
Anticoagulant Potential of Modified Sulfated Exopolysaccharides from Deep-Sea Bacteria: Toward Non-Animal Heparin Alternatives

 , , , , ,
, , , , ,  and
and 
Abstract
1. Introduction

2. Materials and Methods
2.1. Preparation and Characterization of Infernan and Diabolican Derivatives
2.2. Clotting and Chromogenic Assays
2.3. Thrombin Generation Test (TGT)
2.4. Molecular Docking
3. Results
3.1. Characteristics of Diabolican and Infernan Derivatives
3.2. Clotting Assays with Diabolican and Infernan Derivatives
3.3. Thrombin Generation Assay of Infernan and Diabolican Derivatives
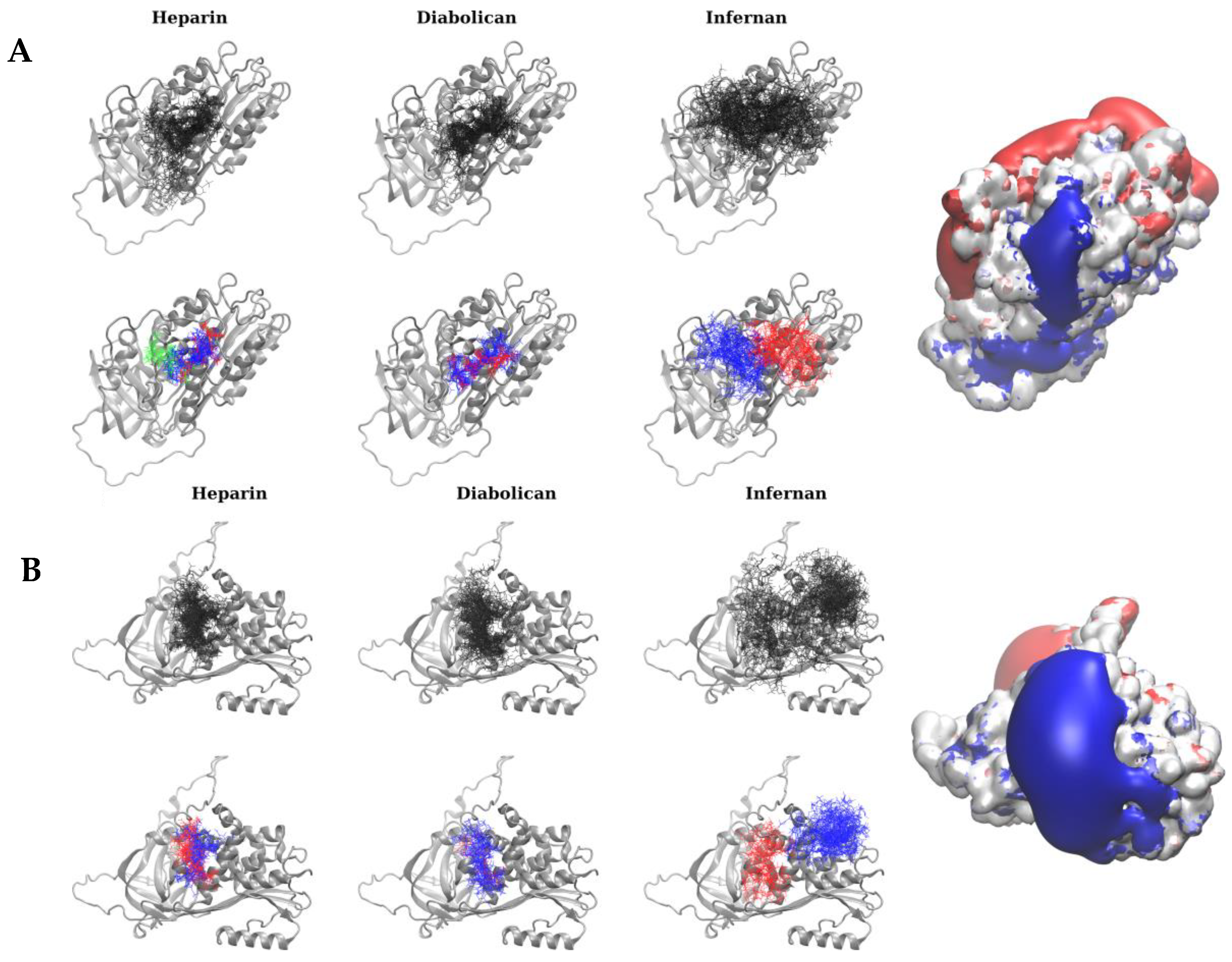
3.4. Molecular Docking Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| APTT | activated partial thromboplastin time |
| AT | antithrombin |
| CTI | corn trypsin inhibitor |
| Da | Daltons |
| DMF | dimethylformamide |
| EPS | exopolysaccharides |
| ETP | endogenous thrombin potential |
| F | factor |
| GAG | glycosaminoglycan |
| GlcA | β-D-glucuronic |
| GlcN | α-D-glucosamine |
| HPAEC | high-performance anin-exchange chromatography |
| HCII | heparin cofactor II |
| HMW | high molecular weight |
| HPSEC-M | high pressure size exclusion chromatography with multi angle light scattering |
| ACS | |
| IdoA | α-L-iduronic |
| LMW | low molecular weight |
| MW | molecular weight |
| NaCl | saline solution |
| PPP | poor platelet plasma |
| RMSatd | room mean square atom type deviation |
| S | sulfated |
| TF | tissue factor |
| TG | thrombin generation |
| TGT | thrombin generation test |
| TT | thrombin time |
| UF | unfractionated |
References
- Hao, C.; Xu, H.; Yu, L.; Zhang, L. Heparin: An essential drug for modern medicine. Prog. Mol. Biol. Transl. Sci. 2019, 163, 1–19. [Google Scholar] [PubMed]
- Ersdal-Badju, E.; Lu, A.; Zuo, Y.; Picard, V.; Bock, S.C. Identification of the antithrombin III heparin binding site. J. Biol. Chem. 1997, 272, 19393–19400. [Google Scholar] [CrossRef] [PubMed]
- Blackburn, M.N.; Smith, R.L.; Carson, J.; Sibley, C.C. The heparin-binding site of antithrombin III. Identification of a critical tryptophan in the amino acid sequence. J. Biol. Chem. 1984, 259, 939–941. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, R.D.; Lam, L. Correlation between structure and function of heparin. Proc. Natl. Acad. Sci. USA 1979, 76, 1218–1222. [Google Scholar] [CrossRef]
- Petitou, M.; Duchaussoy, P.; Lederman, I.; Choay, J.; Sinaÿ, P.; Jacquinet, J.C.; Torri, G. Synthesis of heparin fragments. A chemical synthesis of the pentasaccharide O-(2-deoxy-2-sulfamido-6-O-sulfo-alpha-D-glucopyranosyl)-(1-4)-O-(beta-D-glucopyranosyluronic acid)-(1-4)-O-(2-deoxy-2-sulfamido-3,6-di-O-sulfo-alpha-D-glu copyranosyl)-(1-4)-O-(2-O-sulfo-alpha-L-idopyranosyluronic acid)-(1-4)-2-deoxy-2-sulfamido-6-O-sulfo-D-glucopyranose decasodium salt, a heparin fragment having high affinity for antithrombin III. Carbohydr. Res. 1986, 147, 221–236. [Google Scholar]
- Atha, D.H.; Lormeau, J.C.; Petitou, M.; Rosenberg, R.D.; Choay, J. Contribution of 3-O- and 6-O-sulfated glucosamine residues in the heparin-induced conformational change in antithrombin III. Biochemistry 1987, 26, 6454–6461. [Google Scholar] [CrossRef]
- Capila, I.; Linhardt, R.J. Heparin-protein interactions. Angew. Chem. Int. Ed. Engl. 2002, 41, 391–412. [Google Scholar] [CrossRef]
- Hsu, E.; Moosavi, L. Biochemistry, Antithrombin III. In StatPearls [Internet]; StatPearls Publishing: Singapore, 2023. [Google Scholar]
- Gray, E.; Mulloy, B.; Barrowcliffe, T.W. Heparin and low-molecular-weight heparin. Thromb. Haemost. 2008, 99, 807–818. [Google Scholar]
- Tollefsen, D.M.; Pestka, C.A.; Monafo, W.J. Activation of heparin cofactor II by dermatan sulfate. J. Biol. Chem. 1983, 258, 6713–6716. [Google Scholar] [CrossRef]
- Colliec, S.; Fischer, A.M.; Tapon-Bretaudiere, J.; Boisson, C.; Durand, P.; Jozefonvicz, J. Anticoagulant properties of a fucoïdan fraction. Thromb. Res. 1991, 64, 143–154. [Google Scholar] [CrossRef]
- Aláez-Versón, C.R.; Lantero, E.; Fernàndez-Busquets, X. Heparin: New life for an old drug. Nanomedicine 2017, 12, 1727–1744. [Google Scholar] [CrossRef] [PubMed]
- Lever, R.; Page, C.P. Novel drug development opportunities for heparin. Nat. Rev. Drug Discov. 2002, 1, 140–148. [Google Scholar] [CrossRef]
- Lindahl, U. ‘Heparin’—From anticoagulant drug into the new biology. Glycoconj. J. 2000, 17, 597–605. [Google Scholar] [CrossRef] [PubMed]
- Shi, D.; Sheng, A.; Chi, L. Glycosaminoglycan-Protein Interactions and Their Roles in Human Disease. Front. Mol. Biosci. 2021, 8, 639666. [Google Scholar] [CrossRef] [PubMed]
- Torri, G.; Naggi, A. Heparin centenary—An ever-young life-saving drug. Int. J. Cardiol. 2016, 212 (Suppl. 1), S1–S4. [Google Scholar] [CrossRef]
- Folkman, J.; Langer, R.; Linhardt, R.J.; Haudenschild, C.; Taylor, S. Angiogenesis inhibition and tumor regression caused by heparin or a heparin fragment in the presence of cortisone. Science 1983, 221, 719–725. [Google Scholar] [CrossRef]
- Guerrini, M.; Beccati, D.; Shriver, Z.; Naggi, A.; Viswanathan, K.; Bisio, A.; Capila, I.; Lansing, J.C.; Guglieri, S.; Fraser, B.; et al. Oversulfated chondroitin sulfate is a contaminant in heparin associated with adverse clinical events. Nat. Biotechnol. 2008, 26, 669–675. [Google Scholar] [CrossRef]
- Baytas, S.N.; Linhardt, R.J. Advances in the preparation and synthesis of heparin and related products. Drug Discov. Today 2020, 25, 2095–2109. [Google Scholar] [CrossRef]
- Piran, S.; Schulman, S. Treatment of bleeding complications in patients on anticoagulant therapy. Blood 2019, 133, 425–435. [Google Scholar] [CrossRef]
- Liu, J.; Linhardt, R.J. Chemoenzymatic synthesis of heparan sulfate and heparin. Nat. Prod. Rep. 2014, 31, 1676–1685. [Google Scholar] [CrossRef]
- Eftekhari, A.; Kryschi, C.; Pamies, D.; Gulec, S.; Ahmadian, E.; Janas, D.; Davaran, S.; Khalilov, R. Natural and synthetic nanovectors for cancer therapy. Nanotheranostics 2023, 7, 236–257. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Ji, H.; Xu, L.; Cheng, S.; Liu, X.; Li, Y.; Zhong, R.; Zhao, W.; Kizhakkedathu, J.N.; Zhao, C. Self-anticoagulant sponge for whole blood auto-transfusion and its mechanism of coagulation factor inactivation. Nat. Commun. 2023, 14, 4875. [Google Scholar] [CrossRef] [PubMed]
- Lakhdar, M. Calotropis procera (ait) r. Br, a valuable medicine plant: A review. Adv. Biol. Earth Sci. 2023, 8, 27–35. [Google Scholar]
- Cimini, D.; Rosa, M.D.; Schiraldi, C. Production of glucuronic acid-based polysaccharides by microbial fermentation for biomedical applications. Biotechnol. J. 2012, 7, 237–250. [Google Scholar] [CrossRef]
- DeAngelis, P.L. Glycosaminoglycan polysaccharide biosynthesis and production: Today and tomorrow. Appl. Microbiol. Biotechnol. 2012, 94, 295–305. [Google Scholar] [CrossRef]
- Teng, L.; Fu, H.; Deng, C.; Chen, J.; Chen, J. Modulating the SDF-1/CXCL12-induced cancer cell growth and adhesion by sulfated K5 polysaccharides in vitro. Biomed. Pharmacother. 2015, 73, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Colliec-Jouault, S.; Bavington, C.; Delbarre-Ladrat, C. Heparin-like entities from marine organisms. Handb. Exp. Pharmacol. 2012, 207, 423–449. [Google Scholar]
- Schmid, J.; Sieber, V.; Rehm, B. Bacterial exopolysaccharides: Biosynthesis pathways and engineering strategies. Front. Microbiol. 2015, 6, 496. [Google Scholar] [CrossRef]
- Akoumany, K.; Zykwinska, A.; Sinquin, C.; Marchand, L.; Fanuel, M.; Ropartz, D.; Rogniaux, H.; Pipelier, M.; Delbarre-Ladrat, C.; Colliec-Jouault, S. Characterization of New Oligosaccharides Obtained by An Enzymatic Cleavage of the Exopolysaccharide Produced by the Deep-Sea Bacterium Alteromonas infernus Using its Cell Extract. Molecules 2019, 24, 3441. [Google Scholar] [CrossRef]
- Roger, O.; Kervarec, N.; Ratiskol, J.; Colliec-Jouault, S.; Chevolot, L. Structural studies of the main exopolysaccharide produced by the deep-sea bacterium Alteromonas infernus. Carbohydr. Res. 2004, 339, 2371–2380. [Google Scholar] [CrossRef]
- Colliec-Jouault, S.; Esposito, F.; Ledru, H.; Sinquin, C.; Marchand, L.; Fillaudeau, A.; Routier, S.; Buron, F.; Lopin-Bon, C.; Cuenot, S.; et al. Glycosaminoglycan Mimetics Obtained by Microwave-Assisted Sulfation of Marine Bacterium Sourced Infernan Exopolysaccharide. Biomacromolecules 2023, 24, 462–470. [Google Scholar] [CrossRef] [PubMed]
- Esposito, F.; Vessella, G.; Sinquin, C.; Traboni, S.; Iadonisi, A.; Colliec-Jouault, S.; Zykwinska, A.; Bedini, E. Glycosaminoglycan-like sulfated polysaccharides from Vibrio diabolicus bacterium: Semi-synthesis and characterization. Carbohydr. Polym. 2022, 283, 119054. [Google Scholar] [CrossRef] [PubMed]
- Heymann, D.; Ruiz-Velasco, C.; Chesneau, J.; Ratiskol, J.; Sinquin, C.; Colliec-Jouault, S. Anti-Metastatic Properties of a Marine Bacterial Exopolysaccharide-Based Derivative Designed to Mimic Glycosaminoglycans. Molecules 2016, 21, 309. [Google Scholar] [CrossRef] [PubMed]
- Gélébart, P.; Cuenot, S.; Sinquin, C.; Halgand, B.; Sourice, S.; Le Visage, C.; Guicheux, J.; Colliec-Jouault, S.; Zykwinska, A. Microgels based on Infernan, a glycosaminoglycan-mimetic bacterial exopolysaccharide, as BMP-2 delivery systems. Carbohydr. Polym. 2022, 284, 119191. [Google Scholar] [CrossRef]
- Rougeaux, H.; Kervarec, N.; Pichon, R.; Guezennec, J. Structure of the exopolysaccharide of Vibrio diabolicus isolated from a deep-sea hydrothermal vent. Carbohydr. Res. 1999, 322, 40–45. [Google Scholar] [CrossRef]
- Raguénès, G.; Christen, R.; Guezennec, J.; Pignet, P.; Barbier, G. Vibrio diabolicus sp. nov., a new polysaccharide-secreting organism isolated from a deep-sea hydrothermal vent polychaete annelid, Alvinella pompejana. Int. J. Syst. Bacteriol. 1997, 47, 989–995. [Google Scholar] [CrossRef]
- Guezennec, J.; Pignet, P.; Lijour, Y.; Gentric, E.; Ratiskol, J.; Colliec-Jouault, S. Sulfation and depolymerization of a bacterial exopolysaccharide of hydrothermal origin. Carbohydr. Polym. 1998, 37, 19–24. [Google Scholar] [CrossRef]
- Chopin, N.; Sinquin, C.; Ratiskol, J.; Zykwinska, A.; Weiss, P.; Cérantola, S.; Le Bideau, J.; Colliec-Jouault, S. A Direct Sulfation Process of a Marine Polysaccharide in Ionic Liquid. BioMed Res. Int. 2015, 2015, 508656. [Google Scholar] [CrossRef] [PubMed]
- Case, D.A.; Belfon, K.; Ben-Shalom, I.Y.; Brozell, S.R.; Cerutti, D.S.; Cheatham, T.E., III; Cruzeiro, V.W.D.; Darden, T.A.; Duke, R.E.; Giambasu, G.; et al. Amber 2020; University of California: San Francisco, CA, USA, 2020. [Google Scholar]
- Kirschner, K.N.; Yongye, A.B.; Tschampel, S.M.; González-Outeiriño, J.; Daniels, C.R.; Foley, B.L.; Woods, R.J. GLYCAM06: A generalizable biomolecular force field. Carbohydr. J. Comput. Chem. 2008, 29, 622–655. [Google Scholar] [CrossRef]
- Morris, G.M.; Goodsell, D.S.; Halliday, R.S.; Huey, R.; Hart, W.E.; Belew, R.K.; Olson, A.J. Automated docking using a Lamarckian genetic algorithm and an empirical binding free energy function. J. Comput. Chem. 1998, 19, 1639–1662. [Google Scholar] [CrossRef]
- Samsonov, S.A.; Pisabarro, M.T. Computational analysis of interactions in structurally available protein–glycosaminoglycan complexes. Glycobiology 2016, 26, 850–861. [Google Scholar] [CrossRef]
- Uciechowska-Kaczmarzyk, U.; Chauvot de Beauchene, I.; Samsonov, S.A. Docking software performance in protein-glycosaminoglycan systems. J. Mol. Graph. Model. 2019, 90, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Ester, M.; Kriegel, H.P.; Sander, J.; Xu, X. A density-based algorithm for discovering clusters in large spatial databases with noise. In Proceedings of the Second International Conference on Knowledge Discovery and Data Mining, KDD’96, Portland, OR, USA, 2–4 August 1996; AAAI Press: Portland, OR, USA, 1996; pp. 226–231. [Google Scholar]
- Samsonov, S.A.; Gehrcke, J.P.; Pisabarro, M.T. Flexibility and Explicit Solvent in Molecular-Dynamics-Based Docking of Protein–Glycosaminoglycan Systems. J. Chem. Inf. Model. 2014, 54, 582–592. [Google Scholar] [CrossRef]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Case, D.A.; Aktulga, H.M.; Belfon, K.; Ben-Shalom, I.Y.; Berryman, J.T.; Brozell, S.R.; Carvahol, F.S.; Cerutti, D.S.; Cheatham, T.E., III; Cisneros, G.A.; et al. Amber 2025; University of California: San Francisco, CA, USA, 2025. [Google Scholar]
- Meneghetti, M.C.; Hughes, A.J.; Rudd, T.R.; Nader, H.B.; Powell, A.K.; Yates, E.A.; Lima, M.A. Heparan sulfate and heparin interactions with proteins. J. R. Soc. Interface 2015, 12, 0589. [Google Scholar] [CrossRef]
- Santoro, R.C.; Molinari, A.C.; Leotta, M.; Martini, T. Isolated Prolongation of Activated Partial Thromboplastin Time: Not Just Bleeding Risk! Medicina 2023, 59, 1169. [Google Scholar] [CrossRef] [PubMed]
- Restaino, O.F.; Bhaskar, U.; Paul, P.; Li, L.; De Rosa, M.; Dordick, J.S.; Linhardt, R.J. High cell density cultivation of a recombinant E. coli strain expressing a key enzyme in bioengineered heparin production. Appl. Microbiol. Biotechnol. 2013, 97, 3893–3900. [Google Scholar] [CrossRef]
- Pavão, M.S. Glycosaminoglycans analogs from marine invertebrates: Structure, biological effects, and potential as new therapeutics. Front. Cell Infect. Microbiol. 2014, 4, 123. [Google Scholar] [CrossRef]
- Nahain, A.A.; Ignjatovic, V.; Monagle, P.; Tsanaktsidis, J.; Ferro, V. Heparin mimetics with anticoagulant activity. Med. Res. Rev. 2018, 38, 1582–1613. [Google Scholar] [CrossRef]
- Heymann, D.; Muñoz-Garcia, J.; Babuty, A.; Audéon, A.; Ollivier, E.; Papy-Garcia, D.; Chantepie, S.; Zykwinska, A.; Sinquin, C.; Colliec-Jouault, S. A new promising anticancer agent: A glycosaminoglycan-mimetic derived from the marine bacterial infernan exopolysaccharide. Int. J. Biol. Macromol. 2025, 19, 142074. [Google Scholar] [CrossRef]
- Perez, S.; Makshakova, O.; Angulo, J.; Bedini, E.; Bisio, A.; de Paz, J.L.; Fadda, E.; Guerrini, M.; Hricovini, M.; Hricovini, M.; et al. Glycosaminoglycans: What Remains to Be Deciphered? JACS Au 2023, 3, 628–656. [Google Scholar] [CrossRef] [PubMed]
- Choay, J.; Petitou, M.; Lormeau, J.C.; Sinaÿ, P.; Casu, B.; Gatti, G. Structure-activity relationship in heparin: A synthetic pentasaccharide with high affinity for antithrombin III and eliciting high anti-factor Xa activity. Biochem. Biophys. Res. Commun. 1983, 116, 492–499. [Google Scholar] [CrossRef]
- Petitou, M.; Hérault, J.P.; Bernat, A.; Driguez, P.A.; Duchaussoy, P.; Lormeau, J.C.; Herbert, J.M. Synthesis of thrombin-inhibiting heparin mimetics without side effects. Nature 1999, 398, 417–422. [Google Scholar] [CrossRef]
- Colliec-Jouault, S.; Chevolot, L.; Helley, D.; Ratiskol, J.; Bros, A.; Sinquin, C.; Roger, O.; Fischer, A.M. Characterization, chemical modifications and in vitro anticoagulant properties of an exopolysaccharide produced by Alteromonas infernus. Biochim. Biophys. Acta 2001, 1528, 141–151. [Google Scholar] [CrossRef]
- Grover, S.P.; Mackman, N. Anticoagulant SERPINs: Endogenous Regulators of Hemostasis and Thrombosis. Front. Cardiovasc. Med. 2022, 9, 878199. [Google Scholar] [CrossRef]
- Bano, S.; Khan, A.B.; Fatima, S.; Rashid, Q.; Prakash, A.; Gupta, N.; Ahmad, I.; Ansari, S.; Lynn, A.M.; Abid, M.; et al. Mannose 2, 3, 4, 5, 6-O-pentasulfate (MPS): A partial activator of human heparin cofactor II with anticoagulation potential. J. Biomol. Struct. Dyn. 2023, 41, 3717–3727. [Google Scholar] [CrossRef] [PubMed]
- Al-Horani, R.A.; Abdelfadiel, E.I.; Afosah, D.K.; Morla, S.; Sistla, J.C.; Mohammed, B.; Martin, E.J.; Sakagami, M.; Brophy, D.F.; Desai, U.R. A synthetic heparin mimetic that allosterically inhibits factor XIa and reduces thrombosis in vivo without enhanced risk of bleeding. J. Thromb. Haemost. 2019, 17, 2110–2122. [Google Scholar] [CrossRef] [PubMed]
- Buyue, Y.; Misenheimer, T.M.; Sheehan, J.P. Low molecular weight heparin inhibits plasma thrombin generation via direct targeting of factor IXa: Contribution of the serpin-independent mechanism. J. Thromb. Haemost. 2012, 10, 2086–2098. [Google Scholar] [CrossRef]
- Ofosu, F.A.; Blajchman, M.A.; Modi, G.J.; Smith, L.M.; Buchanan, M.R.; Hirsh, J. The importance of thrombin inhibition for the expression of the anticoagulant activities of heparin, dermatan sulphate, low molecular weight heparin and pentosan polysulphate. Br. J. Haematol. 1985, 60, 695–704. [Google Scholar] [CrossRef]
- Barrowcliffe, T.W.; Merton, R.E.; Havercroft, S.J.; Thunberg, L.; Lindahl, U.; Thomas, D.P. Low-affinity heparin potentiates the action of high-affinity heparin oligosaccharides. Thromb. Res. 1984, 34, 125–133. [Google Scholar] [CrossRef]
- Fischer, A.M.; Merton, R.E.; Marsh, N.A.; Williams, S.; Gaffney, P.J.; Barrowcliffe, T.W.; Thomas, D.P. A comparison of pentosan polysulphate and heparin. II: Effects of subcutaneous injection. Thromb. Haemost. 1982, 47, 109–113. [Google Scholar] [CrossRef] [PubMed]
- Buchanan, M.R.; Boneu, B.; Ofosu, F.; Hirsh, J. The relative importance of thrombin inhibition and factor Xa inhibition to the antithrombotic effects of heparin. Blood 1985, 65, 198–201. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Mw g/mol | Ip Mw/Mn | Sulfate % (w/w) |
|---|---|---|---|
| Inf50-Lsulf | 50,000 | 2 | 10 |
| Inf4-Hsulf | 4000 | 2 | 40 |
| Inf20-Hsulf | 20,000 | 2 | 40 |
| Inf50-Hsulf | 50,000 | 2 | 40 |
| Dia20-0sulf | 20,000 | 2 | 0 |
| Dia6-Hsulf | 6000 | 2 | 35 |
| Dia20-Hsulf | 20,000 | 2 | 40 |
| UF heparin | 15,000 | np | np |
| LMW heparin | 5000 | np | np |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Babuty, A.; Zykwinska, A.; Samsonov, S.A.; Candia, N.; Veinstein, C.; Pugnière, M.; Ngo, T.H.G.; Sinquin, C.; Muñoz-Garcia, J.; Colliec-Jouault, S.; et al. Anticoagulant Potential of Modified Sulfated Exopolysaccharides from Deep-Sea Bacteria: Toward Non-Animal Heparin Alternatives. Polysaccharides 2025, 6, 54. https://doi.org/10.3390/polysaccharides6020054
Babuty A, Zykwinska A, Samsonov SA, Candia N, Veinstein C, Pugnière M, Ngo THG, Sinquin C, Muñoz-Garcia J, Colliec-Jouault S, et al. Anticoagulant Potential of Modified Sulfated Exopolysaccharides from Deep-Sea Bacteria: Toward Non-Animal Heparin Alternatives. Polysaccharides. 2025; 6(2):54. https://doi.org/10.3390/polysaccharides6020054
Chicago/Turabian StyleBabuty, Antoine, Agata Zykwinska, Sergey A. Samsonov, Nathalie Candia, Catherine Veinstein, Martine Pugnière, Thi Hong Giang Ngo, Corinne Sinquin, Javier Muñoz-Garcia, Sylvia Colliec-Jouault, and et al. 2025. "Anticoagulant Potential of Modified Sulfated Exopolysaccharides from Deep-Sea Bacteria: Toward Non-Animal Heparin Alternatives" Polysaccharides 6, no. 2: 54. https://doi.org/10.3390/polysaccharides6020054
APA StyleBabuty, A., Zykwinska, A., Samsonov, S. A., Candia, N., Veinstein, C., Pugnière, M., Ngo, T. H. G., Sinquin, C., Muñoz-Garcia, J., Colliec-Jouault, S., & Heymann, D. (2025). Anticoagulant Potential of Modified Sulfated Exopolysaccharides from Deep-Sea Bacteria: Toward Non-Animal Heparin Alternatives. Polysaccharides, 6(2), 54. https://doi.org/10.3390/polysaccharides6020054