A Global Survey of Current Zoo Housing and Husbandry Practices for Fossa: A Preliminary Review



Abstract
:1. Introduction
2. Materials and Methods
2.1. Survey Participants and Selection
2.2. Survey Questionnaire
2.3. Data Analysis
2.3.1. Demographic and Population Characteristics
2.3.2. Reports of Unnatural Behaviour from the Survey Responses
2.3.3. Reports of Reproductive Success from the Survey Responses
3. Results
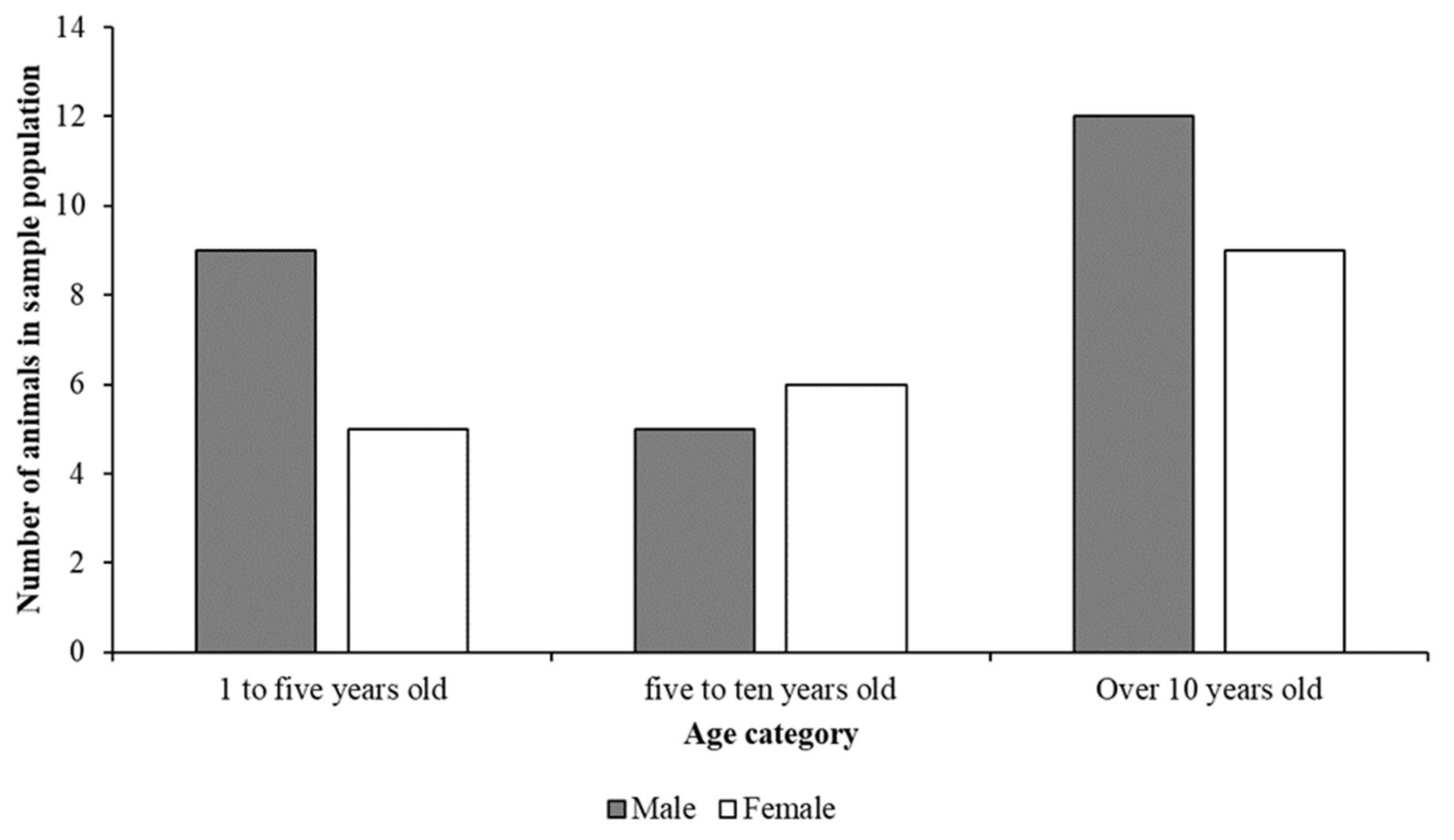
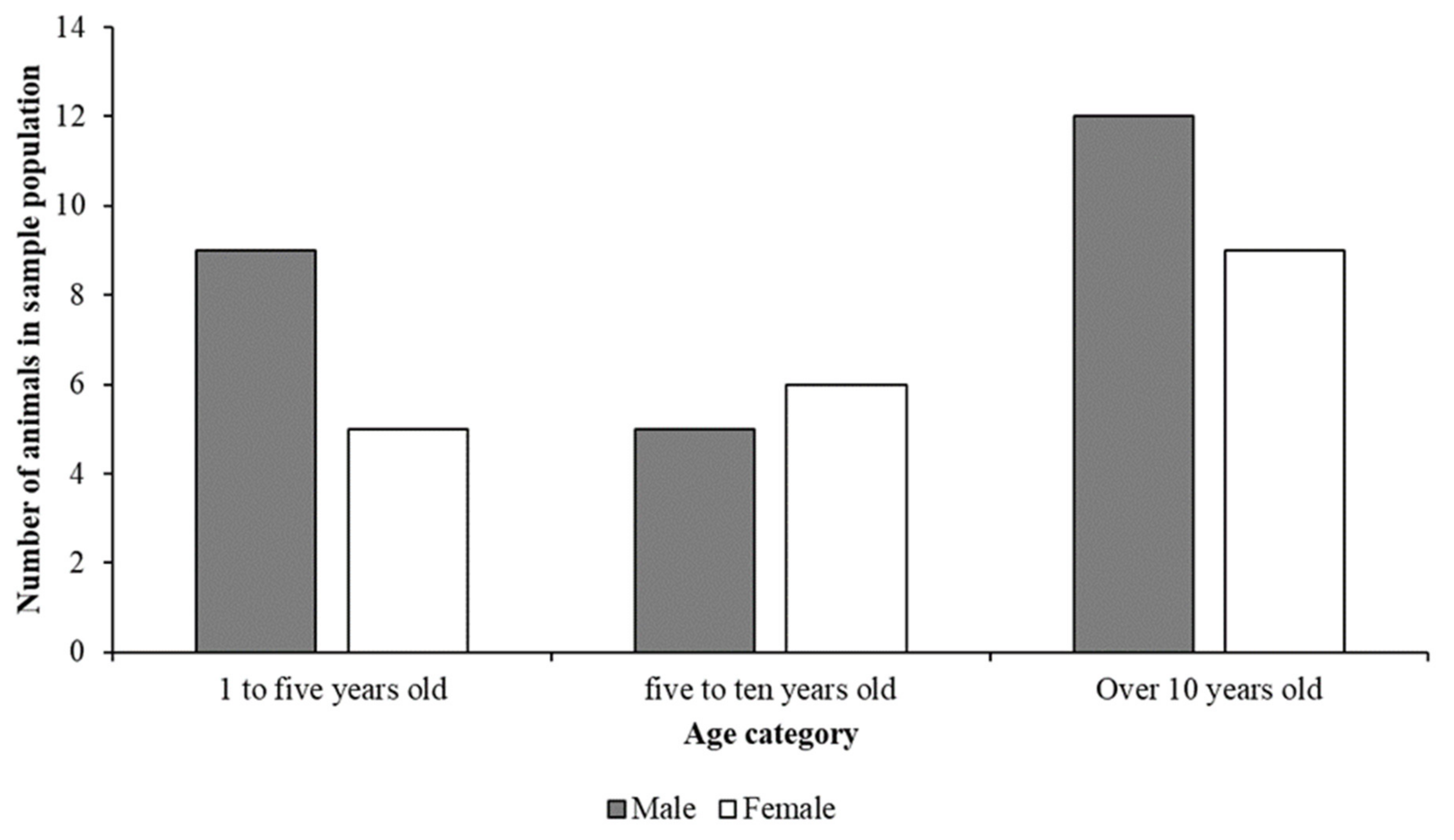
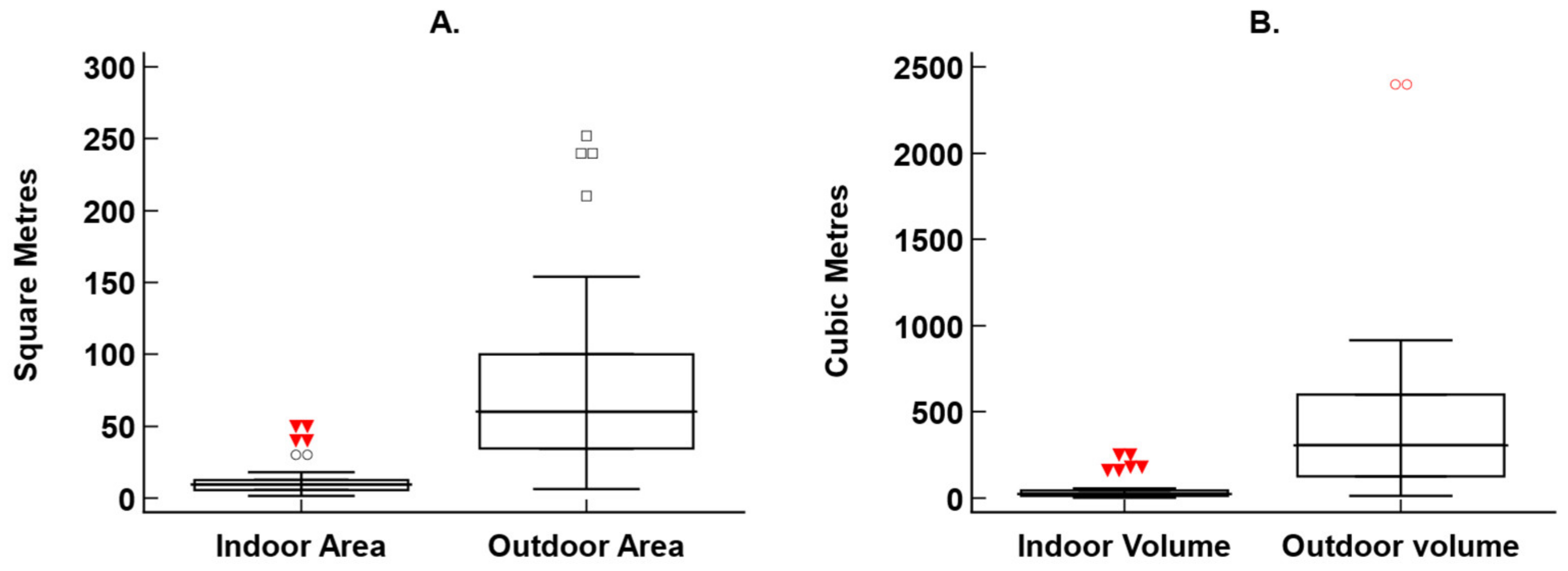
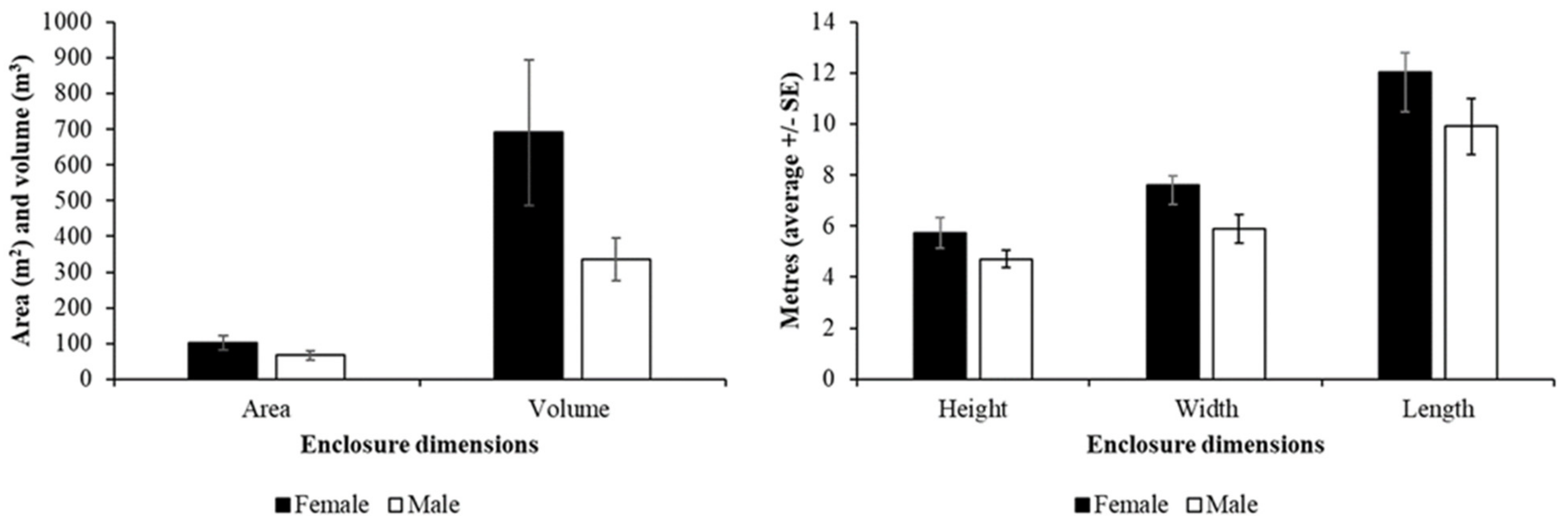
3.1. Demographic and Population Information
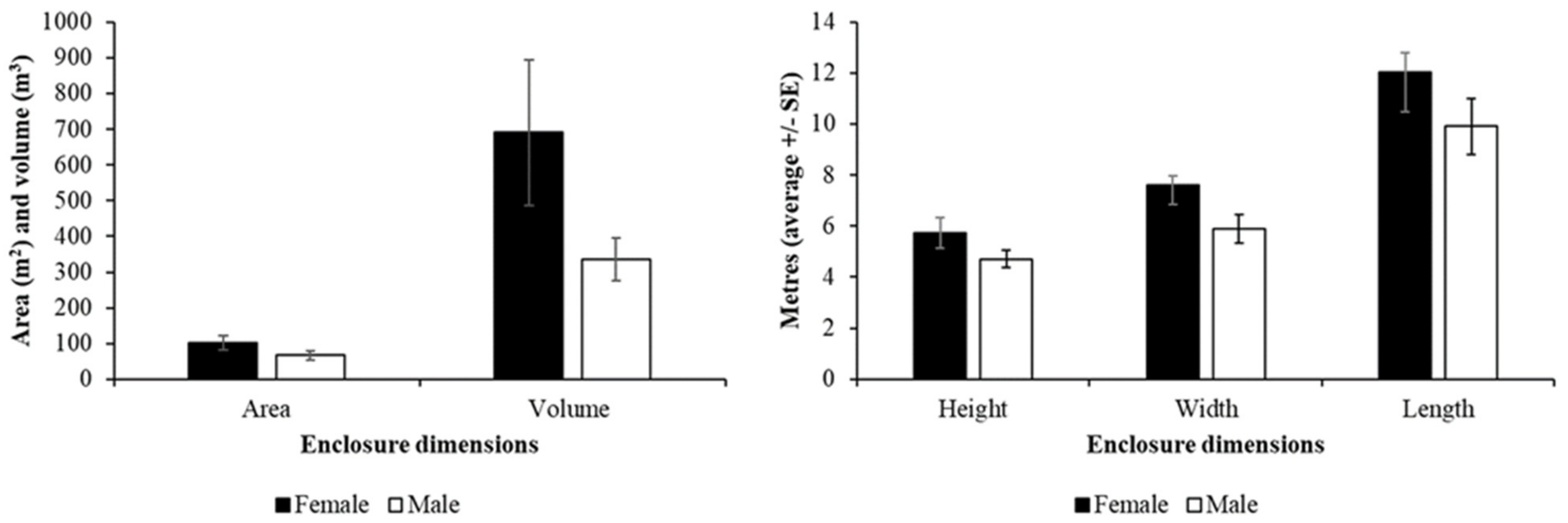
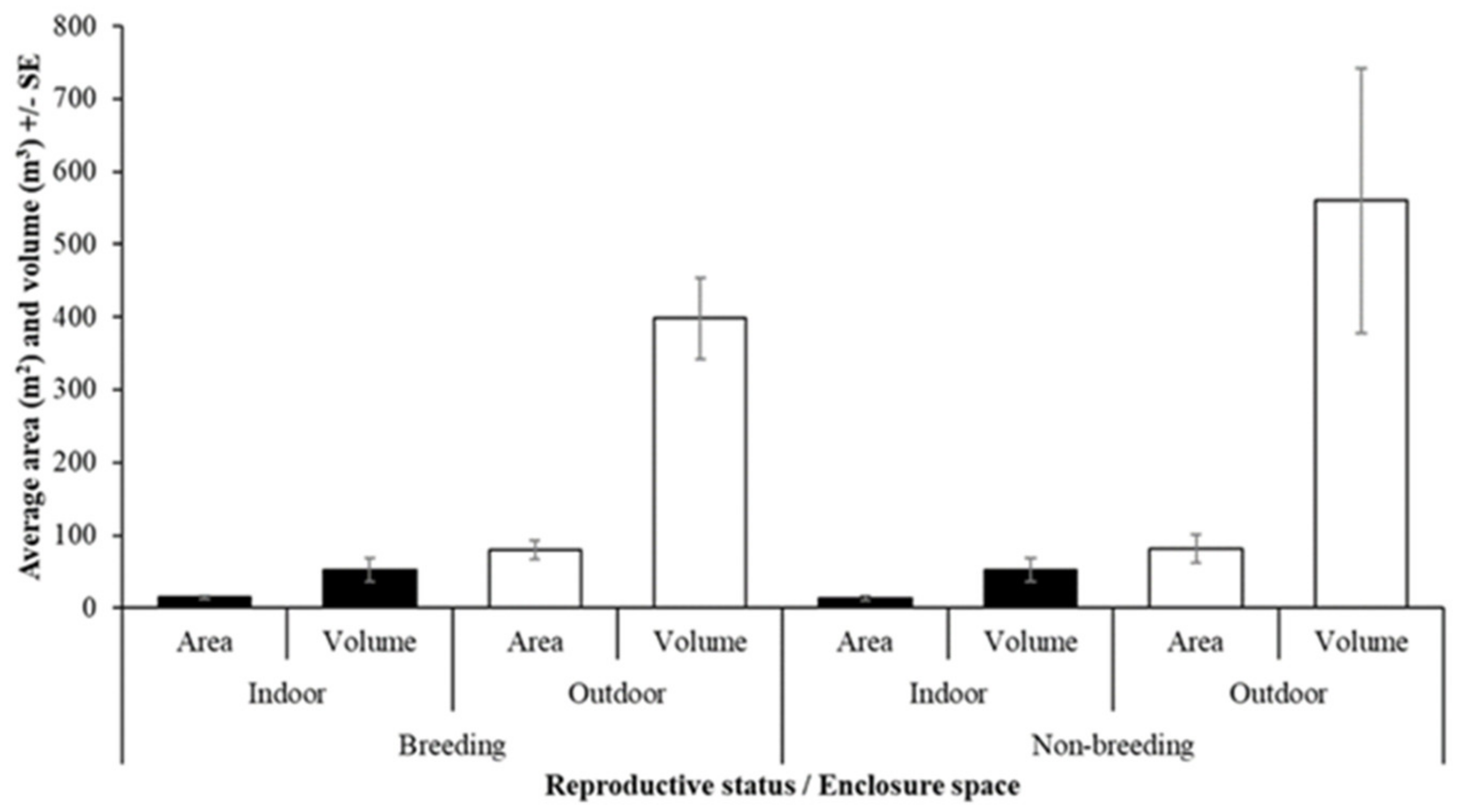
Fossa Housing and Husbandry
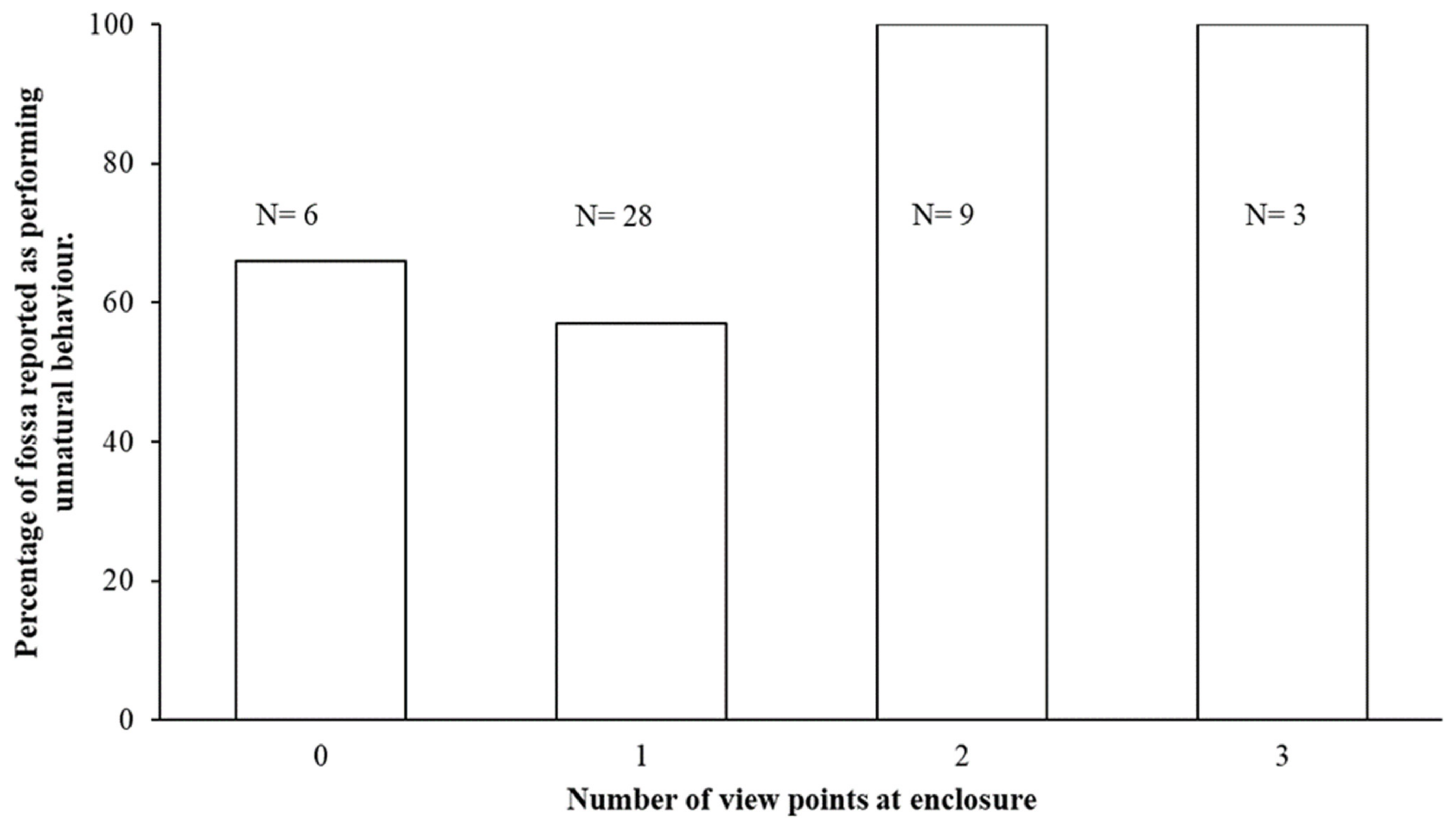
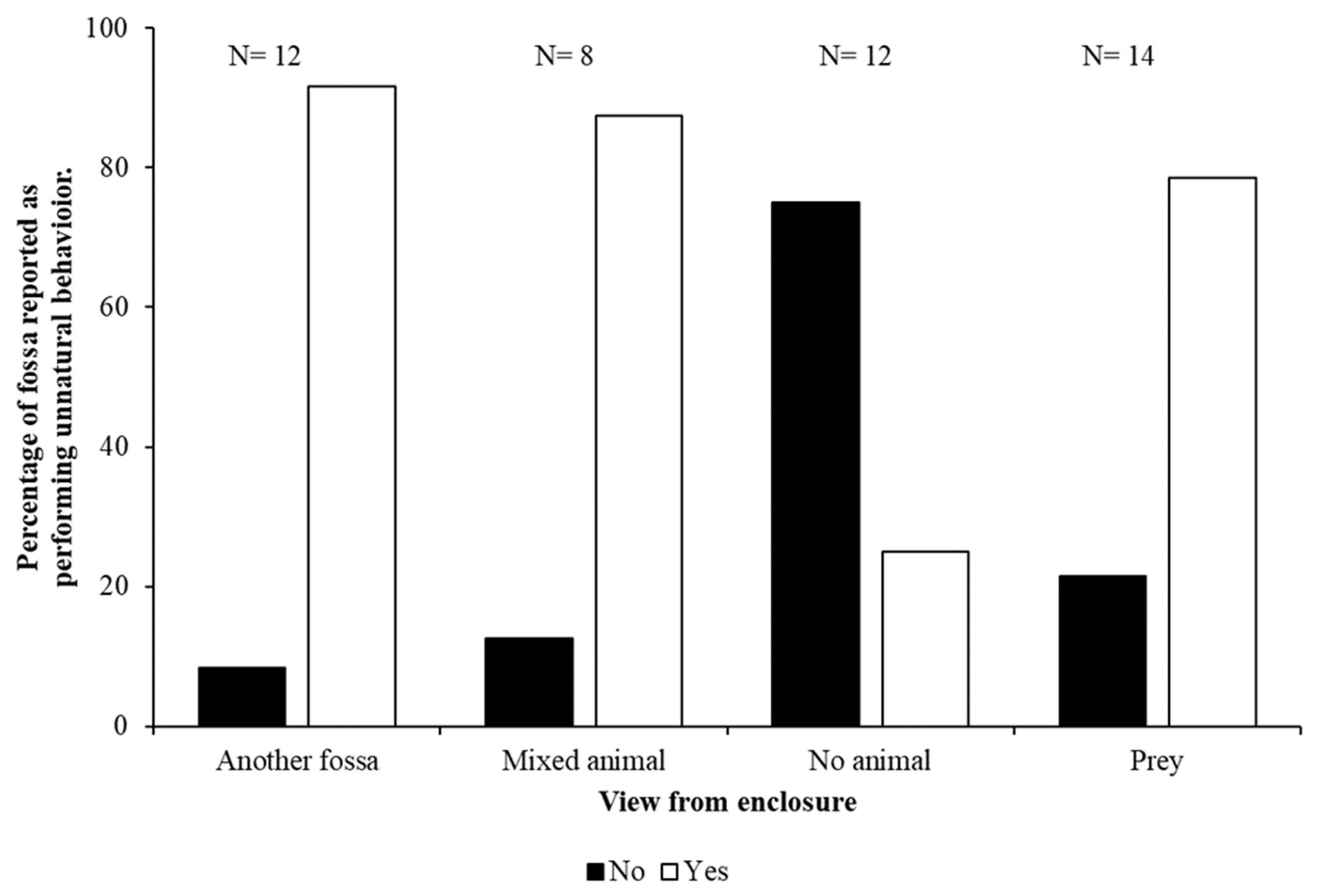
3.2. Reports of Unnatural Behaviour
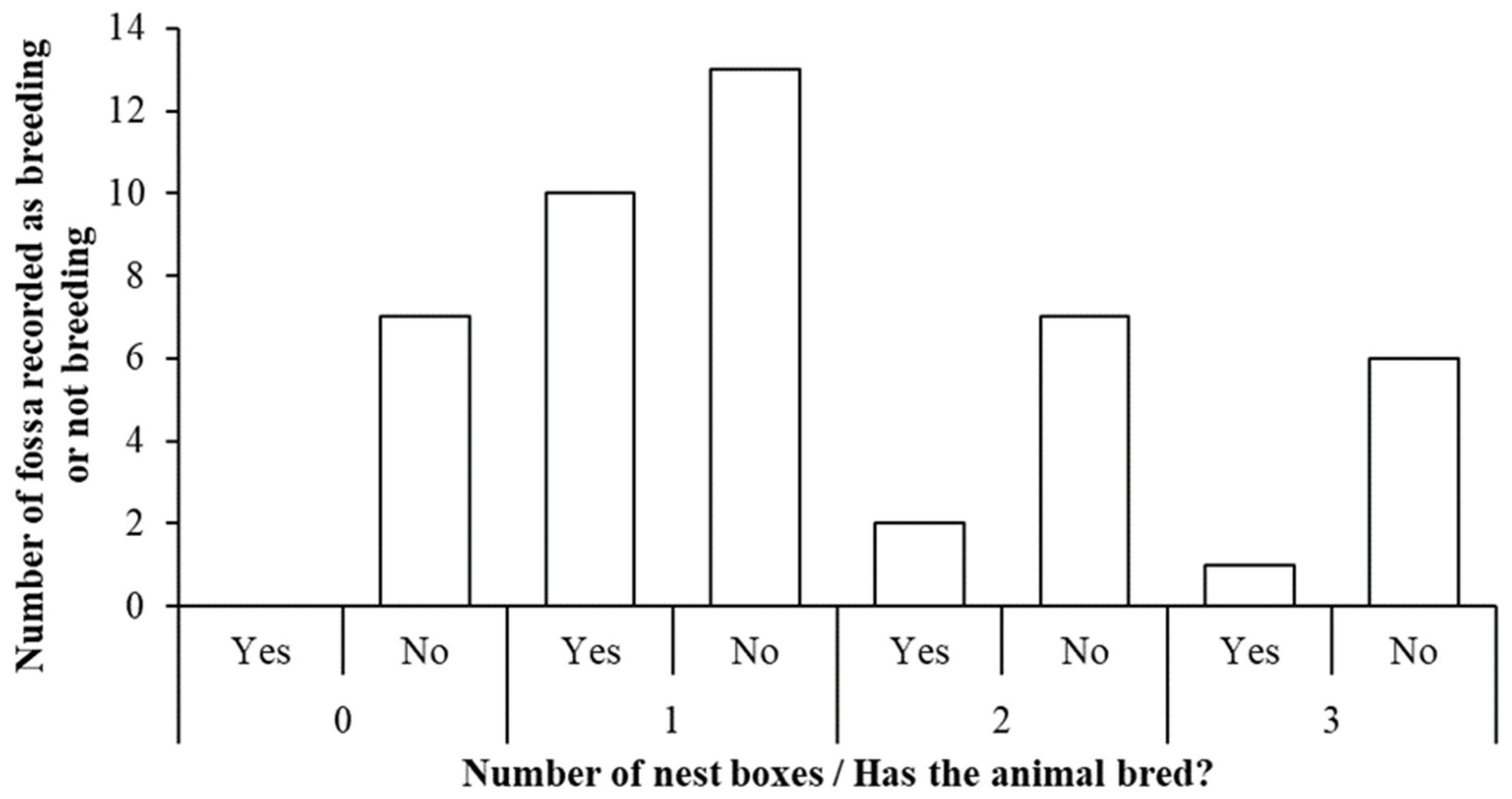
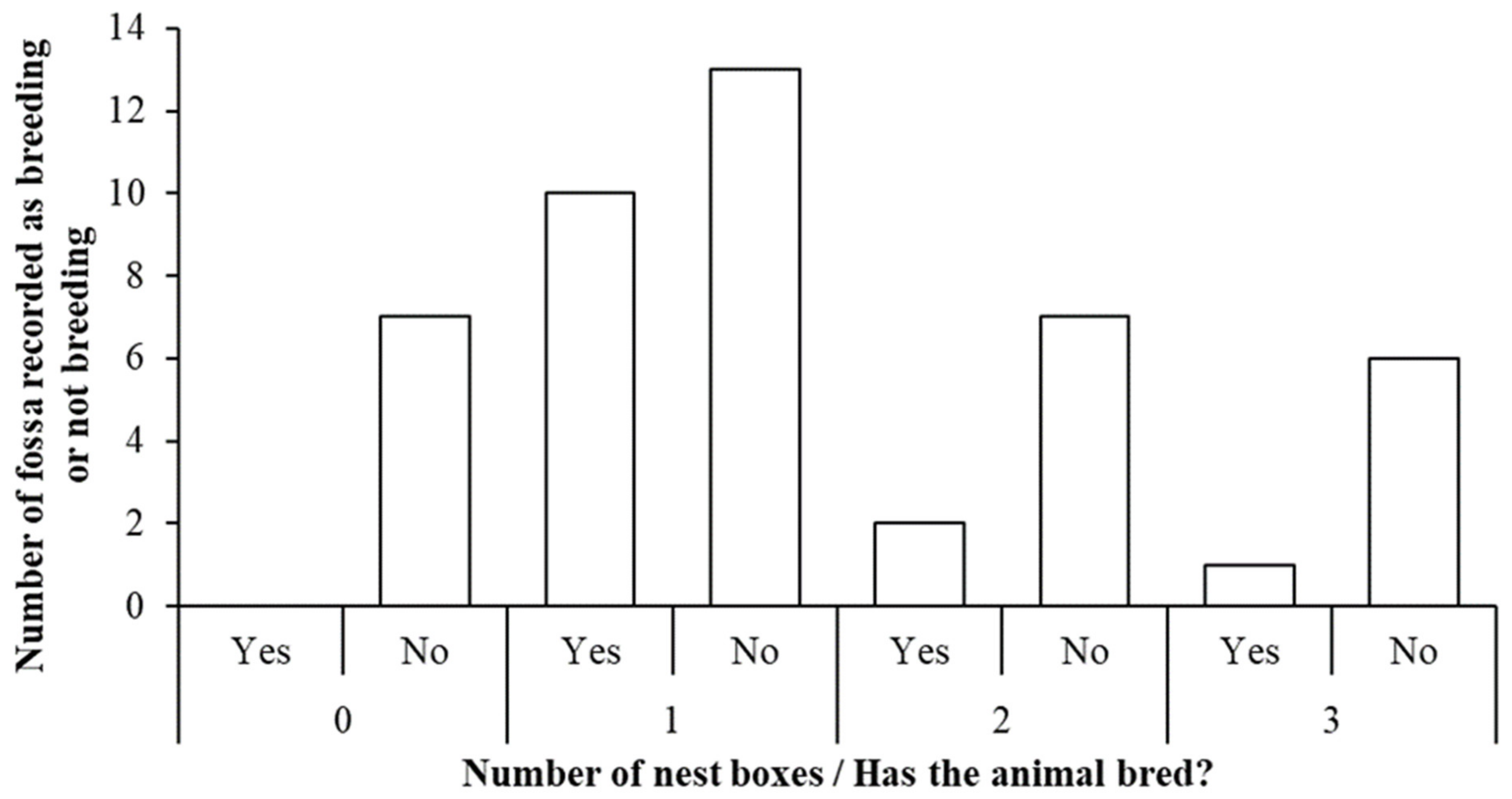
3.3. Breeding
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hosey, G.; Melfi, V.; Pankhurst, S. Zoo Animals: Behaviour, Management, and Welfare; Oxford University Press: New York, NY, USA, 2013. [Google Scholar]
- Ward, S.J.; Sherwen, S.; Clark, F.E. Advances in applied zoo animal welfare science. Appl. Anim. Behav. Sci. 2018, 21 (Suppl. 1), 23–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rose, P.E.; Brereton, J.E.; Rowden, L.J.; De Figueiredo, R.L.; Riley, L. What’s new from the zoo? An analysis of ten years of zoo-themed research output. Palgrave Commun. 2019, 5, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Mellor, E.; Brilot, B.; Collins, S. Abnormal repetitive behaviours in captive birds: A Tinbergian review. Appl. Anim. Behav. Sci. 2018, 198, 109–120. [Google Scholar] [CrossRef] [Green Version]
- Clubb, R.; Mason, G.J. Captivity effects on wide-ranging carnivores. Nature 2003, 425, 473–474. [Google Scholar] [CrossRef] [PubMed]
- Clubb, R.; Mason, G.J. Natural behavioural biology as a risk factor in carnivore welfare: How analysing species differences could help zoos improve enclosures. Appl. Anim. Behav. Sci. 2007, 102, 303–328. [Google Scholar] [CrossRef] [Green Version]
- Kroshko, J.; Clubb, R.; Harper, L.; Mellor, E.; Moehrenschlager, A.; Mason, G. Stereotypic route tracing in captive Carnivora is predicted by species-typical home range sizes and hunting styles. Anim. Behav. 2016, 117, 197–209. [Google Scholar] [CrossRef] [Green Version]
- Carlstead, K. Determining the causes of stereotypic behaviors in zoo carnivores: Toward appropriate enrichment strategies. Second nature: Environmental enrichment for captive animals. In Second Nature: Environmental Enrichment for Captive Animals; Smithsonian Books: Washington, DC, USA, 1998; pp. 172–183. [Google Scholar]
- Veasey, J. Concepts in the care and welfare of captive elephants. Int. Zoo Yearb. 2006, 40, 63–79. [Google Scholar] [CrossRef]
- Morgan, K.N.; Tromborg, C.T. Sources of stress in captivity. Appl. Anim. Behav. Sci. 2007, 102, 262–302. [Google Scholar] [CrossRef]
- Rose, P.E.; Roffe, S.M. A Case Study of Malayan Tapir (Tapirus indicus) Husbandry Practice Across 10 Zoological Collections. Zoo Biol. 2013, 32, 347–356. [Google Scholar] [CrossRef]
- Rowden, L.J.; Rose, P.E. A global survey of banteng (Bos javanicus) housing and husbandry. Zoo Biol. 2016, 35, 546–555. [Google Scholar] [CrossRef] [Green Version]
- Fuller, G.; Kuhar, C.W.; Dennis, P.M.; Lukas, K.E. A Survey of Husbandry Practices for Lorisid Primates in North American Zoos and Related Facilities. Zoo Biol. 2013, 32, 88–100. [Google Scholar] [CrossRef]
- Wall, E.L.; Hartley, M. Assessing enclosure design and husbandry practices for successful keeping and breeding of the Burmese brow antlered deer (Eld’s deer, Rucervus eldii thamin) in European zoos. Zoo Biol. 2017, 36, 201–212. [Google Scholar] [CrossRef]
- Tennant, K.; Segura, V.; Morris, M.; Denninger Snyder, K.; Bocian, D.; Maloney, D.; Maple, T. Achieving optimal welfare for the Nile hippopotamus (Hippopotamus amphibius) in North American zoos and aquariums. Behav. Process. 2018, 156, 51–57. [Google Scholar] [CrossRef]
- Rose, P.; Rowden, L.J. for the Swamp, Catered for in Captivity? A Cross-Institutional Evaluation of Captive Husbandry for Two Species of Lechwe. Animals 2020, 10, 1874. [Google Scholar] [CrossRef]
- Hawkins, F. Cryptoprocta ferox. The IUCN Red List of Threatened Species 2016: e.T5760A45197189. 2016. Available online: https://dx.doi.org/10.2305/IUCN.UK.2016-1.RLTS.T5760A45197189.en (accessed on 19 July 2019).
- Brooke, Z.M.; Bielby, J.; Nambiar, K.; Carbone, C. Correlates of Research Effort in Carnivores: Body Size, Range Size and Diet Matter. PLoS ONE 2014, 9, e93195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerber, B.D.; Karpanty, S.M.; Randrianantenaina, J. The impact of forest logging and fragmentation on carnivore species composition, density, and occupancy in Madagascar’s rainforests. Oryx 2012, 46, 414–422. [Google Scholar] [CrossRef] [Green Version]
- Hawkins, C.E.; Racey, P.A. Food habits of an endangered carnivore, Cryptoprocta ferox, in the dry deciduous forests of western Madagascar. J. Mammal. 2008, 89, 64–74. [Google Scholar] [CrossRef] [Green Version]
- Hawkins, C.; Racey, P. Low population density of a tropical forest carnivore, Cryptoprocta ferox: Implications for protected area management. Oryx 2005, 39, 35–43. [Google Scholar] [CrossRef] [Green Version]
- Lührs, M.-L.; Dammhahn, M. An unusual case of cooperative hunting in a solitary carnivore. J. Ethol. 2009, 28, 379–383. [Google Scholar] [CrossRef] [Green Version]
- Hawkins, C. Cryptoprocta ferox. Fossa, Fosa. In The Natural History of Madagascar; Goodman, S., Benstead, J., Eds.; The University of Chicago Press: Chicago, IL, USA, 2003; pp. 1361–1363. [Google Scholar]
- Lührs, M.-L.; Dammhahn, M.; Kappeler, P. Strength in numbers: Males in a carnivore grow bigger when they associate and hunt cooperatively. Behav. Ecol. 2012, 24, 21–28. [Google Scholar] [CrossRef] [Green Version]
- Veron, G.; Dupré, D.; Lührs, M.-L.; Kappeler, P.M.; Dollar, L.; Pomerantz, J.; Goodman, S.M. Genetic polymorphism and structure of wild and zoo populations of the fosa (Eupleridae, Carnivora), the largest living carnivoran of Madagascar. Mamm. Biol. 2018, 92, 68–77. [Google Scholar] [CrossRef]
- Hawkins, C. The Behaviour and Ecology of the Fossa, Cryptoprocta ferox (Carnivora: Viverridae) in a Dry Deciduous Forest in Western Madagascar; University of Aberdeen: Aberdeen, UK, 1998. [Google Scholar]
- Hawkins, F.; Racey, P.A. A novel mating system in a solitary carnivore: The fossa. J. Zool. 2009, 277, 196–204. [Google Scholar] [CrossRef]
- Hawkins, C.E.; Dallas, J.F.; Fowler, P.A.; Woodroffe, R.; Racey, P.A. Transient masculinization in the fossa, Cryptoprocta ferox (Carnivora, Viverridae). Biol. Reprod. 2002, 66, 610–615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hornsey, T. Breeding the fossa at Suffolk Wildlife Park. Int. Zoo Yearb. 1999, 46, 407–417. [Google Scholar]
- Albignac, R. Breeding the fossa at Montpellier Zoo. Int. Zoo Yearb. 1975, 15, 147–150. [Google Scholar] [CrossRef]
- Krebs, M.; Bryan, C. Population Analysis and Breeding and Transfer Plan Fossa (Cryptoprocta ferox) AZA Species Survival Plan Yellow Program; Omaha Zoo: Omaha, NE, USA, 2012; p. 20. [Google Scholar]
- Species360 Zoological Information Management System (ZIMS). 2018. Available online: https://zims.species360.org/Login.aspx?ReturnUrl=%2f (accessed on 2 March 2017).
- Reiter, J.; Winkler, A. EEP Studbook for the Fossa (Cryptoprocta ferox) Edition 2012; European Association of Zoos and Aquaria: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Pfleiderer, J. Fossa (Cryptoporcta ferox); EEP Annual Report 2017; EAZA: Amsterdam, The Netherlands, 2017. [Google Scholar]
- Krebs, M. Email correspondence. 23 April 2019. [Google Scholar]
- Farris, Z.J.; Gerber, B.D.; Karpanty, S.; Murphy, A.; Andrianjakarivelo, V.; Ratelolahy, F.; Kelly, M.J. When carnivores roam: Temporal patterns and overlap among M adagascar’s native and exotic carnivores. J. Zool. 2015, 296, 45–57. [Google Scholar] [CrossRef]
- McGowan, P.J.; Traylor-Holzer, K.; Leus, K. IUCN guidelines for determining when and how ex situ management should be used in species conservation. Conserv. Lett. 2017, 10, 361–366. [Google Scholar] [CrossRef]
- Kitchener, A.C. Small carnivorans, museums and zoos. Int. Zoo Yearb. 2020. [Google Scholar] [CrossRef]
- AZA Small Carnivore TAG. Mongoose, Meerkat, & Fossa (Herpestidae/Eupleridae) Care Manual; Association of Zoos and Aquariums: Silver Spring, MD, USA, 2011; p. 103. [Google Scholar]
- Reiter, J.; Winkler, A. Husbandry Guidelines for the Fossa (Cryptoprocta ferox); European Association of Zoos and Aquaria: Amsterdam, The Netherlands, 2013. [Google Scholar]
- Winkler, A. Husbandry Guidelines for the Fossa (Cryptoprocta ferox); Prepared for EAZA SCTAG Husbandry Guidelines; Duisberg Zoo: Duisburg, Germany, 2002. [Google Scholar]
- Bennett, C.; Torgerson-White, L.; Fripp, D.; Watters, J.; Petric, A. A multi-institutional assessment of factors influencing locomotion and pacing in captive okapis (Okapia johnstoni). Appl. Anim. Welf. Sci. 2015, 18 (Supp. 1), S43–S61. [Google Scholar] [CrossRef]
- Hogan, L.A.; Tribe, A. Prevalence and cause of stereotypic behaviour in common wombats (Vombatus ursinus) residing in Australian zoos. Appl. Anim. Behav. Sci. 2007, 105, 180–191. [Google Scholar] [CrossRef]
- Boyd, L.E. Time budgets of adult Przewalski horses: Effects of sex, reproductive status and enclosure. Appl. Anim. Behav. Sci. 1998, 21, 19–39. [Google Scholar] [CrossRef]
- Rose, P.E.; Nash, S.M.; Riley, L.M. To pace or not to pace? A review of what abnormal repetitive behaviour tells us about zoo animal management. J. Vet. Sci. 2017, 20, 11–21. [Google Scholar]
- Hosey, G.R. How does the zoo environment affect the behaviour of captive primates? Appl. Anim. Behav. Sci. 2005, 90, 107–109. [Google Scholar] [CrossRef]
- Quadros, S.; Goulart, V.D.; Passos, L.; Vecci, M.A.; Young, R.J. Zoo visitor effect on mammal behaviour: Does noise matter? Appl. Anim. Behav. Sci. 2014, 156, 78–84. [Google Scholar] [CrossRef]
- Sellinger, R.L.; Ha, J.C. The effects of visitor density and intensity on the behaviour of two captive jaguars (Panthera onca). J. Appl. Anim. Welf. Sci. 2015, 8, 233–244. [Google Scholar] [CrossRef]
- Suárez, P.; Recuerda, P.; Arias-de-Reyna, L. Behaviour and welfare: The visitor effect in captive felids. Anim. Welf. 2017, 26, 25–34. [Google Scholar] [CrossRef] [Green Version]
- Boyle, S.A.; Berry, N.; Cayton, J.; Ferguson, S.; Gilgan, A.; Khan, A.; Lam, H.; Leavelle, S.; Mulder, I.; Myers, R.; et al. Widespread Behavioural Responses by Mammals and Fish to Zoo Visitors Highlight Differences between Individual Animals. Animals 2020, 10, 2108. [Google Scholar] [CrossRef]
- Nimon, A.J.; Dalziel, F.R. Cross-species interaction and communication: A study method applied to captive siamang (Hylobates syndactylus) and long-billed corella (Cacatua teuirostris) contacts with humans. Appl. Anim. Behav. Sci. 1992, 22, 261–272. [Google Scholar] [CrossRef]
- Margulis, S.W.; Hoyos, C.; Anderson, M. Effect of felid activity on zoo visitor interest. Zoo Biol. 2003, 22, 587–599. [Google Scholar] [CrossRef]
- Fernandez, E.J.; Tamborski, M.A.; Pickens, S.R.; Timberlake, W. Animal–visitor interactions in the modern zoo: Conflicts and interventions. Appl. Anim. Behav. Sci. 2009, 120, 1–8. [Google Scholar] [CrossRef]
- Mason, G.; Clubb, R.; Latham, N.; Vickery, S. Why and how should we use environmental enrichment to tackle stereotypic behaviour? Appl. Anim. Behav. Sci. 2007, 102, 163–188. [Google Scholar] [CrossRef] [Green Version]
- Cooper, J.J.; Nicol, C.J. The ‘coping’ hypothesis of stereotypic behaviour: A reply to Rushen. Anim. Behav. 1993, 45, 616–618. [Google Scholar] [CrossRef] [Green Version]
- Wechsler, B. Coping and coping strategies: A behavioural view. Appl. Anim. Behav. Sci. 1995, 43, 123–134. [Google Scholar] [CrossRef]
- Maple, T.L.; Perdue, B.M. Zoo Animal Welfare; Springer: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- Kennes, D.; de Rycke, P.H. Influence of performance of stereotypies on plasma corticosterone and leucocyte levels in the bank vole (Clethrionomys glareolus). In Proceedings of the International Congress on Applied Ethology in Farm Animals; Unshelem, G., van Patten, K., Zeeb, K., Eds.; Kuratorium für Technik und Bauwesen in der Landwirtschaft: Darmstadt, Germany, 1988; pp. 238–240. [Google Scholar]
- McGreevy, P.D.; Nicol, C.J. The effect of short-term prevention on the subsequent rate of crib-bitting in Thoroughhred horses. Equine Vet. J. 1998, 27, 30–34. [Google Scholar]
- McBride, S.D.; Cuddeford, D. The putative welfare-reducing effects of preventing equine stereotypic behaviour. Anim. Welf. 2001, 10, 173–189. [Google Scholar]
- Markowitz, H.; Aday, C. Power for captive animals. In Second Nature: Environmental Enrichment for Captive Animals; Shepherdson, D., Forthman, D.L., Eds.; Smithsonian Institution Press: Washington, DC, USA, 1998; p. 47. [Google Scholar]
- Owen, M.A.; Swaisgood, R.R.; Czekala, N.M.; Lindburg, D.G. Enclosure choice and well-being in giant pandas: Is it all about control? Zoo Biol. 2005, 24, 475–481. [Google Scholar] [CrossRef]
- McPhee, M.E.; Carlstead, K. The Importance of Maintaining Natural Behaviours in Captive Mammals. In Wild Mammals in Captivity: Principles and Techniques for Zoo Management; University of Chicago Press: Chicago, IL, USA, 2010; pp. 303–313. [Google Scholar]
- Kurtycz, L.M.; Wagner, K.E.; Ross, S.R. The choice to access outdoor areas affects the behaviour of great apes. J. Appl. Anim. Welf. Sci. 2014, 17, 185–197. [Google Scholar] [CrossRef]
- Ross, S.R. Issues of choice and control in the behaviour of a pair of captive polar bears (Ursus maritimus). Behav. Process. 2006, 73, 117–120. [Google Scholar] [CrossRef] [PubMed]
- Watters, J.V. Searching for behavioral indicators of welfare in zoos: Uncovering anticipatory behavior. Zoo Biol. 2014, 33, 251–256. [Google Scholar] [CrossRef] [PubMed]
- Spruijt, B.M.; Van den Bos, R.; Pijlman, F.T.A. A concept of welfare based on reward evaluating mechanisms in the brain: Anticipatory behaviour as an indicator for the state of reward systems. Appl Anim Behav Sci. 2001, 72, 145–171. [Google Scholar] [CrossRef]
- Jensen, A.L.; Delfour, F.; Carter, T. Anticipatory behavior in captive bottlenose dolphins (Tursiops truncatus): A preliminary study. Zoo Biol. 2013, 32, 436–444. [Google Scholar] [CrossRef]
- Bassett, L.; Buchanan-Smith, H.M. Effects of predictability on the welfare of captive animals. Appl. Anim. Behav. Sci. 2007, 102, 223–245. [Google Scholar] [CrossRef] [Green Version]
- Berridge, K.C. Food reward: Brain substrates of wanting and liking. Neurosci. Biobehav. Rev. 1996, 20, 1–25. [Google Scholar] [CrossRef]
- Wise, R.A. Dopamine and reward: The anhedonia hypothesis 30 years on. Neurotox. Res. 2008, 14, 169–183. [Google Scholar] [CrossRef] [Green Version]
- Berridge, K.C. The debate over dopamine’s role in reward: The case for incentive salience. Psychopharmacology 2007, 191, 391–431. [Google Scholar] [CrossRef]
- Anderson, C.; von Keyserlingk, M.A.G.; Lidfors, L.M.; Weary, D.M. Anticipatory behaviour in animals: A critical review. Anim. Welf. 2020, 29, 231–238. [Google Scholar] [CrossRef]
- Watters, J.V.; Krebs, B.L.; Eschmann, C.L. Assessing Animal Welfare with Behavior: Onward with Caution. J. Zool. Bot. Gard. 2021, 2, 75–87. [Google Scholar] [CrossRef]
- Swaisgood, R.R.; Shepherdson, D.J. Scientific approaches to enrichment and stereotypies in zoo animals: What’s been done and where should we go next? Zoo Biol. 2005, 24, 499–518. [Google Scholar] [CrossRef]
- Carlstead, K.; Shepherdson, D. Alleviating stress in zoo animals with environmental enrichment. In The Biology of Animal Stress: Basic Principles and Implications for Animal Welfare; University of California: Davis, CA, USA, 2000; pp. 337–354. [Google Scholar]
- Neave, H.W.; Webster, J.R.; Zobel, G. Anticipatory behaviour as an indicator of the welfare of dairy calves in different housing environments. PLoS ONE 2021, 16, e0245742. [Google Scholar] [CrossRef] [PubMed]
- Mendl, M.; Burman, O.H.P.; Paul, E.S. An integrative and functional framework for the study of animal emotion and mood. Proc. R. Soc. B 2010, 277, 2895–2904. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Zoo | Number & Sex Ratio Male.Female | Age Range | Housing |
|---|---|---|---|
| 1 | 0.2 | 1–2 yrs | Single sex related pair |
| 2 | 1.1 | 4–10 yrs | Singly |
| 3 | 1.1 | 2–7 yrs | Singly |
| 4 | 1.1 | 10+ yrs | Singly |
| 5 | 0.1 | 10+ yrs | Singly |
| 6 | 0.1 | 10+ yrs | Singly |
| 7 | 1.1 | 10+ yrs | Singly |
| 8 | 2.1 | 6–7 yrs | Single sex related pair (m), singly (f) |
| 9 | 1.0 | 10+ yrs | Singly |
| 10 | 1.1 | 10+ yrs | Singly |
| 11 | 3.1 | 1–10+ yrs | Singly (m, f), single sex related pair (m) |
| 12 | 1.0 | 7–8 yrs | Singly |
| 13 | 2.0 | 4–5 yrs | Singly |
| 14 | 0.1 | 3–4 yrs | Singly |
| 15 | 1.0 | 4–5 yrs | Singly |
| 16 | 1.1 | 10+ yrs | Singly |
| 17 | 1.0 | 10+ yrs | Singly |
| 18 | 1.0 | 10+ yrs | Singly |
| 19 | 1.1 | 9–10+ yrs | Singly |
| 20 | 1.0 | 3–4 yrs | Singly |
| 21 | 1.1 | 8–10+ yrs | Singly |
| 22 | 1.0 | 4–5 yrs | Singly |
| 23 | 1.0 | 10+ yrs | Singly |
| 24 | 1.1 | 6–7 yrs | Singly |
| 25 | 0.1 | 10+ yrs | Singly |
| 26 | 2.3 | 1–10+ yrs | Singly (f), breeding pair (when female without offspring (m.f)), parent (f) w/offspring (m,f) |
| Pacing | Any repetitive locomotion in a fixed pattern, such as back and forth along the same route or pattern e.g., figure of eight. Behaviour appears to have no goal or function. |
| Self-directed | Any self-injurious behaviours, such as biting/chewing/sucking/licking, causing wounds, missing fur or irritations. e.g., excessive grooming, tail/paw biting/sucking. |
| Head rolling | Tossing head in circular motion |
| Anticipatory | Behaviours relating to daily cleaning/feeding routines, e.g., locomotion, vocalisations, waiting at hatch, keeper-directed behaviours |
| Other or Combination | Please outline in as much detail as possible the fossa’s behaviour in the comment box |
| Factor | Indoor Area | Outdoor Area | Outdoor Height |
|---|---|---|---|
| Sex | No significant difference H = 0.64; df = 1; P = 0.425 Female: 11.25 m2 Male: 10 m2 | No significant difference H = 2.14; df = 1; P = 0.144 Female: 94.8 m2 Male: 50 m2 | No significant difference H = 1.62; df = 1; P = 0.203 Female: 6 m Male: 4.5 m |
| Age | Approaches significance H = 5.15; df = 2; P = 0.076 Young: 14.61 m2 Adult: 9 m2 Old adult: 10 m2 | No significant difference H = 0.77; df = 2; P = 0.680 Young: 94.8 m2 Adult: 49.5 m2 Old adult: 73.3 m2 | No significant difference H = 0.92; df = 2; P = 0.631 Young: 4.5 m Adult: 6 m Old adult: 4.75 m |
| Number in enclosure | No significant difference H = 2.95; df = 1; P = 0.09 Single: 9.5 m2 Multiple: 14.61 m2 | No significant difference H = 0.84; df = 1; P = 0.361 Single: 50 m2 Multiple: 94.8 m2 | No significant difference H = 0.22; df = 1; P = 0.641 Single: 5.5 m Multiple: 4.5 m |
| Shut on/off show? | No significant difference H = 0.17; df = 1; P = 0.680 Yes: 10 m2 No: 10.5 m2 | No significant difference H = 0.14; df = 1; P = 0.705 Yes: 76.5 m2 No: 45.5 m2 | No significant difference H = 0.57; df = 1; P = 0.450 Yes: 6 m No: 4 m |
| Escape from public viewing? | No significant difference H = 1.34; df = 1; P = 0.248 Yes: 10 m2 No: 14.61 m2 | No significant difference H = 1.78; df = 1; P = 0.182 Yes: 48 m2 No = 94.8 m2 | No significant difference H = 0.01; df = 1; P = 0.933 Yes: 5 m No: 4.5 m |
| Continent | Significant difference H = 5.50; df = 1; P = 0.02 Europe: 10.3 m2 USA: 6.25 m2 | No significant difference H = 1.64; df = 1; P = 0.20 Europe: 60 m2 USA: 70 m2 | No significant difference H = 1.9; df = 1; P = 0.168 Europe: 6 m USA: 4.5 m |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Harley, J.J.; O’Hara, L.; Rose, P.E. A Global Survey of Current Zoo Housing and Husbandry Practices for Fossa: A Preliminary Review. J. Zool. Bot. Gard. 2021, 2, 388-405. https://doi.org/10.3390/jzbg2030028
Harley JJ, O’Hara L, Rose PE. A Global Survey of Current Zoo Housing and Husbandry Practices for Fossa: A Preliminary Review. Journal of Zoological and Botanical Gardens. 2021; 2(3):388-405. https://doi.org/10.3390/jzbg2030028
Chicago/Turabian StyleHarley, Jessica J., Lisa O’Hara, and Paul E. Rose. 2021. "A Global Survey of Current Zoo Housing and Husbandry Practices for Fossa: A Preliminary Review" Journal of Zoological and Botanical Gardens 2, no. 3: 388-405. https://doi.org/10.3390/jzbg2030028
APA StyleHarley, J. J., O’Hara, L., & Rose, P. E. (2021). A Global Survey of Current Zoo Housing and Husbandry Practices for Fossa: A Preliminary Review. Journal of Zoological and Botanical Gardens, 2(3), 388-405. https://doi.org/10.3390/jzbg2030028