Tempol Alters Antioxidant Enzyme Function, Modulates Multiple Genes Expression, and Ameliorates Hepatic and Renal Impairment in Carbon Tetrachloride (CCl4)-Intoxicated Rats

 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Experimental Animals
2.3. Treatment and Study Design
2.4. Biochemical Assays
2.4.1. Liver Toxicity Assessment
2.4.2. Malondialdehyde (MDA) Estimation
2.4.3. Nitric Oxide (NO) Estimation
2.4.4. Advanced Protein Oxidation Product (APOP) Determination
2.4.5. Estimation of Catalase Activity
2.4.6. Estimation of SOD Enzyme Activity
2.4.7. Myeloperoxidase (MPO) Activity Estimation
2.5. RT-PCR Analysis for Liver Sample for Oxidative Stress and Inflammation Related Genes Expression
2.6. Procedure of Histopathology
2.7. Statistical Analysis
3. Results
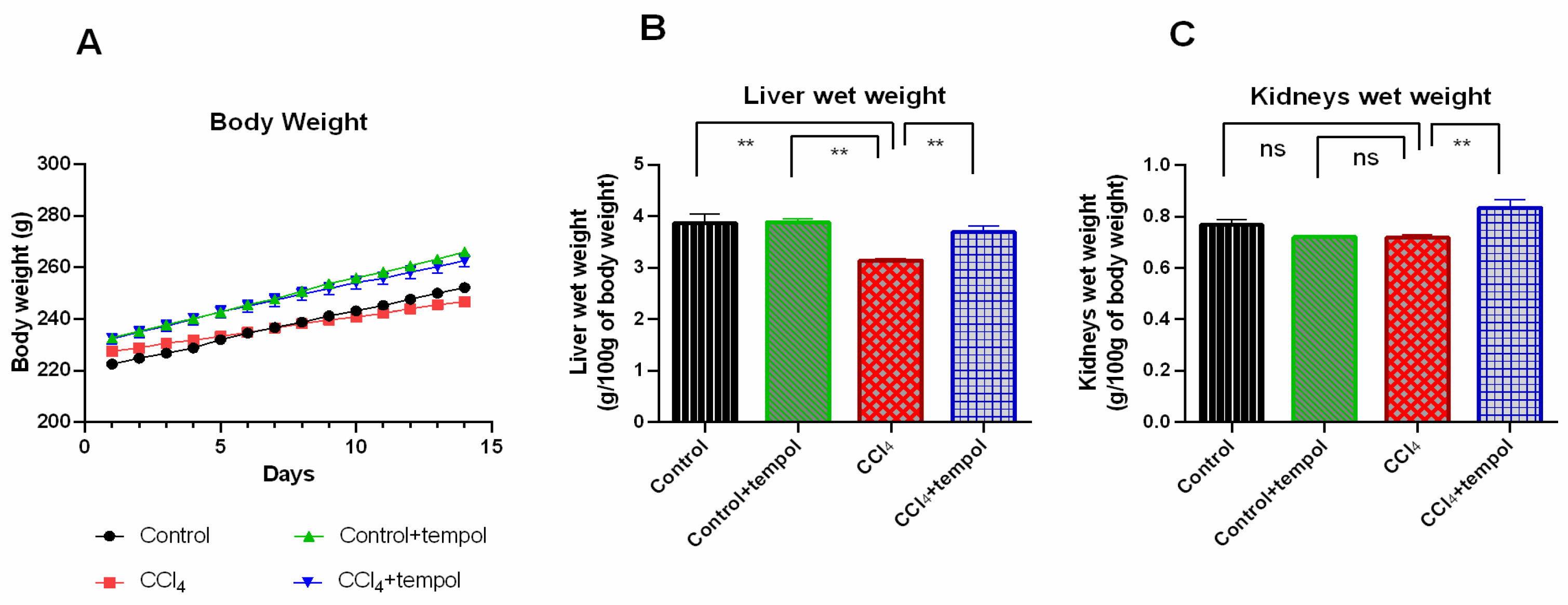
3.1. Tempol Administration Modulated the Body Weight, Liver Wet Weight, and Kidneys Wet Weight in CCl4-Administered Rats
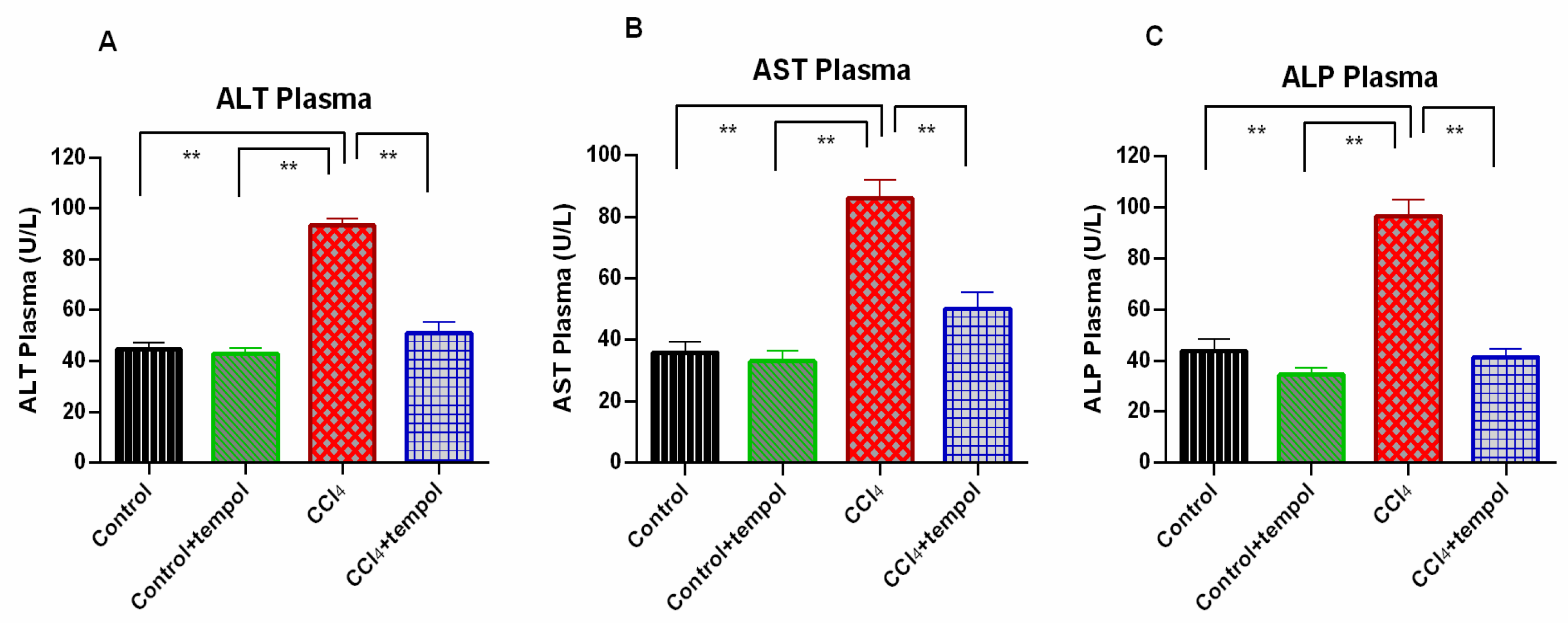
3.2. Tempol Administration Lowered the ALT, AST, and ALP Enzymes Activities in Plasma of CCl4-Administered Rats
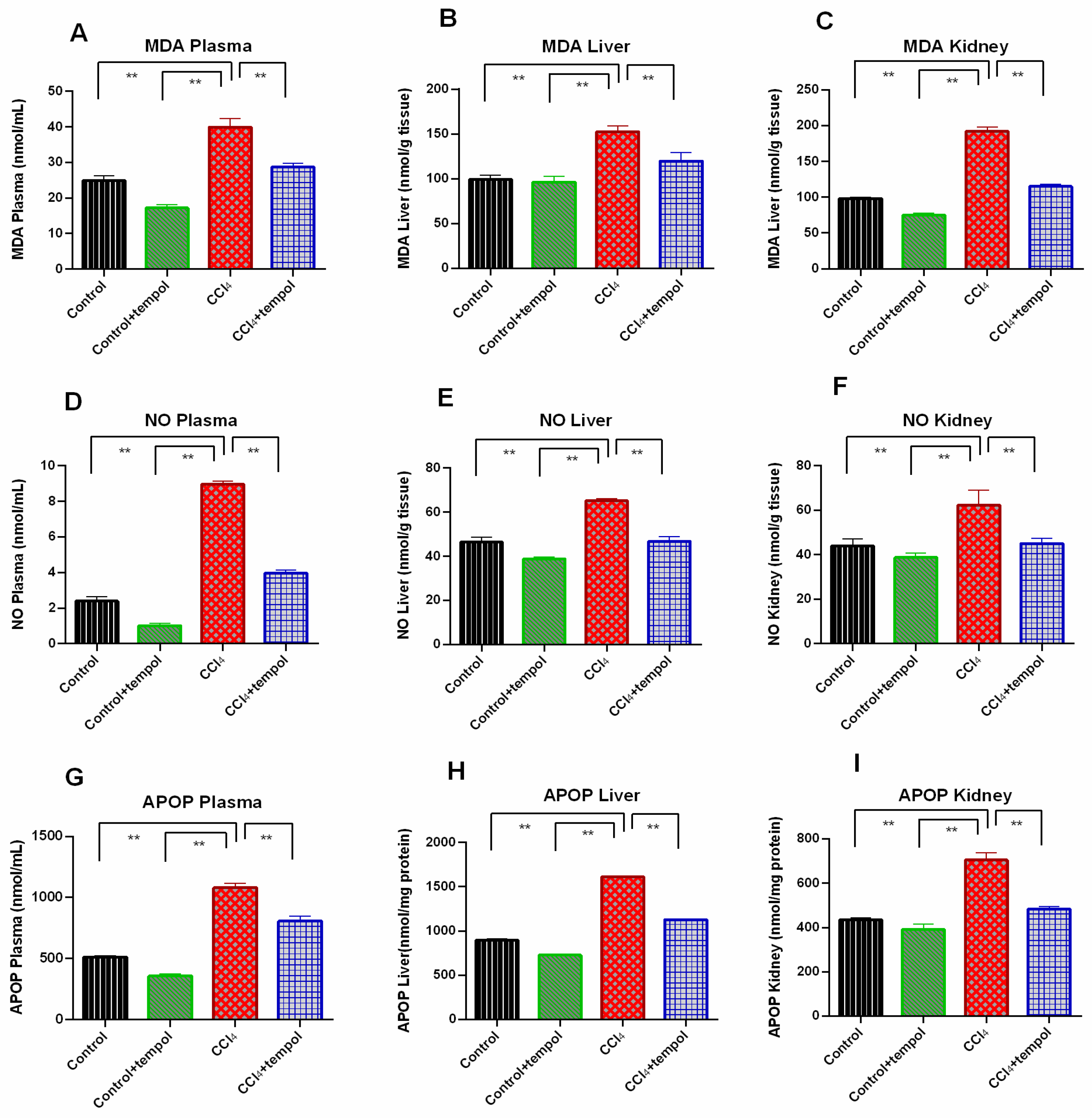
3.3. Tempol Treatment Prevented the Oxidative Stress and Lowered the MDA, NO and AOPP Level in CCl4-Administered Rats
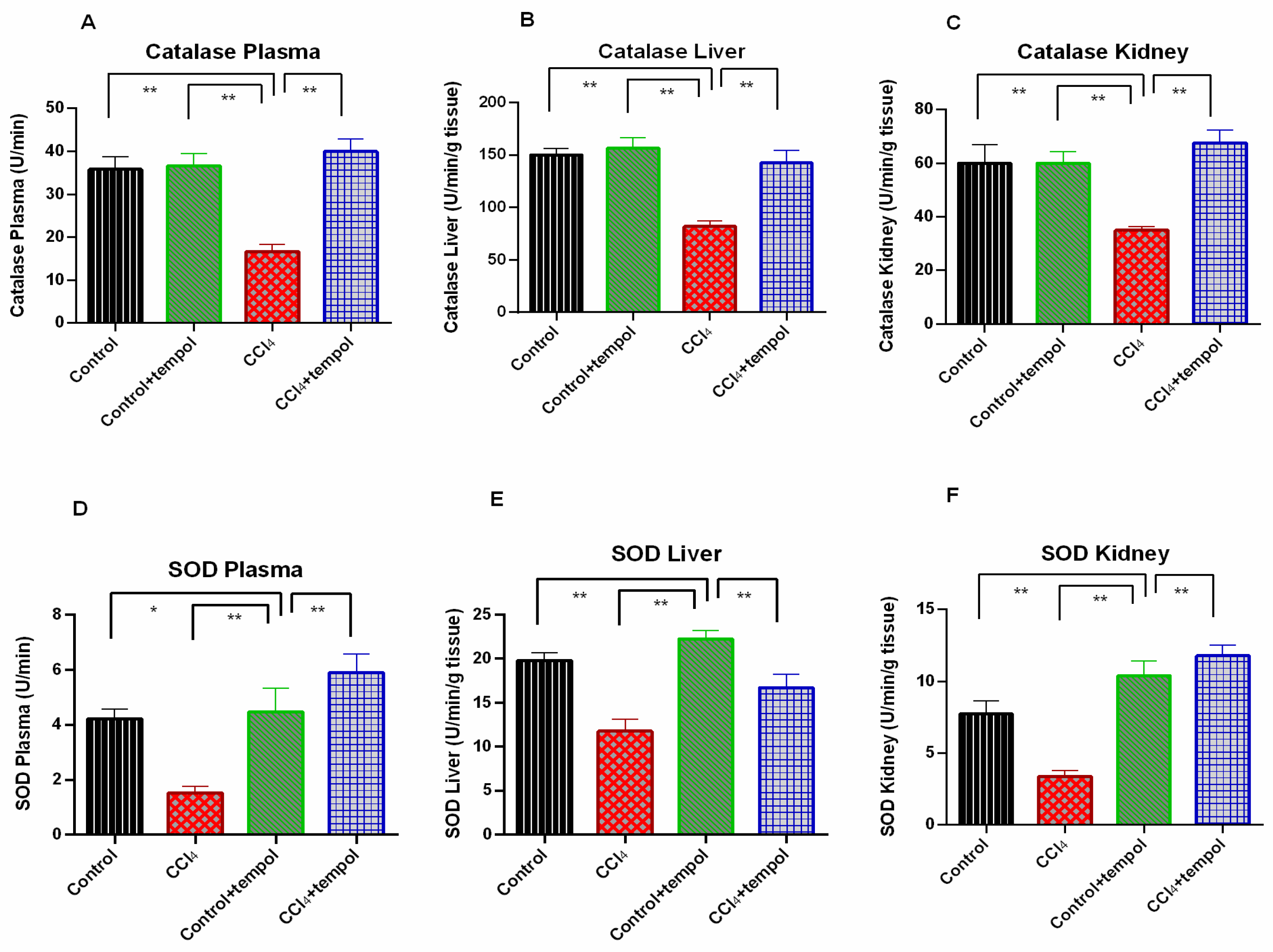
3.4. Tempol Administration in CCl4-Administered rats Restored the Antioxidant Enzymes, Catalase and SOD Activities
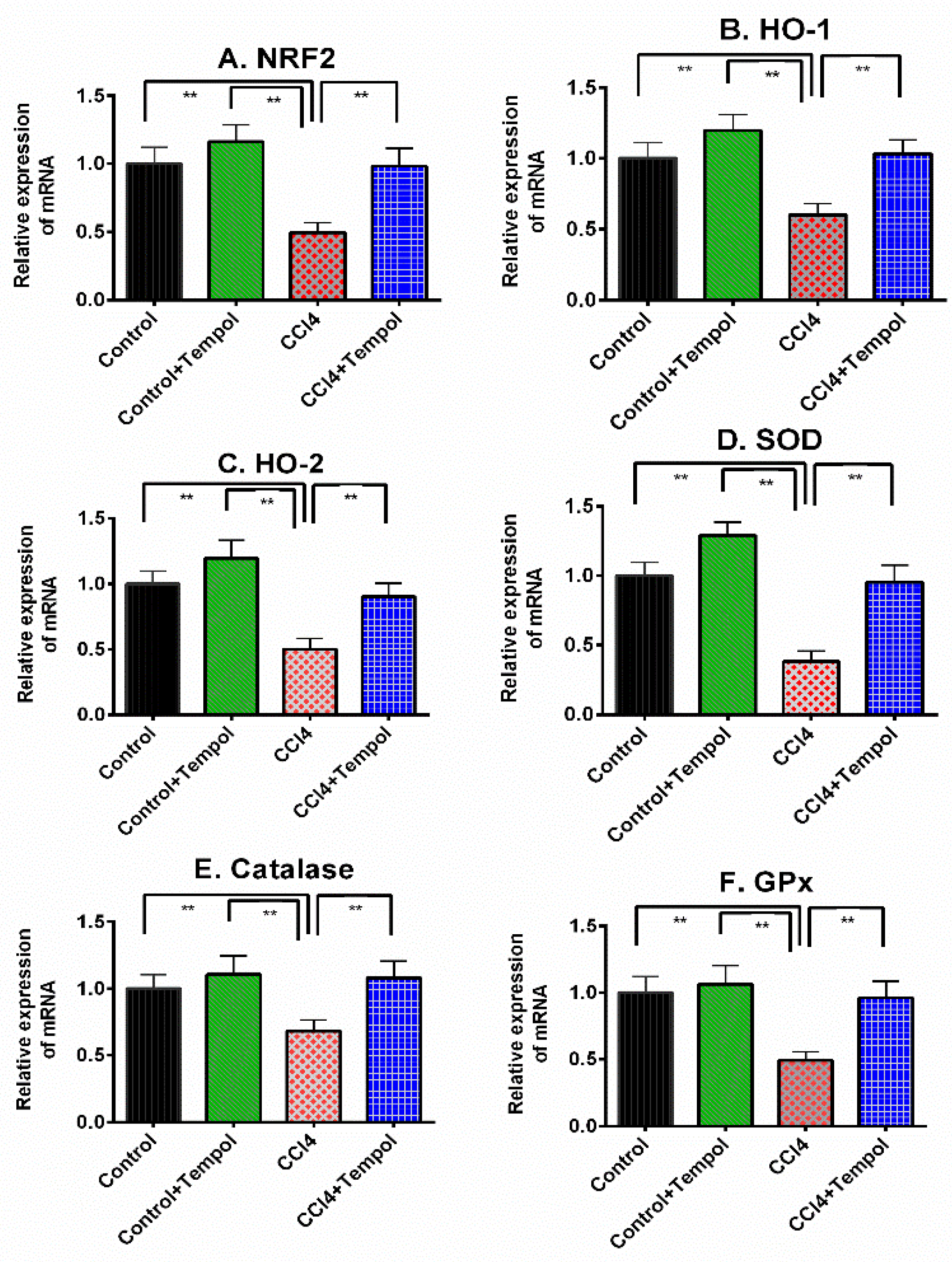
3.5. Tempol Administration Modulated the Antioxidant Genes Expression in Liver of CCl4-Administered Rats
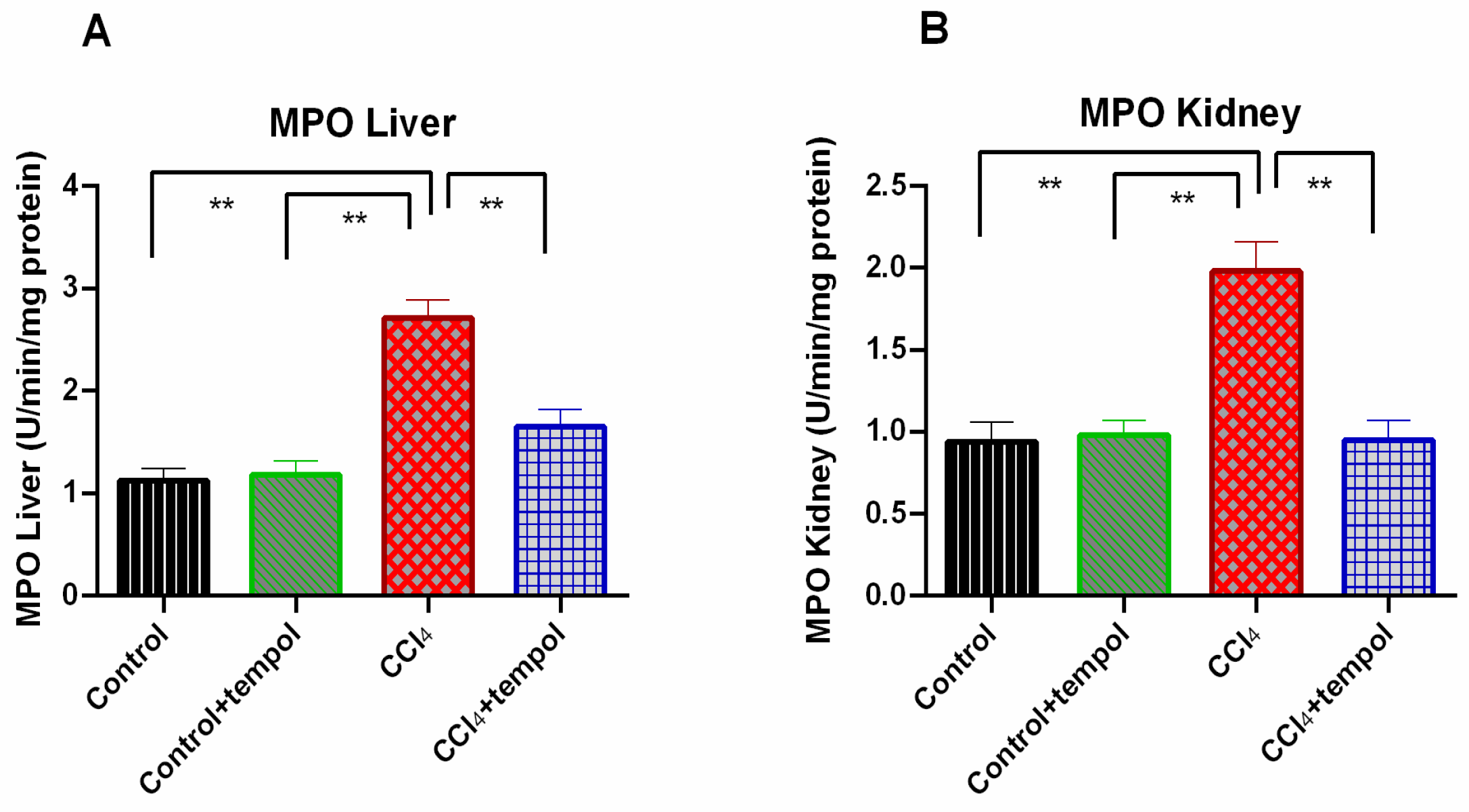
3.6. Tempol Treatment Lowered the MPO Activity in Liver and Kidney of CCl4-Administered Rats
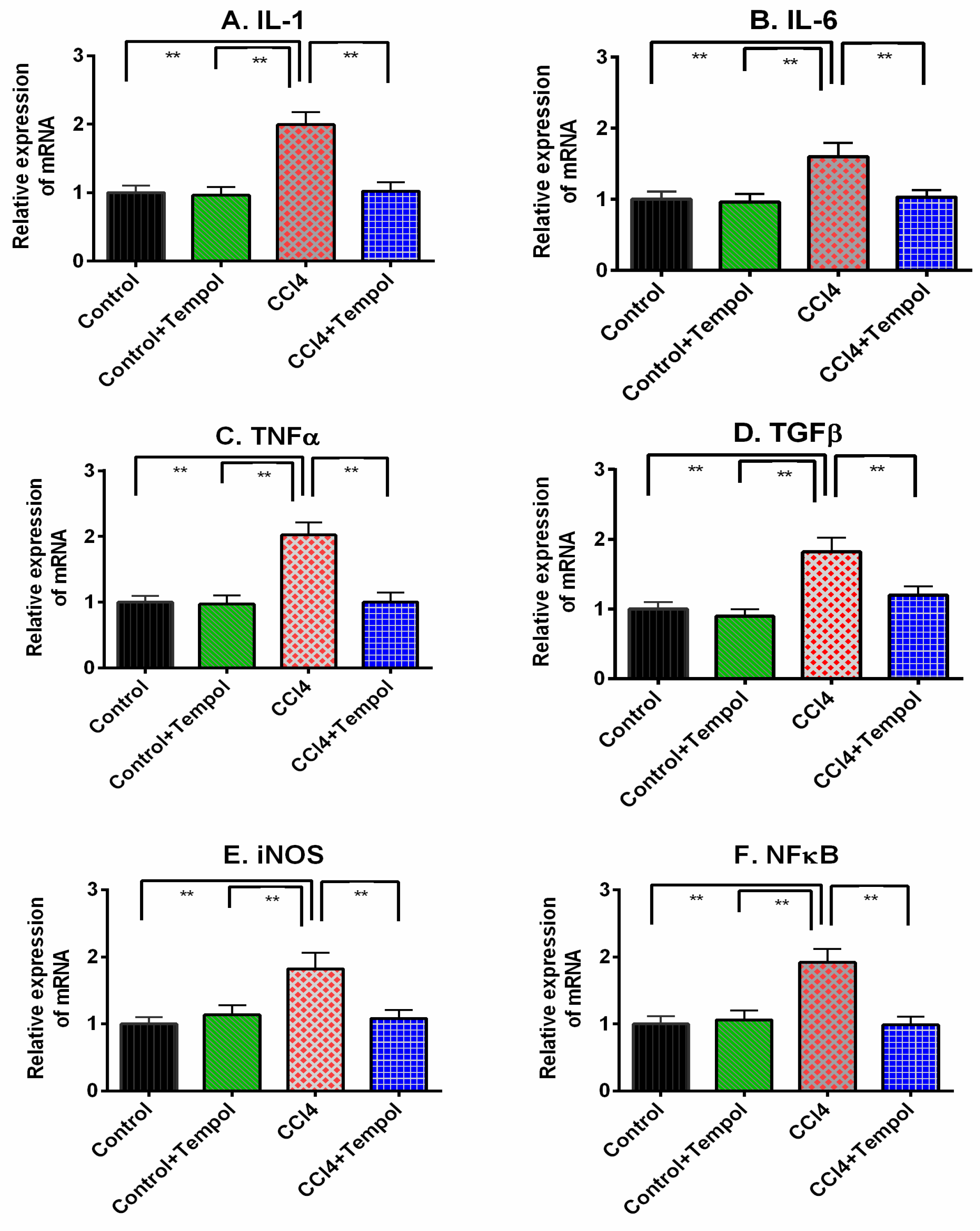
3.7. Tempol Treatment Decreased the Inflammation Related Genes Expression in Liver of CCl4-Administered Rats
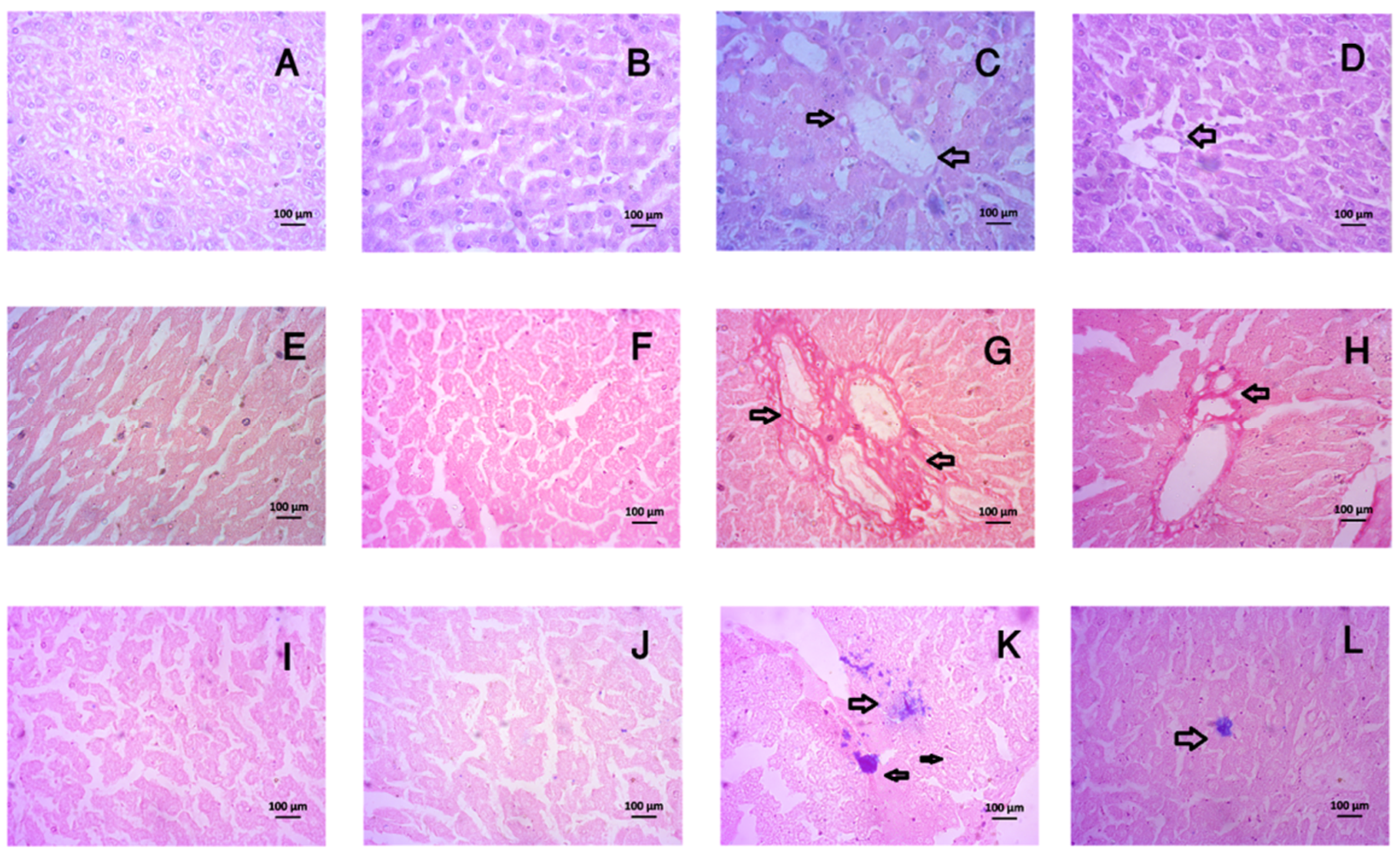
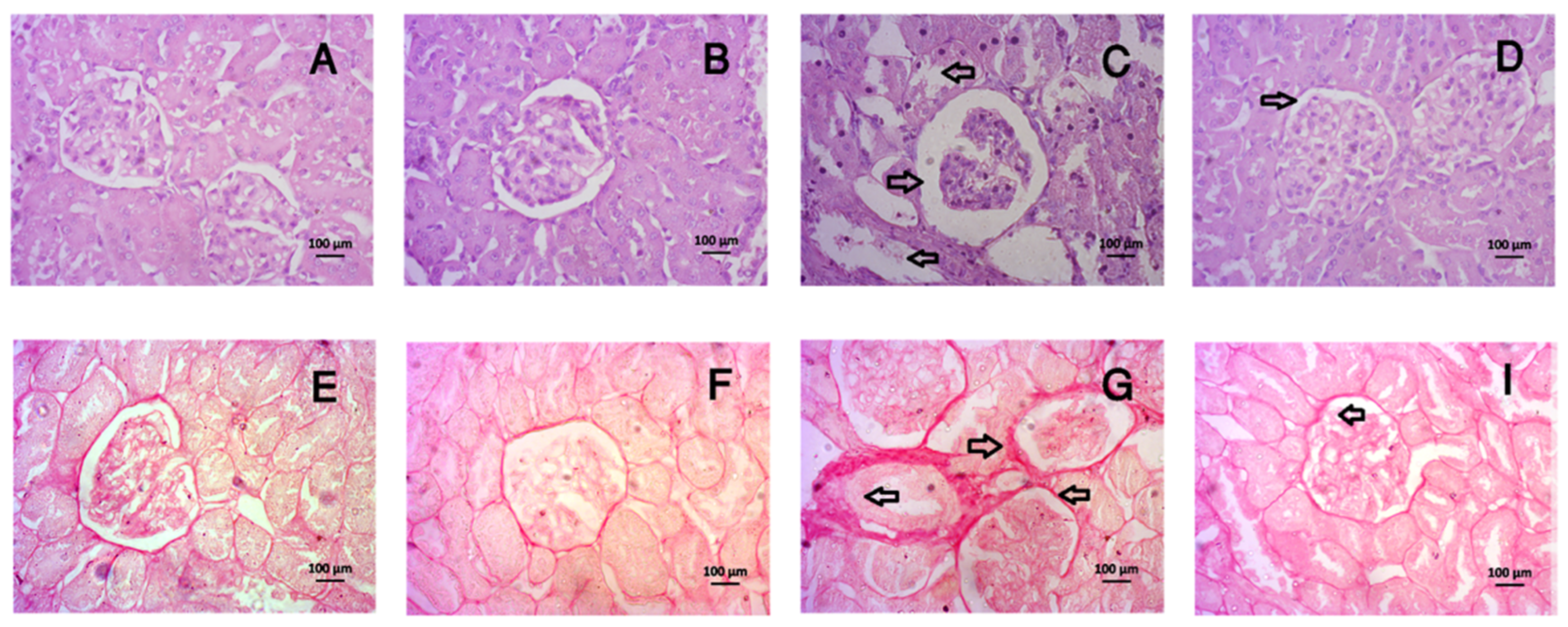
3.8. Tempol Treatment Improved the Histological Abnormalities in Liver and Kidneys of CCl4-Administered Rats
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Paik, J.M.; Golabi, P.; Younossi, Y.; Mishra, A.; Younossi, Z.M. Changes in the Global Burden of Chronic Liver Diseases From 2012 to 2017: The Growing Impact of NAFLD. Hepatology 2020, 72, 1605–1616. [Google Scholar] [CrossRef] [PubMed]
- Sarin, S.K.; Kumar, M.; Eslam, M.; George, J.; Al Mahtab, M.; Akbar, S.M.F.; Jia, J.; Tian, Q.; Aggarwal, R.; Muljono, D.H.; et al. Liver diseases in the Asia-Pacific region: A Lancet Gastroenterology & Hepatology Commission. Gastroenterol. Hepatol. 2020, 5, 167–228. [Google Scholar] [CrossRef]
- Asrani, S.K.; Devarbhavi, H.; Eaton, J.; Kamath, P.S. Burden of liver diseases in the world. J. Hepatol. 2019, 70, 151–171. [Google Scholar] [CrossRef] [PubMed]
- Cichoż-Lach, H.; Michalak, A. Oxidative stress as a crucial factor in liver diseases. World J. Gastroenterol. 2014, 20, 8082–8091. [Google Scholar] [CrossRef] [PubMed]
- Saha, P.; Talukdar, A.D.; Nath, R.; Sarker, S.D.; Nahar, L.; Sahu, J.; Choudhury, M.D. Role of Natural Phenolics in Hepatoprotection: A Mechanistic Review and Analysis of Regulatory Network of Associated Genes. Front. Pharmacol. 2019, 10, 509. [Google Scholar] [CrossRef] [PubMed]
- Uchida, D.; Takaki, A.; Oyama, A.; Adachi, T.; Wada, N.; Onishi, H.; Okada, H. Oxidative stress management in chronic liver diseases and hepatocellular carcinoma. Nutrients 2020, 12, 1576. [Google Scholar] [CrossRef] [PubMed]
- Ha, B.J.; Lee, S.H.; Kim, H.J.; Lee, J.Y. The role of Salicornia herbacea in ovariectomy-induced oxidative stress. Biol. Pharm. Bull. 2006, 29, 1305–1309. [Google Scholar] [CrossRef]
- Muriel, P. Role of free radicals in liver diseases. Hepatol. Int. 2009, 3, 526–536. [Google Scholar] [CrossRef]
- Acharya, P.; Chouhan, K.; Weiskirchen, S.; Weiskirchen, R. Cellular mechanisms of liver fibrosis. Front. Pharmacol. 2021, 12, 671640. [Google Scholar] [CrossRef]
- Ramos-Tovar, E.; Muriel, P. Molecular mechanisms that link oxidative stress, inflammation, and fibrosis in the liver. Antioxidants 2020, 9, 1279. [Google Scholar] [CrossRef]
- Islam, M.A.; Al Mamun, M.A.; Faruk, M.; Ul Islam, M.T.; Rahman, M.M.; Alam, M.N.; Rahman, A.; Reza, H.M.; Alam, M.A. Astaxanthin ameliorates hepatic damage and oxidative stress in carbon tetrachloride-administered rats. Pharmacogn. Res. 2017, 9, S84–S91. [Google Scholar] [CrossRef]
- Adebayo, A.; Yakubu, O.F.; Balogun, T.M. Protective properties of Citrullus lanatus on carbon tetrachloride induced liver damage in rats. Eur. J. Med. Plants 2014, 4, 979–989. [Google Scholar] [CrossRef]
- Abdel-Kader, M.S.; Abulhamd, A.T.; Hamad, A.M.; Alanazi, A.H.; Ali, R.; Alqasoumi, S.I. Evaluation of the hepatoprotective effect of combination between hinokiflavone and Glycyrrhizin against CCl4 induced toxicity in rats. Saudi Pharm. J. 2018, 26, 496–503. [Google Scholar] [CrossRef] [PubMed]
- Boll, M.; Weber, L.W.; Becker, E.; Stampfl, A. Mechanism of carbon tetrachloride-induced hepatotoxicity. Hepatocellular damage by reactive carbon tetrachloride metabolites. Z. Naturforsch. C J. Biosci. 2001, 56, 649–659. [Google Scholar] [CrossRef]
- Lobo, V.; Patil, A.; Phatak, A.; Chandra, N. Free radicals, antioxidants and functional foods: Impact on human health. Pharmacogn. Rev. 2010, 4, 118–126. [Google Scholar] [CrossRef]
- Kurutas, E.B. The importance of antioxidants which play the role in cellular response against oxidative/nitrosative stress: Current state. Nutr. J. 2016, 15, 71. [Google Scholar] [CrossRef] [PubMed]
- Galicia-Moreno, M.; Lucano-Landeros, S.; Monroy-Ramirez, H.C.; Silva-Gomez, J.; Gutierrez-Cuevas, J.; Santos, A.; Armendariz-Borunda, J. Roles of Nrf2 in Liver Diseases: Molecular, Pharmacological, and Epigenetic Aspects. Antioxidants 2020, 9, 980. [Google Scholar] [CrossRef]
- Tang, W.; Jiang, Y.F.; Ponnusamy, M.; Diallo, M. Role of Nrf2 in chronic liver disease. World J. Gastroenterol. 2014, 20, 13079–13087. [Google Scholar] [CrossRef]
- Ahmed, S.M.; Luo, L.; Namani, A.; Wang, X.J.; Tang, X. Nrf2 signaling pathway: Pivotal roles in inflammation. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 585–597. [Google Scholar] [CrossRef]
- Klaassen, C.D.; Reisman, S.A. Nrf2 the rescue: Effects of the antioxidative/electrophilic response on the liver. Toxicol. Appl. Pharmacol. 2010, 244, 57–65. [Google Scholar] [CrossRef]
- Khan, S.; Rahman, M.M.; Kabir, F.; Nahar, K.; Mamun, F.; Lasker, S.; Subhan, N.; Hossain, M.H.; Nahar, L.; Sarker, S.D.; et al. Trichosanthes dioica Roxb. prevents hepatic inflammation and fibrosis in CCl4-induced ovariectomized rats. Clin. Nutr. Exp. 2020, 33, 1–17. [Google Scholar] [CrossRef]
- Wilcox, C.S. Effects of tempol and redox-cycling nitroxides in models of oxidative stress. Pharmacol. Ther. 2010, 126, 119–145. [Google Scholar] [CrossRef]
- Pires, P.W.; Deutsch, C.; McClain, J.L.; Rogers, C.T.; Dorrance, A.M. Tempol, a superoxide dismutase mimetic, prevents cerebral vessel remodeling in hypertensive rats. Microvasc. Res. 2010, 80, 445–452. [Google Scholar] [CrossRef] [PubMed]
- Ebenezer, P.J.; Mariappan, N.; Elks, C.M.; Haque, M.; Francis, J. Diet-induced renal changes in Zucker rats are ameliorated by the superoxide dismutase mimetic TEMPOL. Obesity 2009, 17, 1994–2002. [Google Scholar] [CrossRef] [PubMed]
- Rafikova, O.; Salah, E.M.; Tofovic, S.P. Renal and metabolic effects of tempol in obese ZSF1 rats-distinct role for superoxide and hydrogen peroxide in diabetic renal injury. Metab. Clin. Exp. 2008, 57, 1434–1444. [Google Scholar] [CrossRef] [PubMed]
- Fiordaliso, F.; De Angelis, N.; Bai, A.; Cuccovillo, I.; Salio, M.; Serra, D.M.; Bianchi, R.; Razzetti, R.; Latini, R.; Masson, S. Effect of beta-adrenergic and renin-angiotensin system blockade on myocyte apoptosis and oxidative stress in diabetic hypertensive rats. Life Sci. 2007, 81, 951–959. [Google Scholar] [CrossRef]
- Matsuhashi, T.; Karbowski, M.; Liu, X.; Usukura, J.; Wozniak, M.; Wakabayashi, T. Complete suppression of ethanol-induced formation of megamitochondria by 4-hydroxy-2,2,6,6-tetramethyl-piperidine-1-oxyl (4-OH-TEMPO). Free Radic. Biol. Med. 1998, 24, 139–147. [Google Scholar] [CrossRef]
- Blonder, J.M.; McCalden, T.A.; Hsia, C.J.; Billings, R.E. Polynitroxyl albumin plus tempol attenuates liver injury and inflammation after hepatic ischemia and reperfusion. Life Sci. 2000, 67, 3231–3239. [Google Scholar] [CrossRef]
- Wei, Y.; Clark, S.E.; Thyfault, J.P.; Uptergrove, G.M.; Li, W.; Whaley-Connell, A.T.; Ferrario, C.M.; Sowers, J.R.; Ibdah, J.A. Oxidative stress-mediated mitochondrial dysfunction contributes to angiotensin II-induced nonalcoholic fatty liver disease in transgenic Ren2 rats. Am. J. Clin. Pathol. 2009, 174, 1329–1337. [Google Scholar] [CrossRef]
- Mamun, F.; Rahman, M.M.; Zamila, M.; Subhan, N.; Hossain, H.; Raquibul Hasan, S.M.; Alam, M.A.; Haque, M.A. Polyphenolic compounds of litchi leaf augment kidney and heart functions in 2K1C rats. J. Funct. Foods. 2020, 64, 103662. [Google Scholar] [CrossRef]
- Tracey, W.R.; Tse, J.; Carter, G. Lipopolysaccharide-induced changes in plasma nitrite and nitrate concentrations in rats and mice: Pharmacological evaluation of nitric oxide synthase inhibitors. J. Pharmacol. Exp. Ther. 1995, 272, 1011–1015. [Google Scholar] [PubMed]
- Witko-Sarsat, V.; Friedlander, M.; Capeillere-Blandin, C.; Nguyen-Khoa, T.; Nguyen, A.T.; Zingraff, J.; Jungers, P.; Descamps-Latscha, B. Advanced oxidation protein products as a novel marker of oxidative stress in uremia. Kidney Int. 1996, 49, 1304–1313. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.M.; Ferdous, K.U.; Roy, S.; Nitul, I.A.; Mamun, F.; Hossain, M.H.; Subhan, N.; Alam, M.A.; Haque, M.A. Polyphenolic compounds of amla prevent oxidative stress and fibrosis in the kidney and heart of 2K1C rats. Food Sci. Nutr. 2020, 8, 3578–3589. [Google Scholar] [CrossRef] [PubMed]
- Bradley, P.P.; Priebat, D.A.; Christensen, R.D.; Rothstein, G. Measurement of cutaneous inflammation: Estimation of neutrophil content with an enzyme marker. J. Investig. Dermatol. 1982, 78, 206–209. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; You, Y.; Yoon, H.G.; Lee, Y.H.; Kim, K.; Lee, J.; Kim, M.S.; Kim, J.C.; Jun, W. Hepatoprotective effects of fermented Curcuma longa L. on carbon tetrachloride-induced oxidative stress in rats. Food Chem. 2014, 151, 148–153. [Google Scholar] [CrossRef] [PubMed]
- Abouzied, M.M.; Eltahir, H.M.; Taye, A.; Abdelrahman, M.S. Experimental evidence for the therapeutic potential of tempol in the treatment of acute liver injury. Mol. Cell. Biochem. 2016, 411, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Boonruamkaew, P.; Chonpathompikunlert, P.; Nagasaki, Y. Redox nanoparticle therapeutics for acetaminophen-induced hepatotoxicity in mice. Oxidative Med. Cell. Longev. 2016, 2016, 4984597. [Google Scholar] [CrossRef]
- Conde de la Rosa, L.; Goicoechea, L.; Torres, S.; Garcia-Ruiz, C.; Fernandez-Checa, J.C. Role of oxidative stress in liver disorders. Livers 2022, 2, 283–314. [Google Scholar] [CrossRef]
- Cuzzocrea, S.; McDonald, M.C.; Mazzon, E.; Dugo, L.; Lepore, V.; Fonti, M.T.; Ciccolo, A.; Terranova, M.L.; Caputi, A.P.; Thiemermann, C. Tempol, a membrane-permeable radical scavenger, reduces dinitrobenzene sulfonic acid-induced colitis. Eur. J. Pharmacol. 2000, 406, 127–137. [Google Scholar] [CrossRef]
- Jing, L.; Li, Q.; He, L.; Sun, W.; Jia, Z.; Ma, H. Protective effect of tempol against hypoxia-induced oxidative stress and apoptosis in H9c2 cells. Med. Sci. Monit. Basic Res. 2017, 23, 159–165. [Google Scholar] [CrossRef]
- Samaiya, P.K.; Narayan, G.; Kumar, A.; Krishnamurthy, S. Tempol (4 hydroxy-tempo) inhibits anoxia-induced progression of mitochondrial dysfunction and associated neurobehavioral impairment in neonatal rats. J. Neurol. Sci. 2017, 375, 58–67. [Google Scholar] [CrossRef]
- Xiong, Y.; Singh, I.N.; Hall, E.D. Tempol protection of spinal cord mitochondria from peroxynitrite-induced oxidative damage. Free Radic. Res. 2009, 43, 604–612. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zhang, L.; Ouyang, X.; Jiang, Z.; Xie, Z.; Fan, L.; Zhu, D.; Li, L. Advanced oxidation protein products play critical roles in liver diseases. Eur. J. Clin. Investig. 2019, 49, e13098. [Google Scholar] [CrossRef] [PubMed]
- Ozenirler, S.; Erkan, G.; Konca Degertekin, C.; Ercin, U.; Cengiz, M.; Bilgihan, A.; Yilmaz, G.; Akyol, G. The relationship between advanced oxidation protein products (AOPP) and biochemical and histopathological findings in patients with nonalcoholic steatohepatitis. J. Dig. Dis. 2014, 15, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Han, T.; Tian, J.; Zhu, Z.-Y.; Liu, Y.; Li, Y.; Xiao, S.-X.; Li, Y.; Feng, Y.-Y. Monitoring oxidative stress in acute-on-chronic liver failure by advanced oxidation protein products. Hepatol. Res. 2012, 42, 171–180. [Google Scholar] [CrossRef]
- Miah, P.; Mohona, S.B.S.; Rahman, M.M.; Subhan, N.; Khan, F.; Hossain, H.; Sharker, S.M.; Alam, M.A. Supplementation of cumin seed powder prevents oxidative stress, hyperlipidemia and non-alcoholic fatty liver in high fat diet fed rats. Biomed. Pharmacother. 2021, 141, 111908. [Google Scholar] [CrossRef]
- Yasmin, T.; Rahman, M.M.; Khan, F.; Kabir, F.; Nahar, K.; Lasker, S.; Islam, M.D.; Hossain, M.M.; Hasan, R.; Rana, S.; et al. Metformin treatment reverses high fat diet- induced non-alcoholic fatty liver diseases and dyslipidemia by stimulating multiple antioxidant and anti-inflammatory pathways. Biochem. Biophys. Rep. 2021, 28, 101168. [Google Scholar] [CrossRef]
- Selim, S.; Akter, N.; Nayan, S.I.; Chowdhury, F.I.; Saffoon, N.; Khan, F.; Ahmed, K.S.; Ahmed, M.I.; Hossain, M.M.; Alam, M.A. Flacourtia indica fruit extract modulated antioxidant gene expression, prevented oxidative stress and ameliorated kidney dysfunction in isoprenaline administered rats. Biochem. Biophys. Rep. 2021, 26, 101012. [Google Scholar] [CrossRef]
- Elsawy, H.; Badr, G.M.; Sedky, A.; Abdallah, B.M.; Alzahrani, A.M.; Abdel-Moneim, A.M. Rutin ameliorates carbon tetrachloride (CCl4)-induced hepatorenal toxicity and hypogonadism in male rats. PeerJ 2019, 7, e7011. [Google Scholar] [CrossRef]
- Bataille, A.M.; Manautou, J.E. Nrf2: A potential target for new therapeutics in liver disease. Clin. Pharmacol. Ther. 2012, 92, 340–348. [Google Scholar] [CrossRef]
- Ma, Q. Role of Nrf2 in oxidative stress and toxicity. Annu. Rev. Pharmacol. Toxicol. 2013, 53, 401–426. [Google Scholar] [CrossRef] [PubMed]
- Chami, B.; San Gabriel, P.T.; Kum-Jew, S.; Wang, X.; Dickerhof, N.; Dennis, J.M.; Witting, P.K. The nitroxide 4-methoxy-tempo inhibits the pathogenesis of dextran sodium sulfate-stimulated experimental colitis. Redox Biol. 2020, 28, 101333. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name of Gene | Type | Sequence |
|---|---|---|
| Nrf-2 | Forward | 5′-CCC AGCACA TCC AGACAGAC-3′ |
| Reverse | 5′-TATCCAGGGCAAGCGACT C-3′ | |
| Heme oxigenase-1 (HO-1) | Forward | 5′-TGCTCGCATGAACACTCTG-3′ |
| Reverse | 5′-TCCTCTGTCAGCAGTGCCT-3′ | |
| Heme oxigenase-2 (HO-2) | Forward | 5′-CACCACTGCACTTTACTTCA-3′ |
| Reverse | 5′-AGTGCTGGGGAGTTTTAGTG-3′ | |
| MnSOD | Forward | 5′-GCTCTAATCACGACCCACT-3′ |
| Reverse | 5′-CATTCTCCCAGTTGATTACATTC-3′ | |
| Catalase | Forward | 5′-ATTGCCGTCCGATTCTCC-3′ |
| Reverse | 5′-CCAGTTACCATCTTCAGTGTAG-3′ | |
| Glutathione peroxidase (GPx) | Forward | 5′-GGGCAAAGAAGATTCCAGGTT-3′ |
| Reverse | 5′-GGACGGCTTCATCTTCAGTGA-3′ | |
| IL-1 | Forward | 5′-ATGCCTCGTGCTGTCTGACC-3′ |
| Reverse | 5′-CCATCTTTAGGAAGACACGGGTT-3′ | |
| IL-6 | Forward | 5′-AGCGATGATGCACTGTCAGA-3′ |
| Reverse | 5′-GGTTTGCCGAGTAGACCTCA-3′ | |
| TNF-α | Forward | 5′-ATGTGGAACTGGCAGAGGAG-3′ |
| Reverse | 5′-CCACGAGCAGGAATGAGAAGAG-3′ | |
| TGF-β | Forward | 5′-AAGAAGTCACCCGCGTGCTA-3′ |
| Reverse | 5′-TGTGTGATGTCTTTGGTTTTGTC-3′ | |
| iNOS | Forward | 5′-TGGTCCAACCTGCAGGTCTTC-3′ |
| Reverse | 5′-CAGTAATGGCCGACCTGATGTTG-3′ | |
| NF-кB | Forward | 5′-TGTGAAGAAGCGAGACCTGGAG-3′ |
| Reverse | 5′-GGCACGGTTATCAAAAATCGGATG-3′ | |
| β-Actin | Forward | 5′-GCGAGAAGATGACCCAGATC-3′ |
| Reverse | 5′-GGATAGCACAGCCTGGATAG-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jahan, I.; Islam, M.D.; Sarif, S.; Amena, I.J.; Shuvo, A.U.H.; Akter, N.; Chowdhury, F.I.; Akter, R.; Ahmed, I.; Khan, F.; et al. Tempol Alters Antioxidant Enzyme Function, Modulates Multiple Genes Expression, and Ameliorates Hepatic and Renal Impairment in Carbon Tetrachloride (CCl4)-Intoxicated Rats. Livers 2023, 3, 105-120. https://doi.org/10.3390/livers3010010
Jahan I, Islam MD, Sarif S, Amena IJ, Shuvo AUH, Akter N, Chowdhury FI, Akter R, Ahmed I, Khan F, et al. Tempol Alters Antioxidant Enzyme Function, Modulates Multiple Genes Expression, and Ameliorates Hepatic and Renal Impairment in Carbon Tetrachloride (CCl4)-Intoxicated Rats. Livers. 2023; 3(1):105-120. https://doi.org/10.3390/livers3010010
Chicago/Turabian StyleJahan, Ishrat, Md. Didarul Islam, Sumaia Sarif, Israt Jahan Amena, Asif Ul Haque Shuvo, Noushin Akter, Faizul Islam Chowdhury, Raushanara Akter, Iqbal Ahmed, Ferdous Khan, and et al. 2023. "Tempol Alters Antioxidant Enzyme Function, Modulates Multiple Genes Expression, and Ameliorates Hepatic and Renal Impairment in Carbon Tetrachloride (CCl4)-Intoxicated Rats" Livers 3, no. 1: 105-120. https://doi.org/10.3390/livers3010010
APA StyleJahan, I., Islam, M. D., Sarif, S., Amena, I. J., Shuvo, A. U. H., Akter, N., Chowdhury, F. I., Akter, R., Ahmed, I., Khan, F., Subhan, N., & Alam, M. A. (2023). Tempol Alters Antioxidant Enzyme Function, Modulates Multiple Genes Expression, and Ameliorates Hepatic and Renal Impairment in Carbon Tetrachloride (CCl4)-Intoxicated Rats. Livers, 3(1), 105-120. https://doi.org/10.3390/livers3010010