
High-Avidity Anti-Filovirus IgG Elicited Using Protein Subunit Vaccines Does Not Correlate with Protection

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
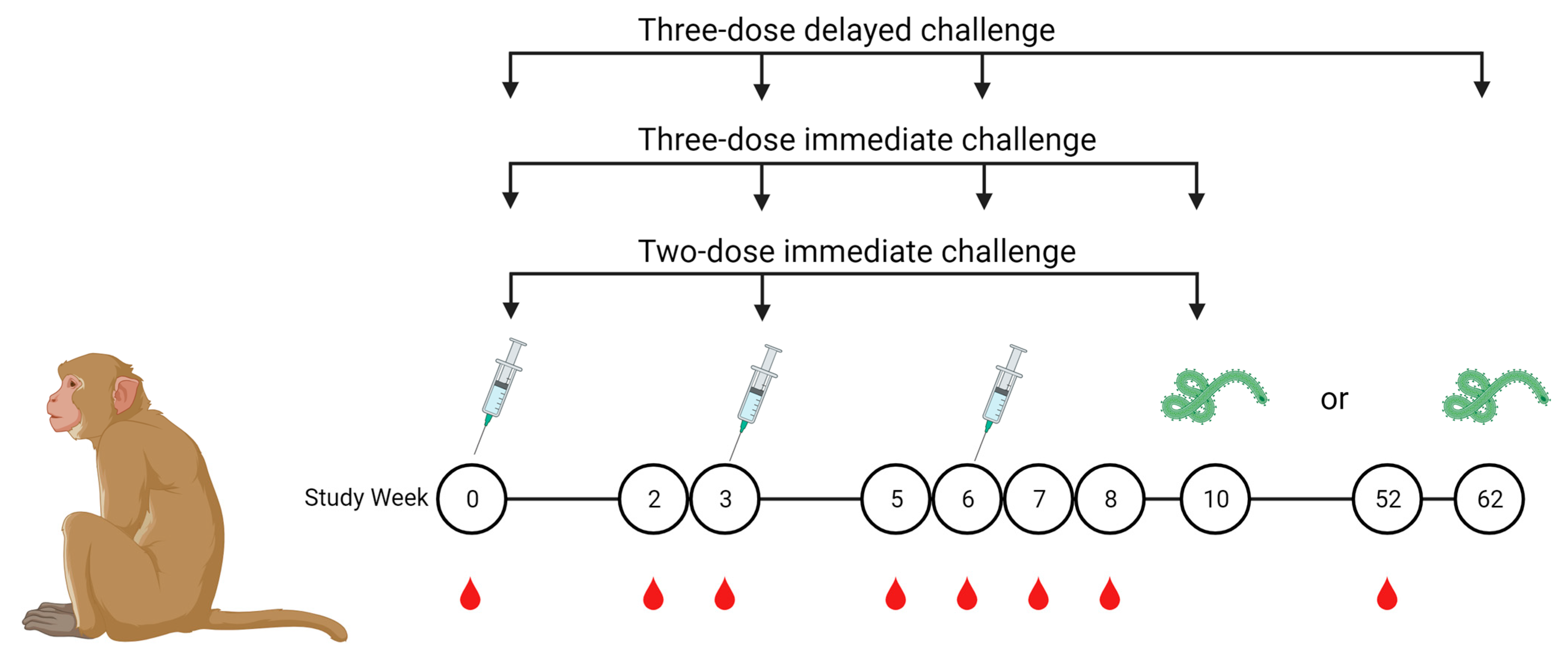
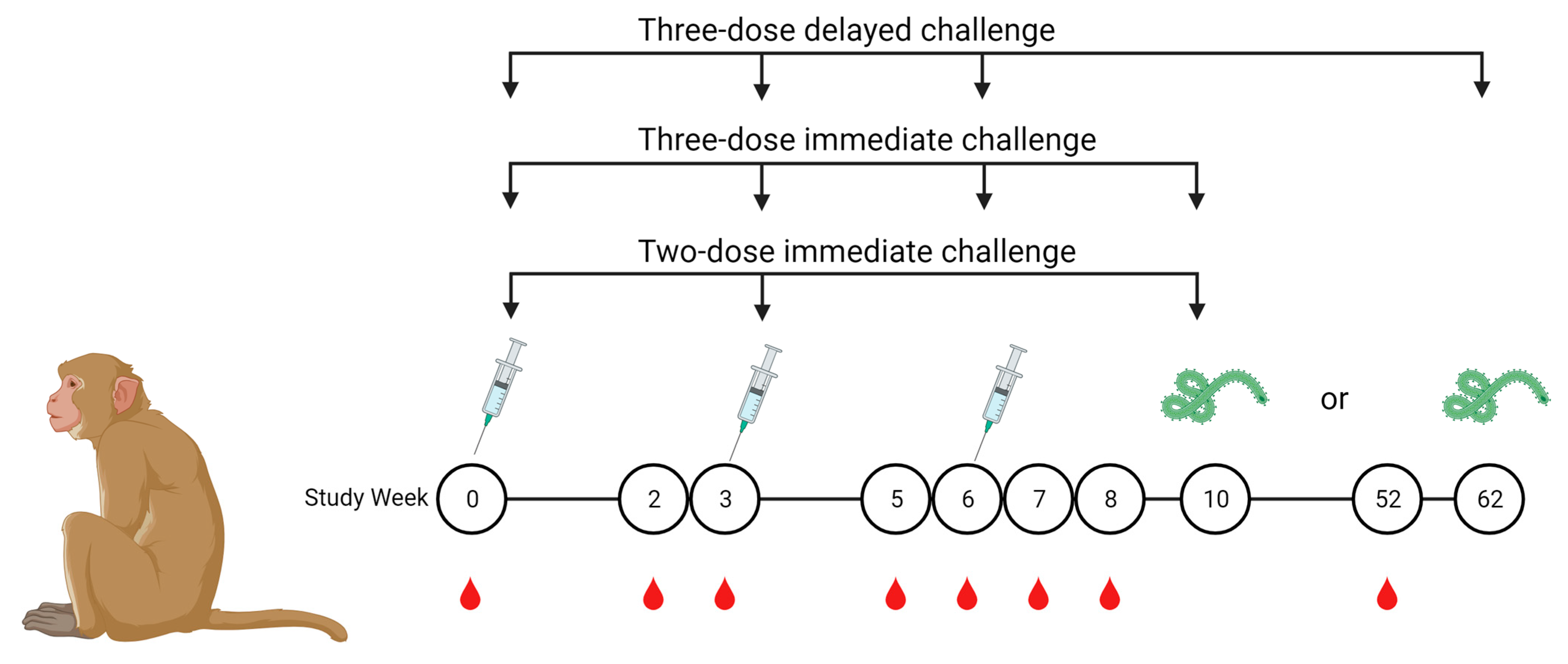
2.1. Vaccines and Nonhuman Primate Immunizations
2.2. Quantification of Antigen-Specific IgG
2.3. Avidity Assay
2.4. Statistical Analysis
3. Results
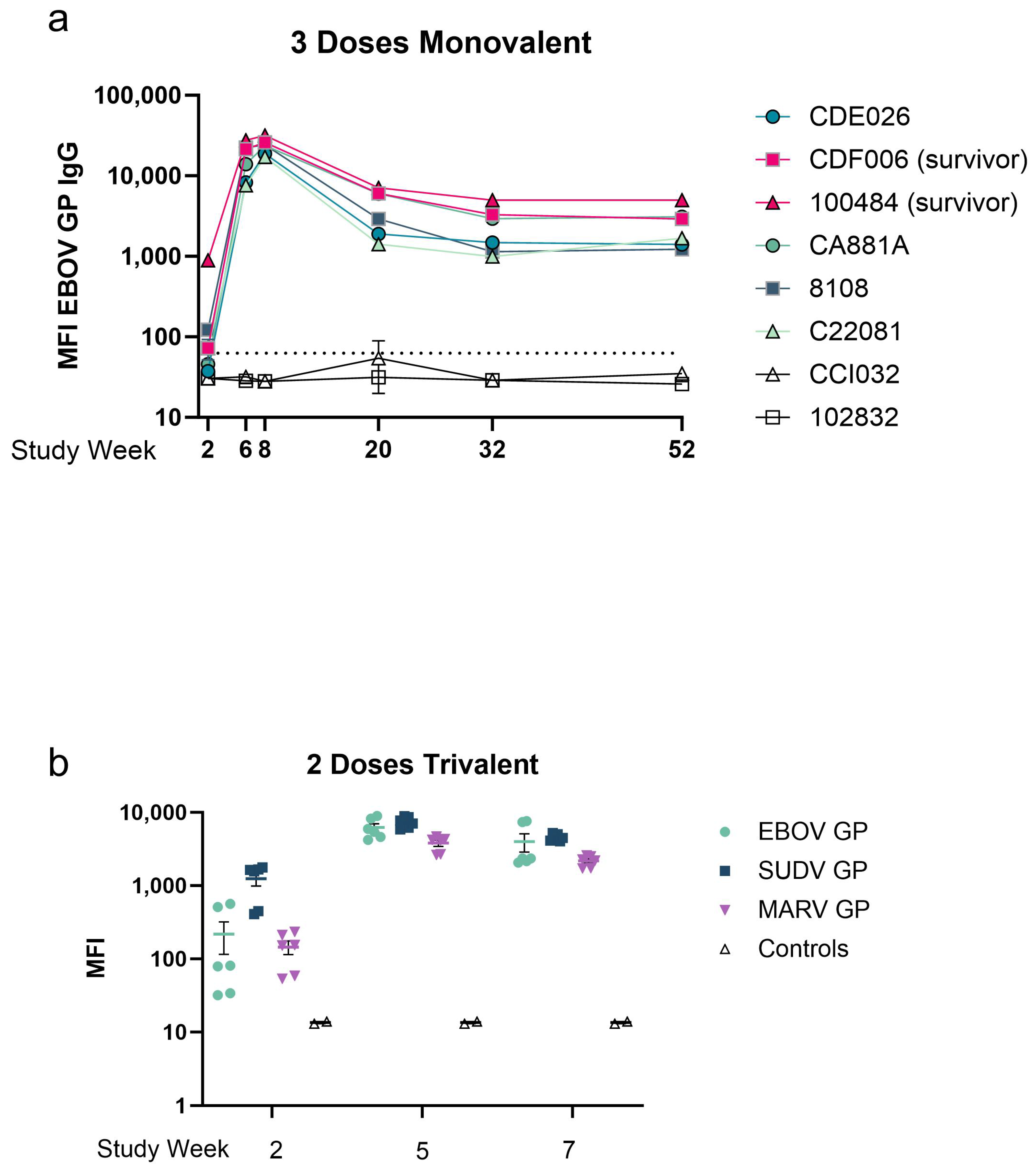
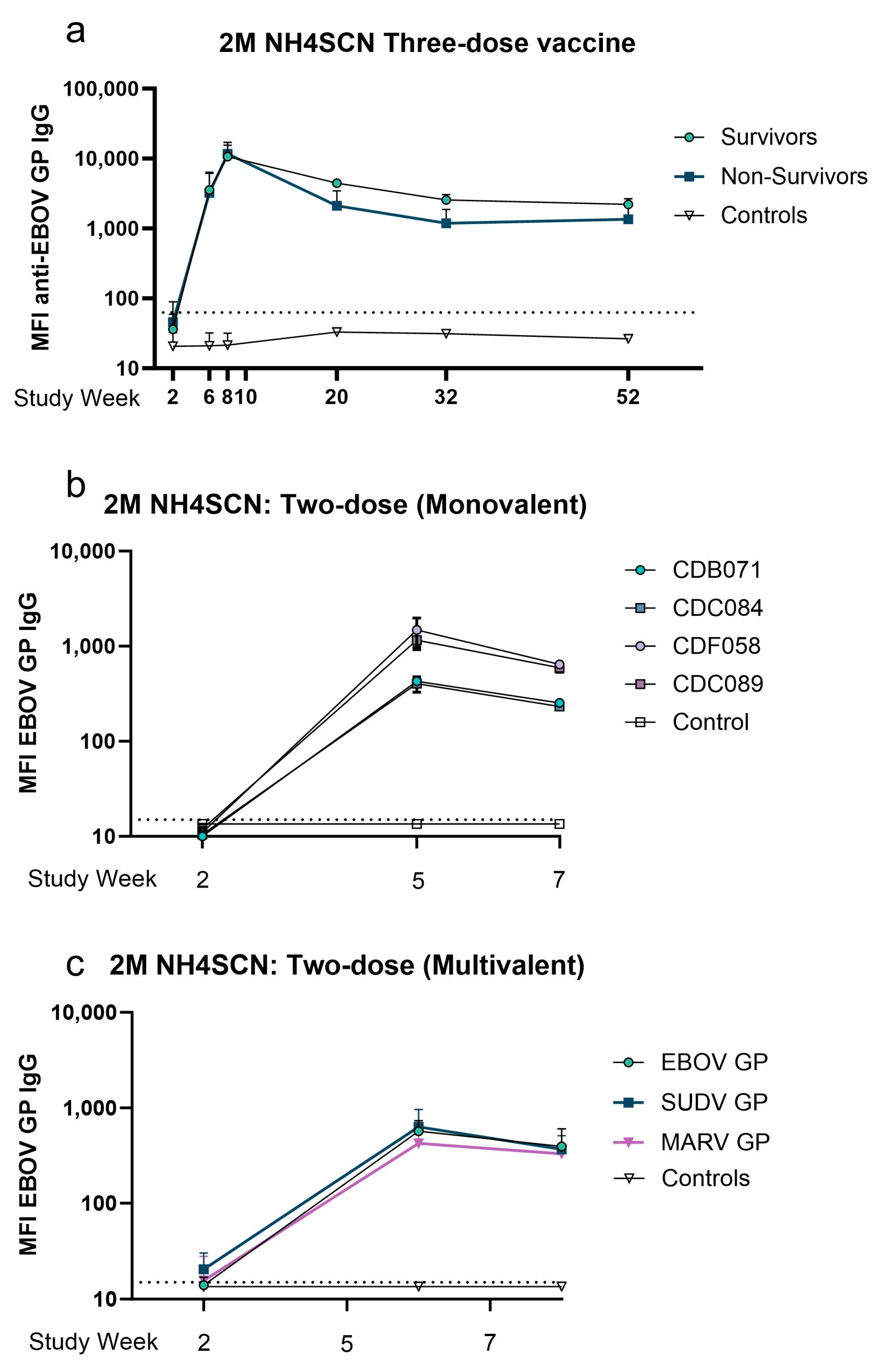
3.1. Avidity Maturation Peaks with Three Doses of Vaccine
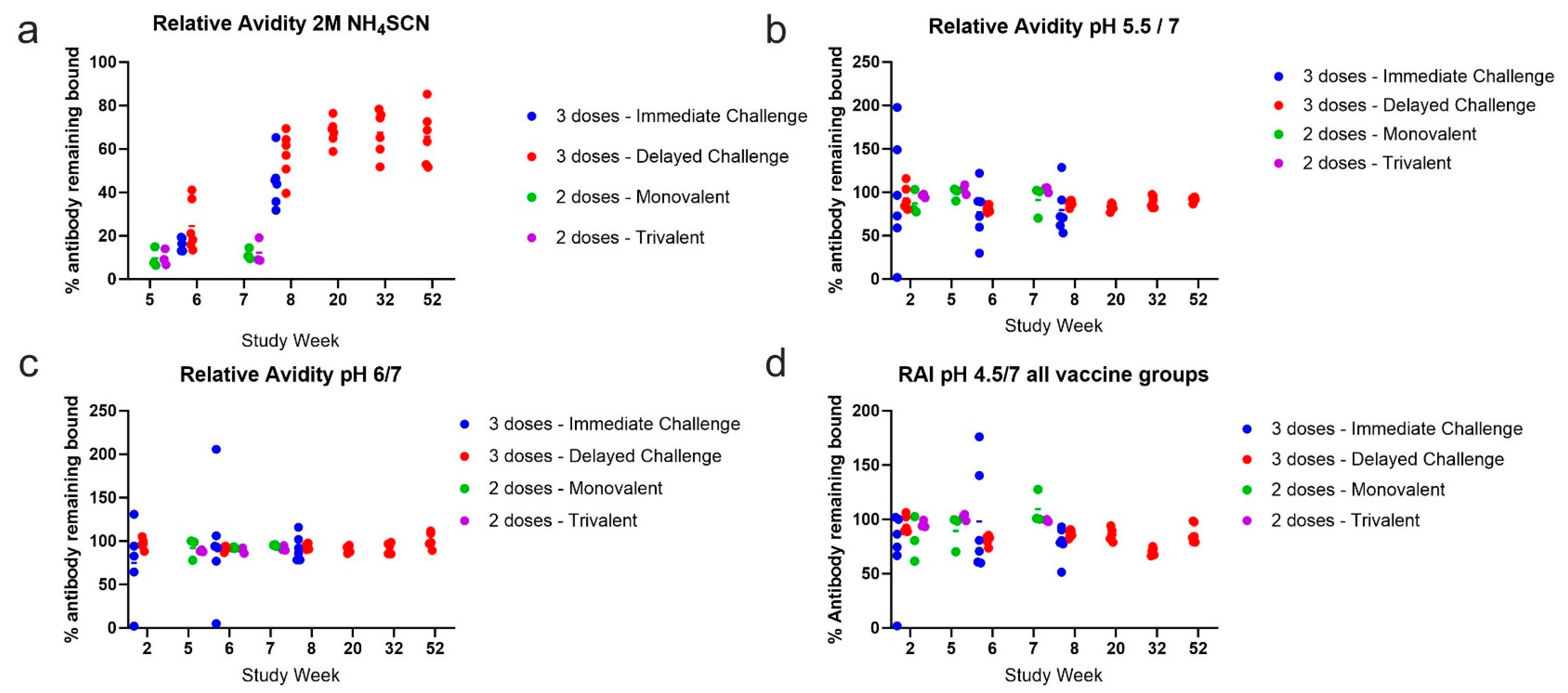
3.2. Avidity of GP-Specific Antibodies from Cynomolgus Macaques Increases with Increasing Number of Vaccine Doses
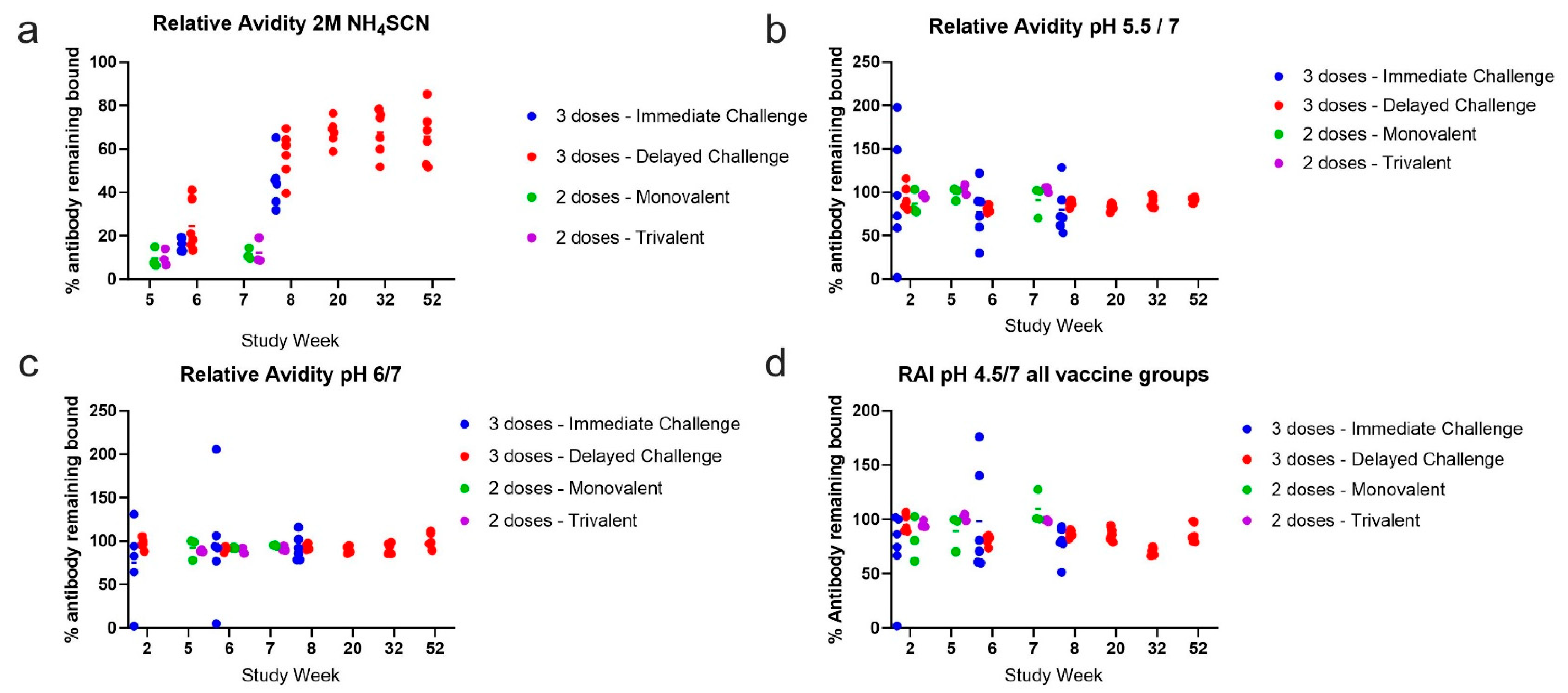
3.3. Antibody Avidity Remains High during a One-Year Rest Period Prior to Viral Challenge
3.4. A Two-Dose Recombinant EBOV GP Vaccine Regimen Generates Moderate Antigen-Specific Avidity
3.5. Avidity of Cross-Reactive Antibody
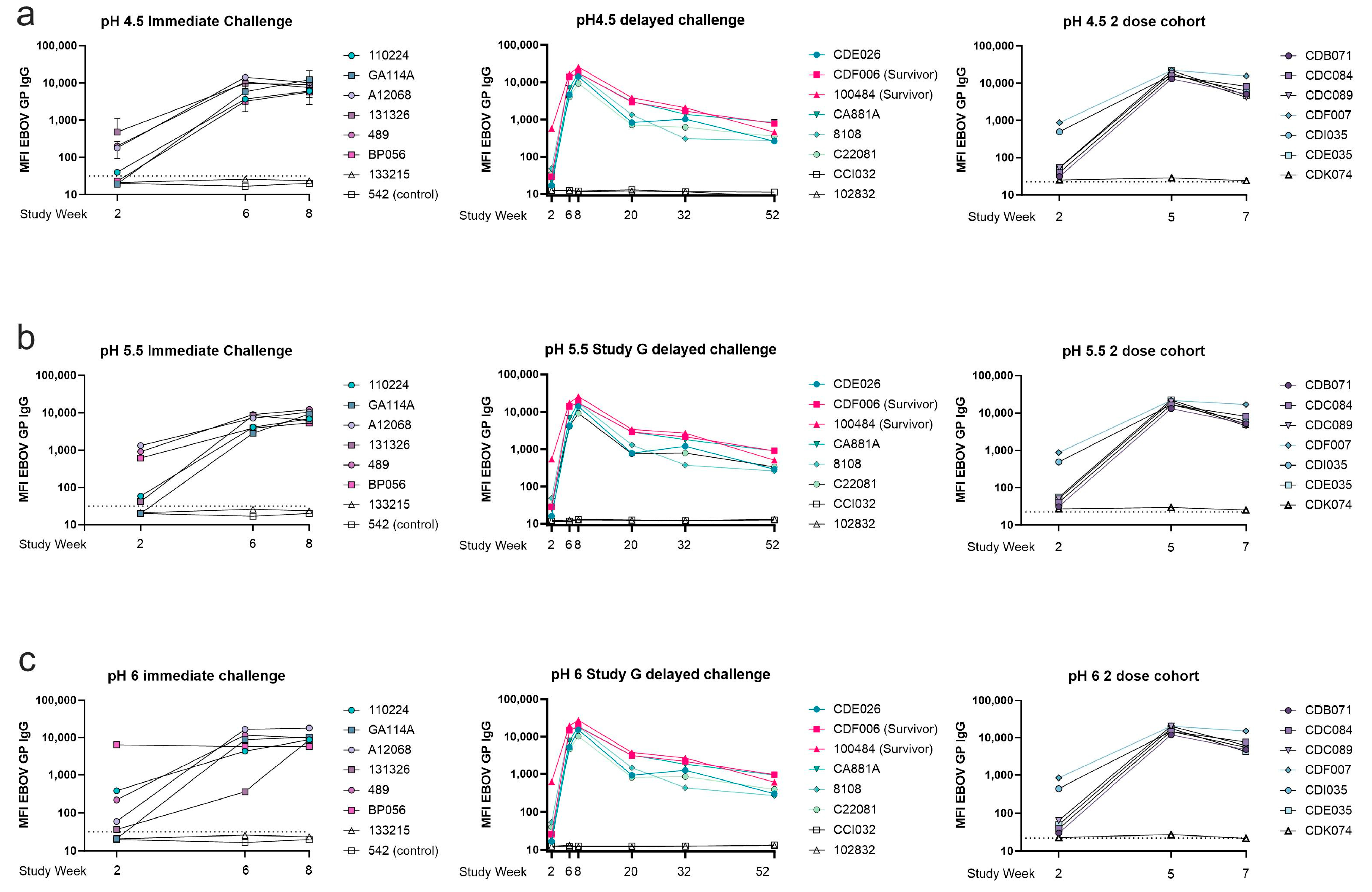
3.6. Binding at Low pH
3.7. Relative Avidity Index
3.8. Avidity Does Not Correlate with Survival or Survival Day
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
References
- Longet, S.; Mellors, J.; Carroll, M.W.; Tipton, T. Ebolavirus: Comparison of Survivor Immunology and Animal Models in the Search for a Correlate of Protection. Front. Immunol. 2020, 11, 599568. [Google Scholar] [CrossRef] [PubMed]
- Henao-Restrepo, A.M.; Camacho, A.; Longini, I.M.; Watson, C.H.; Edmunds, W.J.; Egger, M.; Carroll, M.W.; Dean, N.E.; Diatta, I.; Doumbia, M.; et al. Efficacy and effectiveness of an rVSV-vectored vaccine in preventing Ebola virus disease: Final results from the Guinea ring vaccination, open-label, cluster-randomised trial (Ebola Ca Suffit!). Lancet 2017, 389, 505–518. [Google Scholar] [CrossRef] [PubMed]
- Milligan, I.D.; Gibani, M.M.; Sewell, R.; Clutterbuck, E.A.; Campbell, D.; Plested, E.; Nuthall, E.; Voysey, M.; Silva-Reyes, L.; McElrath, M.J.; et al. Safety and Immunogenicity of Novel Adenovirus Type 26- and Modified Vaccinia Ankara-Vectored Ebola Vaccines: A Randomized Clinical Trial. JAMA 2016, 315, 1610–1623. [Google Scholar] [CrossRef] [PubMed]
- Medaglini, D.; Santoro, F.; Santoro, C.A. Siegrist, Correlates of vaccine-induced protective immunity against Ebola virus disease. Semin. Immunol. 2018, 39, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Khurana, S.; Ravichandran, S.; Hahn, M.; Coyle, E.M.; Stonier, S.W.; Zak, S.E.; Kindrachuk, J.; Davey, R.T.; Dye, J.M.; Chertow, D.S. Longitudinal Human Antibody Repertoire against Complete Viral Proteome from Ebola Virus Survivor Reveals Protective Sites for Vaccine Design. Cell Host Microbe 2020, 27, 262–276. [Google Scholar] [CrossRef] [PubMed]
- Lehrer, A.T.; Chuang, E.; Namekar, M.; Williams, C.A.; Wong, T.A.S.; Lieberman, M.M.; Granados, A.; Misamore, J.; Yalley-Ogunro, J.; Andersen, H.; et al. Recombinant Protein Filovirus Vaccines Protect Cynomolgus Macaques from Ebola, Sudan, and Marburg Viruses. Front. Immunol. 2021, 12, 703986. [Google Scholar] [CrossRef] [PubMed]
- Markham, A. REGN-EB3: First Approval. Drugs 2021, 81, 175–178. [Google Scholar] [CrossRef] [PubMed]
- Gaudinski, M.R.; Coates, E.E.; Novik, L.; Widge, A.; Houser, K.V.; Burch, E.; Holman, L.A.; Gordon, I.J.; Chen, G.L.; Carter, C.; et al. Safety, tolerability, pharmacokinetics, and immunogenicity of the therapeutic monoclonal antibody mAb114 targeting Ebola virus glycoprotein (VRC 608): An open-label phase 1 study. Lancet 2019, 393, 889–898. [Google Scholar] [CrossRef] [PubMed]
- Misasi, J.; Gilman, M.S.; Kanekiyo, M.; Gui, M.; Cagigi, A.; Mulangu, S.; Corti, D.; Ledgerwood, J.E.; Lanzavecchia, A.; Cunningham, J.; et al. Structural and molecular basis for Ebola virus neutralization by protective human antibodies. Science 2016, 351, 1343–1346. [Google Scholar] [CrossRef] [PubMed]
- Tran, E.E.; Nelson, E.A.; Bonagiri, P.; Simmons, J.A.; Shoemaker, C.J.; Schmaljohn, C.S.; Kobinger, G.P.; Zeitlin, L.; Subramaniam, S.; White, J.M. Mapping of Ebolavirus Neutralization by Monoclonal Antibodies in the ZMapp Cocktail Using Cryo-Electron Tomography and Studies of Cellular Entry. J. Virol. 2016, 90, 7618–7627. [Google Scholar] [CrossRef] [PubMed]
- Prevail II Writing Group. A Randomized, Controlled Trial of ZMapp for Ebola Virus Infection. N. Engl. J. Med. 2016, 375, 1448–1456. [Google Scholar] [CrossRef] [PubMed]
- Lehrer, A.T.; Lehrer, A.T.; Wong, T.A.S.; Lieberman, M.M.; Johns, L.; Medina, L.; Feldmann, F.; Feldmann, H.; Marzi, A. Recombinant subunit vaccines protect guinea pigs from lethal Ebola virus challenge. Vaccine 2019, 37, 6942–6950. [Google Scholar] [CrossRef] [PubMed]
- Lehrer, A.T.; Wong, T.A.S.; Lieberman, M.M.; Humphreys, T.; Clements, D.E.; Bakken, R.R.; Hart, M.K.; Pratt, W.D.; Dye, J.M. Recombinant proteins of Zaire ebolavirus induce potent humoral and cellular immune responses and protect against live virus infection in mice. Vaccine 2018, 36, 3090–3100. [Google Scholar] [CrossRef] [PubMed]
- Haun, B.K.; Kamara, V.; Dweh, A.S.; Garalde-Machida, K.; Forkay, S.S.; Takaaze, M.; Namekar, M.; Wong, T.A.S.; Bell-Gam Woto, A.E.; Humphreys, P.; et al. Serological evidence of Ebola virus exposure in dogs from affected communities in Liberia: A preliminary report. PLoS Negl. Trop. Dis. 2019, 13, e0007614. [Google Scholar] [CrossRef] [PubMed]
- Babirye, P.; Musubika, C.; Kirimunda, S.; Downing, R.; Lutwama, J.J.; Mbidde, E.K.; Weyer, J.; Paweska, J.T.; Joloba, M.L.; Wayengera, M. Identity and validity of conserved B cell epitopes of filovirus glycoprotein: Towards rapid diagnostic testing for Ebola and possibly Marburg virus disease. BMC Infect. Dis. 2018, 18, 498. [Google Scholar] [CrossRef] [PubMed]
- Warfield, K.L.; Howell, K.A.; Vu, H.; Geisbert, J.; Wong, G.; Shulenin, S.; Sproule, S.; Holtsberg, F.W.; Leung, D.W.; Amarasinghe, G.K.; et al. Role of Antibodies in Protection Against Ebola Virus in Nonhuman Primates Immunized with Three Vaccine Platforms. J. Infect. Dis. 2018, 218 (Suppl. S5), S553–S564. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Formulation | Doses | Challenge | n | Survivors | Non-Survivors |
|---|---|---|---|---|---|
| 25 μg EBOV GP + CoVaccine HTTM | 3 | EBOV—week 10 challenge | 6 | 6 | 0 |
| 25 μg EBOV GP + CoVaccine HTTM | 3 | EBOV—week 62 challenge | 6 | 2 | 4 |
| 25 μg EBOV GP + CoVaccine HTTM | 2 | EBOV | 4 | 3 | 1 |
| 25 μg EBOV GP + 25 μg MARV GP + 25 μg SUDV GP + CoVaccine HTTM | 2 | EBOV | 3 | 0 | 3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Williams, C.A.; Wong, T.A.S.; Lieberman, M.M.; Yalley-Ogunro, J.; Cabus, M.; Nezami, S.; Paz, F.; Andersen, H.; Geisbert, T.W.; Lehrer, A.T. High-Avidity Anti-Filovirus IgG Elicited Using Protein Subunit Vaccines Does Not Correlate with Protection. Immuno 2023, 3, 358-374. https://doi.org/10.3390/immuno3040022
Williams CA, Wong TAS, Lieberman MM, Yalley-Ogunro J, Cabus M, Nezami S, Paz F, Andersen H, Geisbert TW, Lehrer AT. High-Avidity Anti-Filovirus IgG Elicited Using Protein Subunit Vaccines Does Not Correlate with Protection. Immuno. 2023; 3(4):358-374. https://doi.org/10.3390/immuno3040022
Chicago/Turabian StyleWilliams, Caitlin A., Teri Ann S. Wong, Michael M. Lieberman, Jake Yalley-Ogunro, Mehtap Cabus, Sara Nezami, Fabian Paz, Hanne Andersen, Thomas W. Geisbert, and Axel T. Lehrer. 2023. "High-Avidity Anti-Filovirus IgG Elicited Using Protein Subunit Vaccines Does Not Correlate with Protection" Immuno 3, no. 4: 358-374. https://doi.org/10.3390/immuno3040022
APA StyleWilliams, C. A., Wong, T. A. S., Lieberman, M. M., Yalley-Ogunro, J., Cabus, M., Nezami, S., Paz, F., Andersen, H., Geisbert, T. W., & Lehrer, A. T. (2023). High-Avidity Anti-Filovirus IgG Elicited Using Protein Subunit Vaccines Does Not Correlate with Protection. Immuno, 3(4), 358-374. https://doi.org/10.3390/immuno3040022