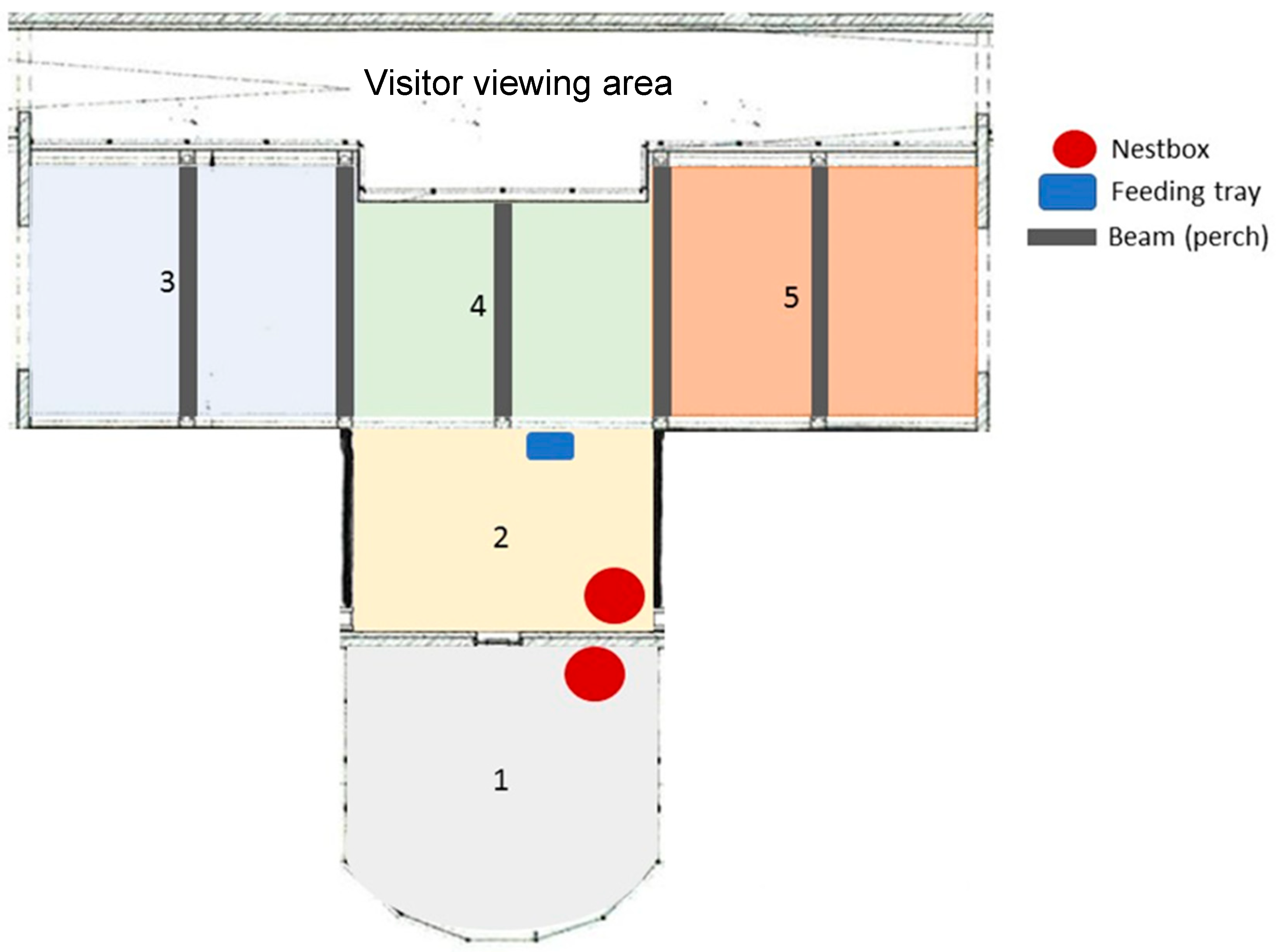
2.1. Study Population and Location
2.2. Data Collection
2.3. Data Analysis
3. Results
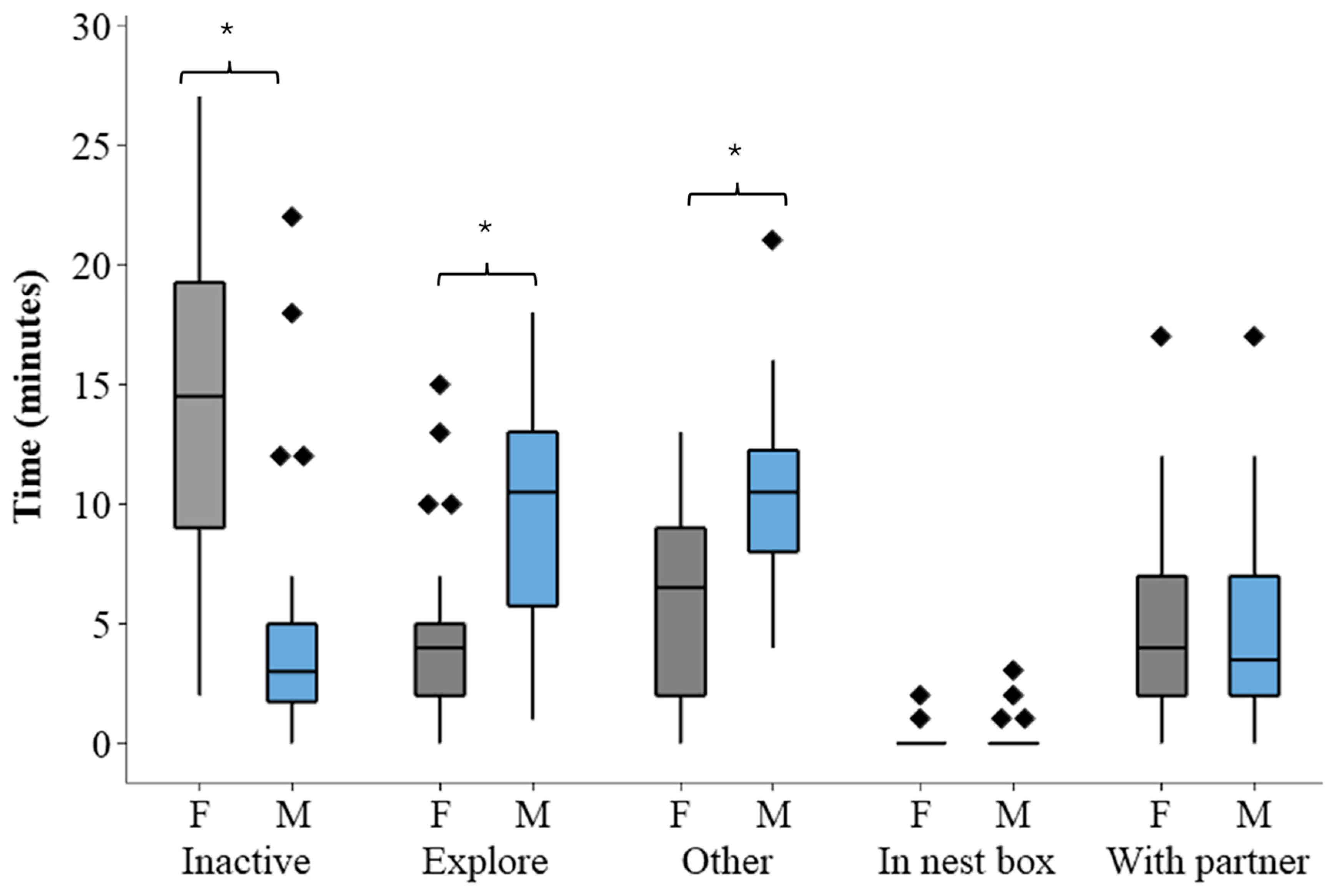
3.1. State Behaviours
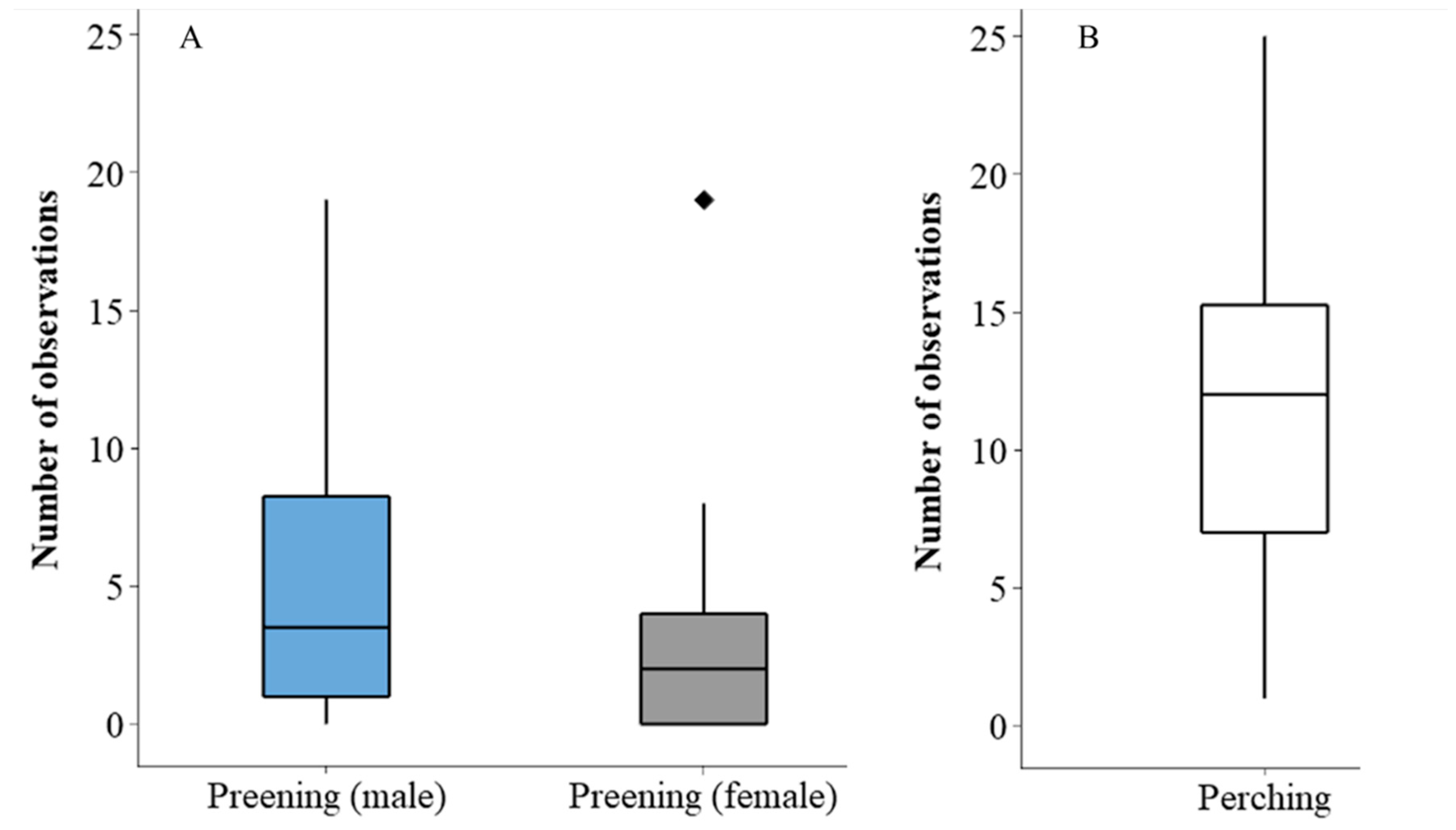
3.2. Event (Courtship) Behaviours
3.3. Event (Maintenance and Interaction) Behaviours
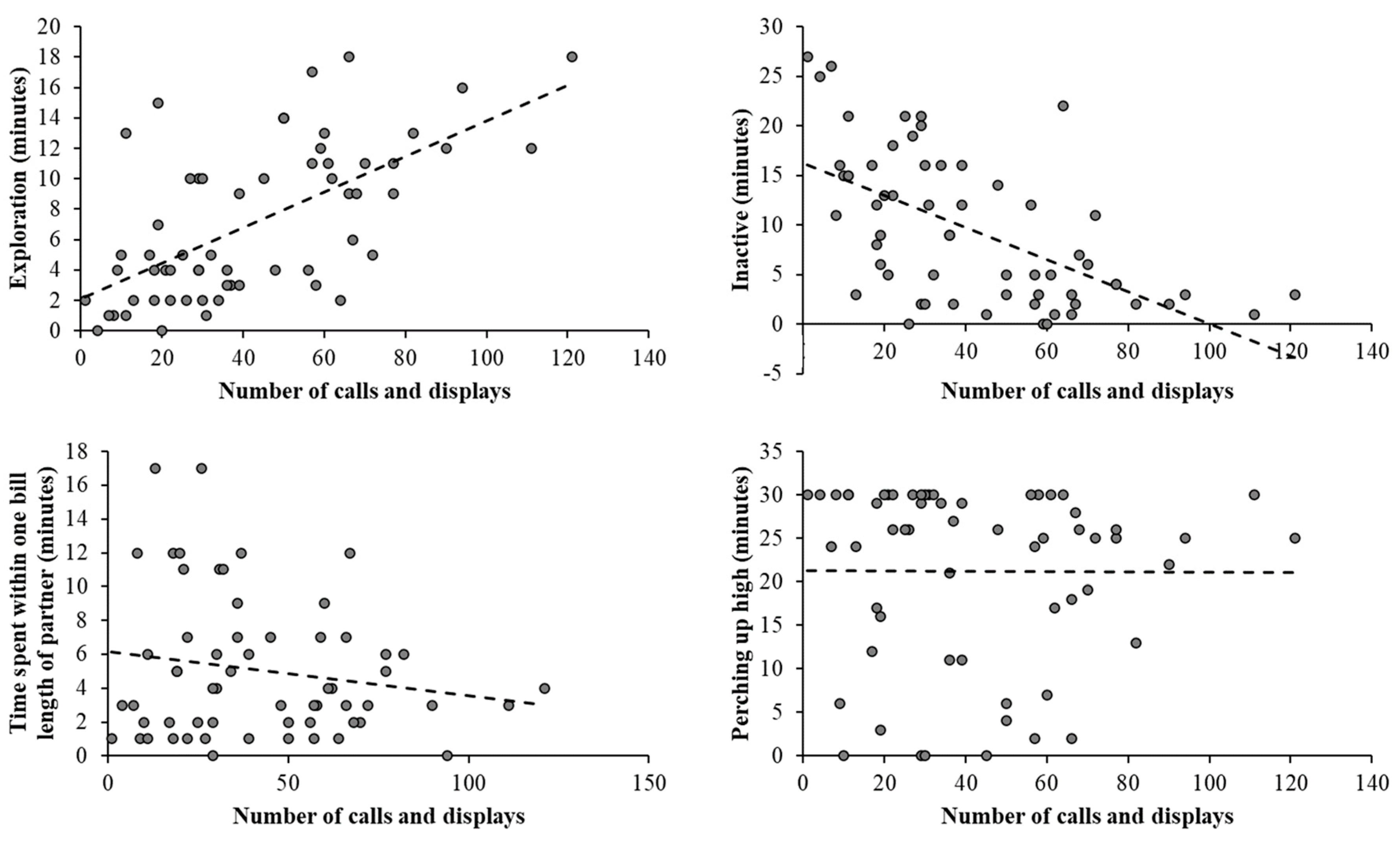
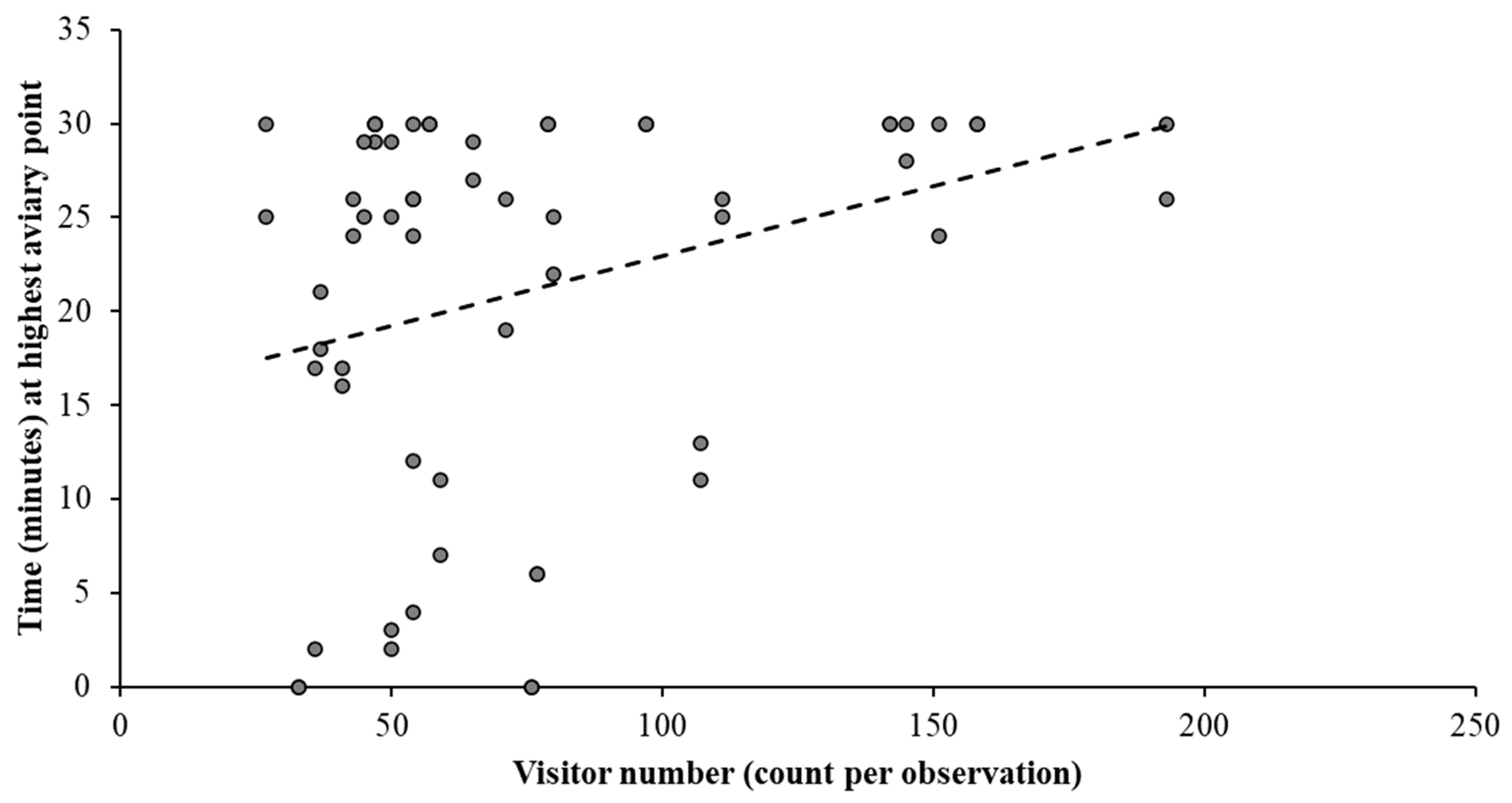
3.4. Behavioural and Environmental Impacts on Courtship Display
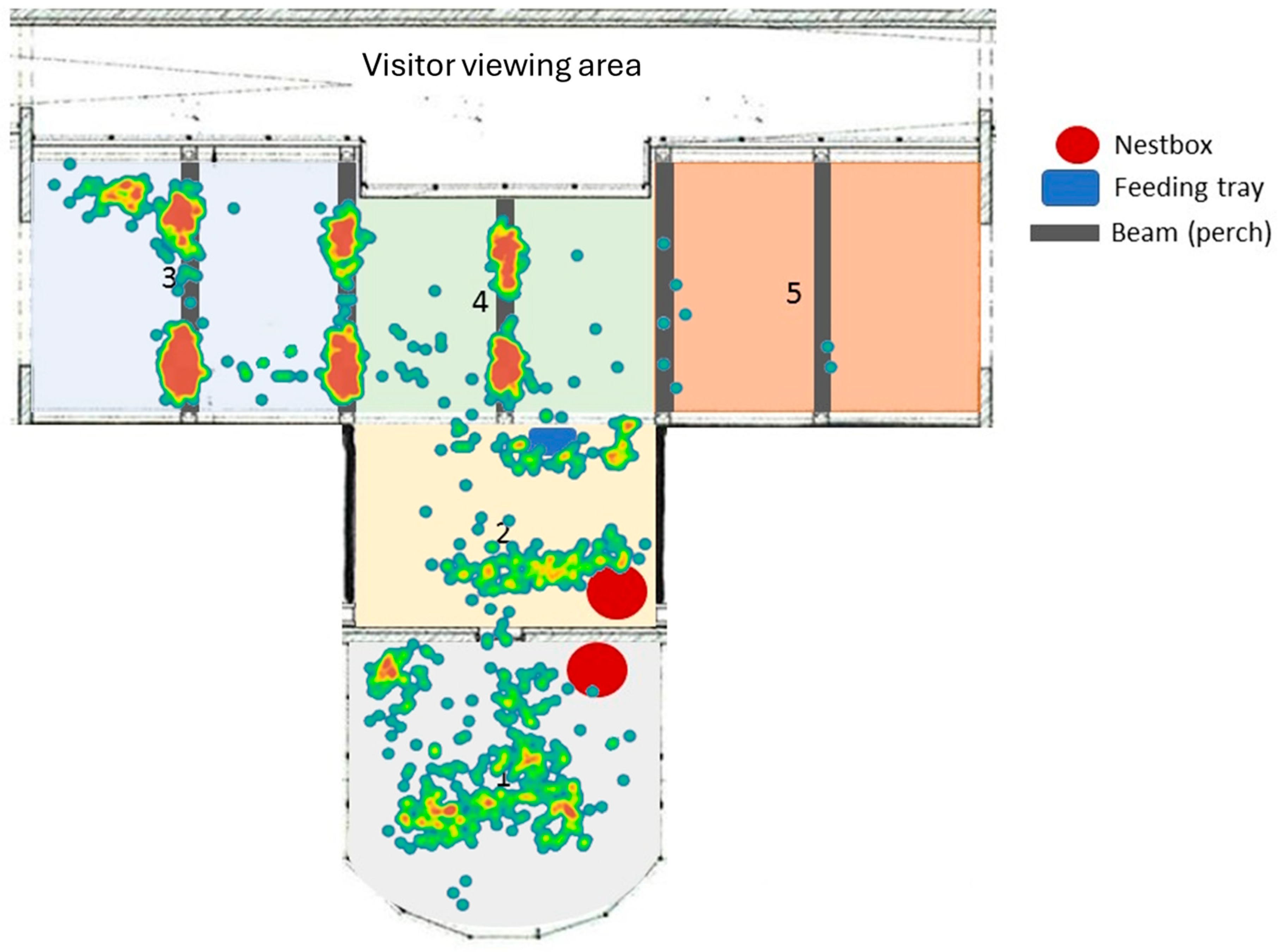
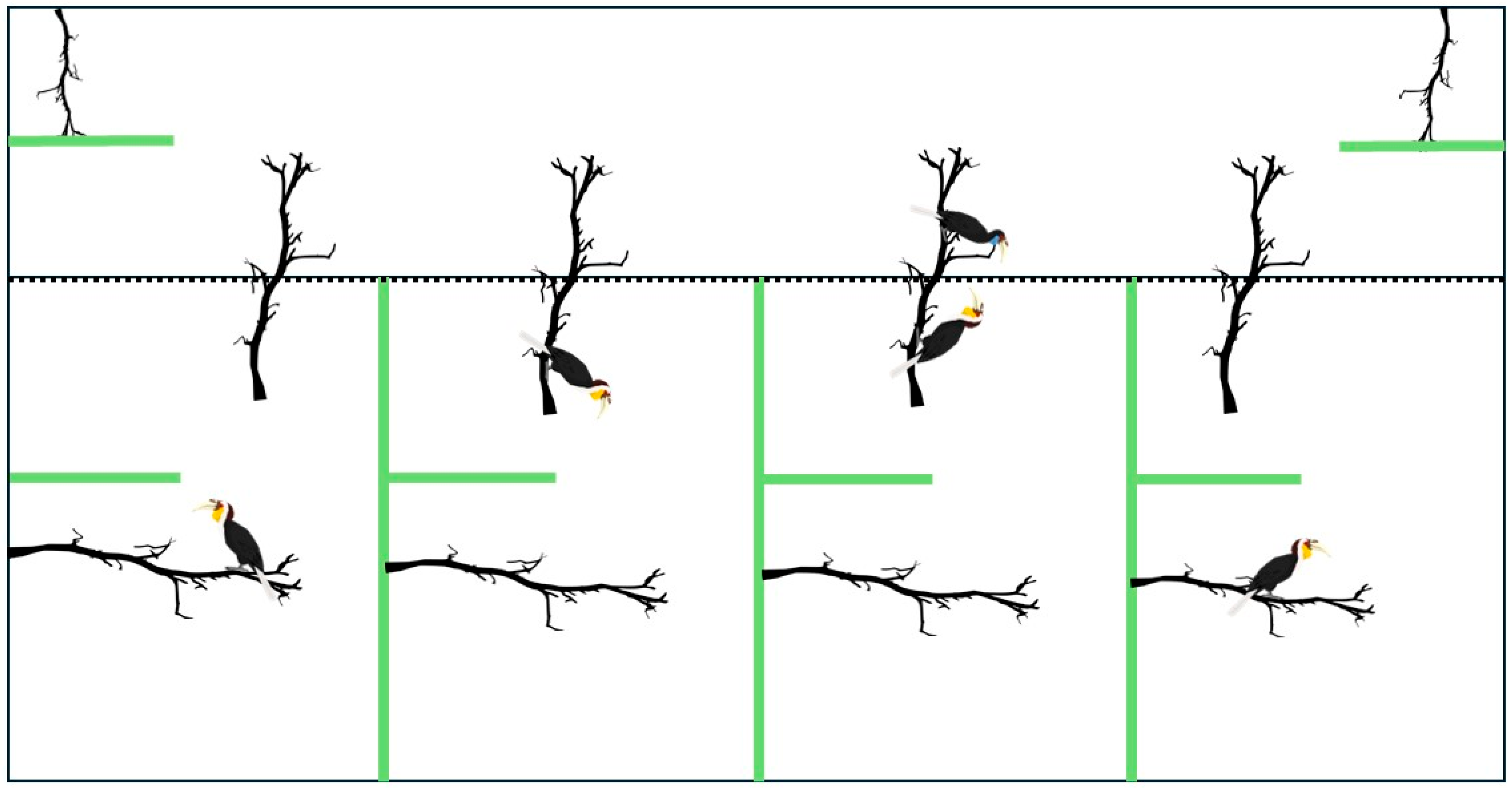
3.5. Enclosure Usage
4. Discussion
4.1. Behavioural Outcomes
4.2. Enclosure and Space Use
4.3. Research Extensions and Husbandry Developments
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kemp, A.C. Hornbills (Bucerotiformes). In Handbook of the Birds of the World; del Hoyo, J., Elliot, A., Sargatal, J., Eds.; Lynx Edicions: Barcelona, Spain, 2001; Volume 6, pp. 436–523. [Google Scholar]
- Beilby, J. The behavioural biology of hornbills, toucans, and kingfishers. In The Behavioural Biology of Zoo Animals; Rose, P.E., Ed.; CRC Press: Boca Raton, FL, USA, 2022; pp. 185–196. [Google Scholar]
- Rose, P.E.; Scales, J.S.; Brereton, J.E. Why the “visitor effect” is complicated. Unraveling individual animal, visitor number, and climatic influences on behavior, space use and interactions with keepers- a case study on captive hornbills. Front. Vet. Sci. 2020, 7, 236. [Google Scholar] [CrossRef]
- Kozlowski, C.P.; Bauman, K.L.; Asa, C.S. Reproductive behavior of the great hornbill (Buceros bicornis). Zoo Biol. 2015, 34, 328–334. [Google Scholar] [CrossRef] [PubMed]
- Brouwer, K.; Lammers, J.; Aparici Plaza, D.; Van Der Meer, R.; Hausen, N. EAZA Hornbill Taxon Advisory Group Regional Collection Plan; EAZA: Amsterdam, The Netherlands, 2020; p. 350. [Google Scholar]
- Galama, W.; King, C.E.; Brouwer, K. EAZA Hornbill Husbandry and Management Guidelines; EAZA: Amsterdam, The Netherlands, 2002. [Google Scholar]
- Kinnaird, M.F.; O’Brien, T.G.; Laman, T. The Ecology and Conservation of Asian Hornbills: Farmers of the Forest; University of Chicago Press: Chicago, IL, USA, 2008. [Google Scholar]
- Kemp, A. The Hornbills: Bucerotiformes (Bird Families of the World); Oxford University Press: Oxford, UK, 1995. [Google Scholar]
- Haimoff, E.H. A spectrographic analysis of the loud calls of helmeted hornbills Rhinoplax vigil. Ibis 1987, 129, 319–326. [Google Scholar] [CrossRef]
- Kemp, A.C.; Boesman, P.F.D. Wreathed Hornbill—Rhyticeros undulatus. Available online: https://birdsoftheworld.org/bow/species/wrehor1/cur/introduction (accessed on 23 October 2024).
- Kemp, A.C.; Kirwan, G.M. Blyth’s Hornbill—Rhyticeros plicatus. Available online: https://birdsoftheworld.org/bow/species/blyhor1/cur/introduction (accessed on 23 October 2024).
- Naniwadekar, R.; Mishra, C.; Isvaran, K.; Datta, A. Gardeners of the forest: Hornbills govern the spatial distribution of large seeds. J. Avian Biol. 2021, 52. [Google Scholar] [CrossRef]
- Zheng, X.; Zhang, L.; Yang, Z.; Chan, B.P.L. Flocking of hornbills observed in Tongbiguan Nature Reserve, Yunnan, China. Hornbill Nat. Hist. Conserv. 2020, 1, 42–44. [Google Scholar]
- Species360. Zoological Information Management System. Available online: https://zims.species360.org/Main.aspx (accessed on 3 July 2024).
- BirdLife International. Rhyticeros undulatus. IUCN Red List of Threatened Species. 2018, p. e.T22682528A132400385. Available online: https://datazone.birdlife.org/species/factsheet/wreathed-hornbill-rhyticeros-undulatus (accessed on 23 October 2023).
- Hadiprakarsa, Y.-Y.; Kinnaird, M.F. Foraging characteristics of an assemblage of four Sumatran hornbill species. Bird Conserv. Int. 2004, 14, S53–S62. [Google Scholar] [CrossRef]
- Groot, K.; Brereton, J.E.; King, C.E.; Rose, P.E. A preliminary global investigation into potential impacts on successful captive breeding for two species of Rhyticeros hornbill. Zoo Biol. 2024, 43, 405–415. [Google Scholar] [CrossRef] [PubMed]
- Mallapur, A.; Qureshi, Q.; Chellam, R. Enclosure design and space utilization by Indian leopards (Panthera pardus) in four zoos in southern India. J. Appl. Anim. Welf. Sci. 2002, 5, 111–124. [Google Scholar] [CrossRef] [PubMed]
- Rose, P.E.; Robert, R. Evaluating the activity patterns and enclosure usage of a little-studied zoo species, the sitatunga (Tragelaphus spekii). J. Zoo Aquar. Res. 2013, 1, 14–19. [Google Scholar]
- Forthman Quick, D.L.; Pappas, T.C. Enclosure utilization, activity budgets, and social behavior of captive chamois (Rupicapra rupicapra) during the rut. Zoo Biol. 1986, 5, 281–292. [Google Scholar] [CrossRef]
- Bateson, M.; Martin, P. Measuring Behaviour: An Introductory Guide, 4th ed.; Cambridge University Press: Cambridge, UK, 2021. [Google Scholar]
- Wark, J.D.; Cronin, K.A.; Niemann, T.; Shender, M.A.; Horrigan, A.; Kao, A.; Ross, M.R. Monitoring the behavior and habitat use of animals to enhance welfare using the ZooMonitor app. Anim. Behav. Cogn. 2019, 6, 158–167. [Google Scholar] [CrossRef]
- Cooper, M.; Jordan, L. Random time-activity budgets in captive Southern Ground Hornbill Bucorvus leadbeateri: Commentary. S. Afr. J. Sci. 2013, 109, 1–2. [Google Scholar] [CrossRef]
- Lehner, P.N. Sampling methods in behavior research. Poult. Sci. 1992, 71, 643–649. [Google Scholar] [CrossRef] [PubMed]
- RStudio Team. RStudio: Integrated Development for R. Available online: http://www.rstudio.com (accessed on 12 October 2024).
- R Core Team R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2023.
- Minitab 22.1. Minitab Statistical Software. Minitab: State College, PA, USA, 2024; Available online: www.minitab.com (accessed on 12 October 2024).
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; Sage: Thousand Oaks, CA, USA, 2019. [Google Scholar]
- Kleiber, C.; Zeileis, A. Applied Econometrics with R; Springer: New York, NY, USA, 2008. [Google Scholar]
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S, 4th ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Brereton, J.E.; Myhill, M.N.G.; Shora, J.A. Investigating the effect of enrichment on the behavior of zoo-housed southern ground hornbills. J. Zool. Bot. Gard. 2021, 2, 600–609. [Google Scholar] [CrossRef]
- Readyhough, T.S.; Joseph, S.L.; Vyas, K.; Schreier, A.L. The effects of Zoo Lights on animal welfare: A case study of great Indian hornbills at Denver Zoo. Zoo Biol. 2022, 41, 263–270. [Google Scholar] [CrossRef]
- Reilly, S.E. Breeding the rhinoceros hornbill buceros rhinoceros at the Audubon Park and Zoological Garden. Int. Zoo Yearb. 1988, 27, 263–269. [Google Scholar] [CrossRef]
- Jaroenwatee, W.; Pongsupath, N.; Wongyuen, N.; Suwanpugdee, A. A study on behavior in breeding season and develop nest box of great hornbill (Buceros bicornis linaeus) in Dusit Zoo. J. Agric. Res. Ext. 2018, 35, 963–971. [Google Scholar]
- Leighton, M. Hornbill social dispersion: Variations on a monogamous theme. In Ecological Aspects of Social Evolution; Rubenstein, D.I., Wrangham, R.W., Eds.; Princeton University Press: Princeton, NJ, USA, 1986; pp. 108–130. [Google Scholar]
- Policht, R.; Petrů, M.; Lastimoza, L.; Suarez, L. Potential for the use of vocal individuality as a conservation research tool in two threatened Philippine hornbill species, the Visayan hornbill and the rufous-headed hornbill. Bird Conserv. Int. 2009, 19, 83–97. [Google Scholar] [CrossRef]
- Sibarani, M.C.; Utoyo, L.; Pratama, R.D.; Danus, M.A.; Sudrajat, R.; Surahmat, F.; Marthy, W. Long-term monitoring of nesting behavior and nesting habitat of four sympatric hornbill species in a Sumatran lowland tropical rainforest of Bukit Barisan Selatan National Park. Hornbill Nat. Hist. Conserv. 2020, 1, 17–29. [Google Scholar]
- Poonswad, P.; Tsuji, A. Ranges of males of the great hornbill Buceros bicornis, brown hornbill Ptilolaemus tickelli and wreathed hornbill Rhyticeros undulatus in Khao Yai National Park, Thailand. Ibis 1994, 136, 79–86. [Google Scholar] [CrossRef]
- Ross, S.R. Issues of choice and control in the behaviour of a pair of captive polar bears (Ursus maritimus). Behav. Process. 2006, 73, 117–120. [Google Scholar] [CrossRef]
- Thicks, S.F. Is there a visitor effect on Abyssinian ground hornbills (Bucorvus abyssinicus), Papuan wreathed hornbills (Aceros plicatus), wrinkled hornbills (Aceros corrugatus) and toco toucans (Ramphastos toco) in a captive zoo environment? Plymouth Stud. Sci. 2008, 1, 30–55. [Google Scholar]
- Phillips, C.; Peck, D. The effects of personality of keepers and tigers (Panthera tigris tigris) on their behaviour in an interactive zoo exhibit. Appl. Anim. Behav. Sci. 2007, 106, 244–258. [Google Scholar] [CrossRef]
- Friard, O.; Gamba, M. BORIS: A free, versatile open-source event-logging software for video/audio coding and live observations. Methods Ecol. Evol. 2016, 7, 1325–1330. [Google Scholar] [CrossRef]
- Weber, W.D.; Fisher, H.S. An inexpensive wireless video recording system for continuous behavioral observations. BioRxiv 2019. [Google Scholar] [CrossRef]
- Huyghe, M. EAZA Best Practice Guidelines Aegypius Monachus—Cinereous Vulture; European Association of Zoos and Aquaria: Amsterdam, The Netherlands, 2023. [Google Scholar]
- Rose, P.E.; Freeman, M.; Hickey, I.; Kelly, R.; Greenwell, P.J. Considering what animals “need to do” in enclosure design: Questions on bird flight and aviaries. Birds 2024, 5, 586–603. [Google Scholar] [CrossRef]
- Rimlinger, D.; Theule, J.; Bass, K. Breeding history and husbandry of the superb bird-of-paradise (Lophorina superba). Zoo Biol. 2021, 40, 485–490. [Google Scholar] [CrossRef]
- Theule, J.; Rimlinger, D. Breeding history and husbandry of the raggiana bird-of-paradise (Paradisaea raggiana). Zoo Biol. 2023, 42, 162–170. [Google Scholar] [CrossRef]
- Rose, P.E. Evidence for aviculture: Identifying research needs to advance the role of ex situ bird populations in conservation initiatives and collection planning. Birds 2021, 2, 77–95. [Google Scholar] [CrossRef]
- Both, C.; Dingemanse, N.J.; Drent, P.J.; Tinbergen, J.M. Pairs of extreme avian personalities have highest reproductive success. J. Anim. Ecol. 2005, 74, 667–674. [Google Scholar] [CrossRef]
- Wark, J.D.; Cronin, K.A. The behavior patterns of giraffes (Giraffa camelopardalis) housed across 18 US zoos. PeerJ 2024, 12, e18164. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| State Behaviours | Event Behaviours | Enclosure Use | On Same Perch as Partner | Visitor Number |
|---|---|---|---|---|
| Inactive. The individual is perched and does not move. | Calling/displaying. The individual is jumping from one side to another on a horizontal perch, while opening and closing its wings simultaneously. The bird emits species-specific vocalisations. | On ground. Individual is located on the ground | Count of occurrences of when individuals are located on the same perch more than one beak length apart. Used as an indication of social space use. | The cumulative number of visitors seen during a 30 min observation period. |
| Exploration. The individual is actively looking around or is in locomotion. | Allofeeding. The individual is attempting to feed their partner, both unsuccessful and successful attempts. | Middle level. Individual is located between the ground and the height of the aviary beams. | ||
| Other active. All other activities not listed including feeding, cleaning their beaks. | Aggressive behaviour (towards a conspecific). Individual is showing aggressive behaviour towards their partner (e.g., biting, pecking, jabbing). | Up high. Individual is located on or above the height of the aviary beams. | ||
| At nest box. The individual is looking into the nest box or is inside the nest box. | Allopreening. Manipulation of partner bird’s feathers. | |||
| With partner. As a measure of social association. A bird is within a beak’s length of their partner. Should one bird be trying to perch close to their partner, but the other bird moves away, we recorded this as “with partner” for the instigator but not for the bird that was choosing otherwise. | Preening. Manipulation of own feathers. | |||
| Interaction with any other species. Any other interactions observed between the hornbill and another bird species within the enclosure, for example aggression. |
| Predictor | Estimate | SE | Z Value | p Value |
|---|---|---|---|---|
| Sex (male) | 0.367 | 0.164 | 2.232 | 0.026 |
| Within one beak length of partner (min) | −0.071 | 0.017 | −4.232 | <0.001 |
| Perching high up (min) | 0.018 | 0.006 | 2.874 | 0.004 |
| Inactivity (min) | −0.064 | 0.013 | −4.922 | <0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Groot, K.; Rose, P. Examining Activity, Courtship, and Social Behaviour in Zoo-Housed Wreathed Hornbills: Potential Insights into Pair Compatibility. J. Zool. Bot. Gard. 2025, 6, 10. https://doi.org/10.3390/jzbg6010010
Groot K, Rose P. Examining Activity, Courtship, and Social Behaviour in Zoo-Housed Wreathed Hornbills: Potential Insights into Pair Compatibility. Journal of Zoological and Botanical Gardens. 2025; 6(1):10. https://doi.org/10.3390/jzbg6010010
Chicago/Turabian StyleGroot, Kees, and Paul Rose. 2025. "Examining Activity, Courtship, and Social Behaviour in Zoo-Housed Wreathed Hornbills: Potential Insights into Pair Compatibility" Journal of Zoological and Botanical Gardens 6, no. 1: 10. https://doi.org/10.3390/jzbg6010010
APA StyleGroot, K., & Rose, P. (2025). Examining Activity, Courtship, and Social Behaviour in Zoo-Housed Wreathed Hornbills: Potential Insights into Pair Compatibility. Journal of Zoological and Botanical Gardens, 6(1), 10. https://doi.org/10.3390/jzbg6010010