
Perfluoroalkyl Carboxylic Acids Interact with the Human Bile Acid Transporter NTCP

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cell Culture and Uptake Experiments
2.3. Statistical Analysis
3. Results
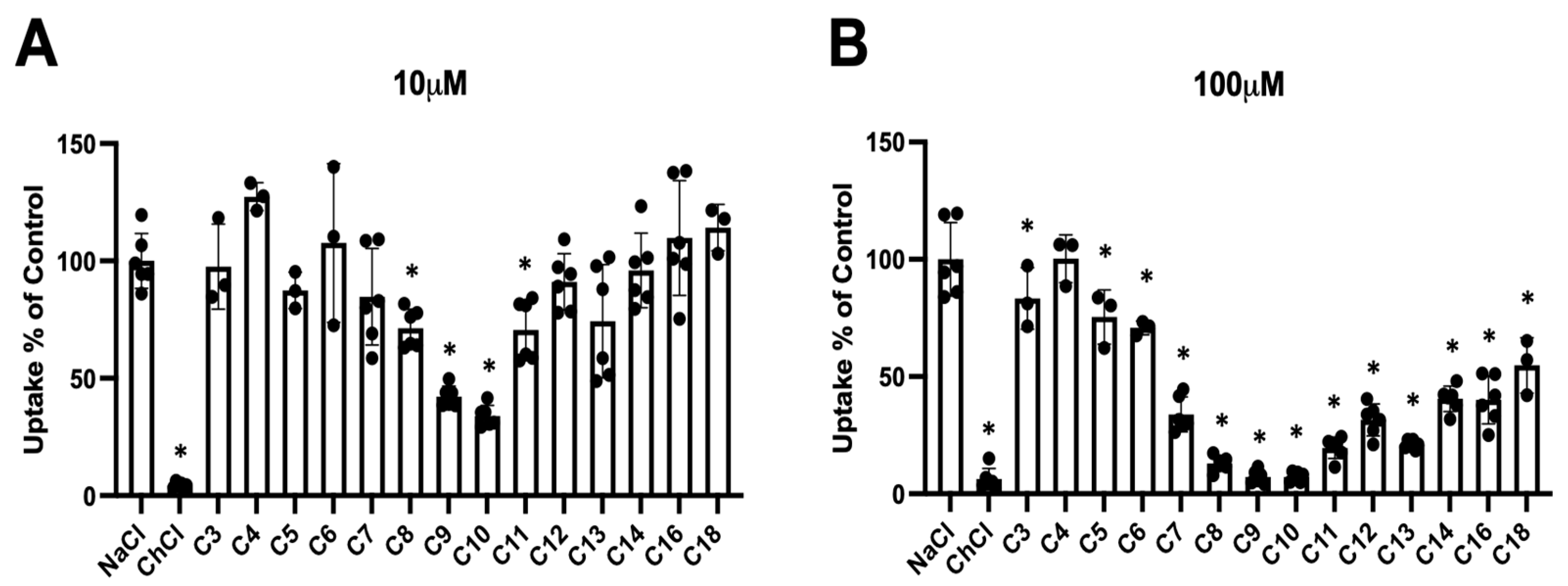
3.1. PFCAs Can Inhibit Taurocholate Uptake Mediated by the Human Na+/Taurocholate Cotransporting Polypeptide (NTCP)
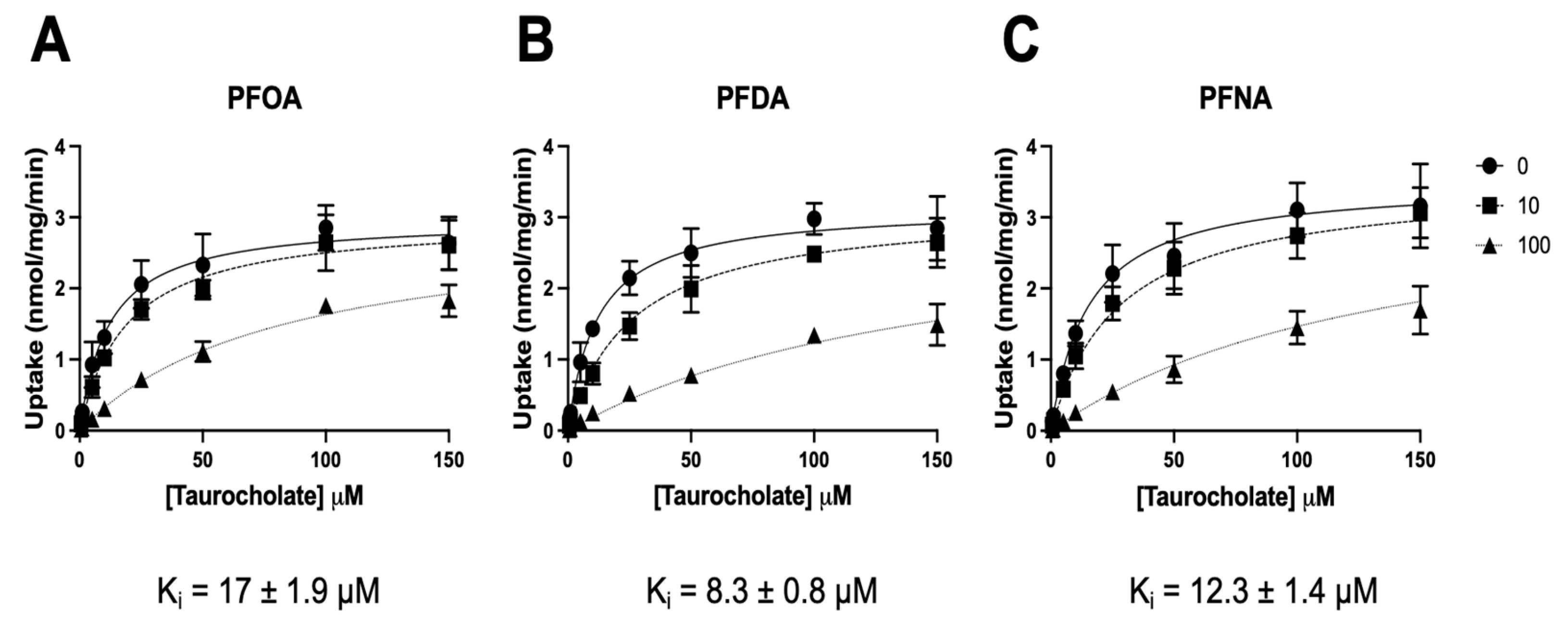
3.2. Determination of Half-Maximal Inhibitory Concentrations for PFOA, PFNA, and PFDA
3.3. Inhibition Kinetics of NTCP-Mediated Taurocholate Transport for PFOA, PFNA, and PFDA
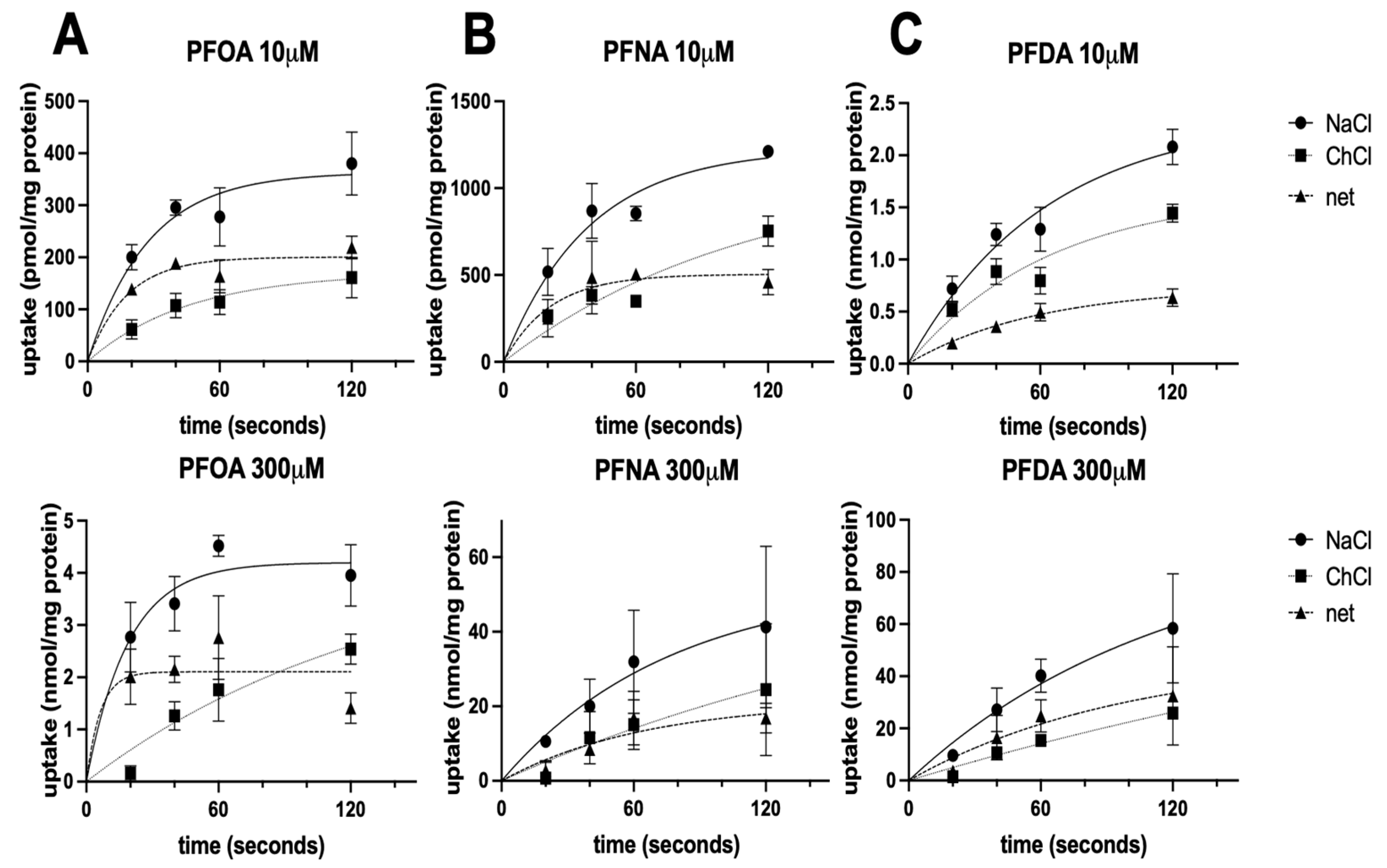
3.4. Time Dependency of NTCP-Mediated PFOA, PFNA, and PFDA Uptake
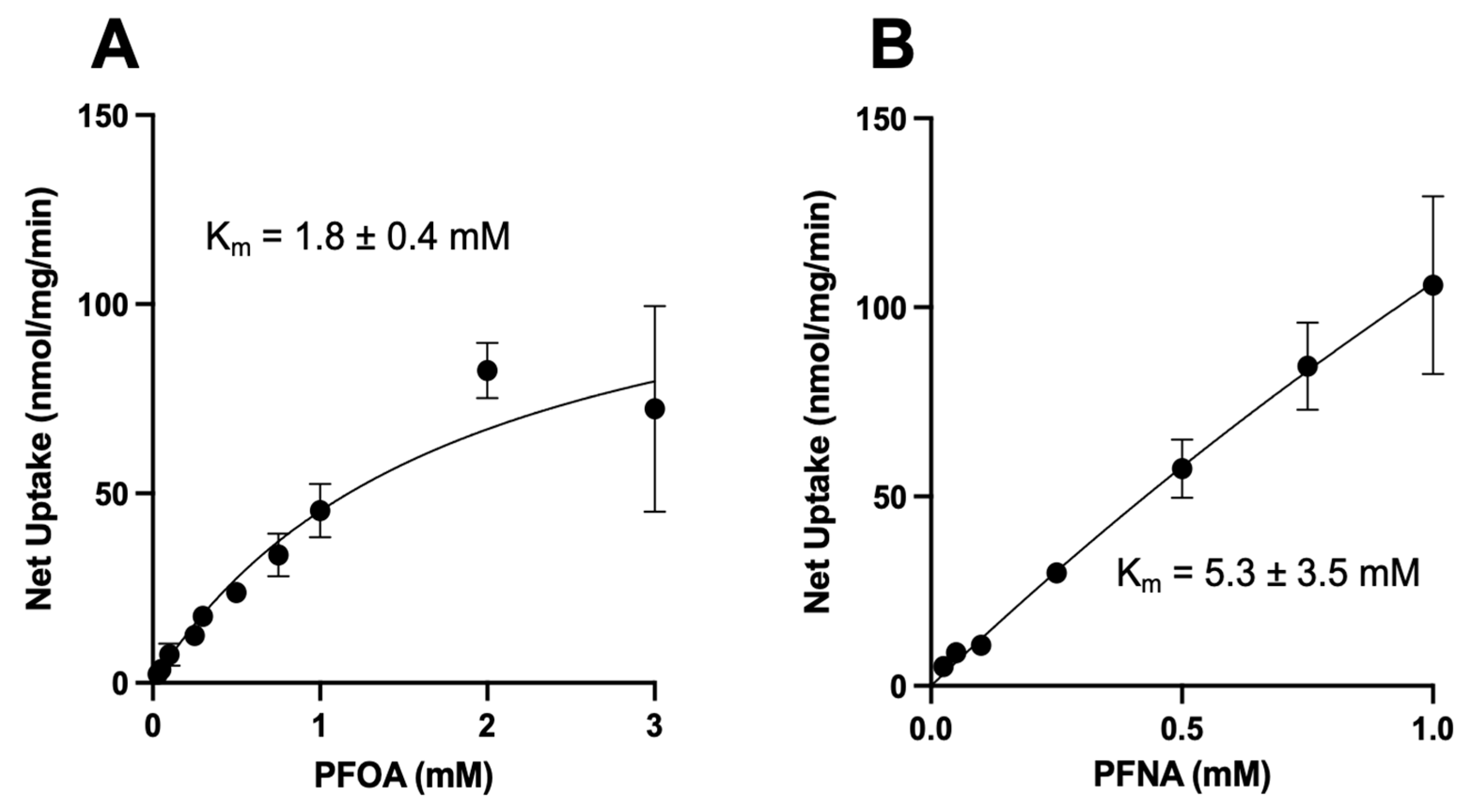
3.5. Kinetics of NTCP-Mediated PFOA and PFNA Uptake
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kissa, E. Fluorinated Surfactants and Repellents, 2nd ed.; rev. and expanded ed.; Marcel Dekker: New York, NY, USA, 2001; Volume 97. [Google Scholar]
- Prevedouros, K.; Cousins, I.T.; Buck, R.C.; Korzeniowski, S.H. Sources, fate and transport of perfluorocarboxylates. Environ. Sci. Technol. 2006, 40, 32–44. [Google Scholar] [CrossRef]
- Fromme, H.; Tittlemier, S.A.; Volkel, W.; Wilhelm, M.; Twardella, D. Perfluorinated compounds—Exposure assessment for the general population in Western countries. Int. J. Hyg. Environ. Health 2009, 212, 239–270. [Google Scholar] [CrossRef] [PubMed]
- Lau, C.; Anitole, K.; Hodes, C.; Lai, D.; Pfahles-Hutchens, A.; Seed, J. Perfluoroalkyl acids: A review of monitoring and toxicological findings. Toxicol. Sci. 2007, 99, 366–394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Death, C.; Bell, C.; Champness, D.; Milne, C.; Reichman, S.; Hagen, T. Per- and polyfluoroalkyl substances (PFAS) in livestock and game species: A review. Sci. Total Environ. 2021, 774, 144795. [Google Scholar] [CrossRef] [PubMed]
- Sunderland, E.M.; Hu, X.C.; Dassuncao, C.; Tokranov, A.K.; Wagner, C.C.; Allen, J.G. A review of the pathways of human exposure to poly- and perfluoroalkyl substances (PFASs) and present understanding of health effects. J. Expo. Sci. Environ. Epidemiol. 2019, 29, 131–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Silva, A.O.; Armitage, J.M.; Bruton, T.A.; Dassuncao, C.; Heiger-Bernays, W.; Hu, X.C.; Karrman, A.; Kelly, B.; Ng, C.; Robuck, A.; et al. PFAS Exposure Pathways for Humans and Wildlife: A Synthesis of Current Knowledge and Key Gaps in Understanding. Environ. Toxicol. Chem. 2021, 40, 631–657. [Google Scholar] [CrossRef]
- CDC. National Report on Human Exposure to Environmental Chemicals. Available online: https://www.cdc.gov/exposurereport/index.html (accessed on 8 August 2021).
- Knutsen, H.K.; Alexander, J.; Barregard, L.; Bignami, M.; Bruschweiler, B.; Ceccatelli, S.; Cottrill, B.; Dinovi, M.; Edler, L.; Grasl-Kraupp, B.; et al. Risk to human health related to the presence of perfluorooctane sulfonic acid and perfluorooctanoic acid in food. EFSA J. 2018, 16, e05194. [Google Scholar] [CrossRef]
- Chang, S.C.; Das, K.; Ehresman, D.J.; Ellefson, M.E.; Gorman, G.S.; Hart, J.A.; Noker, P.E.; Tan, Y.M.; Lieder, P.H.; Lau, C.; et al. Comparative pharmacokinetics of perfluorobutyrate in rats, mice, monkeys, and humans and relevance to human exposure via drinking water. Toxicol. Sci. 2008, 104, 40–53. [Google Scholar] [CrossRef] [Green Version]
- Johnson, J.D.; Gibson, S.J.; Ober, R.E. Cholestyramine-enhanced fecal elimination of carbon-14 in rats after administration of ammonium [14C]perfluorooctanoate or potassium [14C]perfluorooctanesulfonate. Fundam. Appl. Toxicol. 1984, 4, 972–976. [Google Scholar] [CrossRef]
- Andersen, M.E.; Butenhoff, J.L.; Chang, S.C.; Farrar, D.G.; Kennedy, G.L., Jr.; Lau, C.; Olsen, G.W.; Seed, J.; Wallace, K.B. Perfluoroalkyl acids and related chemistries—Toxicokinetics and modes of action. Toxicol. Sci. 2008, 102, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Genuis, S.J.; Birkholz, D.; Ralitsch, M.; Thibault, N. Human detoxification of perfluorinated compounds. Public Health 2010, 124, 367–375. [Google Scholar] [CrossRef]
- Genuis, S.J.; Curtis, L.; Birkholz, D. Gastrointestinal Elimination of Perfluorinated Compounds Using Cholestyramine and Chlorella pyrenoidosa. ISRN Toxicol. 2013, 2013, 657849. [Google Scholar] [CrossRef] [Green Version]
- Kennedy, G.L., Jr.; Butenhoff, J.L.; Olsen, G.W.; O’Connor, J.C.; Seacat, A.M.; Perkins, R.G.; Biegel, L.B.; Murphy, S.R.; Farrar, D.G. The toxicology of perfluorooctanoate. Crit. Rev. Toxicol. 2004, 34, 351–384. [Google Scholar] [CrossRef]
- Claro da Silva, T.; Polli, J.E.; Swaan, P.W. The solute carrier family 10 (SLC10): Beyond bile acid transport. Mol. Asp. Med. 2013, 34, 252–269. [Google Scholar] [CrossRef] [Green Version]
- Hagenbuch, B.; Stieger, B. The SLCO (former SLC21) superfamily of transporters. Mol. Asp. Med. 2013, 34, 396–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, W.; Zitzow, J.D.; Weaver, Y.; Ehresman, D.J.; Chang, S.C.; Butenhoff, J.L.; Hagenbuch, B. Organic Anion Transporting Polypeptides Contribute to the Disposition of Perfluoroalkyl Acids in Humans and Rats. Toxicol. Sci. 2017, 156, 84–95. [Google Scholar] [CrossRef] [Green Version]
- Zhao, W.; Zitzow, J.D.; Ehresman, D.J.; Chang, S.C.; Butenhoff, J.L.; Forster, J.; Hagenbuch, B. Na+/Taurocholate Cotransporting Polypeptide and Apical Sodium-Dependent Bile Acid Transporter Are Involved in the Disposition of Perfluoroalkyl Sulfonates in Humans and Rats. Toxicol. Sci. 2015, 146, 363–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weaver, Y.M.; Ehresman, D.J.; Butenhoff, J.L.; Hagenbuch, B. Roles of rat renal organic anion transporters in transporting perfluorinated carboxylates with different chain lengths. Toxicol. Sci. 2010, 113, 305–314. [Google Scholar] [CrossRef] [Green Version]
- Grosser, G.; Muller, S.F.; Kirstgen, M.; Doring, B.; Geyer, J. Substrate Specificities and Inhibition Pattern of the Solute Carrier Family 10 Members NTCP, ASBT and SOAT. Front. Mol. Biosci. 2021, 8, 689757. [Google Scholar] [CrossRef] [PubMed]
- Meier, P.J.; Eckhardt, U.; Schroeder, A.; Hagenbuch, B.; Stieger, B. Substrate specificity of sinusoidal bile acid and organic anion uptake systems in rat and human liver. Hepatology 1997, 26, 1667–1677. [Google Scholar] [CrossRef]
- Friesema, E.C.; Docter, R.; Moerings, E.P.; Stieger, B.; Hagenbuch, B.; Meier, P.J.; Krenning, E.P.; Hennemann, G.; Visser, T.J. Identification of thyroid hormone transporters. Biochem. Biophys. Res. Commun. 1999, 254, 497–501. [Google Scholar] [CrossRef] [PubMed]
- Olsen, G.W.; Burris, J.M.; Burlew, M.M.; Mandel, J.H. Epidemiologic assessment of worker serum perfluorooctanesulfonate (PFOS) and perfluorooctanoate (PFOA) concentrations and medical surveillance examinations. J. Occup. Environ. Med. 2003, 45, 260–270. [Google Scholar] [CrossRef]
- Emmett, E.A.; Shofer, F.S.; Zhang, H.; Freeman, D.; Desai, C.; Shaw, L.M. Community exposure to perfluorooctanoate: Relationships between serum concentrations and exposure sources. J. Occup. Environ. Med. 2006, 48, 759–770. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Fletcher, T.; Mucs, D.; Scott, K.; Lindh, C.H.; Tallving, P.; Jakobsson, K. Half-lives of PFOS, PFHxS and PFOA after end of exposure to contaminated drinking water. Occup. Environ. Med. 2018, 75, 46–51. [Google Scholar] [CrossRef] [Green Version]
- Fenton, S.E.; Ducatman, A.; Boobis, A.; DeWitt, J.C.; Lau, C.; Ng, C.; Smith, J.S.; Roberts, S.M. Per- and Polyfluoroalkyl Substance Toxicity and Human Health Review: Current State of Knowledge and Strategies for Informing Future Research. Environ. Toxicol. Chem. 2021, 40, 606–630. [Google Scholar] [CrossRef]
- Zhang, Y.; Beesoon, S.; Zhu, L.; Martin, J.W. Biomonitoring of perfluoroalkyl acids in human urine and estimates of biological half-life. Environ. Sci. Technol. 2013, 47, 10619–10627. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Concentration of PFCA | Km (μM) | Vmax (nmol/mg·min) | Ki (μM) | |
|---|---|---|---|---|
| PFOA | 0 | 12 (8.7–16) | 2.9 (2.7–3.2) | 17 (14–21) |
| 10 μM | 20 (16–25) | 3.0 (2.8–3.2) | ||
| 100 μM | 73 (60–92) | 2.8 (2.6–3.1) | ||
| PFDA | 0 | 12 (9.4–15) | 3.2 (2.9–3.4) | 8.3 (6.9–10) |
| 10 μM | 29 (23–36) | 3.2 (3.0–3.4) | ||
| 100 μM | 99 (73–139) | 2.5 (2.2–3.0) | ||
| PFNA | 0 | 16 (12–22) | 3.5 (3.2–3.8) | 12 (10–15) |
| 10 μM | 24 (19–30) | 3.5 (3.2–3.7) | ||
| 100 μM | 118 (78–192) | 3.1 (2.5–4.1) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruggiero, M.J.; Miller, H.; Idowu, J.Y.; Zitzow, J.D.; Chang, S.-C.; Hagenbuch, B. Perfluoroalkyl Carboxylic Acids Interact with the Human Bile Acid Transporter NTCP. Livers 2021, 1, 221-229. https://doi.org/10.3390/livers1040017
Ruggiero MJ, Miller H, Idowu JY, Zitzow JD, Chang S-C, Hagenbuch B. Perfluoroalkyl Carboxylic Acids Interact with the Human Bile Acid Transporter NTCP. Livers. 2021; 1(4):221-229. https://doi.org/10.3390/livers1040017
Chicago/Turabian StyleRuggiero, Melissa J., Haley Miller, Jessica Y. Idowu, Jeremiah D. Zitzow, Shu-Ching Chang, and Bruno Hagenbuch. 2021. "Perfluoroalkyl Carboxylic Acids Interact with the Human Bile Acid Transporter NTCP" Livers 1, no. 4: 221-229. https://doi.org/10.3390/livers1040017
APA StyleRuggiero, M. J., Miller, H., Idowu, J. Y., Zitzow, J. D., Chang, S.-C., & Hagenbuch, B. (2021). Perfluoroalkyl Carboxylic Acids Interact with the Human Bile Acid Transporter NTCP. Livers, 1(4), 221-229. https://doi.org/10.3390/livers1040017