
Impact of Dietary Sugars on β-Cell Function

 ,
,  , , and
, , and 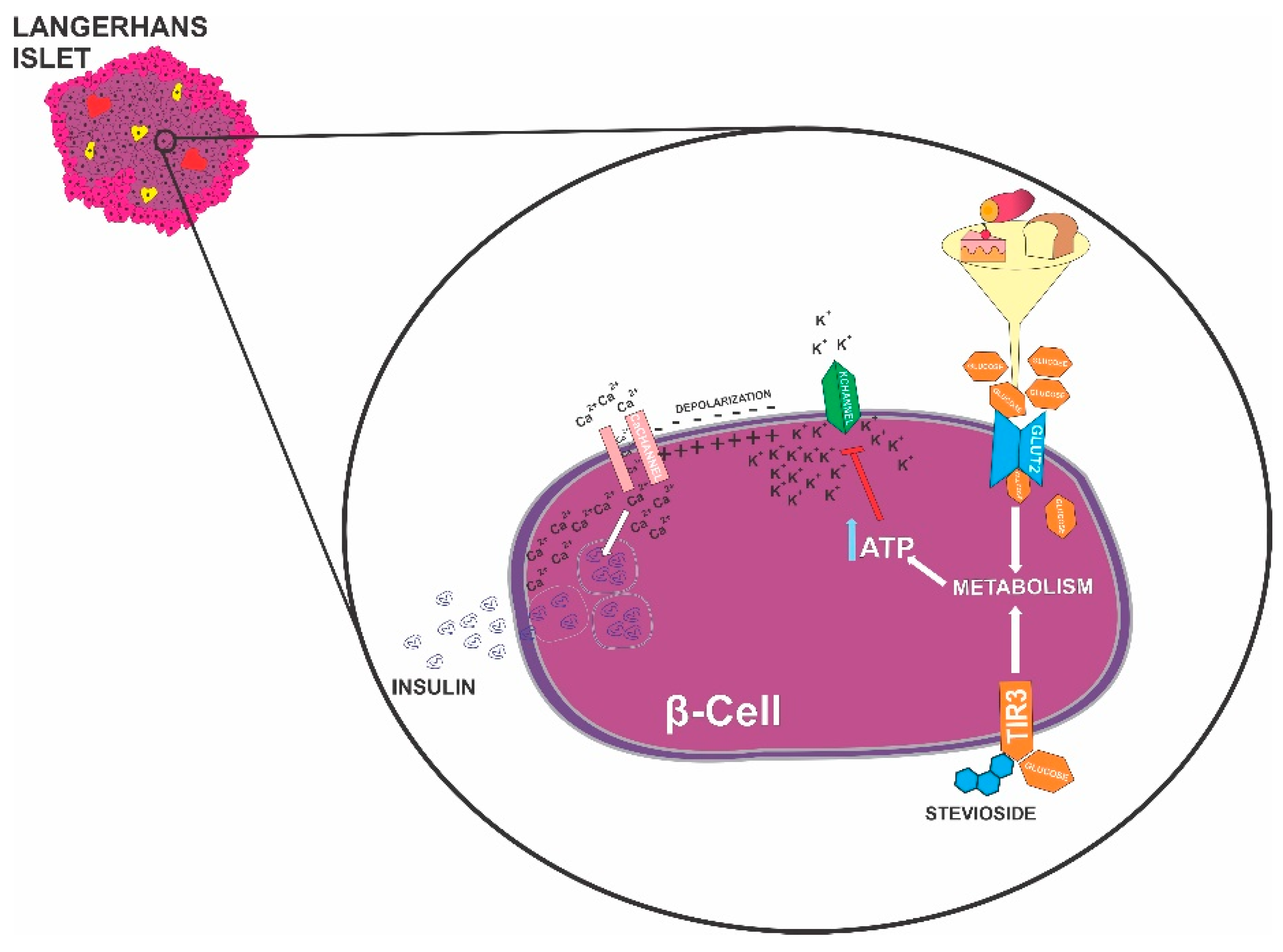{kind=link}
Abstract
1. Introduction
2. Endocrine Pancreas
3. β-Cells Burn Sugar to Provide Fuel to other Cells
4. Great Conflict: Fuel Hypothesis vs. Glucose Receptor
5. β-Cells Sense Sweet, Bitter, Umami, and Salty Taste
6. Sugar Sources Potentially Transport Poisons or Medicines to β-Cells
7. Environment as Vectorial to β-Cell
8. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ashcroft, F.M.; Rorsman, P. Diabetes mellitus and the beta cell: The last ten years. Cell 2012, 148, 1160–1171. [Google Scholar] [CrossRef]
- Eizirik, D.L.; Pasquali, L.; Cnop, M. Pancreatic beta-cells in type 1 and type 2 diabetes mellitus: Different pathways to failure. Nat. Rev. Endocrinol. 2020, 16, 349–362. [Google Scholar] [CrossRef] [PubMed]
- McKeown, N.M.; Meigs, J.B.; Liu, S.; Saltzman, E.; Wilson, P.W.; Jacques, P.F. Carbohydrate nutrition, insulin resistance, and the prevalence of the metabolic syndrome in the Framingham Offspring Cohort. Diabetes Care 2004, 27, 538–546. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.N.; Ventura, E.E.; Weigensberg, M.J.; Ball, G.D.; Cruz, M.L.; Shaibi, G.Q.; Goran, M.I. The relation of sugar intake to beta cell function in overweight Latino children. Am. J. Clin. Nutr. 2005, 82, 1004–1010. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, P.E.; Rorsman, P. Oscillations, intercellular coupling, and insulin secretion in pancreatic beta cells. PLoS Biol. 2006, 4, e49. [Google Scholar] [CrossRef]
- Bock, T.; Pakkenberg, B.; Buschard, K. Genetic background determines the size and structure of the endocrine pancreas. Diabetes 2005, 54, 133–137. [Google Scholar] [CrossRef] [PubMed]
- Cnop, M.; Welsh, N.; Jonas, J.C.; Jorns, A.; Lenzen, S.; Eizirik, D.L. Mechanisms of pancreatic beta-cell death in type 1 and type 2 diabetes: Many differences, few similarities. Diabetes 2005, 54 (Suppl. 2), S97–S107. [Google Scholar] [CrossRef] [PubMed]
- Rorsman, P.; Ashcroft, F.M. Pancreatic Beta-Cell Electrical Activity and Insulin Secretion: Of Mice and Men. Physiol. Rev. 2018, 98, 117–214. [Google Scholar] [CrossRef]
- Mourad, N.I.; Nenquin, M.; Henquin, J.C. Metabolic amplifying pathway increases both phases of insulin secretion independently of beta-cell actin microfilaments. Am. J. Physiol. Cell Physiol. 2010, 299, C389–C398. [Google Scholar] [CrossRef]
- Teraoku, H.; Lenzen, S. Dynamics of Insulin Secretion from EndoC-betaH1 beta-Cell Pseudoislets in Response to Glucose and Other Nutrient and Nonnutrient Secretagogues. J. Diabetes Res. 2017, 2017, 2309630. [Google Scholar] [CrossRef]
- Curry, D.L. Effects of mannose and fructose on the synthesis and secretion of insulin. Pancreas 1989, 4, 2–9. [Google Scholar] [CrossRef] [PubMed]
- Sener, A.; Malaisse, W.J. L-leucine and a nonmetabolized analogue activate pancreatic islet glutamate dehydrogenase. Nature 1980, 288, 187–189. [Google Scholar] [CrossRef]
- Malaisse, W.J. Insulin release: The receptor hypothesis. Diabetologia 2014, 57, 1287–1290. [Google Scholar] [CrossRef] [PubMed]
- von Molitor, E.; Riedel, K.; Krohn, M.; Rudolf, R.; Hafner, M.; Cesetti, T. An alternative pathway for sweet sensation: Possible mechanisms and physiological relevance. Pflugers Arch. 2020, 472, 1667–1691. [Google Scholar] [CrossRef]
- Abudula, R.; Jeppesen, P.B.; Rolfsen, S.E.; Xiao, J.; Hermansen, K. Rebaudioside A potently stimulates insulin secretion from isolated mouse islets: Studies on the dose-, glucose-, and calcium-dependency. Metabolism 2004, 53, 1378–1381. [Google Scholar] [CrossRef] [PubMed]
- Philippaert, K.; Pironet, A.; Mesuere, M.; Sones, W.; Vermeiren, L.; Kerselaers, S.; Pinto, S.; Segal, A.; Antoine, N.; Gysemans, C.; et al. Steviol glycosides enhance pancreatic beta-cell function and taste sensation by potentiation of TRPM5 channel activity. Nat. Commun. 2017, 8, 14733. [Google Scholar] [CrossRef] [PubMed]
- Kyriazis, G.A.; Soundarapandian, M.M.; Tyrberg, B. Sweet taste receptor signaling in beta cells mediates fructose-induced potentiation of glucose-stimulated insulin secretion. Proc. Natl. Acad. Sci. USA 2012, 109, E524–E532. [Google Scholar] [CrossRef]
- Malaisse, W.J.; Vanonderbergen, A.; Louchami, K.; Jijakli, H.; Malaisse-Lagae, F. Effects of artificial sweeteners on insulin release and cationic fluxes in rat pancreatic islets. Cell Signal. 1998, 10, 727–733. [Google Scholar] [CrossRef]
- Nakagawa, Y.; Nagasawa, M.; Yamada, S.; Hara, A.; Mogami, H.; Nikolaev, V.O.; Lohse, M.J.; Shigemura, N.; Ninomiya, Y.; Kojima, I. Sweet taste receptor expressed in pancreatic beta-cells activates the calcium and cyclic AMP signaling systems and stimulates insulin secretion. PLoS ONE 2009, 4, e5106. [Google Scholar] [CrossRef]
- Alam, Y.H.; Kim, R.; Jang, C. Metabolism and Health Impacts of Dietary Sugars. J. Lipid Atheroscler. 2022, 11, 20–38. [Google Scholar] [CrossRef]
- Oberhauser, L.; Jimenez-Sanchez, C.; Madsen, J.G.S.; Duhamel, D.; Mandrup, S.; Brun, T.; Maechler, P. Glucolipotoxicity promotes the capacity of the glycerolipid/NEFA cycle supporting the secretory response of pancreatic beta cells. Diabetologia 2022, 65, 705–720. [Google Scholar] [CrossRef]
- Rizkalla, S.W. Health implications of fructose consumption: A review of recent data. Nutr. Metab. 2010, 7, 82. [Google Scholar] [CrossRef]
- Chandrasekara, A.; Josheph Kumar, T. Roots and Tuber Crops as Functional Foods: A Review on Phytochemical Constituents and Their Potential Health Benefits. Int. J. Food Sci. 2016, 2016, 3631647. [Google Scholar] [CrossRef]
- Cherbut, C.; Bruley des Varannes, S.; Schnee, M.; Rival, M.; Galmiche, J.P.; Delort-Laval, J. Involvement of small intestinal motility in blood glucose response to dietary fibre in man. Br. J. Nutr. 1994, 71, 675–685. [Google Scholar] [CrossRef]
- Lattimer, J.M.; Haub, M.D. Effects of dietary fiber and its components on metabolic health. Nutrients 2010, 2, 1266–1289. [Google Scholar] [CrossRef]
- Schulze, M.B.; Liu, S.; Rimm, E.B.; Manson, J.E.; Willett, W.C.; Hu, F.B. Glycemic index, glycemic load, and dietary fiber intake and incidence of type 2 diabetes in younger and middle-aged women. Am. J. Clin. Nutr. 2004, 80, 348–356. [Google Scholar] [CrossRef]
- Chattopadhyay, S.; Raychaudhuri, U.; Chakraborty, R. Artificial sweeteners—A review. J. Food Sci. Technol. 2014, 51, 611–621. [Google Scholar] [CrossRef] [PubMed]
- Lourenco, S.C.; Moldao-Martins, M.; Alves, V.D. Antioxidants of Natural Plant Origins: From Sources to Food Industry Applications. Molecules 2019, 24, 4132. [Google Scholar] [CrossRef]
- Matsuoka, T.; Kajimoto, Y.; Watada, H.; Kaneto, H.; Kishimoto, M.; Umayahara, Y.; Fujitani, Y.; Kamada, T.; Kawamori, R.; Yamasaki, Y. Glycation-dependent, reactive oxygen species-mediated suppression of the insulin gene promoter activity in HIT cells. J. Clin. Investig. 1997, 99, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Wali, J.A.; Raubenheimer, D.; Senior, A.M.; Le Couteur, D.G.; Simpson, S.J. Cardio-metabolic consequences of dietary carbohydrates: Reconciling contradictions using nutritional geometry. Cardiovasc. Res. 2021, 117, 386–401. [Google Scholar] [CrossRef] [PubMed]
- Gautam, D.; Han, S.J.; Hamdan, F.F.; Jeon, J.; Li, B.; Li, J.H.; Cui, Y.; Mears, D.; Lu, H.; Deng, C.; et al. A critical role for beta cell M3 muscarinic acetylcholine receptors in regulating insulin release and blood glucose homeostasis in vivo. Cell Metab. 2006, 3, 449–461. [Google Scholar] [CrossRef]
- Lundquist, I.; Ericson, L.E. beta-Adrenergic insulin release and adrenergic innervation of mouse pancreatic islets. Cell Tissue Res. 1978, 193, 73–85. [Google Scholar] [CrossRef]
- Bray, G.A. Obesity, a disorder of nutrient partitioning: The MONA LISA hypothesis. J. Nutr. 1991, 121, 1146–1162. [Google Scholar] [CrossRef]
- Vozarova de Courten, B.; Weyer, C.; Stefan, N.; Horton, M.; DelParigi, A.; Havel, P.; Bogardus, C.; Tataranni, P.A. Parasympathetic blockade attenuates augmented pancreatic polypeptide but not insulin secretion in Pima Indians. Diabetes 2004, 53, 663–671. [Google Scholar] [CrossRef] [PubMed]
- Papazoglou, I.; Lee, J.H.; Cui, Z.; Li, C.; Fulgenzi, G.; Bahn, Y.J.; Staniszewska-Goraczniak, H.M.; Pinol, R.A.; Hogue, I.B.; Enquist, L.W.; et al. A distinct hypothalamus-to-beta cell circuit modulates insulin secretion. Cell Metab. 2022, 34, 285–298.e7. [Google Scholar] [CrossRef]
- Gaur, A.; Pal, G.K.; Pal, P. Role of Ventromedial Hypothalamus in Sucrose-Induced Obesity on Metabolic Parameters. Ann. Neurosci 2021, 28, 39–46. [Google Scholar] [CrossRef]
- Zhu, W.; Sahar, N.E.; Javaid, H.M.A.; Pak, E.S.; Liang, G.; Wang, Y.; Ha, H.; Huh, J.Y. Exercise-Induced Irisin Decreases Inflammation and Improves NAFLD by Competitive Binding with MD2. Cells 2021, 10, 3306. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.; Chen, N.; Kang, X.; Hu, Y.; Shi, S. Irisin alleviates FFA induced beta-cell insulin resistance and inflammatory response through activating PI3K/AKT/FOXO1 signaling pathway. Endocrine 2021, 75, 740–751. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.L.; Li, S.J.; Lee, T.I.; Lee, T.W.; Chung, C.C.; Kao, Y.H.; Chen, Y.J. Sugar Fructose Triggers Gut Dysbiosis and Metabolic Inflammation with Cardiac Arrhythmogenesis. Biomedicines 2021, 9, 728. [Google Scholar] [CrossRef]
- Almeida, D.L.; Pavanello, A.; Saavedra, L.P.; Pereira, T.S.; de Castro-Prado, M.A.A.; de Freitas Mathias, P.C. Environmental monitoring and the developmental origins of health and disease. J. Dev. Orig. Health Dis. 2019, 10, 608–615. [Google Scholar] [CrossRef]
- Fabricio, G.; Malta, A.; Chango, A.; De Freitas Mathias, P.C. Environmental Contaminants and Pancreatic Beta-Cells. J. Clin. Res. Pediatr. Endocrinol. 2016, 8, 257–263. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Malta, A.; Saavedra, L.P.J.; Raposo, S.R.; Lopes, G.K.G.; Fernandes, M.D.; Barbosa, L.F.; Almeida, D.L.; Mathias, P.C.d.F. Impact of Dietary Sugars on β-Cell Function. Diabetology 2023, 4, 178-183. https://doi.org/10.3390/diabetology4020017
Malta A, Saavedra LPJ, Raposo SR, Lopes GKG, Fernandes MD, Barbosa LF, Almeida DL, Mathias PCdF. Impact of Dietary Sugars on β-Cell Function. Diabetology. 2023; 4(2):178-183. https://doi.org/10.3390/diabetology4020017
Chicago/Turabian StyleMalta, Ananda, Lucas Paulo Jacinto Saavedra, Scarlett Rodrigues Raposo, Gabriel Kian Guimarães Lopes, Maryana Debossan Fernandes, Letícia Ferreira Barbosa, Douglas Lopes Almeida, and Paulo Cezar de Freitas Mathias. 2023. "Impact of Dietary Sugars on β-Cell Function" Diabetology 4, no. 2: 178-183. https://doi.org/10.3390/diabetology4020017
APA StyleMalta, A., Saavedra, L. P. J., Raposo, S. R., Lopes, G. K. G., Fernandes, M. D., Barbosa, L. F., Almeida, D. L., & Mathias, P. C. d. F. (2023). Impact of Dietary Sugars on β-Cell Function. Diabetology, 4(2), 178-183. https://doi.org/10.3390/diabetology4020017