Abstract
Visitors to zoos can have positive, neutral, or negative relationships with zoo animals. This makes human–animal interactions (HAIs) an essential component of welfare and an important consideration in species selection for zoo exhibits and in enclosure designs. We measured the effect of visitors on reptiles by comparing open and closed periods during the lockdowns in response to the COVID-19 pandemic in the UK in a low-resolution dataset for thirteen species of reptiles and a high-resolution dataset focussing on just one of these. Scan sampling on thirteen reptile species (two chelonians and eleven squamates) showed species-specific differences in response to the presence/absence of visitors, with most taxa being only weakly affected. High-resolution scan sampling via video footage of an off-show and on-show enclosure was carried out for tokay geckos (Gekko gecko) over the open and closed periods. In this part of the study, tokay geckos were significantly more visible during zoo closure than when visitors were present on-exhibit, but there was no change in off-show animals, indicating the effect of visitors as opposed to other factors, such as seasonality, which applied equally to both on- and off-show animals. The high-resolution study showed that a significant effect was present for tokay geckos, even though the low-resolution suggested that they were more weakly affected than other taxa. Our results indicate that, for cryptic species such as this, more intensive sampling may be required to properly understand visitor effects. Our data do not allow the interpretation of effects on welfare but show that such assessments require a species-specific approach.
1. Introduction
Modern zoos have multiple roles, including education, scientific research, species conservation and entertainment [1]. Due to an increase in public concern and greater expectations of captive animals’ welfare [2,3,4], the effects of the zoo environment on welfare states have been under heightened scrutiny [5]. Animal welfare is the state of the animal related to their ability to cope with their environment [5,6]. In response, zoos have strengthened efforts to develop ways in which their welfare can be assessed [7,8,9], including investigating the effects of human–animal interactions (HAI).
Human–animal interactions and relationships are omnipresent in a zoo setting and can be beneficial to both human and animal participants. However, they may also create welfare risks and ethical dilemmas [10]. Most HAI research has been predominately on livestock [5,10,11]. Where HAI have been studied in a zoo setting, this is often mammal-focused, especially primate-focused, and the results highlight some negative effects of visitors on animal behaviours [4,10,12] but, also, positive impacts [4,13,14].
Zoo animals frequently encounter both unfamiliar and familiar humans in their environment and may form one of three relationship types: (i) a negative relationship, where humans cause the animal to be fearful and avoidant, (ii) a neutral relationship, where no consequences are caused by interactions with humans and can lead to habituation, or (iii) a positive relationship, where an interaction with humans could cause positive emotions [4].
In addition to their long-standing reputation for being ‘behaviourally simple’ [15], reptiles, a nonmonophyletic functional group including all ectothermic amniotes (that is, squamates, crocodilians, chelonians and tuataras), and, specifically, their behaviours, are under-researched compared to mammalian groups, complicating the development of an effective welfare assessment [16]. Although these groups do not represent a monophyletic clade, their overlapping physiological constraints and ecology means that reptile requirements in captivity are similar, and it is practically useful to treat them as one group; the term ‘reptile’ is used to this end in the present work. Some studies have shown that captivity stress and related chronic stress in reptiles can lead to abnormal behaviours such as freezing behaviour, avoidance, aggression, vigilance, behavioural complexity and decreased exploratory behaviour [17]. However, the presence of such/abnormal behavioural signs does not infer poor health and welfare and vice versa with normal behavioural signs (normal behaviour can be described as natural behaviours within their appropriate context and range) [18]. Equally, neutral and positive effects of visitors have been shown in reptiles, with neutral interactions occurring in Nile crocodiles (Crocodylus niloticus [19]) and some species, such as varanid lizards, exhibiting a range of positive behaviours, including play behaviour, while zoo visitors are present [20].
Additionally, the ecological characteristics of a species, such as the use of antipredation tactics or camouflage, may be used to partially predict visitor effects among species. In the absence of such data for reptiles, a recent study comparing amphibian (the only other terrestrial ectothermic vertebrate group) visibility during the COVID-19 lockdown periods and open periods found that all studied amphibians showed either an increase in visibility or maintained a consistent visibility for times of full lockdown and a decrease when visitors were present. However, visitors’ presence had a greater effect on camouflaged species rather than aposematic species, with golden mantella (Mantella aurantiaca) and golden poison dart frogs (Phyllobates terribilis) habituating quicker to changes in visitor numbers than their camouflaged counterparts [21].
The unusual, prolonged closure and reopening periods of UK zoos during the 2020 lockdowns amidst the COVID-19 pandemic provided an opportunity to assess the effects of long-term absences of visitors on reptilian behaviours at ZSL London Zoo. This study aims to expand the understanding of human–animal interactions by comparing the visibility of a range of species during periods of visitors’ presence and absence in a rapid assessment case study context similar to that provided for amphibians by Boultwood et al. [21].
2. Materials and Methods
To evaluate the effects of visitors on the behaviours of reptiles, two separate studies were conducted. A transect was established for low-resolution scan sampling of 2 chelonian, 5 snake and 6 lizard species (Table 1) over two lockdown periods, and high-resolution scan sampling was conducted on tokay geckos, Gekko gecko. Enclosures are referred to as ‘open’ (=outside of lockdown) or ‘closed’ (=during lockdown). Both types of samples included the same on-show exhibit of tokay geckos. All animals were maintained with environmental parameters appropriate to their geographic origin and were serviced by keepers at any time between 8:30 a.m. and 5:00 p.m., with the bulk of husbandry between 8:30 a.m. and 12:00 noon. When the exhibits were open, visitors were present between 10:00 a.m. and 5:00 p.m. The exhibits in question were closed to the public from the 16 March until the 4 July 2020, and from the 4 November until the 2 December 2020. Two data collection strategies controlled for the impact of seasonality as opposed to lockdowns. Firstly, we collected data from all exhibits under both open and closed conditions in both the summer and winter. Secondly, we collected data from an on-show exhibit, which was subject to changes in visitor numbers throughout the lockdown alongside a consistent keeper presence, and an off-show enclosure, which was subject to consistent human presence in the form of keeping staff only for one species (tokay geckos; see below). As all data were collected purely observationally, this study did not require ethical review and was registered internally at ZSL as ZSL Projects Database entries ZDZ130 and ZDZ133.

Table 1.
A list of the study species with a brief description of their ecology and numbers of observations in the open and closed states in the broad sampling dataset. The tokay geckos were the same animals used in the focal sampling in the on-show condition.
2.1. Low-Resolution Study
2.1.1. Data Collection
Data were collected between the 13 June 2020 and the 17 December 2020, thus encompassing 2 COVID-19 lockdowns and the subsequent period of reopening. A total of 1643 observations were recorded by a random pool of herpetology keepers and zoo staff (nine staff members, all having undertaken the same familiarisation training for the sampling method) doing visual surveys from the public area of the Reptile and Amphibian House four to five times per day between 8:00 a.m. and 17:00 p.m. GMT. The study focused on 13 reptile species, each housed in a single-species public exhibit, with a total number of individuals per species varying from 1 to 12 (Table 1).
At each enclosure, observers recorded the date and the number of active (the animal engaged in any activity), inactive (the animal not moving other than breathing), and out-of-sight (OOS) individuals. The proportions of animals undertaking these behaviours were then calculated using data on the total number of animals per enclosure. The transect sampling was opportunistic, although often followed a set route, and involved observers surveying exhibits for approximately 10 s. The number of observations in each state (open or closed) is provided in Table 1.
2.1.2. Statistical Analysis
Our primary question was to determine whether the behaviour of our animals was influenced by the absence of visitors caused by the COVID lockdowns and if this effect was species-dependent. Our design was intrinsically pseudo-replicated, because all individuals from the same species were maintained in the same terrarium, as is often the case in a zoo setting. Considering that the same degree of care and effort to offer species-specific captive conditions is put into the creation and maintenance of each terrarium, we thereafter assumed that observed effects could be attributed to intrinsic differences among species.
We used the proportion of active and proportion of out-of-sight individuals as the response variables in generalised linear mixed models (GLMM) with binomial error distributions and logit link functions [35]. For each response variable, we first fitted a full model with the enclosure status (open/closed) as a main effect, with a correlated random intercept and enclosure status slope per species, and with the month (integer) as a random effect to control for differences in intercepts due to seasonality. As all individuals from the same species were housed together in the same terrarium, uncorrelated intercepts per species could not be modelled without involving pseudo-replication, which would have resulted in biased estimates and increased type I errors [36].
The significance of the correlated random intercept and slope of the enclosure status within species was assessed using log-likelihood ratio tests (LRTs) on models fitted by the restricted maximum likelihood [37]. Inference about the fixed effects was carried out by stepwise deletion and comparisons between model candidates in an information-theoretic approach using the Akaike information criterion (AIC). Our final models’ assumptions were verified graphically. Their adequacy was evaluated through dispersion statistics, approximated as the residual deviance divided by the residual degrees of freedom. Their fit was assessed using marginal and conditional R2 metrics [38].
Parameter estimates were all calculated using Laplace approximation [39]. Conditional modes and their variances were computed and plotted to get insights on the predicted effects of the enclosure status in each species and of their uncertainty. The analysis was conducted using the packages lme4 [40], MuMIn [41] and lattice [42] in R version 4.1.0 software [43] and is available by open-access at https://github.com/LeaFieschiMeric/lockdown_on_reptiles (accessed on 21 October 2021).
2.2. High-Resolution Study
2.2.1. Data Collection
Focused observations were collected from tokay geckos in order to trial a more intensive data collection method. Geckos were observed in the same on-show exhibit included in the low-resolution study and in an off-show exhibit in a room into which visitors were never permitted. Both enclosures were maintained with similar environmental parameters and enclosure designs appropriate for the species. The on-show exhibit held three animals (Table 1), and the off-show exhibit, which was smaller, held a single animal.
Timelapse cameras (Plotwatcher Pro, Day 6 Outdoors, Columbus, GA, USA) collected data for 5 days across the closed and open treatments from two lockdown cycles. Lockdown 1 commenced the 11 April 2020, reopened to visitors commencing the 15 June 2020, a second open period commencing the 26 October 2020 and a second lockdown period commencing the 9 November 2020. Two cameras were used to incorporate the entirety of the on-show enclosure, and one camera was used to do the same in the off-show enclosure. Cameras were set to photograph every 5 s during daylight periods. Using GameFinder software (Day 6 Outdoors), instantaneous scan sampling conducted by a single observer (IK) occurred every 10 min between 10:00 a.m. and 17:00 p.m. GMT., recording the number of visible geckos. The presence of a gecko was defined as any whole body part visible (head, body and limb). Where two cameras were used, the videos were cross-referenced to avoid double counting of animals present where the fields of view overlapped.
2.2.2. Statistical Analysis
The total number of geckos seen each day in each enclosure was divided by the number of geckos in the respective enclosures to generate a standardised visibility score for each day; these data were analysed to compare the open and closed periods. Data were analysed using randomisation, which is suited to studies with small sample sizes, in order to avoid pseudo-replication [44,45]. A randomisation test with 10,000 iterations of simulated cases assigned randomly to open or closed categories in proportions reflecting the real dataset and using the residual of the baseline and treatment means as the test statistic was used to analyse the visibility scores, performed in Microsoft Excel® for Windows 365® (following References [44,45,46]).
3. Results
3.1. Low-Resolution Study
Model selection based on LRTs and parsimonious AIC comparison (Supplementary Materials Table S1) led to final models of identical structures for both response variables: a fixed effect of the enclosure status and a correlated random intercept and enclosure status slope per species (Equation (1)):
where Yij is the proportion of active (or out-of-sight) individuals during the jth observation of the ith species and follows a binomial distribution of parameters yij and ni. Logit(yij) is the latent value for the jth observation of the ith species; it is defined by the enclosure status x, by the species-specific random intercept and random enclosure status slope effects b0i and b1i, by the overall fixed intercept and fixed enclosure status slope β0 and β1 and by the random error eij. The random intercept and slope, b0i and b1i, and the random error term are assumed to come from normal distributions with a mean of 0 and the respective variances σ2b0, σ2b1 and σ2e. Both the graphical assessment of the residuals, the absence of over- or under-dispersion and the conditional R2 (Table 2) in our final models suggest an acceptable fit.
Table 2.
Regression parameters estimates on the log-odds scale, with their standard errors and z-values, for both final binomial GLMMs, along with estimated measures of the variance and dispersion (ĉ). The confidence intervals are given on the scale of the linear predictor.
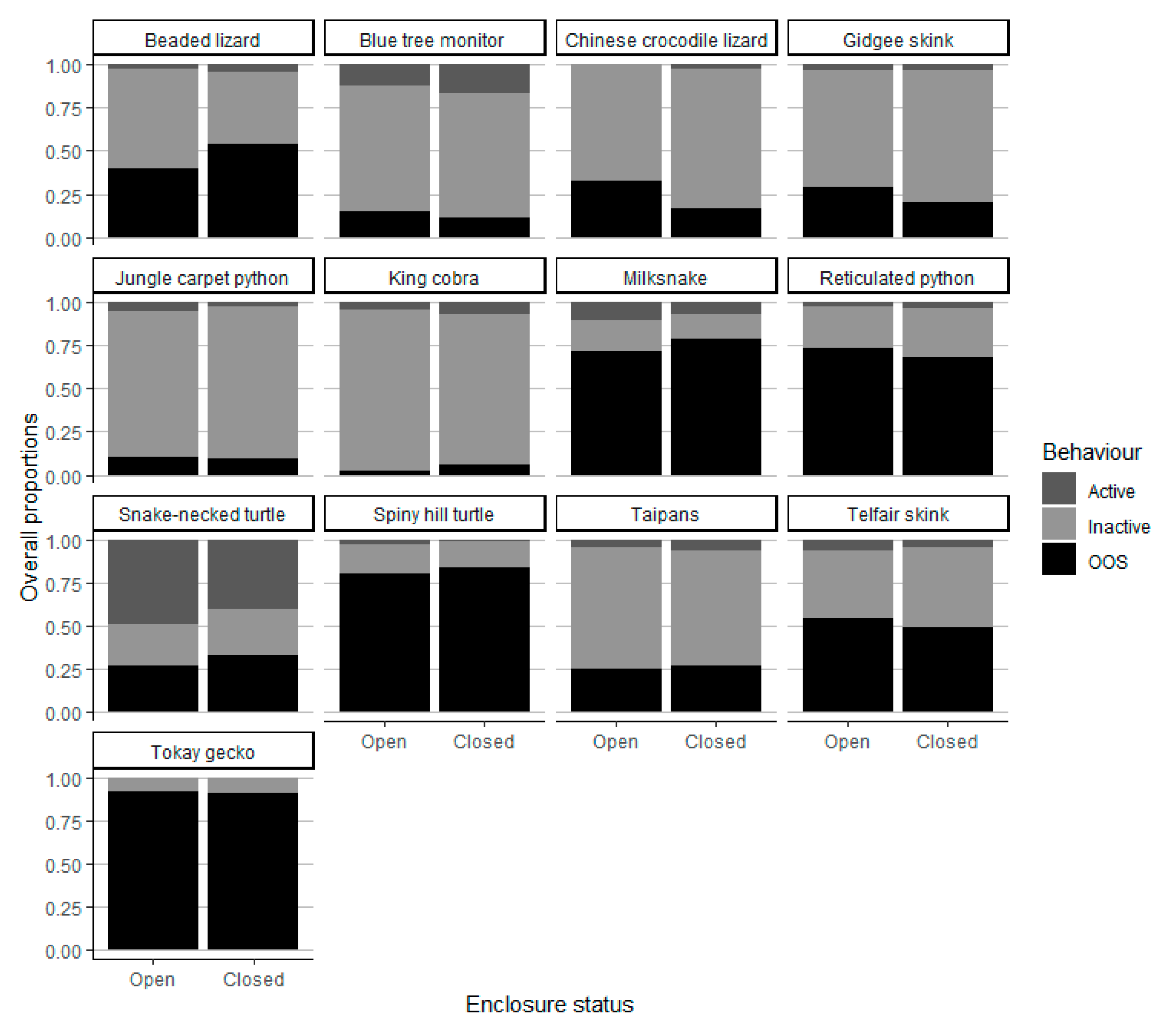
Figure 1 shows the relative proportions of observations in the active, inactive and OOS states for each species in the open and closed zoo periods.

Figure 1.
Proportion of active, inactive and out-of-sight individuals with visitors present (Open) and absent (Closed) in our 13 species of interest.
3.1.1. Proportion of Active Individuals
In general, the proportion of active individuals was slightly lower under the open conditions (odds ratio = 1.09; i.e., animals were 1.09 times less likely to be active when visitors were present). However, our final model had a substantial explanatory power (conditional R2 = 0.51), while its fixed effects structure alone accounted for little of the phenotypic variance (marginal R2 < 0.001), suggesting that the effect of the presence of visitors on the activity levels was very different between species (Table 2 and Figure 1).
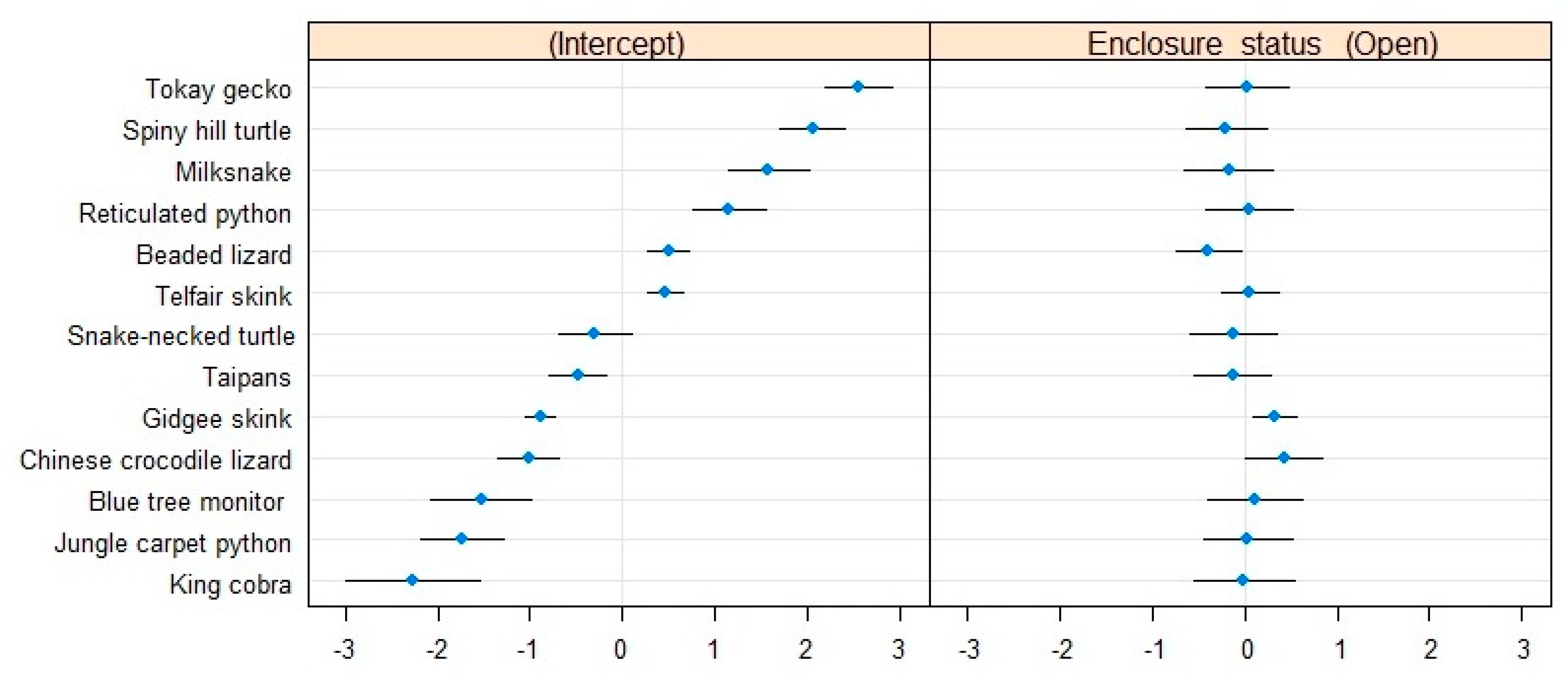
The between-species variance (σ0 = 1.19) suggested a baseline difference in activity between species (Table 2). Indeed, the conditional modes showed that the snake-necked turtle was, in general, very active compared to the other species, which could be a result of its aquatic lifestyle. Conversely, tokay geckos were the least active animals in the collection (Figure 2).
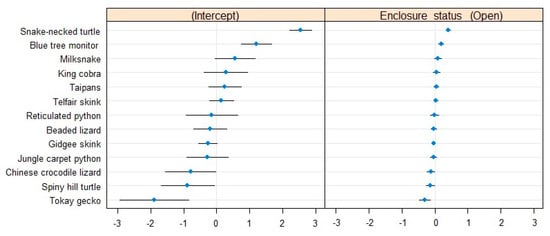
Figure 2.
Conditional modes and variances associated with the species levels in our model of the proportion of active individuals. The difference in intercept (left) shows how different species are in their baseline activity levels relative to the rest of the collection. The difference in slope (right) indicates the strength of the effect of the presence of visitors on the activity of each species relative to the rest of the collection.
The effect of the presence of visitors on the proportion of active individuals was species-specific, but its stability was comparable across species. The snake-necked turtle and the milksnake seemed to be more active under open conditions, whereas the blue tree monitor was more active when the zoo was closed. Most of the rest of the sampled collection seemed relatively less impacted by the visitors. Interestingly, the magnitude of the effect of the presence of visitors on activity levels seems directly dependent on the baseline activity levels of the species (Figure 2). As such, the tokay geckos were the least affected species by the presence/absence of visitors in their activity levels, but they also had extremely low baseline activity levels in general.
3.1.2. Proportion of Individuals Out-of-Sight
Overall, the proportion of individuals hiding was slightly higher under the open condition (odds ratio = 1.09). Again, the enclosure status alone accounted for little of the total phenotypic variance (marginal R2 < 0.001) compared to the random intercept and slope (conditional R2 = 0.63), suggesting that the effect of the presence of visitors on the visibility of animals was very different between species (Table 1 and Figure 1).
The between-species variance (σ0 = 2.22) corroborated the graphically observable global difference in the proportion of hiding individuals between species (Figure 1). The conditional modes indicated that the king cobra was the most often visible animal in the collection, whereas tokay geckos usually remained out-of-sight. Gidgee skinks, telfair skinks and Chinese crocodile lizards had average positions amongst the collection in terms of visibility, but they were the most stable, i.e., the most predictable in their proportions of visible individuals (Figure 3). Beaded and Chinese crocodile lizards appeared to be the species most influenced by the presence of visitors, with beaded lizards hiding less when the zoo was opened and Chinese crocodile lizards conversely hiding more in the presence of the public. None of the pythons from our collection responded differently to the open or closed conditions (Figure 3). The negative effect of the presence of visitors on the visibility of gidgee skinks was the most consistent, suggesting that they were relatively stable in their behavioural reactions to visitors (Figure 1 and Figure 3).
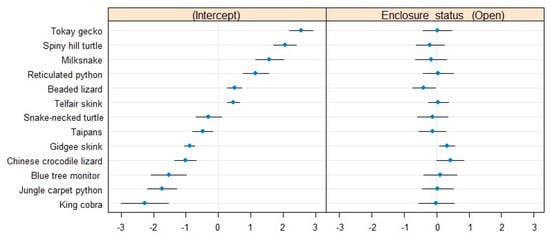
Figure 3.
Conditional modes and variances associated with the species levels in our model of the proportion of out-of-sight individuals. The difference in intercept (left) shows how different species are in their baseline visibility levels relative to the rest of the collection. The difference in slope (right) indicates the strength of the effect of the presence of visitors on the visibility of each species relative to the rest of the collection.
3.2. High-Resolution Study
For the on-show exhibit, the mean (SD) standardised gecko visibility scores were 4.715 (4.68) and 0.82 (2.27) for the lockdown and open treatments, respectively. For the off-show exhibit, the mean (SD) standardised gecko visibility scores were 12.4 (11.3) and 29.8 (10.13) for the lockdown and open treatments, respectively.
There was a significant effect of the treatment (open or closed) in the on-show exhibit (10,000 iterations; p = 0.04) but not the off-show exhibit (10,000 iterations; p = 0.08).
4. Discussion
The design of our study is intrinsically pseudo-replicated and so does not allow us to distinguish an effect of differences among enclosures from differences among species or from individual animals/groups of animals. This is a limitation imposed by the zoo context of the study, where the exhibits and stocking densities could not be controlled and where replicates were not available. In this way, this work was intended as a case study to highlight potential visitor impacts on a poorly studied group and to encourage further, more detailed research, as well as to provide some evidence base for collection planning and welfare mitigation in this group of animals [21]. As the enclosures were designed to best cater to the needs of the animals living in them, we chose to assume that we could consider the observed effects to be attributable to intrinsic differences between species or at least individual representative of those species used here, rather than enclosure effects. We henceforth used ‘species’ as short-hand for the ‘species–individual–exhibit’ combination.
The baseline proportions of active and of out-of-sight individuals were species-specific. Tokay geckos and spiny hill turtles were generally the least active and least visible animals in our sample, which was likely a result of both the general activity levels and more crepuscular or nocturnal diel cycles (also, see below). Reptiles are rarely strictly nocturnal, crepuscular or diurnal, and even taxa with nocturnally skewed activity patterns such as Tokay geckos routinely exhibit activity including basking and hunting during the day (or data to demonstrate this fact—many non-zero observations were recorded). However, in the rest of the species from our collection, there was no obvious pattern linking activity and visibility; in fact, the most active (snake-necked turtle and blue tree monitor) were not the most visible (king cobra and jungle carpet python) species. This baseline difference in levels of activity and of visibility should be taken into account when establishing protocols in the future. It also adds evidence for the necessity to consider reptile species independently rather than treating them as one homogenous group. Taking this baseline into account, our results suggest that visitors’ presence can have an impact on the behaviour of captive reptiles. Overall, the proportions of active and of visible individuals decreased by 1.09 times in the presence of visitors. The strength of this effect was the same in both response variables considered here.
The effect of the presence of the public on the activity levels and the visibility of animals was also species-specific. Only a few species strongly modulated their activity levels in response to the presence of visitors, while most did not or did so only very slightly. Through focused sampling, we detected a clear effect of the lockdown period in geckos housed on-show, with on-show geckos being significantly more visible when the zoo was closed to the public.
Changes in reptile behaviours may be affected by environmental parameters, which could confound the effect of visitor presence [19]. However, the internal environmental parameters of the enclosures were artificially controlled. Secondly, data collection was designed to help control for any effect of extrinsic seasonal cues, such as an indirect photoperiod from external windows. By collecting transect data in both the summer and winter, the seasonal variations were evened out across the observation periods. Moreover, the comparison of tokay gecko data between the off- and on-show exhibits, where any extrinsic seasonal cues were the same but exposure to changes in visitor presence was not, further supports the effect of visitors as a primary driver of changes in animal behaviour. Although this comparison could be made for just one species, there is no reason to expect a different effect for other species.
Broad sampling identified a weaker impact of visitors on the visibility of on-show tokay geckos compared to the other species considered in our study (Figure 3), although OOS reduced during lockdown in this dataset, as well as the high-resolution sampling (Figure 1). Yet, focused sampling using cameras detected that this impact was significant. Indeed, the use of cameras to collect data throughout the day rather than only at four to five snapshot moments may have allowed much more activity to be detected and the effect of visitors more clearly identified. The majority of data for the broad transect dataset for the geckos was OOS, which may have meant that more subtle patterns could be detected with sampling every 10 min. For cryptic taxa like tokay geckos, more sampling effort might be needed to detect the effects: this might have a bearing on the other taxa in this study, such as spiny hill turtles, which were, like the tokay geckos, frequently OOS and where a weaker effect of lockdown was detected. It is important to note that tokay geckos coexist in close proximity to humans in the wild, often dwelling in human buildings. Anecdotal observations (personal observation C.J.M. and K.C.C.) indicated a strong negative response of geckos to humans in the zoo, whereby approaching an enclosure while a gecko was visible led to a rapid retreat of the animal into a refugium. The propensity to coexist with humans may just as easily be due to a higher sensitivity to human presence allowing animals to stay out of sight as to be due to a tolerance for human presence. In either case, our data detected a significant reduction in gecko activity in the presence of human visitors.
These outcomes reflect the diversity of responses to the lockdown identified in a range of taxa by other authors [19,21,28,47]. Common with the only other reptile-based study of the impact of lockdown ([19]; saltwater crocodiles), the majority of reptile species in this study showed only a small, if any, response to changes in visitors’ presence. However, the stronger effects seen in some taxa highlighted that some reptiles may be sensitive to human presence. It is important to note that, like the amphibian study of Boultwood et al. [21], we were unable to collect detailed behavioural data reflecting those collected by some other authors for our species [19,47,48] as a result of resource constraints during lockdown, and therefore, some behavioural changes that did not affect activity or visibility may have been undetected. It is important to note that, although we demonstrated effects of visitors on some species, our data did not allow us to attribute this to positive or negative changes in their welfare state. For example, beaded lizards, which hid less during visitors’ presence, may be looking for interspecific interactions with the visitors and deriving a positive welfare change from this. Alternatively, increased activity may be an escape response to the presence of visitors, which would result in a negative welfare change. In order to better understand the welfare impacts of visitors on specific species, additional behavioural data, potentially combined with a validated stress hormone analysis, would be useful.
The influence of visitor presence depends on the baseline activity of the species; the higher the general baseline activity levels, the stronger the influence of the presence or absence of visitors. This suggests that activity may not be an effective metric for all species or individuals, as less active taxa may respond to positive or negative changes in welfare in other ways. Interestingly, this relationship was not observed in the proportion of out-of-sight individuals: for example, even though the king cobra had the highest baseline visibility of all the species in our study, it was less affected by the presence of visitors than the gidgee skinks or the Chinese crocodile lizards, which generally hid more often.
All enclosures did not have the same proportions of hiding spots, as the exhibits were designed to cater to the best practice husbandry of each species in question within the constraints of the available space, and the size of each exhibit proportionate to the size of animals housed within was not equal. This could explain why beaded lizards, telfair and gidgee skinks had more stable baseline proportions of out-of-sight individuals compared to the other species. This observation could also arise from the unequal number of individuals per species represented, as these three lizard species corresponded to the most numerous populations of animals included in our study. As previously discussed, the context of this case study precluded controlling for these variables.
We could not test for ecological or phylogenetic covariates (e.g., Reference [21]), as our taxon list was opportunistically selected and contained a large number of potential groups (circadian cycle, phylogeny, habitat type, natural diet, group vs. individual housed, geographic origin, etc.), and no clear groupings made sense to investigate over the others. The numbers of visitors have shown significant differences in HAI studies without zoo closures, for example, harbour seals, siamang and white-cheeked gibbons spent an increased amount of time hiding or present in off-show areas when visitor numbers were higher than normal [49]. Logistical constraints and privacy laws, which prohibited the use of recording equipment to count visitors, meant that we were unable to include visitor numbers in our models. Ticket sales were not an accurate representation of the number of visitors viewing individual exhibits, which was affected by many factors [50], and so we elected not to use this as a variable [51]. Finally, as our data were collected by staff observers and cameras requiring light to function, the observations were limited to the daytime. Although this was aligned with the parts of the day that visitors would also be present, spill-over effects of human presences may exist. Extension of the observations into the crepuscular and nocturnal periods may have detected such behavioural changes in species that are active at these times, and this should be a focus of future works in this field.
5. Conclusions
Like other visitor effect studies carried out during the COVID-19 pandemic, the results of this study provided evidence of the influence of visitors’ presence on the behaviour of captive reptiles, forming a foundation for further expansion in visitor impact research in this class.
We showed that captive reptiles differ in their baseline activity and hiding levels. Moreover, our results suggested that their behaviour can be differentially influenced by the presence of visitors, with species-specific responses varying from increases in activity or visibility levels (respectively, in snake-necked turtle and beaded lizards) to a reduction in these same behaviours in other species (respectively, in blue tree monitor and Chinese crocodile lizards), while many species seem to be unaffected (like pythons, for example). For cryptic species, rapid scan sampling approaches may be limited to detect differences (as discussed regarding the tokay geckos in this study), and for species with naturally low activity levels, the selection of different behaviours for assessment may be necessary. Overall, our case study demonstrated the variety of reptile responses to visitor presences and highlighted the dearth of research in this area. Our work reinforced the need for further species-specific research on welfare in captive reptiles and for the development of refined protocols dictating the interactions of different taxa with humans, as well as for the development of terrariums allowing species to respond appropriately to human presences.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/jzbg2040048/s1, Table S1: Model selection: Results of log-likelihood ratio tests (LRTs) comparing the full model to nested models after deletion of the random slope, and AIC comparison between the fixed effects structures. Models with ΔAIC < 2 were considered equivalent, so the final model chosen (marked by *) was the most parsimonious.
Author Contributions
Conceptualisation C.J.M., K.C.C., L.J.R., I.A.T.K. and L.M.C.; methodology, C.J.M., K.C.C., L.M.C. and L.J.R.; formal analysis, C.J.M. and L.F.-M.; investigation, C.J.M., K.C.C., L.J.R., I.A.T.K., L.F.-M. and L.M.C.; data curation, L.F.-M.; writing—original draft preparation, C.J.M., K.C.C., L.J.R., I.A.T.K., L.F.-M. and L.M.C.; writing—review and editing C.J.M., K.C.C., L.J.R., I.A.T.K., L.F.-M. and L.M.C. and supervision, C.J.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Ethical review and approval were waived for this study, as the methods were purely observational and did not entail any experimental changes to the animals’ management or well-being.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are available at https://github.com/LeaFieschiMeric/lockdown_on_reptiles (accessed on 21 October 2021).
Acknowledgments
The authors would like to thank all members (Ben Tapley, Daniel Kane, Francesca Servini, Unnar Aevarsson, Charlotte Ellis and Joe Capon) of the Herpetology Section at ZSL London Zoo for assistance in collecting the data and for continuous high-quality animal care undertaken under extremely difficult circumstances; to Ben Tapley for commenting on the MS; to Ella Meekins for helping with data input and to Julia Reis (Roehampton University) for her support for the project.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Mason, P. Roles of the modern zoo: Conflicting or complementary? Tour. Rev. Int. 2007, 11, 251–263. [Google Scholar] [CrossRef]
- Tribe, A.; Booth, R. Assessing the role of zoos in wildlife conservation. Hum. Dimen. Wildl. 2003, 8, 65–74. [Google Scholar] [CrossRef]
- Carr, N.; Cohen, S. The public face of zoos: Images of entertainment, education and conservation. Anthrozoös 2011, 24, 175–189. [Google Scholar] [CrossRef] [Green Version]
- Sherwen, S.; Hemsworth, P. The visitor effect on zoo animals: Implications and opportunities for zoo animal welfare. Animals 2019, 9, 366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ward, S.; Sherwen, S.; Clark, F. Advances in applied zoo animal welfare science. J. Appl. Anim. Welf. Sci. 2018, 21, 23–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broom, D.M. Indicators of poor welfare. Brit. Vet. J. 1986, 142, 524–526. [Google Scholar] [CrossRef]
- Whitham, J.; Wielebnowski, N. New directions for zoo animal welfare science. Appl. Anim. Behav. Sci. 2013, 147, 247–260. [Google Scholar] [CrossRef]
- Barber, J. Programmatic approaches to assessing and improving animal welfare in zoos and aquariums. Zoo Biol. 2009, 28, 519–530. [Google Scholar] [CrossRef] [PubMed]
- Hill, S.; Broom, D. Measuring zoo animal welfare: Theory and practice. Zoo Biol. 2009, 28, 531–544. [Google Scholar] [CrossRef] [PubMed]
- Learmonth, M.J. Human–Animal Interactions in Zoos: What Can Compassionate Conservation, Conservation Welfare and Duty of Care Tell Us about the Ethics of Interacting, and Avoiding Unintended Consequences? Animals 2020, 10, 2037. [Google Scholar] [CrossRef]
- Hosey, G. A preliminary model of human–animal relationships in the zoo. Appl. Anim. Behav. Sci. 2008, 109, 105–127. [Google Scholar] [CrossRef] [Green Version]
- Sherwen, S.; Magrath, M.; Butler, K.; Hemsworth, P. Little penguins, Eudyptula minor, show increased avoidance, aggression and vigilance in response to zoo visitors. Appl. Anim. Behav. Sci. 2015, 168, 71–76. [Google Scholar] [CrossRef]
- Eltorai, A.; Sussman, R. The “Visitor effect” and captive black-tailed prairie dog. Zool. Gart. 2010, 79, 109–120. [Google Scholar] [CrossRef]
- Todd, P.; Macdonald, C.; Coleman, D. Visitor-associated variation in captive Diana monkey (Cercopithecus diana diana) behaviour. Appl. Anim. Behav. Sci. 2007, 107, 162–165. [Google Scholar] [CrossRef]
- Paré, J.; Lentini, A. Reptile geriatrics. Vet. Clin. Exot. Anim. Pract. 2010, 13, 15–25. [Google Scholar] [CrossRef]
- Benn, A.; McLelland, D.; Whittaker, A. A review of welfare assessment methods in reptiles, and preliminary application of the welfare quality protocol to the pygmy blue-tongue skink, Tiliqua adelaidensis, using animal-based measures. Animals 2019, 9, 27. [Google Scholar] [CrossRef] [Green Version]
- Morgan, K.; Tromborg, C. Sources of stress in captivity. Appl. Anim. Behav. Sci. 2007, 102, 262–302. [Google Scholar] [CrossRef]
- Gillingham, J. Normal behaviour. In Health and Welfare of Captive Reptiles; Warwick, C., Frye, F., Murphy, B., Eds.; Chapman & Hall/Kluwer: London, UK, 2004; pp. 131–164. [Google Scholar]
- Riley, A.; Terry, M.; Freeman, H.; Alba, A.; Soltis, J.; Leeds, A. Evaluating the effect of visitor presence on Nile crocodile (Crocodylus niloticus) behavior. J. Zool. Bot. Gard. 2021, 2, 115–129. [Google Scholar] [CrossRef]
- Kane, D.; Davis, A.C.; Michaels, C.J. Play behaviour by captive tree monitors, Varanus macraei and Varanus prasinus. Herpetol. Bull. 2019, 149, 28–31. [Google Scholar] [CrossRef]
- Boultwood, J.; O’Brien, M.; Rose, P. Bold Frogs or Shy Toads? How did the COVID-19 closure of zoological organisations affect amphibian activity? Animals 2021, 11, 1982. [Google Scholar] [CrossRef]
- Chelodina mccordi. The IUCN Red List of Threatened Species. 2019. Available online: https://www.iucnredlist.org/species/123814489/123814575 (accessed on 9 August 2021).
- Egernia stokesii. The IUCN Red List of Threatened Species. Available online: https://www.iucnredlist.org/species/62246/101743684 (accessed on 19 October 2021).
- Gekko gecko. The IUCN Red List of Threatened Species. 2019. Available online: https://www.iucnredlist.org/species/195309/2378260 (accessed on 13 October 2021).
- O’Shea, M. Lizards of the World: A Guide to Every Family; Ivy Press: London, UK, 2021; pp. 200–234. [Google Scholar]
- Goetz, M. Husbandry and breeding of the spiny turtle Heosemys spinosa (GRAY, 1931) at the Durrell Wildlife Conservation Trust. Radiata 2007, 16, 2–15. [Google Scholar]
- Lampropeltis triangulum. The IUCN Red List of Threatened Species. 2019. Available online: https://www.iucnredlist.org/species/197493/2490171 (accessed on 19 October 2021).
- Leiolopisma telfairii. The IUCN Red List of Threatened Species. 2018. Available online: https://www.iucnredlist.org/species/11409/152276731 (accessed on 19 October 2021).
- Malayopython reticulatus. The IUCN Red List of Threatened Species. 2018. Available online: https://www.iucnredlist.org/species/183151/1730027 (accessed on 19 October 2021).
- Morelia spilota. The IUCN Red List of Threatened Species. 2017. Available online: https://www.iucnredlist.org/species/62232/21649539 (accessed on 19 October 2021).
- Ophiophagus hannah. The IUCN Red List of Threatened Species. 2012. Available online: https://www.iucnredlist.org/species/177540/1491874 (accessed on 19 October 2021).
- Oxyuranus microlepidotus. The IUCN Red List of Threatened Species 2018. Available online: https://www.iucnredlist.org/species/42493150/42493160 (accessed on 13 October 2021).
- Huang, H.; Wang, H.; Li, L.; Wu, Z.; Chen, J. Genetic diversity and population demography of the Chinese crocodile lizard (Shinisaurus crocodilurus) in China. PLoS ONE 2014, 9, e91570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennett, D. International trade in the blue tree monitor lizard Varanus macraei. Biawak J. Varanid Biol. Husb. 2015, 9, 50–57. [Google Scholar]
- Bolker, B.M.; Brooks, M.E.; Clark, C.J.; Geange, S.W.; Poulsen, J.R.; Stevens, M.H.H.; White, J.S.S. Generalized linear mixed models: A practical guide for ecology and evolution. Trends Ecol. Evol. 2009, 24, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Millar, R.B.; Anderson, M.J. Remedies for pseudoreplication. Fish. Res. 2004, 70, 397–407. [Google Scholar] [CrossRef]
- Harrison, X.A.; Donaldson, L.; Correa-Cano, M.E.; Evans, J.; Fisher, D.N.; Goodwin, C.E.; Robinson, B.S.; Hodgson, D.J.; Inger, R. A brief introduction to mixed effects modelling and multi-model inference in ecology. PeerJ 2018, 6, e4794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, P.C. Extension of Nakagawa & Schielzeth’s R2GLMM to random slopes models. Methods Ecol. Evol. 2014, 5, 944–946. [Google Scholar] [PubMed] [Green Version]
- Raudenbush, S.W.; Yang, M.L.; Yosef, M. Maximum likelihood for generalized linear models with nested random effects via high-order, multivariate Laplace approximation. J. Comput. Graph. Stat. 2000, 9, 141–157. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Soft. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Mu-MIn: Multi-Model Inference. R Package Version 1.43.17. 2020. Available online: http://R-Forge.R-project.org/projects/mumin/ (accessed on 17 June 2021).
- Sarkar, D. Lattice: Multivariate Data Visualization with R; Springer: New York, NY, USA, 2008; Available online: http://lmdvr.r-forge.r-project.org (accessed on 17 June 2021).
- The R Foundation. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: http://www.R-project.org/ (accessed on 17 June 2021).
- Dugard, P.; File, P.; Todman, J.; Todman, J.B. Single-Case and Small-n Experimental Designs: A Practical Guide To Randomization Tests; Routledge: New York, NY, USA, 2012; pp. 11–130. [Google Scholar]
- Tanious, R.; Onghena, P. Randomized Single-Case Experimental Designs in Healthcare Research: What, Why, and How? Healthcare 2019, 7, 143–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michaels, C.J.; Gini, B.F.; Clifforde, L. A persistent abnormal repetitive behaviour in a false water cobra (Hydrodynastes gigas). Anim. Welf. 2020, 29, 371–378. [Google Scholar] [CrossRef]
- Williams, E.; Carter, A.; Rendle, J.; Ward, S. Impacts of COVID-19 on animals in zoos: A longitudinal multi-species analysis. J. Zool. Bot. Gard. 2021, 2, 130–145. [Google Scholar] [CrossRef]
- Williams, E.; Carter, A.; Rendle, J.; Ward, S. Understanding impacts of zoo visitors: Quantifying behavioural changes of two popular zoo species during COVID-19 closures. Appl. Anim. Behav. Sci. 2021, 236, 105253. [Google Scholar] [CrossRef]
- Stevens, J.; Thyssen, A.; Laevens, H.; Vervaecke, H. The influence of zoo visitor numbers on the behaviour of harbour seals. J. Zoo Aquar. Res. 2013, 1, 31–34. [Google Scholar] [CrossRef]
- Marcellini, D.L.; Jenssen, T.A. Visitor behavior in the National Zoo’s reptile house. Zoo Biol. 1988, 7, 329–338. [Google Scholar] [CrossRef]
- Bonnie, K.E.; Ang, M.Y.; Ross, S.R. Effects of crowd size on exhibit use by and behavior of chimpanzees (Pan troglodytes) and Western lowland gorillas (Gorilla gorilla) at a zoo. Appl. Anim. Behav. Sci. 2016, 178, 102–110. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).