
Oral Administration of 5-Aminolevulinic Acid Does Not Ameliorate Autoimmune Diabetes in NOD Mice
 , , , , , , ,
, , , , , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Procedures
2.2. Administration of 5-ALA
2.3. Monitoring the Development of Spontaneous Diabetes
2.4. Measurement of Insulin Autoantibodies
2.5. Histology
2.6. Islet Isolation
2.7. Flow Cytometry
2.8. Adoptive Transfer Experiments
2.9. Cell Culture and Carboxyfluorescein Succinimidyl Ester (CFSE) Labeling
2.10. Statistical Analysis
3. Results
3.1. Oral Administration of 5-ALA Did Not Prevent the Development of Diabetes in NOD Mice
3.2. The IAA Production and Severity of Insulitis Were Not Suppressed by 5-ALA Administration in NOD Mice
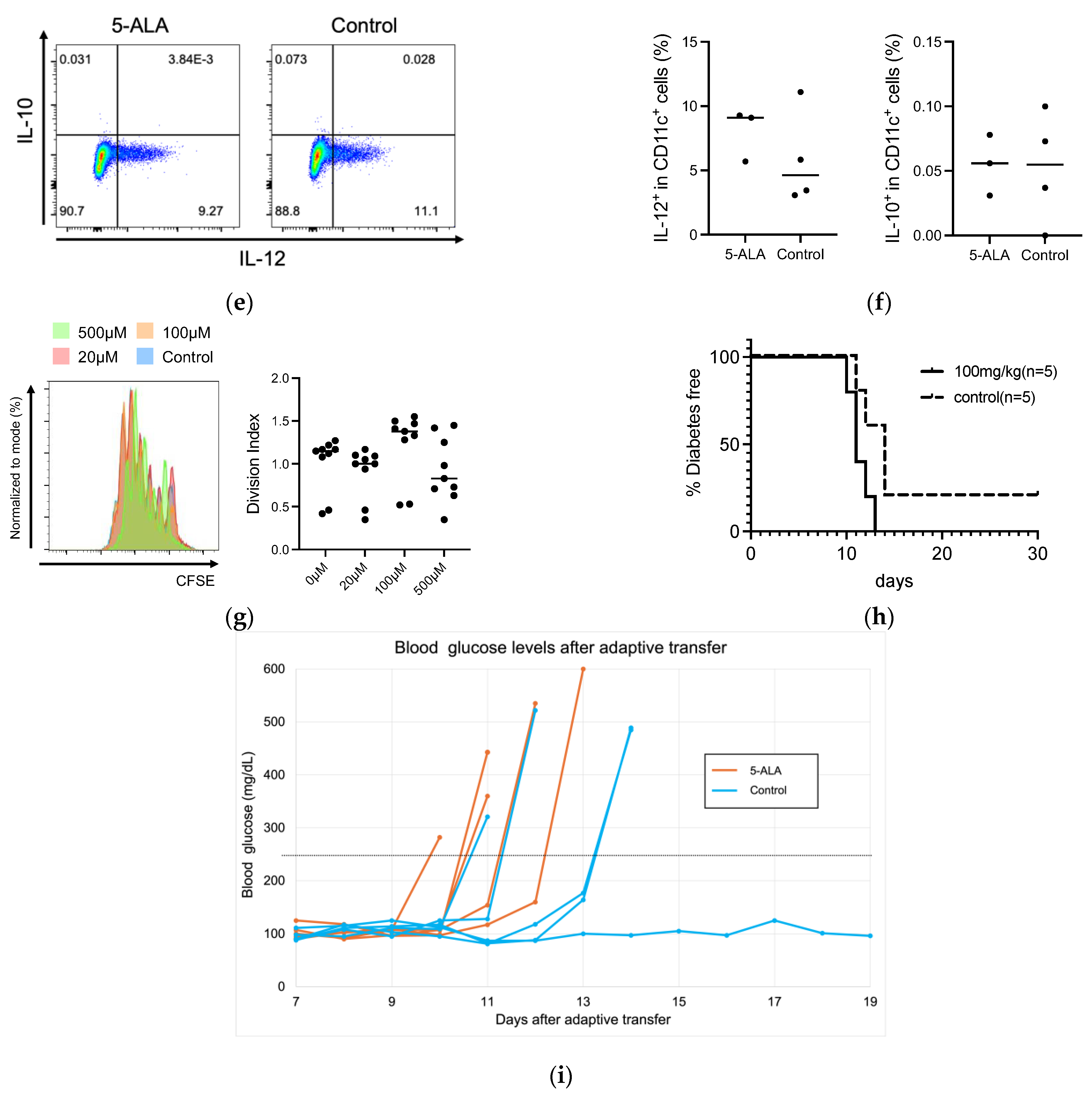
3.3. HO-1 Expression and Effector Function in Immune Cells Remained Unchanged by Short-Term 5-ALA Administration
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 5-ALA | 5-Aminolevulinic Acid |
| CFSE | Carboxyfluorescein succinimidyl ester |
| CoPP | Cobalt protoporphyrin IX |
| HO-1 | Home oxygenase-1 |
| IAA | Insulin autoantibody |
| IHC | Immunohistochemistry |
| NOD | Nonobese diabetic |
| SFC | Sodium ferrous citrate |
| T1D | Type 1 diabetes |
References
- Bluestone, J.A.; Herold, K.; Eisenbarth, G. Genetics, pathogenesis and clinical interventions in type 1 diabetes. Nature 2010, 464, 1293–1300. [Google Scholar] [CrossRef] [PubMed]
- Bluestone, J.A.; Buckner, J.H.; Herold, K.C. Immunotherapy: Building a bridge to a cure for type 1 diabetes. Science 2021, 373, 510–516. [Google Scholar] [CrossRef]
- Herold, K.C.; Bundy, B.N.; Long, S.A.; Bluestone, J.A.; DiMeglio, L.A.; Dufort, M.J.; Gitelman, S.E.; Gottlieb, P.A.; Krischer, J.P.; Linsley, P.S.; et al. An Anti-CD3 Antibody, Teplizumab, in Relatives at Risk for Type 1 Diabetes. N. Engl. J. Med. 2019, 381, 603–613. [Google Scholar] [CrossRef]
- Ramos, E.L.; Dayan, C.M.; Chatenoud, L.; Sumnik, Z.; Simmons, K.M.; Szypowska, A.; Gitelman, S.E.; Knecht, L.A.; Niemoeller, E.; Tian, W.; et al. Teplizumab and beta-Cell Function in Newly Diagnosed Type 1 Diabetes. N. Engl. J. Med. 2023, 389, 2151–2161. [Google Scholar] [CrossRef] [PubMed]
- Rehani, P.R.; Iftikhar, H.; Nakajima, M.; Tanaka, T.; Jabbar, Z.; Rehani, R.N. Safety and Mode of Action of Diabetes Medications in comparison with 5-Aminolevulinic Acid (5-ALA). J. Diabetes Res. 2019, 2019, 4267357. [Google Scholar] [CrossRef] [PubMed]
- Higashikawa, F.; Noda, M.; Awaya, T.; Tanaka, T.; Sugiyama, M. 5-aminolevulinic acid, a precursor of heme, reduces both fasting and postprandial glucose levels in mildly hyperglycemic subjects. Nutrition 2013, 29, 1030–1036. [Google Scholar] [CrossRef]
- Fujino, M.; Nishio, Y.; Ito, H.; Tanaka, T.; Li, X.K. 5-Aminolevulinic acid regulates the inflammatory response and alloimmune reaction. Int. Immunopharmacol. 2016, 37, 71–78. [Google Scholar] [CrossRef]
- Nishio, Y.; Fujino, M.; Zhao, M.; Ishii, T.; Ishizuka, M.; Ito, H.; Takahashi, K.; Abe, F.; Nakajima, M.; Tanaka, T.; et al. 5-Aminolevulinic acid combined with ferrous iron enhances the expression of heme oxygenase-1. Int. Immunopharmacol. 2014, 19, 300–307. [Google Scholar] [CrossRef]
- Moreau, A.; Hill, M.; Thebault, P.; Deschamps, J.Y.; Chiffoleau, E.; Chauveau, C.; Moullier, P.; Anegon, I.; Alliot-Licht, B.; Cuturi, M.C. Tolerogenic dendritic cells actively inhibit T cells through heme oxygenase-1 in rodents and in nonhuman primates. FASEB J. 2009, 23, 3070–3077. [Google Scholar] [CrossRef]
- Chauveau, C.; Remy, S.; Royer, P.J.; Hill, M.; Tanguy-Royer, S.; Hubert, F.X.; Tesson, L.; Brion, R.; Beriou, G.; Gregoire, M.; et al. Heme oxygenase-1 expression inhibits dendritic cell maturation and proinflammatory function but conserves IL-10 expression. Blood 2005, 106, 1694–1702. [Google Scholar] [CrossRef]
- Listopad, J.; Asadullah, K.; Sievers, C.; Ritter, T.; Meisel, C.; Sabat, R.; Docke, W.D. Heme oxygenase-1 inhibits T cell-dependent skin inflammation and differentiation and function of antigen-presenting cells. Exp. Dermatol. 2007, 16, 661–670. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, A.; Wafula, P.O.; Teles, A.; El-Mousleh, T.; Linzke, N.; Zenclussen, M.L.; Langwisch, S.; Heinze, K.; Wollenberg, I.; Casalis, P.A.; et al. Blockage of heme oxygenase-1 abrogates the protective effect of regulatory T cells on murine pregnancy and promotes the maturation of dendritic cells. PLoS ONE 2012, 7, e42301. [Google Scholar] [CrossRef]
- Al-Huseini, L.M.; Aw Yeang, H.X.; Hamdam, J.M.; Sethu, S.; Alhumeed, N.; Wong, W.; Sathish, J.G. Heme oxygenase-1 regulates dendritic cell function through modulation of p38 MAPK-CREB/ATF1 signaling. J. Biol. Chem. 2014, 289, 16442–16451. [Google Scholar] [CrossRef] [PubMed]
- George, J.F.; Braun, A.; Brusko, T.M.; Joseph, R.; Bolisetty, S.; Wasserfall, C.H.; Atkinson, M.A.; Agarwal, A.; Kapturczak, M.H. Suppression by CD4+CD25+ regulatory T cells is dependent on expression of heme oxygenase-1 in antigen-presenting cells. Am. J. Pathol. 2008, 173, 154–160. [Google Scholar] [CrossRef]
- Campbell, N.K.; Fitzgerald, H.K.; Malara, A.; Hambly, R.; Sweeney, C.M.; Kirby, B.; Fletcher, J.M.; Dunne, A. Naturally derived Heme-Oxygenase 1 inducers attenuate inflammatory responses in human dendritic cells and T cells: Relevance for psoriasis treatment. Sci. Rep. 2018, 8, 10287. [Google Scholar] [CrossRef]
- Yang, Y.; Zhang, X.; Xu, M.; Wu, X.; Zhao, F.; Zhao, C. Quercetin attenuates collagen-induced arthritis by restoration of Th17/Treg balance and activation of Heme Oxygenase 1-mediated anti-inflammatory effect. Int. Immunopharmacol. 2018, 54, 153–162. [Google Scholar] [CrossRef]
- Geisel, J.; Bruck, J.; Glocova, I.; Dengler, K.; Sinnberg, T.; Rothfuss, O.; Walter, M.; Schulze-Osthoff, K.; Rocken, M.; Ghoreschi, K. Sulforaphane protects from T cell-mediated autoimmune disease by inhibition of IL-23 and IL-12 in dendritic cells. J. Immunol. 2014, 192, 3530–3539. [Google Scholar] [CrossRef] [PubMed]
- Ghoreschi, K.; Bruck, J.; Kellerer, C.; Deng, C.; Peng, H.; Rothfuss, O.; Hussain, R.Z.; Gocke, A.R.; Respa, A.; Glocova, I.; et al. Fumarates improve psoriasis and multiple sclerosis by inducing type II dendritic cells. J. Exp. Med. 2011, 208, 2291–2303. [Google Scholar] [CrossRef]
- Yan, S.C.; Wang, Y.J.; Li, Y.J.; Cai, W.Y.; Weng, X.G.; Li, Q.; Chen, Y.; Yang, Q.; Zhu, X.X. Dihydroartemisinin Regulates the Th/Treg Balance by Inducing Activated CD4+ T cell Apoptosis via Heme Oxygenase-1 Induction in Mouse Models of Inflammatory Bowel Disease. Molecules 2019, 24, 2475. [Google Scholar] [CrossRef]
- Hu, C.M.; Lin, H.H.; Chiang, M.T.; Chang, P.F.; Chau, L.Y. Systemic expression of heme oxygenase-1 ameliorates type 1 diabetes in NOD mice. Diabetes 2007, 56, 1240–1247. [Google Scholar] [CrossRef]
- Li, M.; Peterson, S.; Husney, D.; Inaba, M.; Guo, K.; Kappas, A.; Ikehara, S.; Abraham, N.G. Long-lasting expression of HO-1 delays progression of type I diabetes in NOD mice. Cell Cycle 2007, 6, 567–571. [Google Scholar] [CrossRef]
- Simon, T.; Pogu, J.; Remy, S.; Brau, F.; Pogu, S.; Maquigneau, M.; Fonteneau, J.F.; Poirier, N.; Vanhove, B.; Blancho, G.; et al. Inhibition of effector antigen-specific T cells by intradermal administration of heme oxygenase-1 inducers. J. Autoimmun. 2017, 81, 44–55. [Google Scholar] [CrossRef] [PubMed]
- Saitoh, S.; Okano, S.; Nohara, H.; Nakano, H.; Shirasawa, N.; Naito, A.; Yamamoto, M.; Kelly, V.P.; Takahashi, K.; Tanaka, T.; et al. 5-aminolevulinic acid (ALA) deficiency causes impaired glucose tolerance and insulin resistance coincident with an attenuation of mitochondrial function in aged mice. PLoS ONE 2018, 13, e0189593. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Yang, X.; Zhu, P.; Fujino, M.; Ito, H.; Takahashi, K.; Nakajima, M.; Tanaka, T.; Wang, J.; Zhuang, J.; et al. Combination of 5-aminolevulinic acid and iron prevents skin fibrosis in murine sclerodermatous graft-versus-host disease. Exp. Dermatol. 2018, 27, 1104–1111. [Google Scholar] [CrossRef] [PubMed]
- Koga, T.; Sumiyoshi, R.; Tsuji, Y.; Kodama, K.; Endo, Y.; Furukawa, K.; Kawakami, A. Efficacy and safety of 5-aminolevulinic acid in adult-onset Still’s disease: A preclinical study in mice and a pilot study in humans. Clin. Immunol. 2023, 257, 109846. [Google Scholar] [CrossRef]
- Jhala, G.; Chee, J.; Trivedi, P.M.; Selck, C.; Gurzov, E.N.; Graham, K.L.; Thomas, H.E.; Kay, T.W.; Krishnamurthy, B. Perinatal tolerance to proinsulin is sufficient to prevent autoimmune diabetes. JCI Insight 2016, 1, e86065. [Google Scholar] [CrossRef]
- Akazawa, S.; Kobayashi, M.; Kuriya, G.; Horie, I.; Yu, L.; Yamasaki, H.; Okita, M.; Nagayama, Y.; Matsuyama, T.; Akbari, M.; et al. Haploinsufficiency of interferon regulatory factor 4 strongly protects against autoimmune diabetes in NOD mice. Diabetologia 2015, 58, 2606–2614. [Google Scholar] [CrossRef]
- Graham, K.L.; Fynch, S.; Papas, E.G.; Tan, C.; Kay, T.W.H.; Thomas, H.E. Isolation and Culture of the Islets of Langerhans from Mouse Pancreas. Bio-Protoc. 2016, 6, e1840. [Google Scholar] [CrossRef]
- Niri, T.; Inoue, S.I.; Akazawa, S.; Nishikido, S.; Miwa, M.; Kobayashi, M.; Yui, K.; Okita, M.; Kawakami, A.; Abiru, N. Essential role of interferon-regulatory factor 4 in regulating diabetogenic CD4+ T and innate immune cells in autoimmune diabetes in NOD mice. Clin. Exp. Immunol. 2024, 219, uxae093. [Google Scholar] [CrossRef]
- Jin, N.; Wang, Y.; Crawford, F.; White, J.; Marrack, P.; Dai, S.; Kappler, J.W. N-terminal additions to the WE14 peptide of chromogranin A create strong autoantigen agonists in type 1 diabetes. Proc. Natl. Acad. Sci. USA 2015, 112, 13318–13323. [Google Scholar] [CrossRef]
- Pearson, J.A.; Wong, F.S.; Wen, L. The importance of the Non Obese Diabetic (NOD) mouse model in autoimmune diabetes. J. Autoimmun. 2016, 66, 76–88. [Google Scholar] [CrossRef] [PubMed]
- Andre, I.; Gonzalez, A.; Wang, B.; Katz, J.; Benoist, C.; Mathis, D. Checkpoints in the progression of autoimmune disease: Lessons from diabetes models. Proc. Natl. Acad. Sci. USA 1996, 93, 2260–2263. [Google Scholar] [CrossRef] [PubMed]
- Delovitch, T.L.; Singh, B. The nonobese diabetic mouse as a model of autoimmune diabetes: Immune dysregulation gets the NOD. Immunity 1997, 7, 727–738. [Google Scholar] [CrossRef] [PubMed]
- Abiru, N.; Yu, L.; Miao, D.; Maniatis, A.K.; Liu, E.; Moriyama, H.; Eisenbarth, G.S. Transient insulin autoantibody expression independent of development of diabetes: Comparison of NOD and NOR strains. J. Autoimmun. 2001, 17, 1–6. [Google Scholar] [CrossRef]
- Yang, X.; Ma, Z.; Tan, X.; Shi, Y.; Yuan, M.; Chen, G.; Luo, X.; Hou, L. Adoptive transfer of immature dendritic cells with high HO-1 expression delays the onset of T1DM in NOD mice. Life Sci. 2023, 335, 122273. [Google Scholar] [CrossRef]
- Passeri, L.; Andolfi, G.; Bassi, V.; Russo, F.; Giacomini, G.; Laudisa, C.; Marrocco, I.; Cesana, L.; Di Stefano, M.; Fanti, L.; et al. Tolerogenic IL-10-engineered dendritic cell-based therapy to restore antigen-specific tolerance in T cell mediated diseases. J. Autoimmun. 2023, 138, 103051. [Google Scholar] [CrossRef]
- Waibel, M.; Wentworth, J.M.; So, M.; Couper, J.J.; Cameron, F.J.; MacIsaac, R.J.; Atlas, G.; Gorelik, A.; Litwak, S.; Sanz-Villanueva, L.; et al. Baricitinib and beta-Cell Function in Patients with New-Onset Type 1 Diabetes. N. Engl. J. Med. 2023, 389, 2140–2150. [Google Scholar] [CrossRef]
- Pescovitz, M.D.; Greenbaum, C.J.; Krause-Steinrauf, H.; Becker, D.J.; Gitelman, S.E.; Goland, R.; Gottlieb, P.A.; Marks, J.B.; McGee, P.F.; Moran, A.M.; et al. Rituximab, B-lymphocyte depletion, and preservation of beta-cell function. N. Engl. J. Med. 2009, 361, 2143–2152. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nishikido, S.; Akazawa, S.; Niri, T.; Inoue, S.-I.; Matsuda, K.; Aoshi, T.; Nakashima, M.; Haraguchi, A.; Horie, I.; Kobayashi, M.; et al. Oral Administration of 5-Aminolevulinic Acid Does Not Ameliorate Autoimmune Diabetes in NOD Mice. Diabetology 2025, 6, 62. https://doi.org/10.3390/diabetology6070062
Nishikido S, Akazawa S, Niri T, Inoue S-I, Matsuda K, Aoshi T, Nakashima M, Haraguchi A, Horie I, Kobayashi M, et al. Oral Administration of 5-Aminolevulinic Acid Does Not Ameliorate Autoimmune Diabetes in NOD Mice. Diabetology. 2025; 6(7):62. https://doi.org/10.3390/diabetology6070062
Chicago/Turabian StyleNishikido, Shinpei, Satoru Akazawa, Tetsuro Niri, Shin-Ichi Inoue, Katsuya Matsuda, Taiki Aoshi, Masahiro Nakashima, Ai Haraguchi, Ichiro Horie, Masakazu Kobayashi, and et al. 2025. "Oral Administration of 5-Aminolevulinic Acid Does Not Ameliorate Autoimmune Diabetes in NOD Mice" Diabetology 6, no. 7: 62. https://doi.org/10.3390/diabetology6070062
APA StyleNishikido, S., Akazawa, S., Niri, T., Inoue, S.-I., Matsuda, K., Aoshi, T., Nakashima, M., Haraguchi, A., Horie, I., Kobayashi, M., Okita, M., Kawakami, A., & Abiru, N. (2025). Oral Administration of 5-Aminolevulinic Acid Does Not Ameliorate Autoimmune Diabetes in NOD Mice. Diabetology, 6(7), 62. https://doi.org/10.3390/diabetology6070062