
Lycium barbarum for Health and Longevity: A Review of Its Biological Significance

 , and
, and
Abstract
1. Introduction
2. Methodology
3. Phytochemistry
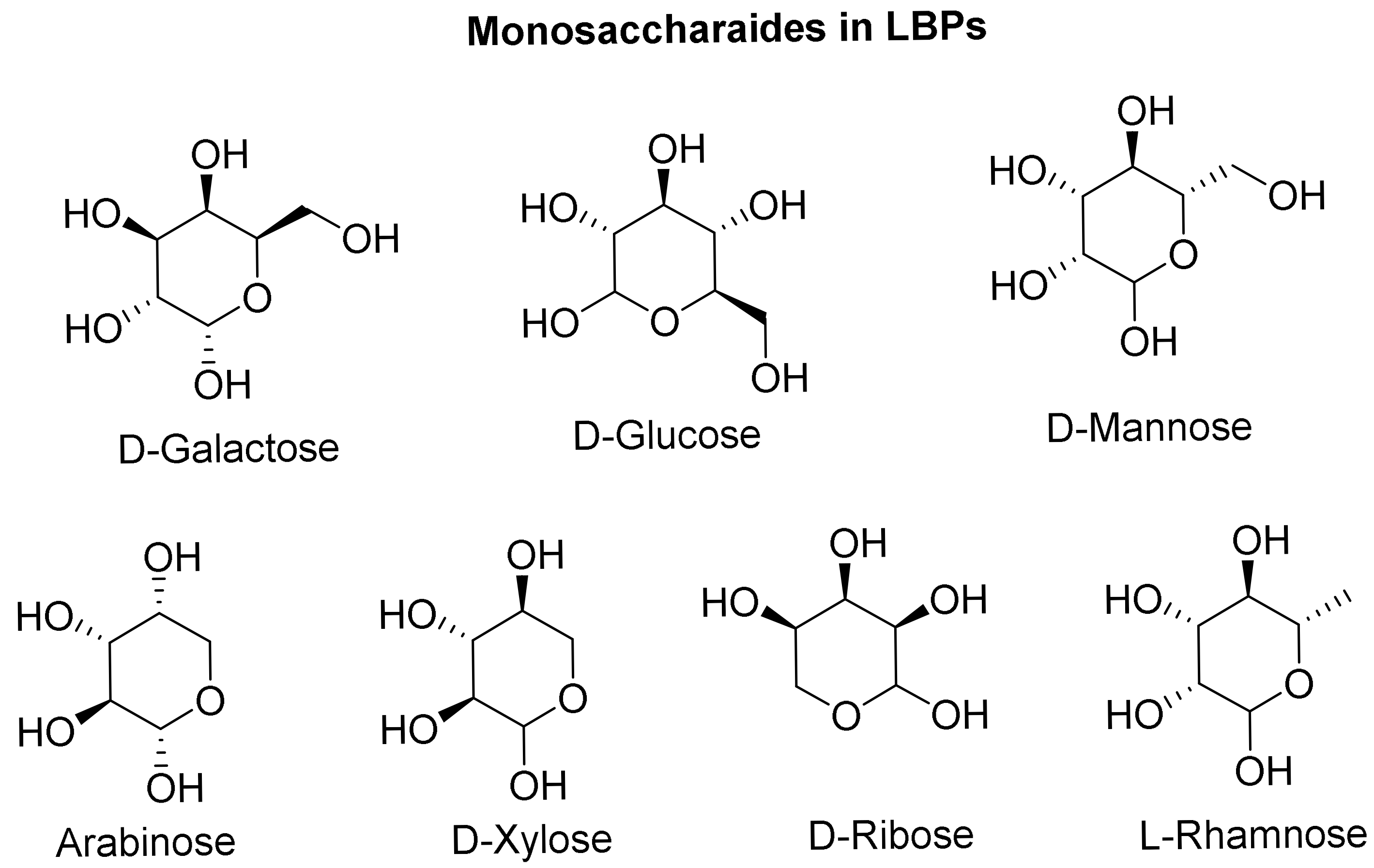
3.1. Polysaccharides
3.2. Polyphenols
3.3. Alkaloids
3.4. Carotenoids
4. Biological Properties
4.1. Cardiovascular Benefits
4.2. Glycaemic Control and Anti-Diabetic Activity
4.2.1. Obesity and Insulin Sensitivity
4.2.2. Glycaemic Control and Diabetes Management
4.2.3. Effects on Lipid Metabolism
4.2.4. Obesity-Related Bone Health
4.3. Antioxidant Activity

{kind=link}
{kind=link}
{kind=link}
| Model | Main Bioactive(s) | Key Findings | Mechanism/Pathway | Refs. |
|---|---|---|---|---|
| In vitro | Flavonoids, Zeaxanthin, LBP | Effective free radical scavenging | H-donation, metal chelation | [103] |
| In vitro | Phenolics, Flavonoids, Anthocyanins | Strong correlation with antioxidant capacity | Assay-based correlation | [104] |
| In vitro | Polyphenols, Organic acids | High TPC, FRAP, TBCC levels | Direct ROS scavenging | [51] |
| In vitro | Zeaxanthin, Carotenoids | ABTS/FRAP correlations | Antioxidant capacity | [74] |
| In vitro | Aqueous extract | ↑ GSH, ↓ lipid peroxidation and protein carbonyls | Antioxidant enzyme activation | [105] |
| In vitro | LBP | ↓ Apoptosis, ↑ cell protection against H2O2 | Nrf2 pathway | [106] |
| In vivo | LBP | ↑ SOD, ↑ CAT, ↑ GPx, ↓ MDA | Enzyme-mediated ROS defence | [107] |
| In vivo | LBP | Protected liver/kidney from ROS damage | Antioxidant defence | [108] |
4.4. Anti-Inflammatory Activity
4.5. Immunomodulatory Effects
4.6. Anticancer Activity
4.6.1. Inhibition of Cancer Cell Proliferation
4.6.2. Anti-Metastatic Properties
4.6.3. Synergistic Effects with Chemotherapy
4.6.4. Mitigation of Chemotherapy-Induced Toxicity
4.7. Hepatoprotective Activity
4.8. Antimicrobial Effect
4.9. Prebiotic Activity
4.10. Neuroprotective Effects
4.11. Anti-Aging Activity
4.11.1. Glycation and Oxidative Stress Reduction
4.11.2. Stem Cell Promotion and Tissue Regeneration
4.11.3. Anti-Apoptotic Effects
4.12. Ocular Health
4.12.1. Glaucoma
4.12.2. Retinitis Pigmentosa
4.12.3. Age-Related Macular Degeneration
4.12.4. Ocular Hypertension
4.12.5. Transient Retinal Ischaemia
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ABP | Achyranthes bidentata polysaccharide |
| ABTS | 2,2′-Azino-bis(3-ethylbenzothiazoline-6-sulfonic acid |
| ALT | Alanine aminotransferase |
| AMD | Age-related macular degeneration |
| AOH | Acute ocular hypertension |
| ARE | Antioxidant response element |
| ARPE | Arising retinal pigment epithelia |
| AST | Aspartate aminotransferase |
| ATF6 | Activating transcription factor 6 |
| ATGL | Adipose triglyceride lipase |
| ATK | Protein kinase B (also known as Akt) |
| B-ALP | Bone-alkaline phosphatase |
| Bax | Bcl-2-associated X protein |
| BclxL | B-cell lymphoma-extra large |
| CAT | Catalase |
| CDK | Cyclin-dependent kinase |
| Chrdl1 | Chordin-like 1 |
| CIH | Chronic intermittent hypoxia |
| CONT | Complete optic nerve transection |
| CPT1 | Carnitine palmitoyltransferase 1 |
| CVD | Cardiovascular disease |
| CypD | Cyclophilin D |
| CRP | C-reactive protein |
| CUPRAC | Cupric ion reducing antioxidant capacity |
| DPPH | 2,2-Diphenyl-1-picrylhydrazyl |
| eNOS | Endothelial nitric oxide synthase |
| ERK1/2 | Extracellular signal-regulated kinases 1 and 2 |
| FAS | Fatty acid synthase |
| FC | Folin–Ciocalteu |
| FGFR1 | Fibroblast growth factor receptor 1 |
| Fsp27 | Fat-specific protein 27 |
| FRAP | Ferric reducing antioxidant power |
| GAE | Gallic acid equivalents |
| GC | Gas chromatography |
| GSH | Glutathione |
| GPx | Glutathione peroxidase |
| GRK2 | G protein-coupled receptor kinase 2 |
| HO-1 | Haem oxygenase 1 |
| hPDLSCs | Human periodontal ligament stem cells |
| HPLC | High-performance liquid chromatography |
| IFN | Interferon |
| IGF | Insulin-like growth factor |
| IL | Interleukin |
| I/R | Ischaemia/reperfusion |
| JNK | c-Jun N-terminal kinase |
| L. Barbarum | Lycium Barbarum |
| LBLF | L. Barbarum leaf flavonoids |
| LBP | L. Barbarum polysaccharide |
| LC | Liquid chromatography |
| MAPK | Mitogen-activated protein kinase |
| MDA | Malondialdehyde |
| MDSCs | Myeloid-derived suppressor cells |
| mfERG | Multifocal electroretinography |
| MIC | Minimum inhibitory concentration |
| MMP-9 | Matrix metalloproteinase 9 |
| MS/MS | Tandem mass spectrometry |
| NASH | Non-alcoholic steatohepatitis |
| NF-κB | Nuclear factor kappa-light-chain-enhancer of activated B cells |
| NLRP3 | NOD-like receptor protein 3 |
| NMR | Nuclear magnetic resonance |
| NO | Nitric oxide |
| Nrf2 | Nuclear factor erythroid 2-related factor 2 |
| PI3K | Phosphoinositide 3-kinase |
| PONT | Partial optic nerve transection |
| PPAR-α | Peroxisome proliferator-activated receptor alpha |
| PPARγ | Peroxisome proliferator-activated receptor gamma |
| RCC | Renal cell carcinoma |
| RGC | Retinal ganglion cell |
| ROS | Reactive oxygen species |
| RSM | Response surface methodology |
| RP | Retinitis pigmentosa |
| SCFAs | Short-chain fatty acids |
| SIRT1 | Sirtuin 1 |
| SIRT3 | Sirtuin 3 |
| sONE | An antisense mRNA |
| SOD | Superoxide dismutase |
| STZ | Streptozotocin |
| TBCC | Total bioactive compound content |
| TFC | Total flavonoid content |
| TLR | Toll-like receptor |
| TNF | Tumour necrosis factor |
| TOR | Target of rapamycin |
| TPC | Total phenolic content |
| TQ | Triple quadrupole |
| TXNIP | Thioredoxin-interacting protein |
| UPLC | Ultra performance liquid chromatography |
| UVB | Ultraviolet B radiation |
References
- Matos, L.C.; Machado, J.P.; Monteiro, F.J.; Greten, H.J. Understanding traditional chinese medicine therapeutics: An overview of the basics and clinical applications. Healthcare 2021, 9, 257. [Google Scholar] [CrossRef] [PubMed]
- Smith, H.F.; Laitman, J.T. The yin, yang, and anatomy of traditional Chinese medicine in The Anatomical Record. Anat. Rec. 2023, 306, 2915–2919. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Fang, M.; Hu, Y.; Wang, X. Four types of traditional Chinese medicine inducing epileptic seizures. Seizure 2012, 21, 311–315. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Wei, Y.; Wang, Y.; Gao, F.; Chen, Z. Lycium barbarum: A traditional Chinese herb and a promising anti-aging agent. Aging Dis. 2017, 8, 778–791. [Google Scholar] [CrossRef]
- German Federal Institute for Risk Assessment. Superfoods—Super Good? German Federal Institute for Risk Assessment: Berlin, Germany, 2020. [Google Scholar]
- Vidović, B.B.; Milinčić, D.D.; Marčetić, M.D.; Djuriš, J.D.; Ilić, T.D.; Kostić, A.Ž.; Pešić, M.B. Health benefits and applications of goji berries in functional food products development: A review. Antioxidants 2022, 11, 248. [Google Scholar] [CrossRef]
- Yao, R.; Heinrich, M.; Zou, Y.; Reich, E.; Zhang, X.; Chen, Y.; Weckerle, C.S. Quality variation of Goji (fruits of Lycium spp.) in China: A comparative morphological and metabolomic analysis. Front. Pharmacol. 2018, 9, 151. [Google Scholar] [CrossRef]
- Wang, H.; Li, J.; Tao, W.; Zhang, X.; Gao, X.; Yong, J.; Zhao, J.; Zhang, L.; Li, Y.; Duan, J.-A. Lycium ruthenicum studies: Molecular biology, phytochemistry and pharmacology. Food Chem. 2018, 240, 759–766. [Google Scholar] [CrossRef]
- Rubio, R.M.; García, M.R.; Barroso, N.N.; Iñiguez, F.M.S.; Gómez, M.J.R.; Magro, P.C. Classification of goji berry (Lycium barbarum L.) varieties according to physicochemical and bioactive signature. Eur. Food Res. Technol. 2024, 251, 355–365. [Google Scholar] [CrossRef]
- Ju, Y.; Liu, H.; Niu, S.; Kang, L.; Ma, L.; Li, A.; Zhao, Y.; Yuan, Y.; Zhao, D. Optimizing geographical traceability models of Chinese Lycium barbarum: Investigating effects of region, cultivar, and harvest year on nutrients, bioactives, elements and stable isotope composition. Food Chem. 2025, 467, 142286. [Google Scholar] [CrossRef]
- Bertoldi, D.; Cossignani, L.; Blasi, F.; Perini, M.; Barbero, A.; Pianezze, S.; Montesano, D. Characterisation and geographical traceability of Italian goji berries. Food Chem. 2019, 275, 585–593. [Google Scholar] [CrossRef]
- Yao, R.; Heinrich, M.; Weckerle, C.S. The genus Lycium as food and medicine: A botanical, ethnobotanical and historical review. J. Ethnopharmacol. 2018, 212, 50–66. [Google Scholar] [CrossRef] [PubMed]
- Ju, Y.; Wang, Y.; Ma, L.; Kang, L.; Liu, H.; Ma, X.; Zhao, D. Comparative analysis of polyphenols in Lycium barbarum fruits using UPLC-IM-QTOF-MS. Molecules 2023, 28, 4930. [Google Scholar] [CrossRef] [PubMed]
- Ilić, T.; Dodevska, M.; Marčetić, M.; Božić, D.; Kodranov, I.; Vidović, B. Chemical characterization, antioxidant and antimicrobial properties of goji berries cultivated in Serbia. Foods 2020, 9, 1614. [Google Scholar] [CrossRef] [PubMed]
- Pires, T.C.S.P.; Dias, M.I.; Barros, L.; Calhelha, R.C.; Alves, M.J.; Santos-Buelga, C.; Ferreira, I.C.F.R. Phenolic compounds profile, nutritional compounds and bioactive properties of Lycium barbarum L.: A comparative study with stems and fruits. Ind. Crops Prod. 2018, 122, 574–581. [Google Scholar] [CrossRef]
- Lu, Y.; Guo, S.; Zhang, F.; Yan, H.; Qian, D.-W.; Wang, H.-Q.; Jin, L.; Duan, J.-A. Comparison of functional components and antioxidant activity of Lycium barbarum L. fruits from different regions in China. Molecules 2019, 24, 2228. [Google Scholar] [CrossRef]
- Liu, J.; Shi, X.; Lin, H.; He, C.; Li, Q.; Shen, G.; Feng, J. Geographical origin identification and quality comparison of Ningxia goji berries (Lycium barbarum L.) by NMR-based techniques. J. Food Compos. Anal. 2023, 119, 105258. [Google Scholar] [CrossRef]
- Berisha, A.; Alexa, E.-A.; Kelleher, R.; Zhang, T. From botany to bedside: A review of the health benefits of Lycium barbarum as a functional food. Explor. Foods Foodomics 2025, 3, 101070. [Google Scholar] [CrossRef]
- Ma, R.-H.; Zhang, X.-X.; Thakur, K.; Zhang, J.-G.; Wei, Z.-J. Research progress of Lycium barbarum L. as functional food: Phytochemical composition and health benefits. Curr. Opin. Food Sci. 2022, 47, 100871. [Google Scholar] [CrossRef]
- Sicari, V.; Romeo, R.; Mincione, A.; Santacaterina, S.; Tundis, R.; Loizzo, M.R. Ciabatta bread incorporating goji (Lycium barbarum L.): A new potential functional product with impact on human health. Foods 2023, 12, 566. [Google Scholar] [CrossRef]
- Blasi, F.; Rocchetti, G.; Montesano, D.; Lucini, L.; Chiodelli, G.; Ghisoni, S.; Baccolo, G.; Simonetti, M.S.; Cossignani, L. Changes in extra-virgin olive oil added with Lycium barbarum L. carotenoids during frying: Chemical analyses and metabolomic approach. Food Res. Int. 2018, 105, 507–516. [Google Scholar] [CrossRef]
- Teixeira, F.; Silva, A.M.; Delerue-Matos, C.; Rodrigues, F. Lycium barbarum berries (Solanaceae) as source of bioactive compounds for healthy purposes: A review. Int. J. Mol. Sci. 2023, 24, 4777. [Google Scholar] [CrossRef] [PubMed]
- Rjeibi, I.; Feriani, A.; Ben Saad, A.; Ncib, S.; Sdayria, J.; Saidi, I.; Souid, S.; Hfaiedh, N.; Allagui, M.S. Phytochemical characterization and bioactivity of Lycium europaeum: A focus on antioxidant, antinociceptive, hepatoprotective and nephroprotective effects. Biomed. Pharmacother. 2017, 95, 1441–1450. [Google Scholar] [CrossRef] [PubMed]
- Long, L.N.; Kang, B.J.; Jiang, Q.; Chen, J.S. Effects of dietary Lycium barbarum polysaccharides on growth performance, digestive enzyme activities, antioxidant status, and immunity of broiler chickens. Poult. Sci. 2020, 99, 744–751. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.; Zhang, G.; Sun, X.; He, S.; Dou, G. Distinct role of Lycium barbarum L. polysaccharides in oxidative stress-related ocular diseases. Pharmaceuticals 2023, 16, 215. [Google Scholar] [CrossRef]
- Potterat, O. Goji (Lycium barbarum and L. chinense): Phytochemistry, pharmacology and safety in the perspective of traditional uses and recent popularity. Planta Med. 2010, 76, 7–19. [Google Scholar] [CrossRef]
- Lopatriello, A.; Previtera, R.; Pace, S.; Werner, M.; Rubino, L.; Werz, O.; Taglialatela-Scafati, O.; Forino, M. NMR-based identification of the major bioactive molecules from an Italian cultivar of Lycium barbarum. Phytochemistry 2017, 144, 52–57. [Google Scholar] [CrossRef]
- Chao, S.; Schreuder, M.; Young, G.; Nakaoka, K.; Moyes, L.; Oberg, C. Preclinical study: Antioxidant levels and immunomodulatory effects of wolfberry juice and other juice mixtures in mice. Jana 2004, 7, 2–8. [Google Scholar]
- Kabir, F.; Katayama, S.; Tanji, N.; Nakamura, S. Antimicrobial effects of chlorogenic acid and related compounds. J. Korean Soc. Appl. Biol. Chem. 2014, 57, 359–365. [Google Scholar] [CrossRef]
- Ho, Y.-S.; Yu, M.-S.; Lai, C.S.-W.; So, K.-F.; Yuen, W.-H.; Chang, R.C.-C. Characterizing the neuroprotective effects of alkaline extract of Lycium barbarum on β-amyloid peptide neurotoxicity. Brain Res. 2007, 1158, 123–134. [Google Scholar] [CrossRef]
- Wawruszak, A.; Czerwonka, A.; Okła, K.; Rzeski, W. Anticancer effect of ethanol Lycium barbarum (Goji berry) extract on human breast cancer T47D cell line. Nat. Prod. Res. 2016, 30, 1993–1996. [Google Scholar] [CrossRef]
- Yu, Z.; Xia, M.; Lan, J.; Yang, L.; Wang, Z.; Wang, R.; Tao, H.; Shi, Y. A comprehensive review on the ethnobotany, phytochemistry, pharmacology and quality control of the genus Lycium in China. Food Funct. 2023, 14, 2998–3025. [Google Scholar] [CrossRef] [PubMed]
- Ma, R.H.; Zhang, X.X.; Ni, Z.J.; Thakur, K.; Wang, W.; Yan, Y.M.; Cao, Y.L.; Zhang, J.G.; Rengasamy, K.R.R.; Wei, Z.J. Lycium barbarum (Goji) as functional food: A review of its nutrition, phytochemical structure, biological features, and food industry prospects. Crit. Rev. Food Sci. Nutr. 2023, 63, 10621–10635. [Google Scholar] [CrossRef] [PubMed]
- Magiera, S.; Zaręba, M. Chromatographic Determination of Phenolic Acids and Flavonoids in Lycium barbarum L. and Evaluation of Antioxidant Activity. Food Anal. Methods 2015, 8, 2665–2674. [Google Scholar] [CrossRef]
- Qiang, X.; Xia, T.; Geng, B.; Zhao, M.; Li, X.; Zheng, Y.; Wang, M. Bioactive components of Lycium barbarum and deep-processing fermentation products. Molecules 2023, 28, 8044. [Google Scholar] [CrossRef]
- Chen, Y.; Yao, F.; Ming, K.; Wang, D.; Hu, Y.; Liu, J. Polysaccharides from Traditional Chinese Medicines: Extraction, Purification, Modification, and Biological Activity. Molecules 2016, 21, 1705. [Google Scholar] [CrossRef]
- Wang, Z.; Pan, H.; Xu, J.; Chang, Y.; Liu, C.; Zhang, Y.; Yang, H.; Duan, C.; Huang, J.; Fu, Y. A sustainable and integrated natural surfactant mediated microwave-assisted extraction technique enhances the extraction of phytochemicals from plants. Ind. Crops Prod. 2022, 184, 115043. [Google Scholar] [CrossRef]
- Perera, C.O.; Alzahrani, M.A.J. Ultrasound as a pre-treatment for extraction of bioactive compounds and food safety: A review. LWT 2021, 142, 111114. [Google Scholar] [CrossRef]
- Song, Y.-R.; Han, A.-R.; Park, S.-G.; Cho, C.-W.; Rhee, Y.-K.; Hong, H.-D. Effect of enzyme-assisted extraction on the physicochemical properties and bioactive potential of lotus leaf polysaccharides. Int. J. Biol. Macromol. 2020, 153, 169–179. [Google Scholar] [CrossRef]
- Gao, S.; Yan, S.; Zhou, Y.; Feng, Y.; Xie, X.; Guo, W.; Shen, Q.; Chen, C. Optimisation of enzyme-assisted extraction of Erythronium sibiricum bulb polysaccharide and its effects on immunomodulation. Glycoconj. J. 2022, 39, 357–368. [Google Scholar] [CrossRef]
- Masci, A.; Carradori, S.; Casadei, M.A.; Paolicelli, P.; Petralito, S.; Ragno, R.; Cesa, S. Lycium barbarum polysaccharides: Extraction, purification, structural characterisation and evidence about hypoglycaemic and hypolipidaemic effects. A review. Food Chem. 2018, 254, 377–389. [Google Scholar] [CrossRef]
- Yin, G.; Dang, Y. Optimization of extraction technology of the Lycium barbarum polysaccharides by Box–Behnken statistical design. Carbohydr. Polym. 2008, 74, 603–610. [Google Scholar] [CrossRef]
- Tian, X.; Liang, T.; Liu, Y.; Ding, G.; Zhang, F.; Ma, Z. Extraction, structural characterization, and biological functions of Lycium barbarum polysaccharides: A review. Biomolecules 2019, 9, 389. [Google Scholar] [CrossRef] [PubMed]
- Xia, F.; Zhou, S.; Sun, D. Surface optimization method to optimize the enzymatic extraction of Lycium barbarum polysaccharides. Chin. Wild Plant Resour. 2017, 36, 27–33. [Google Scholar]
- Kou, R.; Zuo, G.; Liu, J.; Di, D.; Guo, M. Structural properties and hypoglycaemic activity of polysaccharides extracted from the fruits of Lycium barbarum L. using various extraction media. Ind. Crops Prod. 2022, 188, 115725. [Google Scholar] [CrossRef]
- Zhang, Q.; Li, S. Effects of extraction methods on extraction ratio and antioxidant activity of polysaccharide from Lycium barbarum. Agricul. Sci. 2019, 47, 169–173. [Google Scholar]
- Quan, N.; Wang, Y.-D.; Li, G.-R.; Liu, Z.-Q.; Feng, J.; Qiao, C.-L.; Zhang, H.-F. Ultrasound–microwave combined extraction of novel polysaccharide fractions from Lycium barbarum leaves and their in vitro hypoglycemic and antioxidant activities. Molecules 2023, 28, 3880. [Google Scholar] [CrossRef]
- Zhu, L.; Peng, Z.; Zhang, X.; Yang, J.; Lai, X.; Yang, G. Determination of polyphenols in Lycium barbarum leaves by high-performance liquid chromatography–tandem mass spectrometry. Anal. Lett. 2017, 50, 761–776. [Google Scholar] [CrossRef]
- Islam, T.; Yu, X.; Badwal, T.S.; Xu, B. Comparative studies on phenolic profiles, antioxidant capacities and carotenoid contents of red goji berry (Lycium barbarum) and black goji berry (Lycium ruthenicum). Chem. Cent. J. 2017, 11, 59. [Google Scholar] [CrossRef]
- Ebeydulla, R.; Wang, X.; Zhao, L.; Xu, D.; Sun, H.; Ji, B.; Zhou, F. Nutritional and functional components of Chinese wolfberry dried by different drying methods. Food Sci. 2017, 38, 138–142. [Google Scholar]
- Donno, D.; Beccaro, G.L.; Mellano, M.G.; Cerutti, A.K.; Bounous, G. Goji berry fruit (Lycium spp.): Antioxidant compound fingerprint and bioactivity evaluation. J. Funct. Foods 2015, 18, 1070–1085. [Google Scholar] [CrossRef]
- Zhao, W.-H.; Shi, Y.-P. Comprehensive analysis of phenolic compounds in four varieties of goji berries at different ripening stages by UPLC–MS/MS. J. Food Compos. Anal. 2022, 106, 104279. [Google Scholar] [CrossRef]
- Liu, G.-T.; Li, Y.-L.; Wang, J.; Dong, C.-Z.; Deng, M.; Tai, M.; Deng, L.; Che, B.; Lin, L.; Du, Z.-Y.; et al. Improvement of skin barrier dysfunction by phenolic-containing extracts of Lycium barbarum via Nrf2/HO-1 regulation. Photochem. Photobiol. 2022, 98, 262–272. [Google Scholar] [CrossRef] [PubMed]
- Inbaraj, B.S.; Lu, H.; Kao, T.H.; Chen, B.H. Simultaneous determination of phenolic acids and flavonoids in Lycium barbarum Linnaeus by HPLC–DAD–ESI-MS. J. Pharm. Biomed. Anal. 2010, 51, 549–556. [Google Scholar] [CrossRef] [PubMed]
- Forino, M.; Tartaglione, L.; Dell’Aversano, C.; Ciminiello, P. NMR-based identification of the phenolic profile of fruits of Lycium barbarum (goji berries). Isolation and structural determination of a novel N-feruloyl tyramine dimer as the most abundant antioxidant polyphenol of goji berries. Food Chem. 2016, 194, 1254–1259. [Google Scholar] [CrossRef]
- Nardi, G.M.; Farias Januario, A.G.; Freire, C.G.; Megiolaro, F.; Schneider, K.; Perazzoli, M.R.; Do Nascimento, S.R.; Gon, A.C.; Mariano, L.N.; Wagner, G.; et al. Anti-inflammatory activity of berry fruits in mice model of inflammation is based on oxidative stress modulation. Pharmacognosy. Res. 2016, 8, S42–S49. [Google Scholar] [CrossRef]
- Mocan, A.; Vlase, L.; Vodnar, D.C.; Bischin, C.; Hanganu, D.; Gheldiu, A.M.; Oprean, R.; Silaghi-Dumitrescu, R.; Crișan, G. Polyphenolic content, antioxidant and antimicrobial activities of Lycium barbarum L. and Lycium chinense Mill. leaves. Molecules 2014, 19, 10056–10073. [Google Scholar] [CrossRef]
- Arumugam, M.K.; Paal, M.C.; Donohue, T.M.; Ganesan, M.; Osna, N.A.; Kharbanda, K.K. Beneficial effects of betaine: A comprehensive review. Biology 2021, 10, 456. [Google Scholar] [CrossRef]
- Qian, D.; Zhao, Y.; Yang, G.; Huang, L. Systematic review of chemical constituents in the genus Lycium (Solanaceae). Molecules 2017, 22, 911. [Google Scholar] [CrossRef]
- Tian, L.; Zhao, X.; Hu, Z.; Liu, J.; Ma, J.; Fan, Y.; Liu, D. iTRAQ-based proteomics identifies proteins associated with betaine accumulation in Lycium barbarum L. J. Proteom. 2024, 290, 105033. [Google Scholar] [CrossRef]
- Liu, W.; Xia, M.; Yang, L.; Wang, Z.; Wang, R.; Shi, Y. Development and optimization of a method for determining betaine and trigonelline in the fruits of Lycium species by using solid-phase extraction combined with high-performance liquid chromatography-diode array detector. J. Sep. Sci. 2020, 43, 2073–2078. [Google Scholar] [CrossRef]
- Yan, X.; Meng, X.; Pan, S.; Li, Z.; Song, Y. Determination of betaine in dried wolfberry by ion chromatography with pulsed amperometric detection. Food Sci. Technol. 2017, 42, 278–281. [Google Scholar]
- Chao, J.; Li, B.Y. Effects of Lycium barbarum polysaccharides extract and betaine on cell activation and liver fibrosis in rat hepatic stellate HSC-T6 cells. FASEB J. 2018, 31, 793–795. [Google Scholar] [CrossRef]
- Chen, D.; Guo, S.; Zhou, J.; Zhu, Y.; Zhang, F.; Zeng, F.; Duan, R.; Xu, M.; Duan, J.-A. Chemical constituents from Lycium barbarum (Solanaceae) and their chemophenetic significance. Biochem. Syst. Ecol. 2021, 97, 104292. [Google Scholar] [CrossRef]
- Chen, H.; Kong, J.-B.; Zhang, L.; Wang, H.-H.; Cao, Y.-G.; Zeng, M.-N.; Li, M.; Sun, Y.-J.; Du, K.; Xue, G.-M.; et al. Lycibarbarines A–C, three tetrahydroquinoline alkaloids possessing a spiro-heterocycle moiety from the fruits of Lycium barbarum. Org. Lett. 2021, 23, 858–862. [Google Scholar] [CrossRef]
- Kokotkiewicz, A.; Migas, P.; Stefanowicz, J.; Luczkiewicz, M.; Krauze-Baranowska, M. Densitometric TLC analysis for the control of tropane and steroidal alkaloids in Lycium barbarum. Food Chem. 2017, 221, 535–540. [Google Scholar] [CrossRef]
- Maoka, T. Carotenoids as natural functional pigments. J. Nat. Med. 2020, 74, 1–16. [Google Scholar] [CrossRef]
- Böhm, V. Carotenoids. Antioxidants 2019, 8, 516. [Google Scholar] [CrossRef]
- Ceccarini, M.R.; Vannini, S.; Cataldi, S.; Moretti, M.; Villarini, M.; Fioretti, B.; Albi, E.; Beccari, T.; Codini, M. In vitro protective effects of Lycium barbarum berries cultivated in Umbria (Italy) on human hepatocellular carcinoma cells. Biomed. Res. Int. 2016, 2016, 7529521. [Google Scholar] [CrossRef]
- Long, J.T.; Fan, H.X.; Zhou, Z.Q.; Sun, W.Y.; Li, Q.W.; Wang, Y.; Ma, M.; Gao, H.; Zhi, H. The major zeaxanthin dipalmitate derivatives from wolfberry. J. Asian Nat. Prod. Res. 2020, 22, 746–753. [Google Scholar] [CrossRef]
- Ren, R.; Li, Y.; Chen, H.; Wang, Y.; Yang, L.; Su, C.; Zhao, X.; Chen, J.; Ma, X. Carotenoid contents of Lycium barbarum: A novel QAMS analysis, geographical origins discriminant evaluation, and storage stability assessment. Molecules 2021, 26, 5374. [Google Scholar] [CrossRef]
- Yu, J.; Yan, Y.; Zhang, L.; Mi, J.; Yu, L.; Zhang, F.; Lu, L.; Luo, Q.; Li, X.; Zhou, X.; et al. A comprehensive review of goji berry processing and utilization. Food Sci. Nutr. 2023, 11, 7445–7457. [Google Scholar] [CrossRef] [PubMed]
- Inbaraj, B.S.; Lu, H.; Hung, C.F.; Wu, W.B.; Lin, C.L.; Chen, B.H. Determination of carotenoids and their esters in fruits of Lycium barbarum Linnaeus by HPLC-DAD-APCI-MS. J. Pharm. Biomed. Anal. 2008, 47, 812–818. [Google Scholar] [CrossRef] [PubMed]
- Wojdyło, A.; Nowicka, P.; Bąbelewski, P. Phenolic and carotenoid profile of new goji cultivars and their anti-hyperglycemic, anti-aging and antioxidant properties. J. Funct. Foods 2018, 48, 632–642. [Google Scholar] [CrossRef]
- Hsu, H.J.; Huang, R.F.; Kao, T.H.; Inbaraj, B.S.; Chen, B.H. Preparation of carotenoid extracts and nanoemulsions from Lycium barbarum L. and their effects on growth of HT-29 colon cancer cells. Nanotechnology 2017, 28, 135103. [Google Scholar] [CrossRef]
- Langi, P.; Kiokias, S.; Varzakas, T.; Proestos, C. Carotenoids: From plants to food and feed industries. In Microbial Carotenoids: Methods and Protocols; Barreiro, C., Barredo, J.-L., Eds.; Springer: New York, NY, USA, 2018; pp. 57–71. [Google Scholar]
- Wu, H.; Liu, Y.; Hao, Y.; Hou, D.; Yang, R. Lycium barbarum polysaccharide protects cardiomyocytes from hypoxia/reoxygenation injury via activation of SIRT3/CypD signaling. Ann. Transl. Med. 2023, 11, 72. [Google Scholar] [CrossRef]
- Li, Y.; Yang, B.; Zhang, X.; Shen, X.; Ma, Y.; Jing, L. Lycium barbarum polysaccharide antagonizes cardiomyocyte apoptosis by inhibiting the upregulation of GRK2 induced by I/R injury, and salvage mitochondrial fission/fusion imbalance and AKT/eNOS signaling. Cell. Signal. 2022, 92, 110252. [Google Scholar] [CrossRef]
- Xue, S.; Hu, X.; Zhu, L.; Nie, L.; Li, G. Protective functions of Lycium barbarum polysaccharides in H(2)O(2)-injured vascular endothelial cells through anti-oxidation and anti-apoptosis effects. Biomed. Rep. 2019, 11, 207–214. [Google Scholar] [CrossRef]
- Zhang, X.; Yang, X.; Lin, Y.; Suo, M.; Gong, L.; Chen, J.; Hui, R. Anti-hypertensive effect of Lycium barbarum L. with down-regulated expression of renal endothelial lncRNA sONE in a rat model of salt-sensitive hypertension. Int. J. Clin. Exp. Pathol. 2015, 8, 6981–6987. [Google Scholar]
- Huang, R.; Wu, E.; Deng, X. Potential of Lycium barbarum polysaccharide for the control of glucose and lipid metabolism disorders: A review. Int. J. Food Prop. 2022, 25, 673–680. [Google Scholar] [CrossRef]
- Liu, H.; Cui, B.; Zhang, Z. Mechanism of glycometabolism regulation by bioactive compounds from the fruits of Lycium barbarum: A review. Food Res. Int. 2022, 159, 111408. [Google Scholar] [CrossRef]
- Zhou, R.; Liu, Y.; Hu, W.; Yang, J.; Lin, B.; Zhang, Z.; Chen, M.; Yi, J.; Zhu, C. Lycium barbarum polysaccharide ameliorates the accumulation of lipid droplets in adipose tissue via an ATF6/SIRT1-dependent mechanism. Acta Biochim. Biophys. Sin. 2024, 56, 844–856. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Li, Y.; Dai, Y.; Ma, L.; Di, D.; Liu, J. Screening, structural characterization and anti-adipogenesis effect of a water-soluble polysaccharide from Lycium barbarum L. by an activity-oriented approach. Food Biosci. 2023, 53, 102502. [Google Scholar] [CrossRef]
- Yang, F.L.; Wei, Y.X.; Liao, B.Y.; Wei, G.J.; Qin, H.M.; Pang, X.X.; Wang, J.L. Effects of Lycium barbarum polysaccharide on endoplasmic reticulum stress and oxidative stress in obese mice. Front. Pharmacol. 2020, 11, 742. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Ma, L.; Ma, J.; Liu, S.; Fu, J.; Fan, Y.; Liu, Y. Lycium barbarum leaf flavonoids ameliorate high fructose induced insulin resistance in mice by regulating blood glucose and gut microbiota composition. Food Biosci. 2024, 62, 105087. [Google Scholar] [CrossRef]
- Ren, Y.; Wang, K.; Wu, Y.; Li, J.; Ma, J.; Wang, L.; Zhang, C.; Li, J.; Wei, Y.; Yang, Y. Lycium barbarum polysaccharide mitigates high-fat-diet-induced skeletal muscle atrophy by promoting AMPK/PINK1/Parkin-mediated mitophagy. Int. J. Biol. Macromol. 2025, 301, 140488. [Google Scholar] [CrossRef]
- Kou, L.; Du, M.; Zhang, C.; Dai, Z.; Li, X.; Zhang, B. The Hypoglycemic, Hypolipidemic, and Anti-Diabetic Nephritic Activities of Zeaxanthin in Diet-Streptozotocin-Induced Diabetic Sprague Dawley Rats. Appl. Biochem. Biotechnol. 2017, 182, 944–955. [Google Scholar] [CrossRef]
- Li, D.; Zhang, X.; Fan, Y.; Zhang, Y.; Tao, X.; Yang, J. Lycium barbarum polysaccharides improved glucose metabolism in prediabetic mice by regulating duodenal contraction. Nutrients 2023, 15, 4437. [Google Scholar] [CrossRef]
- Wu, H.; Guo, H.; Zhao, R. Effect of Lycium barbarum polysaccharide on the improvement of antioxidant ability and DNA damage in NIDDM rats. Yakugaku Zasshi 2006, 126, 365–371. [Google Scholar] [CrossRef]
- Zhao, X.-q.; Guo, S.; Lu, Y.-y.; Hua, Y.; Zhang, F.; Yan, H.; Shang, E.-x.; Wang, H.-q.; Zhang, W.-h.; Duan, J.-a. Lycium barbarum L. leaves ameliorate type 2 diabetes in rats by modulating metabolic profiles and gut microbiota composition. Biomed. Pharmacother. 2020, 121, 109559. [Google Scholar] [CrossRef]
- Xia, H.; Zhou, B.; Sui, J.; Ma, W.; Wang, S.; Yang, L.; Sun, G. Lycium barbarum polysaccharide regulates lipid metabolism and alters gut microbiota in high-fat diet-induced obese mice. Int. J. Environ. Res. Public Health 2022, 19, 12093. [Google Scholar] [CrossRef]
- Huang, Z.; Ye, Y.; Long, Z.; Qin, H.; Liu, L.; Xu, A.; Li, Z. Lycium barbarum polysaccharides improve lipid metabolism disorders of spotted sea bass Lateolabrax maculatus induced by high lipid diet. Int. J. Biol. Macromol. 2023, 242, 125122. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Li, J.; Fan, Z.; Sun, Z.; Zheng, X.; Zhang, H.; Xu, H.; Wang, L. Dietary Lycium barbarum polysaccharide modulates growth performance, antioxidant capacity, and lipid metabolism in common carp (Cyprinus carpio) fed with high-fat diet. Antioxidants 2024, 13, 540. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.N.; Chi, X.; Yan, L.; Pu, Z.Y.; Yang, J.J.; Zhang, Y.N. Lycium barbarum polysaccharides regulate the gut microbiota to modulate metabolites in high-fat diet-induced obese rats. J. Asian Nat. Prod. Res. 2024, 26, 1115–1129. [Google Scholar] [CrossRef] [PubMed]
- Jing, L.; Hu, B.; Song, Q.H. Lycium barbarum polysaccharide (LBP) inhibits palmitic acid (PA)-induced MC3T3-E1 cell apoptosis by regulating miR-200b-3p/Chrdl1/PPARγ. Food Nutr. Res. 2020, 64, 10-29219. [Google Scholar] [CrossRef]
- Li, Z.-X.; Zhuo, J.-L.; Yang, N.; Gao, M.-B.; Qu, Z.-H.; Han, T. Effect of Lycium barbarum polysaccharide on osteoblast proliferation and differentiation in postmenopausal osteoporosis. Int. J. Mol. Sci. 2024, 271, 132415. [Google Scholar] [CrossRef]
- Piñar-Gutierrez, A.; García-Fontana, C.; García-Fontana, B.; Muñoz-Torres, M. Obesity and bone health: A complex relationship. Int. J. Mol. Sci. 2022, 23, 8303. [Google Scholar] [CrossRef]
- Tan, B.L.; Norhaizan, M.E.; Liew, W.P.; Sulaiman Rahman, H. Antioxidant and oxidative stress: A mutual interplay in age-related diseases. Front. Pharmacol. 2018, 9, 1162. [Google Scholar] [CrossRef]
- Yun, D.; Yan, Y.; Liu, J. Isolation, structure and biological activity of polysaccharides from the fruits of Lycium ruthenicum Murr: A review. Carbohydr. Polym. 2022, 291, 119618. [Google Scholar] [CrossRef]
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative stress: Harms and benefits for human health. Oxid. Med. Cell. Longev. 2017, 2017, 8416763. [Google Scholar] [CrossRef]
- Rumpf, J.; Burger, R.; Schulze, M. Statistical evaluation of DPPH, ABTS, FRAP, and Folin-Ciocalteu assays to assess the antioxidant capacity of lignins. Int. J. Biol. Macromol. 2023, 233, 123470. [Google Scholar] [CrossRef]
- Wang, C.C.; Chang, S.C.; Inbaraj, B.S.; Chen, B.H. Isolation of carotenoids, flavonoids and polysaccharides from Lycium barbarum L. and evaluation of antioxidant activity. Food Chem. 2010, 120, 184–192. [Google Scholar] [CrossRef]
- de Nijs, M.; Crews, C.; Dorgelo, F.; MacDonald, S.; Mulder, P.P.J. Emerging issues on tropane alkaloid contamination of food in Europe. Toxins 2023, 15, 98. [Google Scholar] [CrossRef] [PubMed]
- Skenderidis, P.; Kerasioti, E.; Karkanta, E.; Stagos, D.; Kouretas, D.; Petrotos, K.; Hadjichristodoulou, C.; Tsakalof, A. Assessment of the antioxidant and antimutagenic activity of extracts from goji berry of Greek cultivation. Toxicol. Rep. 2018, 5, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Liang, R.; Zhao, Q.; Zhu, Q.; He, X.; Gao, M.; Wang, Y. Lycium barbarum polysaccharide protects ARPE-19 cells against H2O2-induced oxidative stress via the Nrf2/HO-1 pathway. Mol. Med. Rep. 2021, 24, 769. [Google Scholar] [CrossRef]
- Pehlİvan KarakaŞ, F.; CoŞkun, H.; SoytÜrk, H.; Bozat, B.G. Anxiolytic, antioxidant, and neuroprotective effects of goji berry polysaccharides in ovariectomized rats: Experimental evidence from behavioral, biochemical, and immunohistochemical analyses. Turk. J. Biol. 2020, 44, 238–251. [Google Scholar] [CrossRef]
- Li, X.M. Protective effect of Lycium barbarum polysaccharides on streptozotocin-induced oxidative stress in rats. Int. J. Biol. Macromol. 2007, 40, 461–465. [Google Scholar] [CrossRef]
- Ma, Z.; Du, B.; Li, J.; Yang, Y.; Zhu, F. An insight into anti-inflammatory activities and inflammation related diseases of anthocyanins: A review of both in vivo and in vitro investigations. Int. J. Mol. Sci. 2021, 22, 11076. [Google Scholar] [CrossRef]
- Mitchell, S.; Vargas, J.; Hoffmann, A. Signaling via the NFκB system. Wiley Interdiscip. Rev. Syst. Biol. Med. 2016, 8, 227–241. [Google Scholar] [CrossRef]
- Lee, S.; Jeong, S.; Park, Y.; Seo, H.; You, C.; Hwang, U.; Park, H.; Suh, H.-j. Supplementation of non-fermented and fermented goji berry (Lycium barbarum) improves hepatic function and corresponding lipid metabolism via their anti-inflammatory and antioxidant properties in high fat-fed rats. Appl. Biol. Chem. 2021, 64, 70. [Google Scholar] [CrossRef]
- Xiao, J.; Xing, F.; Huo, J.; Fung, M.L.; Liong, E.C.; Ching, Y.P.; Xu, A.; Chang, R.C.C.; So, K.F.; Tipoe, G.L. Lycium barbarum polysaccharides therapeutically improve hepatic functions in non-alcoholic steatohepatitis rats and cellular steatosis model. Sci. Rep. 2014, 4, 5587. [Google Scholar] [CrossRef]
- Du, M.; Hu, X.; Kou, L.; Zhang, B.; Zhang, C. Lycium barbarum polysaccharide mediated the antidiabetic and antinephritic effects in diet-streptozotocin-induced diabetic Sprague Dawley rats via regulation of NF-κB. Biomed. Res. Int. 2016, 2016, 3140290. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.; Huang, Y.Y.; Chen, H.G.; Zhou, X. Study on the efficacy and mechanism of Lycium barbarum polysaccharide against lead-induced renal Iinjury in mice. Nutrients 2021, 13, 2945. [Google Scholar] [CrossRef] [PubMed]
- Gan, F.; Liu, Q.; Liu, Y.; Huang, D.; Pan, C.; Song, S.; Huang, K. Lycium barbarum polysaccharides improve CCl(4)-induced liver fibrosis, inflammatory response and TLRs/NF-kB signaling pathway expression in wistar rats. Life Sci. 2018, 192, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Li, J.; Yan, J.; Liu, S.; Guo, Y.; Chen, D.; Luo, Q. Lycium barbarum polysaccharides ameliorates renal injury and inflammatory reaction in alloxan-induced diabetic nephropathy rabbits. Life Sci. 2016, 157, 82–90. [Google Scholar] [CrossRef]
- Gan, L.; Zhang, S.H. Effect of Lycium barbarum polysaccharides on anti-tumor activity and immune function. Acta Nutr. Sin. 2003, 25, 200–202. [Google Scholar]
- Deng, X.; Luo, S.; Luo, X.; Hu, M.; Ma, F.; Wang, Y.; Lai, X.; Zhou, L. Polysaccharides from Chinese herbal Lycium barbarum induced systemic and local immune responses in H22 tumor-bearing Mice. J. Immunol. Res. 2018, 2018, 3431782. [Google Scholar] [CrossRef]
- Feng, L.; Xiao, X.; Liu, J.; Wang, J.; Zhang, N.; Bing, T.; Liu, X.; Zhang, Z.; Shangguan, D. Immunomodulatory effects of Lycium barbarum polysaccharide extract and its uptake behaviors at the cellular level. Molecules 2020, 25, 1351. [Google Scholar] [CrossRef]
- Liu, Y.L.; Yin, R.Q.; Liang, S.S.; Duan, Y.L.; Yao, J.H.; Duan, Y.L.; Yang, X.J. Effect of dietary Lycium barbarum polysaccharide on growth performance and immune function of broilers. J. Appl. Poult. Res. 2017, 26, 200–208. [Google Scholar] [CrossRef]
- Qian, L. Modulation of cytokine level and sperm quality of mice by Lycium barbarum polysaccharides. Int. J. Biol. Macromol. 2019, 126, 475–477. [Google Scholar] [CrossRef]
- Ding, Y.; Yan, Y.; Chen, D.; Ran, L.; Mi, J.; Lu, L.; Jing, B.; Li, X.; Zeng, X.; Cao, Y. Modulating effects of polysaccharides from the fruits of Lycium barbarum on the immune response and gut microbiota in cyclophosphamide-treated mice. Food Funct. 2019, 10, 3671–3683. [Google Scholar] [CrossRef]
- Guo, M.; Jin, J.; Zhao, D.; Rong, Z.; Cao, L.-Q.; Li, A.-H.; Sun, X.-Y.; Jia, L.-Y.; Wang, Y.-D.; Huang, L.; et al. Research advances on anti-cancer natural products. Front. Oncol. 2022, 12, 866154. [Google Scholar] [CrossRef] [PubMed]
- Naeem, A.; Hu, P.; Yang, M.; Zhang, J.; Liu, Y.; Zhu, W.; Zheng, Q. Natural products as anticancer agents: Current status and future perspectives. Molecules 2022, 27, 8367. [Google Scholar] [CrossRef] [PubMed]
- Miranda, M.R.; Vestuto, V.; Amodio, G.; Manfra, M.; Pepe, G.; Campiglia, P. Antitumor mechanisms of Lycium barbarum fruit: An overview of in vitro and in vivo potential. Life 2024, 14, 420. [Google Scholar] [CrossRef] [PubMed]
- Kwaśnik, P.; Lemieszek, M.K.; Rzeski, W. Impact of phytochemicals and plant extracts on viability and proliferation of NK cell line NK-92—A closer look at immunomodulatory properties of goji berries extract in human colon cancer cells. Ann. Agric. Environ. Med. 2021, 28, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Georgiev, K.D.; Slavov, I.J.; Iliev, I.A. Antioxidant activity and antiproliferative effects of Lycium barbarum’s (Goji berry) fractions on breast cancer cell lines. Folia Med. 2019, 61, 104–112. [Google Scholar] [CrossRef]
- Gong, G.; Liu, Q.; Deng, Y.; Dang, T.; Dai, W.; Liu, T.; Liu, Y.; Sun, J.; Wang, L.; Liu, Y.; et al. Arabinogalactan derived from Lycium barbarum fruit inhibits cancer cell growth via cell cycle arrest and apoptosis. Int. J. Biol. Macromol. 2020, 149, 639–650. [Google Scholar] [CrossRef]
- Zhang, M.; Chen, H.; Huang, J.; Li, Z.; Zhu, C.; Zhang, S. Effect of Lycium barbarum polysaccharide on human hepatoma QGY7703 cells: Inhibition of proliferation and induction of apoptosis. Life Sci. 2005, 76, 2115–2124. [Google Scholar] [CrossRef]
- Deng, X.; Li, X.; Luo, S.; Zheng, Y.; Luo, X.; Zhou, L. Antitumor activity of Lycium barbarum polysaccharides with different molecular weights: An in vitro and in vivo study. Food Nutr. Res. 2017, 61, 1399770. [Google Scholar] [CrossRef]
- Sanghavi, A.; Srivatsa, A.; Adiga, D.; Chopra, A.; Lobo, R.; Kabekkodu, S.P.; Gadag, S.; Nayak, U.; Sivaraman, K.; Shah, A. Goji berry (Lycium barbarum) inhibits the proliferation, adhesion, and migration of oral cancer cells by inhibiting the ERK, AKT, and CyclinD cell signaling pathways: An in-vitro study. F1000Research 2022, 11, 1563. [Google Scholar] [CrossRef]
- Liu, Y.; Du, Y.; Gao, L.; Ma, L. Effect of Lycium barbarum polysaccharide combined with cisplatin on the proliferation of human lung cancer cells. Pharmacogn. Mag. 2022, 18, 378–385. [Google Scholar]
- Chen, S.; Liang, L.; Wang, Y.; Diao, J.; Zhao, C.; Chen, G.; He, Y.; Luo, C.; Wu, X.; Zhang, Y. Synergistic immunotherapeutic effects of Lycium barbarum polysaccharide and interferon-α2b on the murine Renca renal cell carcinoma cell line in vitro and in vivo. Mol. Med. Rep. 2015, 12, 6727–6737. [Google Scholar] [CrossRef] [PubMed]
- Xin, Y.-F.; Wan, L.-L.; Peng, J.-L.; Guo, C. Alleviation of the acute doxorubicin-induced cardiotoxicity by Lycium barbarum polysaccharides through the suppression of oxidative stress. Food Chem. Toxicol. 2011, 49, 259–264. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Luo, S.; Luo, X.; Hu, M.; Ma, F.; Wang, Y.; Zhou, L.; Huang, R. Fraction from Lycium barbarum polysaccharides reduces immunotoxicity and enhances antitumor activity of doxorubicin in mice. Integr. Cancer Ther. 2018, 17, 860–866. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liang, X.; Li, Y.; Fan, Y.; Li, Y.; Cao, Y.; An, W.; Shi, Z.; Zhao, J.; Guo, S. Changes in metabolome and nutritional quality of Lycium barbarum fruits from three typical growing areas of China as revealed by widely targeted metabolomics. Metabolites 2020, 10, 46. [Google Scholar] [CrossRef]
- Xiao, J.; Zhu, Y.; Liu, Y.; Tipoe, G.L.; Xing, F.; So, K.-F. Lycium barbarum polysaccharide attenuates alcoholic cellular injury through TXNIP-NLRP3 inflammasome pathway. Int. J. Biol. Macromol. 2014, 69, 73–78. [Google Scholar] [CrossRef]
- Gündüz, E.; Dursun, R.; Zengin, Y.; İçer, M.; Durgun, H.M.; Kanıcı, A.; Kaplan, İ.; Alabalık, U.; Gürbüz, H.; Güloğlu, C. Lycium barbarum extract provides effective protection against paracetamol-induced acute hepatotoxicity in rats. Int. J. Clin. Exp. Med. 2015, 8, 7898–7905. [Google Scholar]
- Cheng, D.; Kong, H. The effect of Lycium barbarum polysaccharide on alcohol-induced oxidative stress in rats. Molecules 2011, 16, 2542–2550. [Google Scholar] [CrossRef]
- Sanghavi, A.D.; Chopra, A.; Shah, A.; Lobo, R.; Shenoy, P.A. Antimicrobial, anti-adhesion, anti-biofilm properties of goji berry (Lycium barbarum) against periodontal bacteria: Potential benefits for periodontal diseases. J. Complement. Integr. Med. 2023, 20, 129–136. [Google Scholar] [CrossRef]
- Zhu, W.; Zhou, S.; Liu, J.; McLean, R.J.C.; Chu, W. Prebiotic, immuno-stimulating and gut microbiota-modulating effects of Lycium barbarum polysaccharide. Biomed. Pharmacother. 2020, 121, 109591. [Google Scholar] [CrossRef]
- Guo, L.; Guan, Q.; Duan, W.; Ren, Y.; Zhang, X.J.; Xu, H.Y.; Shi, J.S.; Wang, F.Z.; Lu, R.; Zhang, H.L.; et al. Dietary goji shapes the gut microbiota to prevent liver injury induced by acute alcohol intake. Front. Nutr. 2022, 9, 929776. [Google Scholar] [CrossRef]
- Skenderidis, P.; Mitsagga, C.; Lampakis, D.; Petrotos, K.; Giavasis, I. The effect of encapsulated powder of goji berry (Lycium barbarum) on growth and survival of probiotic bacteria. Microorganisms 2020, 8, 57. [Google Scholar] [CrossRef] [PubMed]
- Orsini, M.; Nascimento, O.J.; Matta, A.P.; Reis, C.H.; de Souza, O.G.; Bastos, V.H.; Moreira, R.; Ribeiro, P.; Fiorelli, S.; Novellino, P.; et al. Revisiting the term neuroprotection in chronic and degenerative diseases. Neurol. Int. 2016, 8, 6311. [Google Scholar] [CrossRef] [PubMed]
- Karvandi, M.S.; Sheikhzadeh Hesari, F.; Aref, A.R.; Mahdavi, M. The neuroprotective effects of targeting key factors of neuronal cell death in neurodegenerative diseases: The role of ER stress, oxidative stress, and neuroinflammation. Front. Cell. Neurosci. 2023, 17, 1105247. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Zhang, X.; Chen, L.; Ren, B.X.; Tang, F.R. Lycium barbarum ameliorates neural damage induced by experimental ischemic stroke and radiation exposure. Front. Biosci. 2023, 28, 38. [Google Scholar] [CrossRef]
- Lam, C.S.; Tipoe, G.L.; So, K.F.; Fung, M.L. Neuroprotective mechanism of Lycium barbarum polysaccharides against hippocampal-dependent spatial memory deficits in a rat model of obstructive sleep apnea. PLoS ONE 2015, 10, e0117990. [Google Scholar] [CrossRef]
- Cao, S.; Du, J.; Hei, Q. Lycium barbarum polysaccharide protects against neurotoxicity via the Nrf2-HO-1 pathway. Exp. Ther. Med. 2017, 14, 4919–4927. [Google Scholar] [CrossRef]
- Yang, Y.; Yu, L.; Zhu, T.; Xu, S.; He, J.; Mao, N.; Liu, Z.; Wang, D. Neuroprotective effects of Lycium barbarum polysaccharide on light-induced oxidative stress and mitochondrial damage via the Nrf2/HO-1 pathway in mouse hippocampal neurons. Int. J. Biol. Macromol. 2023, 251, 126315. [Google Scholar] [CrossRef]
- Hu, X.; Qu, Y.; Chu, Q.; Li, W.; He, J. Investigation of the neuroprotective effects of Lycium barbarum water extract in apoptotic cells and Alzheimer’s disease mice. Mol. Med. Rep. 2018, 17, 3599–3606. [Google Scholar] [CrossRef]
- Yao, X.-l.; Wu, W.-l.; Zheng, M.-y.; Li, W.; Ye, C.-h.; Lu, X.-l. Protective effects of Lycium barbarum extract against MPP(+) -induced neurotoxicity in Caenorhabditis elegans and PC12 cells. Zhong Yao Cai 2011, 34, 1241–1246. [Google Scholar]
- Lp, S.; Rr, U.; Mardhekar, V. Neuroprotective effects of Lycium barbarum in ischemic stroke: Current perspectives. Int. J. Pharmacogn. Chin. Med. 2019, 3, 126315. [Google Scholar] [CrossRef]
- Deng, H.B.; Cui, D.P.; Jiang, J.M.; Feng, Y.C.; Cai, N.S.; Li, D.D. Inhibiting effects of Achyranthes bidentata polysaccharide and Lycium barbarum polysaccharide on nonenzyme glycation in D-galactose induced mouse aging model. Biomed. Environ. Sci. 2003, 16, 267–275. [Google Scholar]
- Tang, R.; Chen, X.; Dang, T.; Deng, Y.; Zou, Z.; Liu, Q.; Gong, G.; Song, S.; Ma, F.; Huang, L.; et al. Lycium barbarum polysaccharides extend the mean lifespan of Drosophila melanogaster. Food Funct. 2019, 10, 4231–4241. [Google Scholar] [CrossRef]
- Li, H.; Li, Z.; Peng, L.; Jiang, N.; Liu, Q.; Zhang, E.; Liang, B.; Li, R.; Zhu, H. Lycium barbarum polysaccharide protects human keratinocytes against UVB-induced photo-damage. Free Radic. Res. 2017, 51, 200–210. [Google Scholar] [CrossRef]
- Meng, J.; Lv, Z.; Chen, X.; Sun, C.; Jin, C.; Ding, K.; Chen, C. LBP1C-2 from Lycium barbarum maintains skeletal muscle satellite cell pool by interaction with FGFR1. iScience 2023, 26, 106573. [Google Scholar] [CrossRef]
- Lai, S.; Liu, C.; Liu, C.; Fan, L.; Li, X.; Yang, Y.; Zhu, Y.; Deng, L.; Xiao, L.; Mu, Y. Lycium barbarum polysaccharide-glycoprotein promotes osteogenesis in hPDLSCs via ERK activation. Oral Dis. 2023, 29, 3503–3513. [Google Scholar] [CrossRef]
- Xia, G.; Xin, N.; Liu, W.; Yao, H.; Hou, Y.; Qi, J. Inhibitory effect of Lycium barbarum polysaccharides on cell apoptosis and senescence is potentially mediated by the p53 signaling pathway. Mol. Med. Rep. 2014, 9, 1237–1241. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, H.; Sheng, X.; Gambino, P.E.; Costello, B.; Bojanowski, K. Protective effect of Fructus Lycii polysaccharides against time and hyperthermia-induced damage in cultured seminiferous epithelium. J. Ethnopharmacol. 2002, 82, 169–175. [Google Scholar] [CrossRef]
- Yu, Y.; Wu, X.; Pu, J.; Luo, P.; Ma, W.; Wang, J.; Wei, J.; Wang, Y.; Fei, Z. Lycium barbarum polysaccharide protects against oxygen glucose deprivation/reoxygenation-induced apoptosis and autophagic cell death via the PI3K/Akt/mTOR signaling pathway in primary cultured hippocampal neurons. Biochem. Biophys. Res. Commun. 2018, 495, 1187–1194. [Google Scholar] [CrossRef]
- Zhou, L.; Pinho, R.; Gu, Y.; Radak, Z. The role of SIRT3 in exercise and aging. Cells 2022, 11, 2596. [Google Scholar] [CrossRef]
- He, M.; Pan, H.; Chang, R.C.; So, K.F.; Brecha, N.C.; Pu, M. Activation of the Nrf2/HO-1 antioxidant pathway contributes to the protective effects of Lycium barbarum polysaccharides in the rodent retina after ischemia-reperfusion-induced damage. PLoS ONE 2014, 9, e84800. [Google Scholar] [CrossRef]
- Li, H.; Liang, Y.; Chiu, K.; Yuan, Q.; Lin, B.; Chang, R.C.; So, K.F. Lycium barbarum (wolfberry) reduces secondary degeneration and oxidative stress, and inhibits JNK pathway in retina after partial optic nerve transection. PLoS ONE 2013, 8, e68881. [Google Scholar] [CrossRef]
- Chu, P.H.; Li, H.Y.; Chin, M.P.; So, K.F.; Chan, H.H. Effect of Lycium barbarum (wolfberry) polysaccharides on preserving retinal function after partial optic nerve transection. PLoS ONE 2013, 8, e81339. [Google Scholar] [CrossRef]
- Li, H.Y.; Huang, M.; Luo, Q.Y.; Hong, X.; Ramakrishna, S.; So, K.F. Lycium barbarum (wolfberry) increases retinal ganglion cell survival and affects both microglia/macrophage polarization and autophagy after rat partial optic nerve transection. Cell Transpl. 2019, 28, 607–618. [Google Scholar] [CrossRef]
- Wang, K.; Xiao, J.; Peng, B.; Xing, F.; So, K.F.; Tipoe, G.L.; Lin, B. Retinal structure and function preservation by polysaccharides of wolfberry in a mouse model of retinal degeneration. Sci. Rep. 2014, 4, 7601. [Google Scholar] [CrossRef]
- Zheng, H.-l.; Li, M.-t.; Zhou, T.; Wang, Y.-y.; Shang, E.-X.; Hua, Y.-q.; Duan, J.-a.; Zhu, Y. Protective effects of Lycium barbarum L. berry extracts against oxidative stress-induced damage of the retina of aging mouse and ARPE-19 cells. Food Funct. 2023, 14, 399–412. [Google Scholar] [CrossRef]
- Mi, X.S.; Feng, Q.; Lo, A.C.; Chang, R.C.; Lin, B.; Chung, S.K.; So, K.F. Protection of retinal ganglion cells and retinal vasculature by Lycium barbarum polysaccharides in a mouse model of acute ocular hypertension. PLoS ONE 2012, 7, e45469. [Google Scholar] [CrossRef]
- Lakshmanan, Y.; Wong, F.S.-Y.; Yu, W.-Y.; Li, S.Z.-C.; Choi, K.-Y.; So, K.-F.; Chan, H.H.-L. Lycium brbarum polysaccharides rescue neurodegeneration in an acute ocular hypertension rat model under pre- and posttreatment conditions. Investig. Ophthalmol. Vis. Sci. 2019, 60, 2023–2033. [Google Scholar] [CrossRef]
- Mi, X.S.; Chiu, K.; Van, G.; Leung, J.W.; Lo, A.C.; Chung, S.K.; Chang, R.C.; So, K.F. Effect of Lycium barbarum Polysaccharides on the expression of endothelin-1 and its receptors in an ocular hypertension model of rat glaucoma. Neural. Regen. Res. 2012, 7, 645–651. [Google Scholar] [CrossRef]
- Mi, X.S.; Feng, Q.; Lo, A.C.Y.; Chang, R.C.; Chung, S.K.; So, K.F. Lycium barbarum polysaccharides related RAGE and Aβ levels in the retina of mice with acute ocular hypertension and promote maintenance of blood retinal barrier. Neural. Regen. Res. 2020, 15, 2344–2352. [Google Scholar] [CrossRef]
- Yang, D.; So, K.-F.; Lo, A.C. Lycium barbarum polysaccharide extracts preserve retinal function and attenuate inner retinal neuronal damage in a mouse model of transient retinal ischaemia. Clin. Exp. Ophthalmol. 2017, 45, 717–729. [Google Scholar] [CrossRef]


| Model | Main Bioactives | Key Findings | Mechanism/Pathway | Refs. |
|---|---|---|---|---|
| In vitro | LBP | ↓ Myocardial damage, ↓ apoptosis, preserved mitochondrial function | SIRT3/CypD pathway | [77] |
| In vivo | LBP | ↓ Infarct size, prevented adverse cardiac remodelling, lowered oxidative stress, improved mitochondrial dynamics | GRK2 expression inhibition; restoration of mitochondrial fission/fusion balance; activation of AKT/eNOS signalling | [78] |
| In vitro | LBP | Enhanced antioxidant defences (↑ SOD, ↑ NO), ↓ oxidative damage (↓ MDA), anti-apoptotic effects | Oxidative stress regulation | [79] |
| In vivo | L. barbarum extract | Normalised blood pressure, ↑ eNOS, ↓ sONE expression | sONE/eNOS pathway modulation | [80] |
| Model | Main Bioactive(s) | Key Findings | Mechanism/Pathway | Refs. | |
|---|---|---|---|---|---|
| A | In vivo | LBP | ↓ Adipocyte lipid accumulation, ↑ insulin sensitivity | ATF6/SIRT1-dependent downregulation of Fsp27 | [83] |
| In vivo | LICP009-2-1 | ↓ Lipid accumulation and hyperlipidemia | Anti-adipogenic activity | [84] | |
| In vivo | LBP | ↓ Blood glucose, ↑ insulin sensitivity, ↑ testosterone | Anti-metabolic disturbance | [85] | |
| In vivo | LBLF | ↓ Oxidative stress, ↑ liver function Improved insulin resistance, optimised gut microbiota | MAPK and retinol metabolism pathways modulation | [86] | |
| In vitro In vivo | LBP | ↓ Obesity-related factors and ↑ muscle-related factors; ↑ glucose metabolism; mitigated ectopic fat and mitochondrial dysfunction | AMPK/PINK1/Parkin-mediated mitophagy; ↑ mitochondrial membrane potential and ATP, ↓ROS | [87] | |
| B | In vivo | Zeaxanthin | Improved blood glucose, lipid profile, nephroprotection | Modulation of inflammatory cytokines and antioxidant enzymes | [88] |
| In vivo | LBP | Improved gut motility, ↑ SCFA production | Neuronal regulation of duodenal contraction | [89] | |
| In vivo | LBP | ↓ Oxidative stress, protected DNA in lymphocytes | Antioxidant protection (↑ SOD, ↓ MDA/NO) | [90] | |
| In vivo | LBL | Restored organ function, improved lipid and glucose metabolism | Modulation of gut microbiota and metabolic disruption reversal | [91] | |
| C | In vivo | LBP | ↓ Cholesterol and triglycerides, promoted weight loss | Gut microbiota modulation (↑ Bacteroidetes, ↓ Firmicutes) | [92] |
| In vivo | LBP | Improved lipid metabolism, ↑ antioxidant capacity | Upregulation of lipid metabolism genes (FAS, PPAR-α, CPT1, ATGL) | [93] | |
| In vivo | LBP | Improved growth, lipid metabolism, antioxidant capacity | Downregulation of lipid metabolism genes (ACC1, PPAR-γ) | [94] | |
| In vivo | LBP | Improved lipid metabolism, alleviated metabolic disorder symptoms, ↓weight gain, ↑ microbial diversity | Modulation of gut microbiota (↑ Firmicutes, ↑ microbial diversity), regulation of >30 differential metabolites and 4 metabolic pathways | [95] | |
| D | In vitro | LBP | Mitigated osteoblast apoptosis | miR-200b-3p/Chrdl1/PPARγ pathway modulation | [96] |
| In vitro | LBP | Promoted osteoblast proliferation via SCFA production | Upregulation of B-ALP and osteocalcin expression | [97] |
| Model | Main Bioactive(s) | Key Findings | Mechanism/Pathway | Refs. |
|---|---|---|---|---|
| In vivo | Methanol extract | ↓ Paw swelling, ↓ ROS, neutrophil migration | Myeloperoxidase inhibition | [56] |
| In vivo | Fermented/non-fermented extracts | Improved hepatic function, lipid metabolism | Anti-inflammatory + antioxidant | [111] |
| In vivo | LBP | ↓ Fat accumulation, fibrosis, inflammation | NF-κB, MAPK, autophagy | [112] |
| In vivo | LBP | ↓ Albuminuria, ↓ inflammatory, ↑ SOD ↓ Cytokines, ↓NF-κB | Renal protection | [113] |
| In vivo | LBP | Anti-inflammatory, anti-apoptotic | Nrf2 pathway | [114] |
| In vivo | LBP | ↓ Oxidative injury, ↓ inflammation markers | TLR/NF-κB inhibition | [115] |
| In vivo | LBP | ↓ ROS, 1CAM-1, improved kidney function | NF-κB and angiotensin downregulation | [116] |
| Model | Main Bioactive(s) | Key Findings | Mechanism/Pathway | Refs. |
|---|---|---|---|---|
| In vivo | LBP | ↓ Tumour size, ↑ macrophage and lymphocyte activity | Immune activation, ↓ lipid peroxidation | [117] |
| In vivo | LBP | ↑ CD8+ T cell infiltration, ↓ T cell exhaustion | Enhanced systemic/local antitumour immunity | [118] |
| In vitro | LBP (>10 kDa) | ↑ Macrophage viability and NO, TNF-α, IL-6 | Cellular uptake via clathrin-mediated endocytosis | [119] |
| In vivo | LBP | ↑ Immune organ indices, IgG, CD4+/CD8+ ratio | Immune stimulation and improved feed efficiency | [24,120] |
| In vivo | LBP | ↑ Cytokines, improved sperm parameters | Immuno-protection in cyclophosphamide model | [121] |
| In vivo | LBP | ↓ Hepatotoxicity, ↑ SCFAs, modulated gut microbiota | Gut–immune axis modulation | [122] |
| In vivo | Juice blends | ↑ Splenic macrophages, spleen weight | Synergistic antioxidant-immune enhancement | [28] |
| Model | Main Bioactive(s) | Key Findings | Mechanism/Pathway | Refs. |
|---|---|---|---|---|
| In vitro | Ethanol extract | ↑ NK cytotoxicity, ↓Proliferation | NK cell activation | [126] |
| In vitro | Ethanol extract | ↓ Proliferation, ↑ Apoptosis, ↑ Bax, ↓ BclxL | Mitochondrial pathway | [31] |
| In vitro | Polyphenol-rich extract | Inhibited proliferation in MCF-7 and MDA-MB-231 cancer cells | Dose-dependent inhibition | [127] |
| In vitro | LBP fractions | Induced G0/G1 arrest, apoptosis, ↓ mito. potential | ↑ Caspase, ↑ MAPK, ↓ Bcl-2 | [128] |
| In vitro | LBP | Induced S phase arrest and apoptosis, ↑ RNA/Ca2+ | Calcium-regulated apoptotic pathways | [129] |
| In vivo | LBP-3 (40–350 kDa) | Tumour suppression, S phase arrest immune support | Immune modulation | [130] |
| In vitro | Ethanol extract | ↓ Proliferation, ↓ migration | ERK1/2, AKT suppression | [131] |
| In vitro | LLB extract | No cytotoxicity or genotoxicity; ↓ DNA damage; ↓ pro-metastatic and ↑tumour suppressor genes | Modulation of oxidative stress, apoptosis, and cancer-related gene expression | [69] |
| In vitro | LBP | Combined LBP and cisplatin inhibited cell proliferation; ↑ apoptosis, ↓ ROS; modulated S/G2-M cell cycle phases | Enhanced apoptosis and cell cycle arrest via cyclin D1-CDK4-Rb pathway | [132] |
| In vitro In vivo | LBP | Combined LBP and IFN-α2b inhibited proliferation, induced apoptosis, ↓ tumour volume, and ↓ MDSC ratio | Synergistic regulation of apoptosis and immune suppression via cyclin D1/c-Myc/Bcl-2 pathway and MDSC modulation | [133] |
| In vitro In vivo | LBP | Reduced cardiac damage (↓ myofibrillar disarrangement), improved conduction abnormalities, and preserved anti-tumour activity of Doxorubicin | Suppression of oxidative stress (↑ SOD, ↑ GSH-Px, ↓ MDA) | [134] |
| Model | Main Bioactive(s) | Key Findings | Mechanism/Pathway | Refs. | |
|---|---|---|---|---|---|
| A | In vitro | LBP | ↓ Oxidative stress, ↓ apoptosis, ↓ inflammation | TXNIP-NLRP3 | [137] |
| In vivo | L. barbarum extract | ↓ ALT, ↓AST | Antioxidant enhancement | [138] | |
| In vivo | LBP | ↓ Fatty liver, ↑ liver enzymes, ↑ antioxidant activity | Hepatic protection and redox modulation | [139] | |
| B | In vitro | Yellow L. barbarum extract | Strong activity against Gram– and Candida | Dose-dependent | [14] |
| In vitro | Hydromethanolic extract | ↑ Antibacterial activity (Gram+ > Gram–) | Disruption of cell membrane integrity | [15] | |
| In vitro | Polyphenols (e.g., chlorogenic acid) | ↓ E. coli growth | Time/dose/temperature-dependent | [29] | |
| In vitro | Ethanolic extract | Effective against periodontal pathogens | Alternative to chlorhexidine | [140] | |
| C | In vivo | LBP | ↑ Firmicutes/Proteobacteria, ↓ Bacteroidetes ↑ Lactobacillus/Akkermansia/Prevotellaceae | Microbiota modulation Gut microbiota enrichment | [141] |
| In vitro | LBP | ↑ L. acidophilus, ↑B. longum (2.5–15% LBP) | Prebiotic-enhanced bacterial growth | [141] | |
| In vivo | L. barbarum | ↓ ALT/AST ↑ Lachnospiraceae, ↑ Ruminococcaceae | Gut–liver axis modulation | [142] | |
| In vitro | Aqueous extract | ↑ Lactobacillus, Bifidobacterium growth | Stimulated probiotic proliferation | [143] |
| Model | Main Bioactive(s) | Key Findings | Mechanism/Pathway | Refs. |
|---|---|---|---|---|
| In vivo | L. barbarum extract | ↓ Oxidative stress and cytokines; ↑ hippocampal neuron survival | PI3K/Akt/GSK-3β, PKCε/Nrf2/HO-1, NR2A/NR2B | [146] |
| In vivo | LBP | ↑ Spatial memory, ↑ neurogenesis; ↓apoptosis and ER stress | Nrf2/HO-1 signalling | [147] |
| In vitro In vivo | LBP | ↓ ROS, ↓ mitochondrial damage; ↑ caspase-3/-9 activity | ↑ Nrf2 and HO-1 expression | [148] |
| In vivo In vitro | L. barbarum | Reversed cognitive impairment; ↓ apoptosis; ↑ antioxidant defence | Nrf2/HO-1 | [149] |
| In vitro In vivo | Water extract | ↑ Cell survival; ↓ROS; ↑ acetylcholine, choline acetyltransferase | Mitochondrial protection and neurotransmitter regulation | [150] |
| In vitro | Alkaline extract | ↓ Caspase-3 activity; ↑ Akt phosphorylation | Anti-apoptotic via Akt signalling | [30] |
| In vivo In vitro | L. barbarum extract | ↓ Dopaminergic neuron loss; ↓ROS; ↑ GSH | Mitochondrial stabilisation, antioxidative action | [151] |
| Model | Main Bioactive(s) | Key Findings | Mechanism/Pathway | Refs. |
|---|---|---|---|---|
| In vivo | ABP and LBP | ↓ glycation end-products, ↑ IL-2, ↑ SOD, ↑ cognitive and motor function | Glycation inhibition; immune and oxidative modulation | [153] |
| In vivo | LBP, LBP-2 (arabinogalactan) | ↑ Lifespan, ↑ SOD/CAT, ↓ MDA | MAPK/TOR/S6K pathway; longevity gene upregulation | [154] |
| In vitro | LBP | ↓ UVB-induced DNA damage and ROS, ↑ Nrf2 activation, ↓ p38 MAPK | Nrf2/ARE; caspase-3, MMP-9 | [155] |
| In vivo In vitro | LBP1C-2 | ↑ Muscle stem cell self-renewal and repair | FGFR1 binding; Spry1 upregulation | [156] |
| In vivo In vitro | LBP | ↑ Osteogenic markers; ↓bone resorption and osteoclasts | ERK1/2 pathway activation | [157] |
| In vivo | LBP | ↓ Senescence, ↓ apoptosis | p53 signalling modulation | [158] |
| In vitro | LBP | ↓ Lipid peroxidation, delayed apoptosis | Oxidative stress reduction | [159] |
| In vitro | LBP | ↓ Oxidative stress, ↓ apoptosis and autophagy | PI3K/Akt/mTOR, Bcl-2/Bax, Caspase-3 | [160] |
| In vivo | LBP | ↑ SIRT3, ↓ CypD acetylation, ↑ mitochondrial protection | SIRT3/CypD pathway activation | [77] |
| Model | Main Bioactive(s) | Key Findings | Mechanism/Pathway | Refs. |
|---|---|---|---|---|
| In vivo | LBP | ↑ Nrf2 and HO-1; ↓ apoptosis; ↑ survival of ganglion cells | Nrf2/HO-1 antioxidant pathway | [162] |
| In vivo | LBP | ↓ oxidative stress; ↓ JNK; ↑ IGF-1; ↓ secondary RGC degeneration | JNK pathway, oxidative stress inhibition | [163] |
| In vivo | LBP | ↑ retinal function and visual signalling | Retinal functional recovery | [164] |
| In vivo | LBP | ↑ M2 microglia/macrophage polarisation; ↓ autophagy; ↑ RGC survival | Immune modulation | [165] |
| In vivo | LBP | Preserved photoreceptor morphology and visual behaviour | NF-κB and HIF-1α inhibition | [166] |
| In vivo | LBW-95E | ↑SOD, ↑GSH, ↑Nrf2; ↓ ROS, inflammation | Antioxidant and anti-inflammatory | [167] |
| In vivo | LBP | ↓ RGC loss; preserved blood-retinal barrier | Downregulation of inflammatory mediators | [168] |
| In vivo | LBP | Preserved inner retinal layer thickness; ↑retinal function | Protection from secondary degeneration | [169] |
| In vivo | LBP | ↓ ET-1 expression; ↑ ETA, ↓ ETB in RGCs | ET-1 signalling modulation | [170] |
| In vivo | LBP | ↓ astrocyte/microglia activation; preserved barrier integrity | Glial reactivity modulation | [171] |
| In vivo | LBP | ↑ viable retinal cells, ERG, ↓ glial activity | Neuroprotection | [172] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, T.; Alexa, E.-A.; Liu, G.; Berisha, A.; Walsh, R.; Kelleher, R. Lycium barbarum for Health and Longevity: A Review of Its Biological Significance. Obesities 2025, 5, 35. https://doi.org/10.3390/obesities5020035
Zhang T, Alexa E-A, Liu G, Berisha A, Walsh R, Kelleher R. Lycium barbarum for Health and Longevity: A Review of Its Biological Significance. Obesities. 2025; 5(2):35. https://doi.org/10.3390/obesities5020035
Chicago/Turabian StyleZhang, Tao, Elena-Alexandra Alexa, Gavin Liu, Alois Berisha, Rhys Walsh, and Robbie Kelleher. 2025. "Lycium barbarum for Health and Longevity: A Review of Its Biological Significance" Obesities 5, no. 2: 35. https://doi.org/10.3390/obesities5020035
APA StyleZhang, T., Alexa, E.-A., Liu, G., Berisha, A., Walsh, R., & Kelleher, R. (2025). Lycium barbarum for Health and Longevity: A Review of Its Biological Significance. Obesities, 5(2), 35. https://doi.org/10.3390/obesities5020035