
Benthic Macrofauna in the Loukkos Estuary, Morocco: Patterns and Environmental Drivers


Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Sampling of Benthic Macrofauna
2.3. Environmental Data
2.4. Statistical Analyses
3. Results
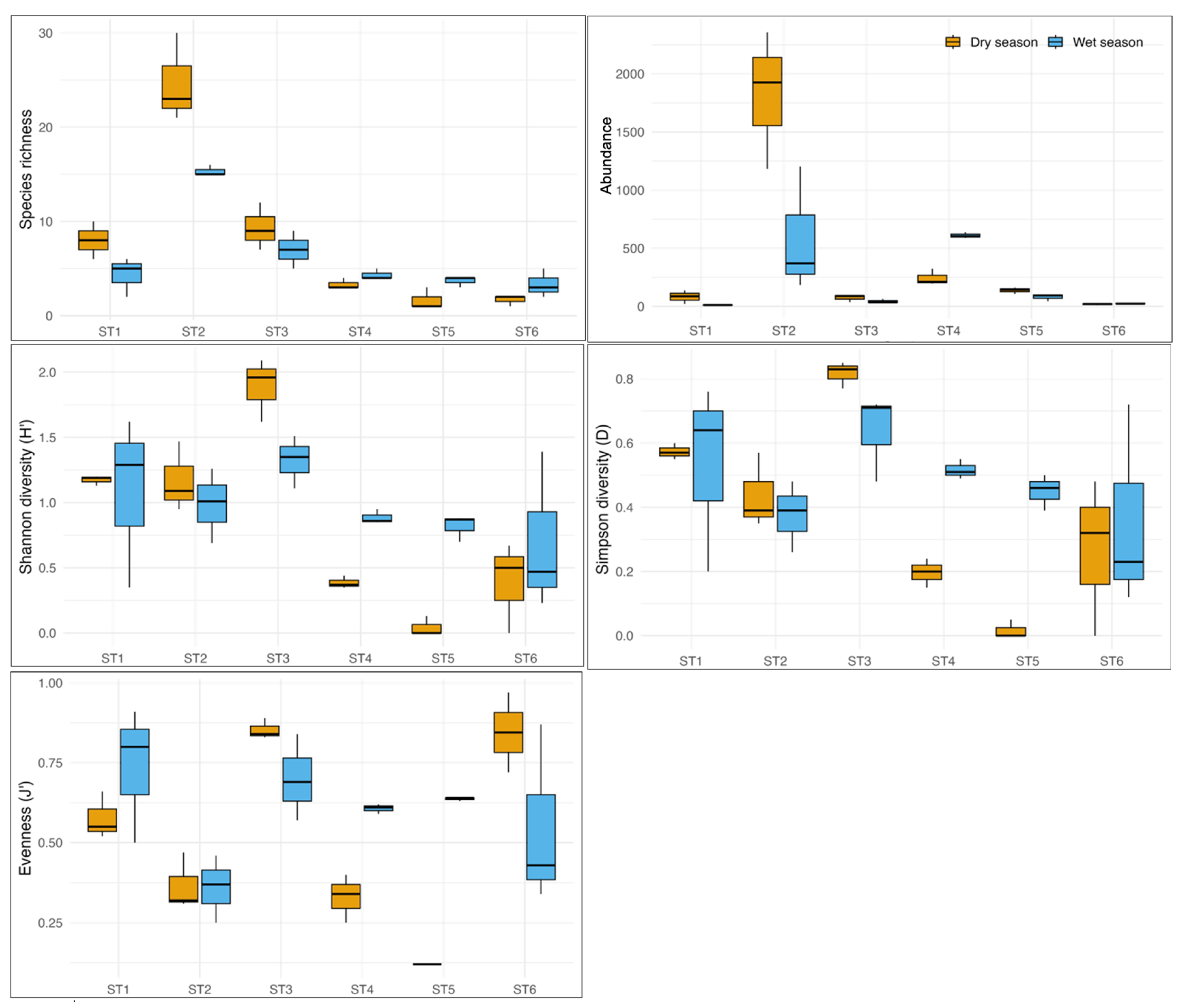
3.1. Structure and Diversity Patterns of Benthic Communities
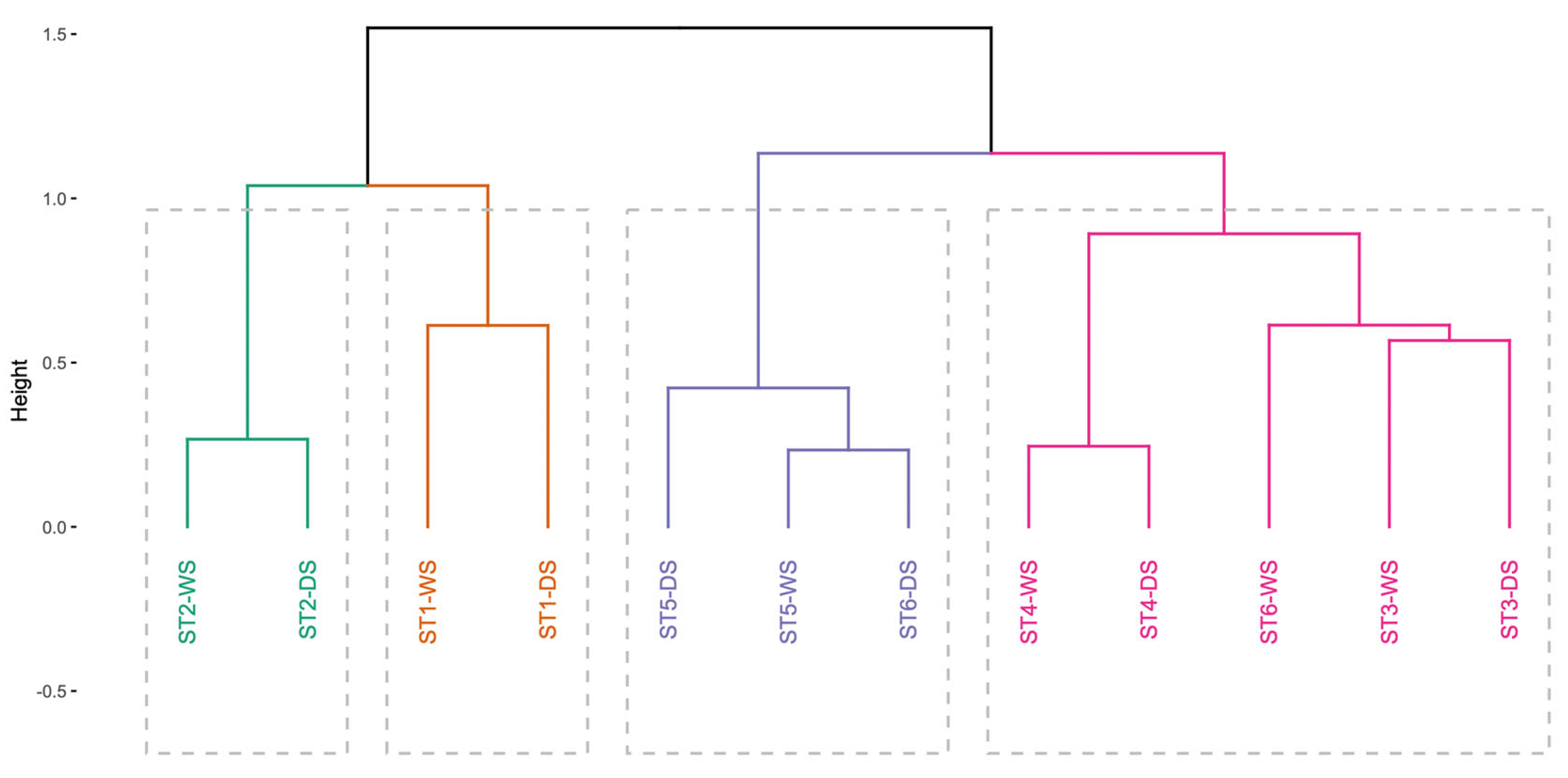
3.2. Spatio-Temporal Organization of Benthic Communities
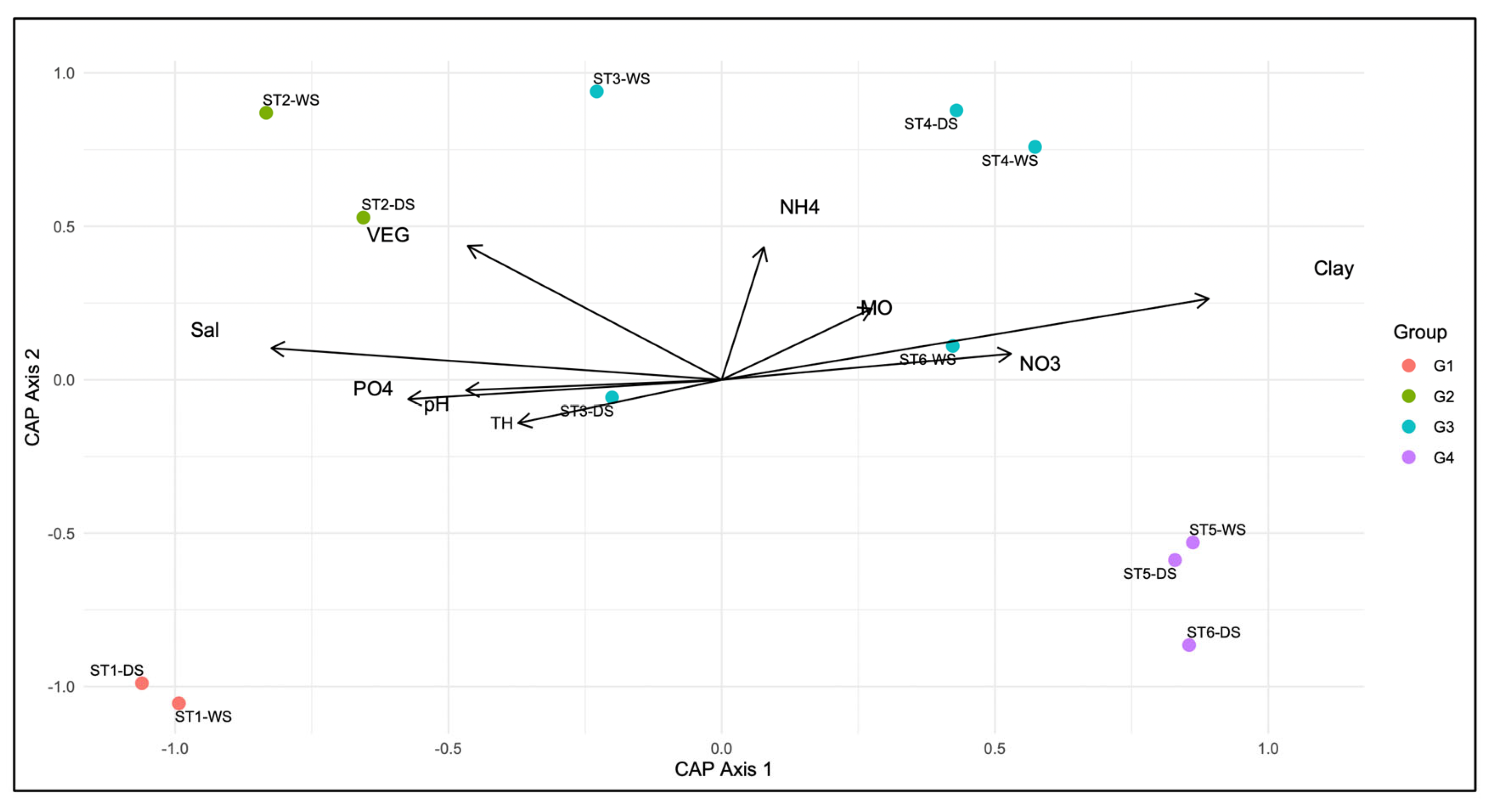
3.3. Environmental Drivers of Benthic Community Structure
4. Discussion
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zwarts, L.; Wanink, J.H. How the food supply harvestable by waders in the Wadden Sea depends on the variation in energy density, body weight, biomass, burying depth and behaviour of tidal-flat invertebrates. Neth. J. Sea Res. 1993, 31, 441–476. [Google Scholar] [CrossRef]
- Touhami, F.; Bazairi, H.; Badaoui, B.; Benhoussa, A. Vertical Distribution of Benthic Macrofauna in Intertidal Habitats Frequented by Shorebirds at Merja Zerga Lagoon. Thalassas 2017, 34, 255–265. [Google Scholar] [CrossRef]
- Touhami, F.; Idrissi, H.R.; Benhoussa, A. Foraging behaviour of wintering shorebirds at Merja Zerga lagoon (Atlantic coast, Morocco). Ostrich 2020, 91, 244–251. [Google Scholar] [CrossRef]
- Chakraborty, A.; Saha, G.; Aditya, G. Macroinvertebrates as engineers for bioturbation in freshwater ecosystem. Environ. Sci. Pollut. Res. 2022, 29, 64447–64468. [Google Scholar] [CrossRef]
- Souza, F.; Gilbert, E.; Brauko, K.; Lorenzi, L.; Machado, E.; Camargo, M. Macrobenthic community responses to multiple environmental stressors in a subtropical estuary. PeerJ 2021, 9, e12427. [Google Scholar] [CrossRef]
- Adesakin, T.A.; Erhomosele, E.I.; Ogunrinola, O.; Oloyede, O.O.; Adedeji, A.A.; Odufuwa, P.T.; Aimienoho, A.; Aduwo, A.I.; Adewumi, E.A. Using benthic macroinvertebrates as bioindicators to evaluate the impact of anthropogenic stressors on water quality and sediment properties of a West African lagoon. Heliyon 2023, 9, e19508. [Google Scholar] [CrossRef]
- Borja, Á.; Franco, J.; Pérez, V. A Marine Biotic Index to Establish the Ecological Quality of Soft-Bottom Benthos Within European Estuarine and Coastal Environments. Mar. Pollut. Bull. 2000, 40, 1100–1114. [Google Scholar] [CrossRef]
- Gao, X.; Li, W.; Zhang, Y.; Song, H.; Li, Y.; Li, H. Integrated Assessment of Ecological Quality Combining Biological and Environmental Data in the Yellow River Estuary. Water 2024, 16, 1615. [Google Scholar] [CrossRef]
- El Morhit, M. Hydrochimie, Éléments Traces Métalliques et Incidences Écotoxicologiques sur les Différentes Composantes d’un Écosystème Estuarien (Bas Loukkos). Ph.D. Thesis, Mohammed V University, Rabat, Morocco, 2009. [Google Scholar]
- Touhami, F.; El Qoraychy, I.; Hammani, O. Assessment of spatial and seasonal variations in water quality in Loukkos Estuary (Atlantic Coast of Morocco). Desalin. Water Treat. 2025, 322, 101240. [Google Scholar] [CrossRef]
- RSIS. Fiche Descriptive Ramsar–Complexe du bas Loukkos (MAROC). Available online: https://rsis.ramsar.org/RISapp/files/RISrep/MA1475RIS_2501_fr.pdf (accessed on 15 April 2025).
- Qninba, H.; Himmi, O.; Benhoussa, A.; El Agbani, M.A.; Thévenot, M. Nouveaux cas de nidification d’oiseaux dans le complexe de zones humides du Bas Loukkos (Nord-Ouest du Maroc). Bull. Inst. Sci. Rabat 2008, 30, 45–50. [Google Scholar]
- El Morhit, M.; Fekhaoui, M.; El Abidi, A.; Yahyaoui, A.; Hamdani, A. Impact des activités humaines sur la dégradation de la qualité des sédiments de l’estuaire de Loukkos (Maroc). Environ. Ing. Dév. 2012, 61, 8–17. [Google Scholar] [CrossRef]
- Carmona, P.; Ruiz, J.M. Geomorphological evolution of the river Loukkos estuary around the Phoenician city of Lixus on the Atlantic Littoral of Morocco. Geoarchaeology 2009, 24, 821–845. [Google Scholar] [CrossRef]
- Hayward, P.J.; Ryland, J.S. Handbook of the Marine Fauna of North-West Europe; Oxford University Press: Oxford, UK, 1995. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2024; Available online: https://www.R-project.org/ (accessed on 19 March 2025).
- Cherkaoui, E.; Bayed, A.; Hily, C. Organisation spatiale des peuplements macrozoobenthiques subtidaux d’un estuaire de la côte atlantique marocaine: L’estuaire du Bou Regreg. Cah. Biol. Mar. 2003, 44, 339–352. [Google Scholar] [CrossRef]
- Chaouti, A.; Azirar, A.; Zourarah, B.; Bayed, A. Patterns of Spatial Subtidal Macrobenthic Community Distribution in Relation to Abiotic Conditions in a North African Estuarine Ecosystem (Northwestern Morocco). Ann. Zool. Fennici 2016, 53, 125–138. [Google Scholar] [CrossRef]
- Bergayou, H.; Annajar, E.M.; Lefrere, L.; Moukrim, A.; Gillet, P. Recovery of an Estuarine Ecosystem after the Stopping of Wastewater Discharge: Intertidal Macrobenthic Community Characterization in the Estuary of Oued Souss (Southwestern Morocco). J. Ecol. Eng. 2019, 20, 133–145. [Google Scholar] [CrossRef]
- Touhami, F.; Bazairi, H.; Badaoui, B.; Morabbi, A.; Benhoussa, A. Structure and spatial organization of macrobenthic fauna of intertidal habitats frequented by wintering shorebirds at Merja Zerga lagoon (Moroccan Ramsar Site). Cah. Biol. Mar. 2019, 60, 41–50. [Google Scholar] [CrossRef]
- El Asri, F.; Errhif, A.; Tamsouri, M.; Martín, D.; Maanan, M.; Zidane, H. Analysis of the structural characteristics and spatial organization of macrobenthic fauna in Oualidia lagoon, Morocco. OpenAlex 2022, 12, 96. [Google Scholar] [CrossRef]
- Warwick, R.; Clarke, K. Comparing the severity of disturbance: A metaanalysis of marine macrobenthic community data. Mar. Ecol. Prog. Ser. 1993, 92, 221–232. [Google Scholar] [CrossRef]
- Ysebaert, T.; Herman, P. Spatial and temporal variation in benthic macrofauna and relationships with environmental variables in an estuarine, intertidal soft-sediment environment. Mar. Ecol. Prog. Ser. 2002, 244, 105–124. [Google Scholar] [CrossRef]
- Hinchey, E.; Schaffner, L.; Hoar, C.; Vogt, B.; Batte, L. Responses of Estuarine Benthic Invertebrates to Sediment Burial: The Importance of Mobility and Adaptation. Hydrobiologia 2006, 556, 85–98. [Google Scholar] [CrossRef]
- Little, S.; Wood, P.J.; Elliott, M. Quantifying salinity-induced changes on estuarine benthic fauna: The potential implications of climate change. Estuar. Coast. Shelf Sci. 2017, 198, 610–625. [Google Scholar] [CrossRef]
- Vafeiadou, A.; Materatski, P.; Adão, H.; De Troch, M.; Moens, T. Food sources of macrobenthos in an estuarine seagrass habitat (Zostera noltii) as revealed by dual stable isotope signatures. Mar. Biol. 2013, 160, 2517–2523. [Google Scholar] [CrossRef]
- Boyé, A.; Legendre, P.; Grall, J.; Gauthier, O. Constancy despite variability: Local and regional macrofaunal diversity in intertidal seagrass beds. J. Sea Res. 2017, 130, 107–122. [Google Scholar] [CrossRef]
- Lundquist, C.; Jones, T.; Parkes, S.; Bulmer, R. Changes in benthic community structure and sediment characteristics after natural recolonisation of the seagrass Zostera muelleri. Sci. Rep. 2018, 8, 31398. [Google Scholar] [CrossRef]
- Pennington, J.T.; Chavez, F. Seasonal fluctuations of temperature, salinity, nitrate, chlorophyll and primary production at station H3/M1 over 1989–1996 in Monterey Bay, California. Deep Sea Res. Part II Top. Stud. Oceanogr. 2000, 47, 947–973. [Google Scholar] [CrossRef]
- Almeida, M.A.; Cunha, M.A.; Alcântara, F. Seasonal change in the proportion of bacterial and phytoplankton production along a salinity gradient in a shallow estuary. In Nutrients and Eutrophication in Estuaries and Coastal Waters; Springer: Dordrecht, The Netherlands, 2002; pp. 251–262. [Google Scholar] [CrossRef]
- Akin, S.; Winemiller, K.O. Seasonal variation in food web composition and structure in a temperate tidal estuary. Estuaries Coasts 2006, 29, 552–567. [Google Scholar] [CrossRef]
- Ge, J.; Chen, J.; Zi, F.; Song, T.; Hu, L.; He, Z.; Wu, L.; Ding, Y.; Li, H. Seasonal Variations in Macrobenthos Communities and Their Relationship with Environmental Factors in the Alpine Yuqu River. Biology 2025, 14, 120. [Google Scholar] [CrossRef]
- Dauvin, J.-C.; Gentil, F. Long-term changes in populations of subtidal bivalves (Abra alba and A. prismatica) from the Bay of Morlaix (Western English Channel). Mar. Biol. 1989, 103, 63–73. [Google Scholar] [CrossRef]
- Haubois, A.-C.; Guarini, J.-M.; Richard, P.; Blanchard, G.F.; Sauriau, P.-G. Spatio–temporal differentiation in the population structure of Hydrobia ulvae on an intertidal mudflat (Marennes-Oléron Bay, France). J. Mar. Biol. Assoc. UK 2002, 82, 605–614. [Google Scholar] [CrossRef]
- Pearson, T.H.; Rosenberg, R. Macrobenthic Succession in Relation to Organic Enrichment and Pollution of the Marine Environment. Oceanogr. Mar. Biol. Annu. Rev. 1978, 16, 229–311. [Google Scholar]
- Dauer, D.M. Biological Criteria, Environmental Health and Estuarine Macrobenthic Community Structure. Mar. Pollut. Bull. 1993, 26, 249–257. [Google Scholar] [CrossRef]
- Bazaïri, H.; Bayed, A.; Glémarec, M.; Hily, C. Spatial Organisation of Macrozoobenthic Communities in Response to Environmental Factors in a Coastal Lagoon of the NW African Coast (Merja Zerga, Morocco). Oceanol. Acta 2003, 26, 457–471. [Google Scholar] [CrossRef]
- Touhami, F.; Bazairi, H.; Badaoui, B.; Bouarour, B.; Benhoussa, A. Merja Zerga Lagoon: Study of the Functional Structure and Bioassessment of the Ecological Quality of Benthic Communities. J. Mater. Environ. Sci. 2017, 8, 4591–4599. [Google Scholar] [CrossRef]
- Attrill, M.J.; Strong, J.A.; Rowden, A.A. Are macroinvertebrate communities influenced by seagrass structural complexity? Ecography 2000, 23, 114–121. [Google Scholar] [CrossRef]
- Boström, C.; Jackson, E.L.; Simenstad, C.A. Seagrass landscapes and their effects on associated fauna: A review. Estuar. Coast. Shelf Sci. 2006, 68, 383–403. [Google Scholar] [CrossRef]
- Fersi, A.; Pezy, J.-P.; Bakalem, A.; Neifar, L.; Dauvin, J.-C. Molluscs from Tidal Channels of the Gulf of Gabès (Tunisia): Quantitative Data and Comparison with Other Lagoons and Coastal Waters of the Mediterranean Sea. J. Mar. Sci. Eng. 2023, 11, 545. [Google Scholar] [CrossRef]
- Gray, J.S.; Wu, R.S.S.; Or, Y.Y. Effects of hypoxia and organic enrichment on the coastal marine environment. Mar. Ecol. Prog. Ser. 2002, 238, 249–279. [Google Scholar] [CrossRef]
- Giangrande, A.; Licciano, M.; Musco, L. Polychaetes as environmental indicators revisited. Mar. Pollut. Bull. 2005, 50, 1153–1162. [Google Scholar] [CrossRef]
- Pampanin, D.M.; Camus, L.; Gomiero, A.; Marangon, I.; Volpato, E.; Nasci, C. Susceptibility to oxidative stress of mussels (Mytilus galloprovincialis) in the Venice Lagoon (Italy). Mar. Pollut. Bull. 2005, 50, 1548–1557. [Google Scholar] [CrossRef]
- Carvalho, S.; Barata, M.; Pereira, F.; Gaspar, M.; Cancela da Fonseca, L.; Pousão-Ferreira, P. Distribution patterns of macrobenthic species in relation to organic enrichment within aquaculture earthen ponds. Mar. Pollut. Bull. 2006, 52, 1573–1584. [Google Scholar] [CrossRef]
- Vesal, S.E.; Auriemma, R.; Libralato, S.; Nasi, F.; De Negro, P. Impacts of organic enrichment on macrobenthic production, productivity, and transfer efficiency: What can we learn from a gradient of sewage effluents? Mar. Pollut. Bull. 2022, 182, 113972. [Google Scholar] [CrossRef]
- Diaz, R.J.; Rosenberg, R. Spreading dead zones and consequences for marine ecosystems. Science 2008, 321, 926–929. [Google Scholar] [CrossRef]
- Whitfield, A.K.; Elliott, M.; Basset, A.; Blaber, S.J.; West, R.J. Paradigms in estuarine ecology—A review of the Remane diagram with a suggested revised model for estuaries. Estuar. Coast. Shelf Sci. 2012, 97, 78–90. [Google Scholar] [CrossRef]
- Belarmino, E.; Cabral, H.; Garcia, A.M. Long-term trends in the functional structure of estuarine fish assemblages in a subtropical estuary and its relationships with local environmental variability, man-made changes, and climatic drivers. Mar. Environ. Res. 2024, 201, 106698. [Google Scholar] [CrossRef] [PubMed]
- Lam-Gordillo, O.; Mosley, L.; Simpson, S.; Welsh, D.; Dittmann, S. Loss of benthic macrofauna functional traits correlates with changes in sediment biogeochemistry along an extreme salinity gradient in the Coorong lagoon, Australia. Mar. Pollut. Bull. 2022, 174, 113202. [Google Scholar] [CrossRef] [PubMed]
- Pascal, L.; Chaillou, G.; Nozais, C.; Cool, J.; Bernatchez, P.; Letourneux, K.; Archambault, P. Benthos response to nutrient enrichment and functional consequences in coastal ecosystems. Mar. Environ. Res. 2022, 175, 105584. [Google Scholar] [CrossRef]
- Manokaran, S.; Joydas, T.V.; Khan, A. Physico-chemical factors regulating marine benthos structure and function. In Ecology and Biodiversity of Benthos; Elsevier: Amsterdam, The Netherlands, 2022; pp. 209–250. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Molluscs | Polychaetes |
| Bivalves | Capitella capitata (Fabricius, 1780) |
| Abra alba (W. Wood, 1802) | Chaetopterus sp. |
| Cerastoderma edule (Linnaeus, 1758) | Diopatra neapolitana (Delle Chiaje, 1841) |
| Donax trunculus (Linnaeus, 1758) | Glycera tridactyla (Schmarda, 1861) |
| Mactra stultorum (Linnaeus, 1758) | Hediste diversicolor (O.F. Müller, 1776) |
| Mytilus galloprovincialis (Lamarck, 1819) | Heteromastus filiformis (Claparède, 1864) |
| Ruditapes decussatus (Linnaeus, 1758) | Lagis koreni (Malmgren, 1866) |
| Scrobicularia plana (da Costa, 1778) | Nephtys hombergii (Lamarck, 1818) |
| Solen marginatus (Pulteney, 1799) | Perinereis cultrifera (Grube, 1840) |
| Venus verrucosa (Linnaeus, 1758) | Prionospio sp. |
| Spisula subtruncata (da Costa, 1778) | Scoloplos armiger (Müller, 1776) |
| Gastropods | Crustaceans |
| Aplysia sp. | Afruca tangeri (Eydoux, 1835) |
| Bittium reticulatum (da Costa, 1778) | Callinectes sapidus (Rathbun, 1896) |
| Bulla striata (Bruguière, 1792) | Carcinus maenas (Linnaeus, 1758) |
| Haminoea navicula (da Costa, 1778) | Cyathura carinata (Krøyer, 1847) |
| Littorina obtusata (Linnaeus, 1758) | Idotea chelipes (Pallas, 1766) |
| Neverita sp. | Gammarus sp. |
| Peringia ulvae (Pennant, 1777) | Melita palmata (Montagu, 1804) |
| Phorcus turbinatus (Born, 1778) | Palaemon elegans (Rathke, 1836) |
| Rissoa parva (da Costa, 1778) | Sphaeroma sp. |
| Steromphala umbilicalis (da Costa, 1778) | Upogebia pusilla (Petagna, 1792) |
| Tritia reticulata (Linnaeus, 1758) | Others |
| Tritia incrassata (Strøm, 1768) | Ophiura ophiura (Linnaeus, 1758) |
| Dolichopodidae larave | |
| Larvae Pisces | |
| Nemertea ind. |
| Group | Stations | SIMPER Analysis | IndVal Analysis | ||
|---|---|---|---|---|---|
| Dissimilarities Between Groups | Similarity Within Group | Dominant Species (Contribution > 5%) | Indicator Species with p-Value and Significance | ||
| G1 | ST1-WS ST1-DS | 100% dissimilarity with G4 >94% with G2 and G3 | 18.6% | C. edule (49.6%) | D. trunculus (p = 0.043 *) |
| S. marginatus (32.1%) | S. armiger (p = 0.043 *) | ||||
| N. hombergii (6.1%) | |||||
| G2 | ST2-WS ST2-DS | 88.45% dissimilarity with G3 >94% with G4 | 48% | B. reticulatum (77.2%) | A. alba (p = 0.025 *) |
| L. koreni (p = 0.025 *) | |||||
| R. parva (p = 0.025 *) | |||||
| B. reticulatum (p = 0.027 *) | |||||
| P. ulvae (p = 0.021 *) | |||||
| T. reticulata (p = 0.015 *) | |||||
| I. chelipes (p = 0.034 *) | |||||
| D. neapolitana (p = 0.031 *) | |||||
| R. decussatus (p = 0.024 *) | |||||
| H. navicula (p = 0.034 *) | |||||
| (Shared with G1) | |||||
| N. hombergii (0.004, **) | |||||
| S. marginatus (0.020 *) | |||||
| G3 | ST3-WS ST3-DS ST4-WS ST4-DS ST6-WS | 65.15% dissimilarity with G4 | 20.7% | S. plana (60.7%) | (Shared with G2) |
| H. diversicolor (22.8%) | C. carinata (p = 0.035 *) | ||||
| C. carinata (8.9%) | C. capitata (p = 0.040 *) | ||||
| G4 | ST5-WS ST5-DS ST6-DS | 100% dissimilarity with G1 >94% with G2 | 34.5% | H. diversicolor (85.9%) | No exclusive indicator species detected |
| D. larvae (9.9%) | |||||
| F | Pr(>F) | % Variance Explained | % Cumulative Variance | |
|---|---|---|---|---|
| Global Model test | 2.2254 | 0.042 | - | - |
| Variables | ||||
| Salinity (Sal) | 5.0110 | 0.004 | 20.85 | 20.85 |
| Vegetation (VEG) | 4.2268 | 0.005 | 17.59 | 38.44 |
| pH | 2.2298 | 0.074 | 9.28 | 47.72 |
| Total hardness (TH) | 1.8067 | 0.163 | 7.52 | 55.24 |
| Phosphate (PO42−) | 1.7600 | 0.170 | 7.32 | 62.56 |
| Nitrate (NO3−) | 1.5345 | 0.237 | 6.39 | 68.95 |
| Clay (%) | 1.4628 | 0.259 | 6.09 | 75.04 |
| Ammonium (NH4+) | 1.3796 | 0.276 | 5.74 | 80.78 |
| Organic matter (MO) | 0.6172 | 0.756 | 2.57 | 83.35 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Touhami, F. Benthic Macrofauna in the Loukkos Estuary, Morocco: Patterns and Environmental Drivers. Ecologies 2025, 6, 53. https://doi.org/10.3390/ecologies6030053
Touhami F. Benthic Macrofauna in the Loukkos Estuary, Morocco: Patterns and Environmental Drivers. Ecologies. 2025; 6(3):53. https://doi.org/10.3390/ecologies6030053
Chicago/Turabian StyleTouhami, Feirouz. 2025. "Benthic Macrofauna in the Loukkos Estuary, Morocco: Patterns and Environmental Drivers" Ecologies 6, no. 3: 53. https://doi.org/10.3390/ecologies6030053
APA StyleTouhami, F. (2025). Benthic Macrofauna in the Loukkos Estuary, Morocco: Patterns and Environmental Drivers. Ecologies, 6(3), 53. https://doi.org/10.3390/ecologies6030053