1. Introduction
The poor nutritional quality of feeder insects is frequently cited as a challenge to the successful maintenance of insectivorous vertebrates in captivity and the root cause of a number of common pathologies found in this context [
1,
2,
3]. Arguably chief among the nutrients of concern is calcium, a deficiency of which is responsible for metabolic pathologies in vertebrates. Among these pathologies, nutritional secondary hyperparathyroidism is responsible for particularly notable morbidity and mortality in captive insectivores [
2]. Calcium is typically deficient in commonly used feeder invertebrates [
1] and may be augmented through ‘gut loading’ with high calcium feed before being offered to predators [
4]. Calcium metabolism in vertebrates is to varying extents dependent on vitamin D
3 and its derivatives, which may be obtained from both diet and endogenous cutaneous photobiosynthesis during exposure to UVB radiation (wavelength 280–320 nm) in combination with heat [
5]. To this end, Oonincx et al. [
6] recently demonstrated that some invertebrate species routinely used as feeder items could be vitamin D
3 enriched through exposure to UVB radiation. Although this is a useful tool for improving nutritional quality of captive insectivore diets, the species selection and duration of UVB exposure used in that study do not necessarily align with the logistic needs of institutions working with insectivores in captivity. We trialed UVB exposure over a shorter time period designed to be more in keeping with the typical time available to enhance nutritional quality in captive husbandry settings, using black field crickets (
Gryllus bimaculatus), a species identified as the most amenable to other forms of nutritional enhancement [
7]. We also tested for any impact of UVB exposure on calcium accumulation, given that in vertebrates UVB exposure improves calcium accumulation via the photobiosynthesis of vitamin D
3 [
2]. Additionally, we quantified the loss of calcium from the guts of crickets after the high calcium gut load diet was removed from the insects in order to better understand the changes in nutritional quality of insects between being offered to predators and actually being consumed.
2. Materials and Methods
All experimental work was carried out at ZSL London Zoo.
Adult (final instar) Gryllus bimaculatus were purchased from Peregrine Live Foods (Magdalen Laver, Ongar, UK) and housed in groups of 75 in twenty 360 × 210 × 160 mm plastic enclosures lined with egg cartons at 25.5–27.5 °C and relative humidity of 45–52%. Water was continuously available, either by provision of a soaked paper towel whist undergoing fasting or mixed in with the feed.
Enclosures were assigned to one of two treatments; UVB+ and UVB−. All enclosures were illuminated by T5 Ultraviolet B lamps (54W, D3+ T5 12%, Arcadia, UK) with reflectors on a photoperiod of 12:12 (light:dark). Mylar film, which absorbs all UV-radiation below 320 nm [
8], was used to cover a portion of each lamp so that half of all enclosures (
n = 10) received no UVB radiation (UVB−) and the other half (
n = 10) received UVB radiation (UVB+), but each lamp illuminated some of both treatments, thereby reducing variation between lamps. Readings were taken at cricket level on top of the egg carton using a solar meter model 6.5 (UV index) and a solar meter model 6.2 (UVB) (both Solar Light Company, Glendale, PA, USA) to confirm irradiation levels 30 min after lamps were turned on (Baines et al., 2016). Crickets were exposed to a mean (SD) UVi of 9.5 (0.67), corresponding to a total irradiance of 292.8(120.9) uW/cm
2. Crickets were able to shelter underneath the cardboard with no UVB exposure but were routinely observed exposed and apparently basking under the lamps. Control enclosures were confirmed to receive no UVB radiation (UVi = 0(0) and 0(0) uW/cm2).
Crickets were fasted for 24 h to clear the gastrointestinal tract (1) with UVB lighting turned off. Lighting was then turned on and crickets were offered ad libitum a commercial gut loading diet (Nutrogrub (Vet Ark Professional, Winchester, Hampshire, UK)) containing 6.5% dry mass (DM) calcium for 48 h. The diet was made into a paste by mixing with reverse osmosis (RO) water and placed in small plastic trays. Crickets were then fasted again for 24 h to simulate time spent in a predator’s enclosure without access to food.
Baseline samples were taken after being fasted and without UVB irradiation for 24 h. This was termed 0 h and marks the start of the experiment. Sample times were then taken from the point at which food was offered to crickets and lighting was turned on (24 h). Crickets were sampled from every container (
n = 10 per treatment; 1 male and 1 female cricket per sample) for mineral analysis (calcium and phosphorus) at 0, 24, 48, and 72 h. For cost reasons, crickets were sampled for vitamin D
3 analysis less frequently; samples (35 male and 35 female crickets) were taken from three random enclosures for each treatment at 0 and 48 h. Equal numbers of males and females were used in each sample to avoid sex associated bias in gut loading and/or irradiation responses. Larger numbers of crickets were sampled for vitamin D
3 analysis due to the higher mass requirements of this process. Euthanasia of sampled crickets was conducted immediately upon sampling following Jayson et al. [
9].
Samples for mineral analysis were shipped on dry ice to Manchester Metropolitan University (UK) following the methods described by Michaels et al. [
3]. Samples for vitamin analysis, where one sample comprised 70 crickets, were sent on dry ice to Sciantec Analytical Services (Stockbridge Technology Centre 16, Selby, North Yorkshire, UK) to be analysed via high performance liquid chromatography (HPLC), SOP Number: S1181.
Data were analysed on a dry matter content (DMC) basis. All data were normally distributed (Shapiro–Wilks test) and had homogenous variance (Levene’s F test).
We used general linear models (GLM) to analyse each mineral in turn. Initially, we used a fully factorial model using type 1 sums of squares including UVB treatment and sample time as factors. If the interaction term was not significant, we removed this and ran a model including only the main effects using type 3 sums of squares.
Post hoc least squares difference tests were used to compare time points after GLMs were performed in the case of a significant effect of time.
Vitamin D3 results were below the detectable limit in all samples (<0.50 IU/g) and so no data were analysed for this parameter.
3. Results
Vitamin D3 was undetectable in all samples and so there was no detectable effect of UVB irradiation on this nutrient in the crickets.
GLM Interaction terms were not significant for calcium and phosphorus and so were excluded from models.
UVB treatment had no significant effect on calcium levels (F
1,75 = 0.299,
p = 0.6), but there was a significant effect of time (F
1,75 = 13.8,
p < 0.001). Calcium was significantly higher at 24 h and 48 h than at 0 h (
p < 0.001;
p < 0.001, respectively) and at 72 h (
p < 0.001;
p < 0.001, respectively), but there was no difference between 24 and 48 h (
p = 0.21) or between 0 and 72 h (
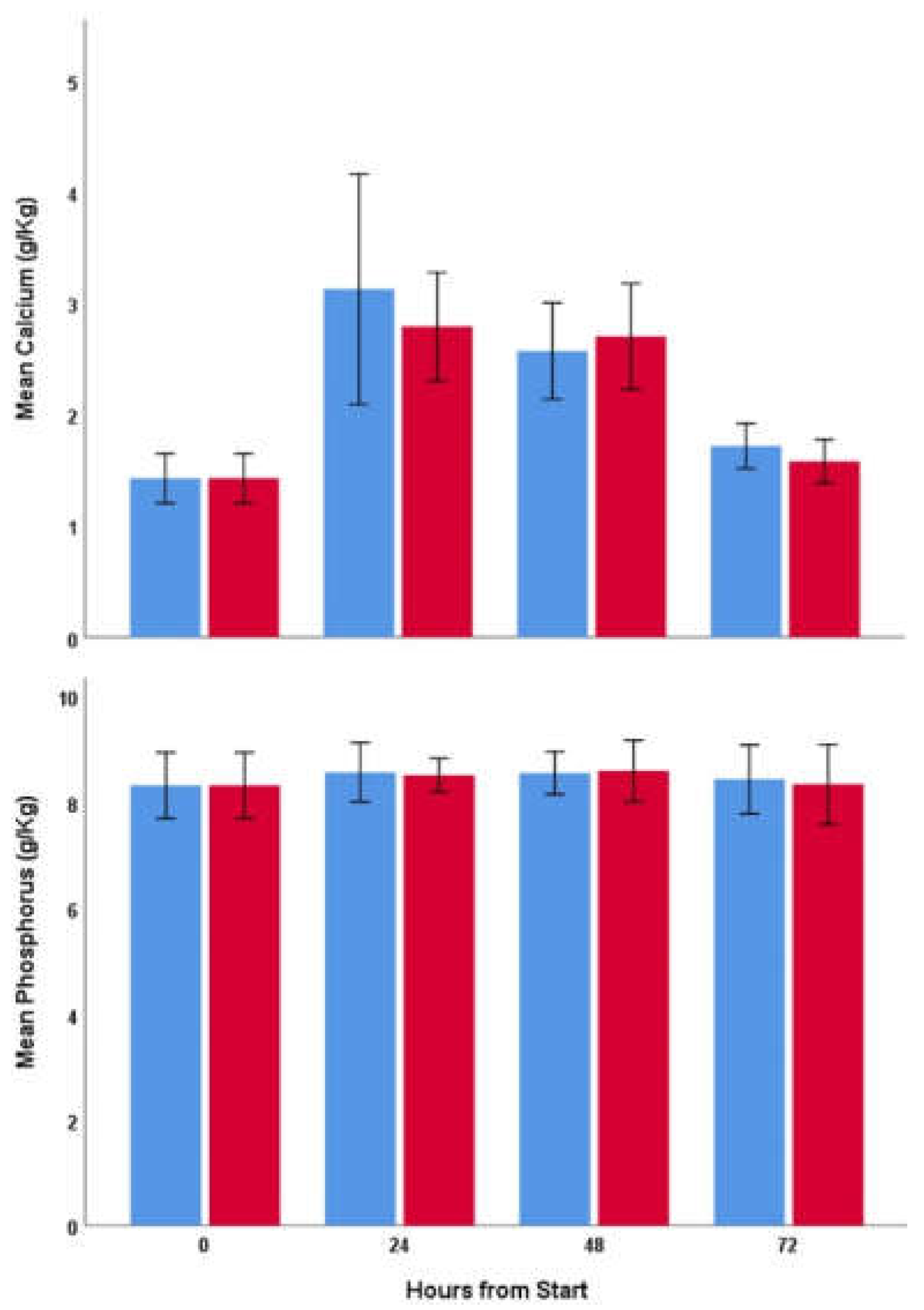
p = 0.48). Mean (SD) calcium content was 1.429 (0.353), 2.962 (1.248), 2.639 (0.702), and 1.650 (0.314) g/Kg at 0, 24, 48, and 72 h, respectively (
Figure 1, top). Neither time nor UVB treatment had significant effects on phosphorus (F
1,75 = 0.365,
p = 0.8; F
1,75 = 0.019,
p = 0.9, respectively;
Figure 1, bottom). Mean (SD) phosphorus content of samples was 8.480 (0.871) g/kg.
4. Discussion
Oonincx et al. [
6] found evidence of vitamin D
3 synthesis in house crickets (
Acheta domestica) after 31 days’ exposure to UVI 3.5. They found a baseline vitamin D
3 content of 41 IU/kg with a tripling to around 120 IU/kg. We found that unirradiated
G. bimaculatus have no detectable Vitamin D
3 content (that is, below 0.50 IU/g), and that irradiation with UVB light also does not potentiate its synthesis, or at least the levels were still undetectable. Even if some increase had occurred below detectable levels, this is meaningless in practical terms as the concentration is too low to have physiological significance. If UVB irradiation did not increase vitamin D3 levels, rather than any change being below the minimum detection threshold, this result may be due to the shorter exposure period in our study (48 h vs. 31 days). However, Oonincx et al. [
6] found higher accumulation of vitamin D
3 with higher levels of UVB irradiation. We exposed crickets to a UVI of approximately 9.5, which was markedly more intense than Oonincx et al. [
6], who used a UVI of up to 3.5 under a similar temperature regimen. Moreover, mammals synthesise vitamin D
3 rapidly in response to UVB exposure, with substantial synthesis taking place over hours to days [
10]. With this in mind, it is surprising that no vitamin D
3 synthesis was detectable in our crickets after a 48 h period of UVB irradiation comprising 24 h of actual exposure. Oonincx et al. [
6] found substantial variation between species of insect in the effect of UVB irradiation, with some species not responding at all. It is possible that
G. bimaculatus is such a species; its black melanin-heavy colouration, which is paired with a tendency to be active in intense sunlight in nature, may contribute to this as it may shield underlying tissues from irradiation.
The life-stage of insects when they are exposed to UV radiation may be important, and our study differed from Oonincx et al. (2018) [
3] in using adult rather than fourth instar nymphs, which may have differing physiological processes. Further work investigating both the duration and life-stage of exposure is urged.
Finally, the insects in this study were able to avoid UVB radiation if they so chose by hiding underneath the egg cartons in their enclosures. Our study enclosures were designed to replicate the conditions under which crickets are typically maintained in order to minimize mortality and to facilitate harvesting and cleaning. Our data are therefore relevant to actual husbandry practice, which was the primary goal of the study, rather than addressing theoretical questions. Nevertheless, G. bimaculatus are drawn to warm, bright conditions and the majority of insects were noted actively exposing themselves to lighting in both treatments, and therefore incidentally to UVB radiation in the experimental treatment. Ultimately, too little is currently understood about vitamin D3 photobiosynthesis in insects to confidently explain our data; they remain useful to guide husbandry practice, however.
Calcium accumulation (
Figure 1, top) in our crickets was similar to that found in other studies using quasi-equal dietary calcium content, with a Ca:P ratio increasing from 0.17 at 0 h to a peak of c. 0.4 after 24–48 h [
11,
12,
13,
14]. This corresponded to a mean (SD) peak calcium content of 2.801(1.018) g/kg compared with 1.429(0.353) g/kg at 0 h, and no change in phosphorus content (
Figure 1, bottom). Neither the peak calcium content, nor the peak Ca:P ratio of 0.4, are sufficient to maintain healthy calcium metabolism in vertebrates (4–12 g/kg DM and 2:1, respectively [
15,
16]); this and the relatively rapid loss of nutritional quality following fasting highlight the importance of combining or replacing gut loading of calcium with supplementary dusting [
3] and ensuring that prey items are consumed rapidly by insectivores.
The loss of calcium content resulting from insects voiding gut contents once food is removed, as is the case when they are placed in an insectivore’s enclosure and are not immediately eaten, has not been reported before in the literature. This is important as many insectivores do not feed immediately and so the actual calcium content ingested may not equate to the calcium content of freshly gut loaded insects. We found a rapid loss of calcium but no change in phosphorus after crickets were fasted (
Figure 1), with neither Ca:P ratio nor calcium content being significantly different from pre-gut loaded levels after 24 h of fasting. This loss of calcium content is not dissimilar to the same phenomenon when insects are dusted with mineral powder [
3], and shows that neither method provides long lasting calcium augmentation, at least in
G. bimaculatus.
Phosphorus content remained comparatively high, similar to data presented by other authors [
1,
3]. As it was not presented in the diet, and as fasting periods were only long enough to influence gut contents rather than tissue composition, phosphorus content was unaffected by either treatment or their interaction.
We hypothesised that if UVB irradiation increased vitamin D3 content of feeder insects, then this may lead to a more rapid accumulation of calcium as vitamin D3 may play a similar role in active calcium uptake from the gut in crickets as it does in vertebrates. As our study animals did not increase vitamin D3 content in the experiment, we were unable to test this hypothesis.
Our experimental design was specifically intended to apply theory to a practical situation found within institutions maintaining collections of insectivores. Oonincx et al. [
6] reported vitamin D
3 increases in insects maintained for relatively long periods of time under UVB lighting, while in most institutions holding live insects as prey items, nutritional improvement is usually undertaken in 24–48 h in order to ensure a constant supply of insects from the producer to the insectivores in question. Ogilvy et al. [
7] demonstrated that
G. bimaculatus crickets outperformed other cricket species in terms of gut loading capabilities, and so this species is often used as an important dietary component in preference to other cricket taxa. Our study was therefore designed to test whether UVB illumination over a practicable timescale of a prey species already known to be superior for nutritional improvement yielded nutritional benefits. Our data suggest that this is not the case and that for this prey species, daytime UVB illumination for 48 h is not a useful addition to standard nutritional improvement protocols. We strongly encourage, nevertheless, the repeat of the study with other prey insect taxa.



{kind=link}
