
Seed Source for Restoration: Little Bluestem (Schizachyrium scoparium (Michx.) Nash) and the Carolina Sandhills


Abstract
1. Introduction
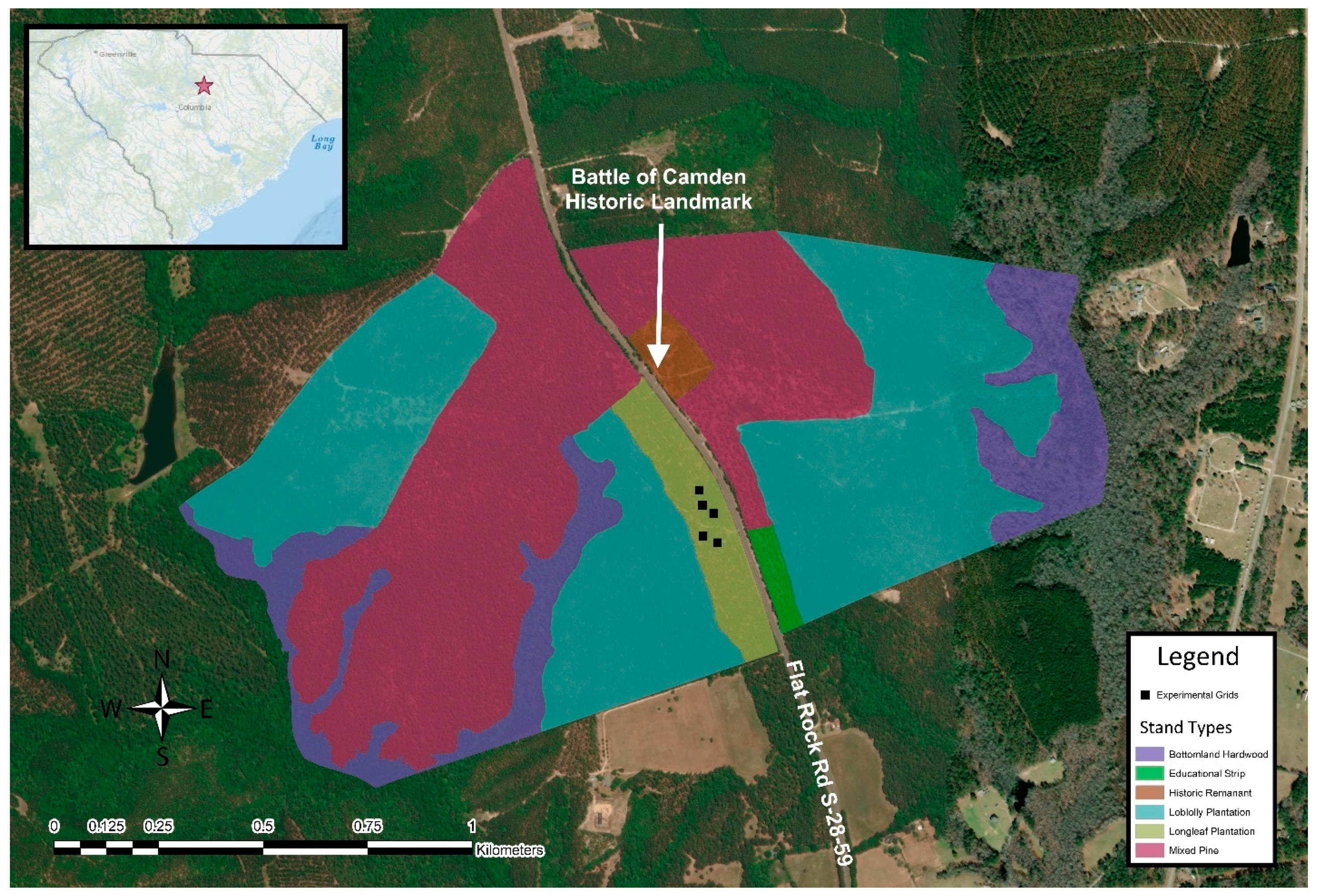
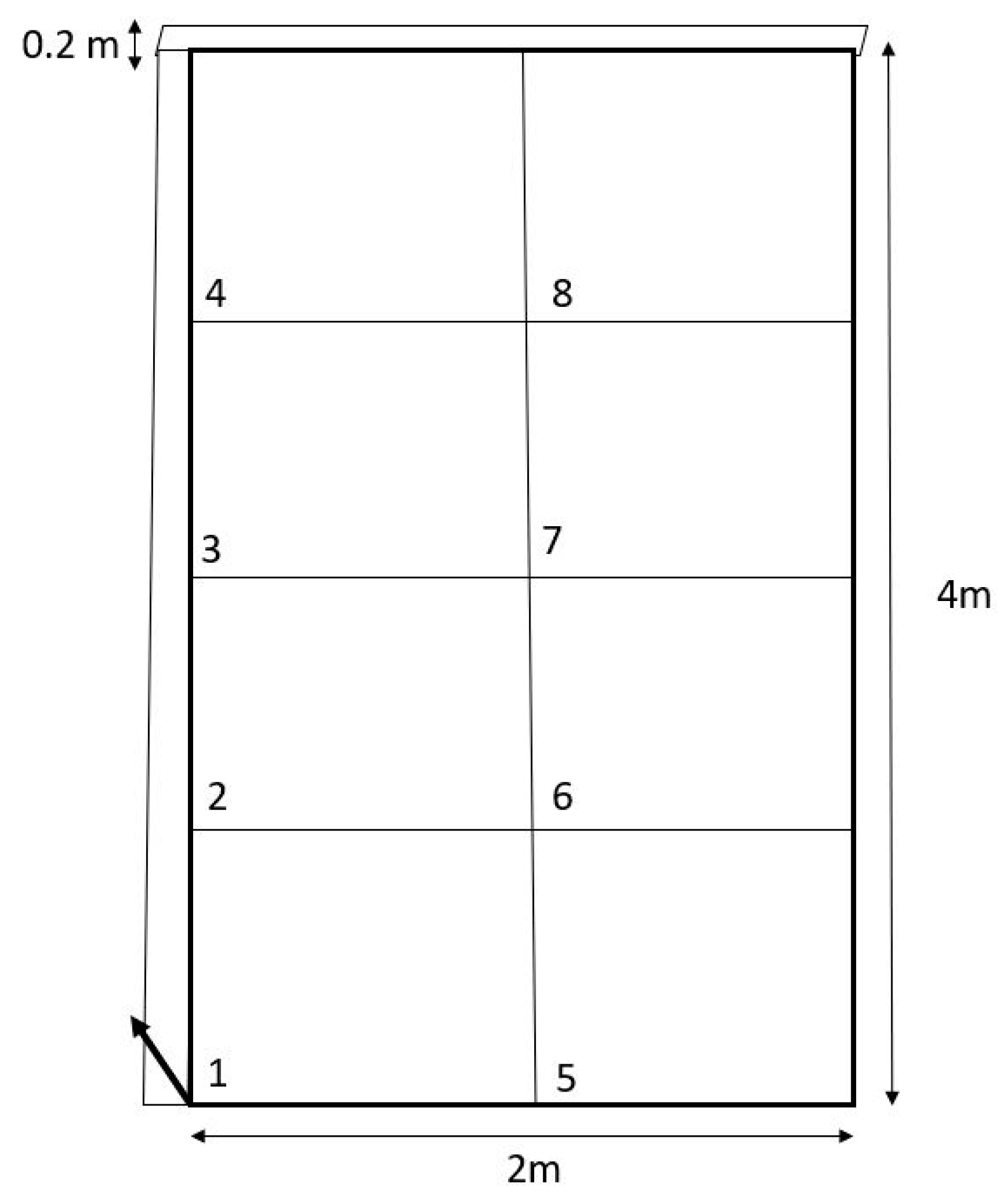
2. Materials and Methods
3. Results
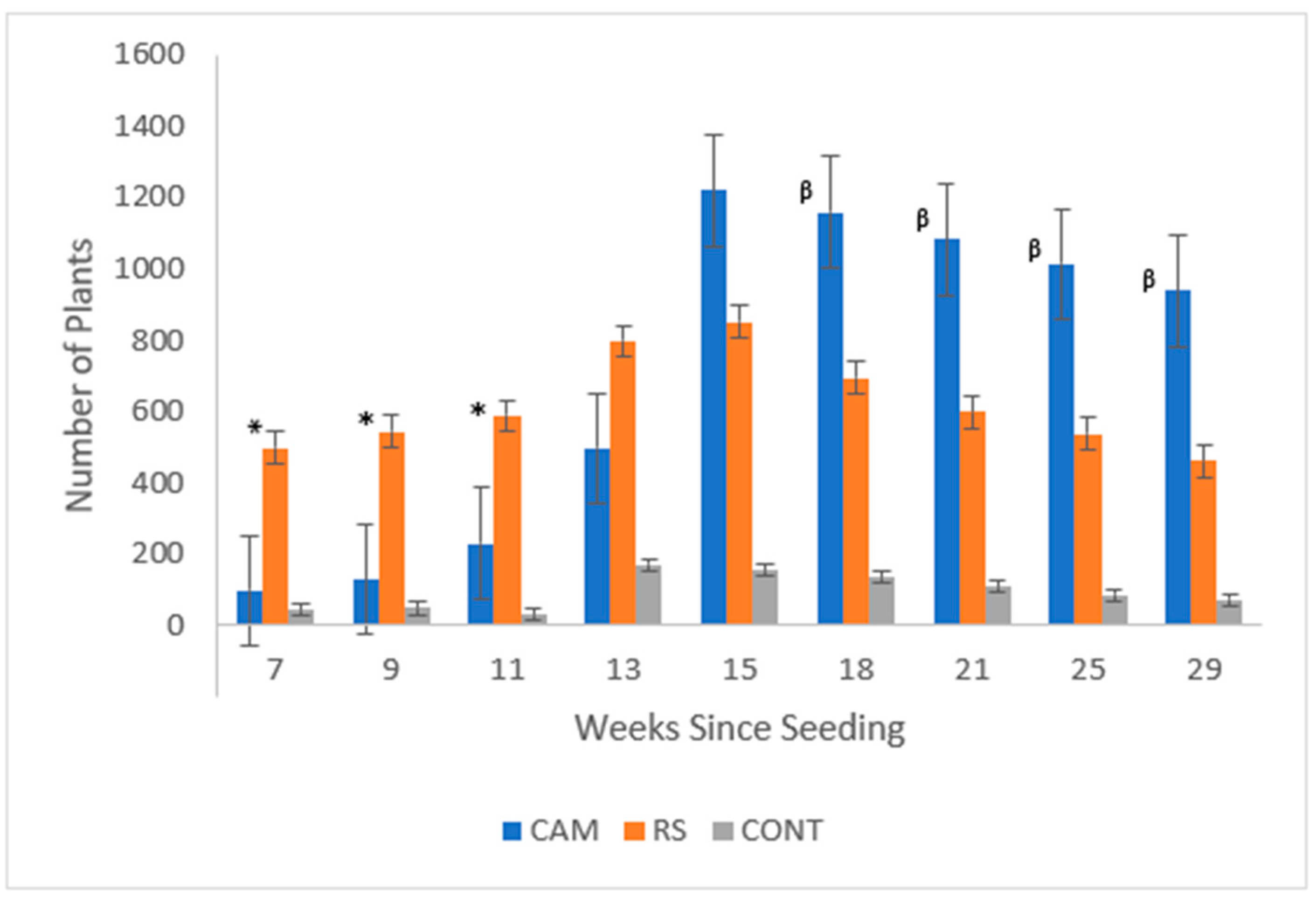
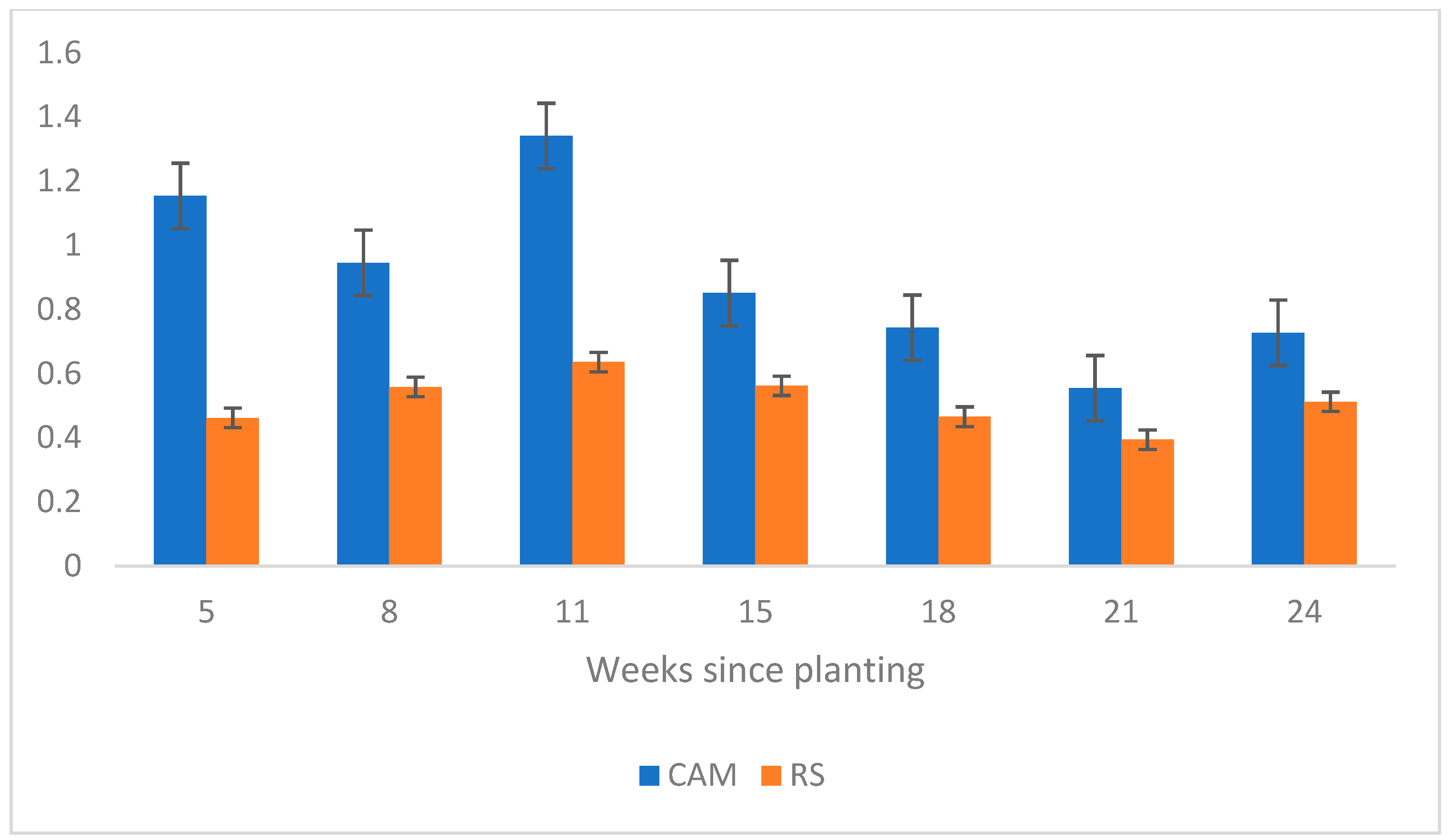
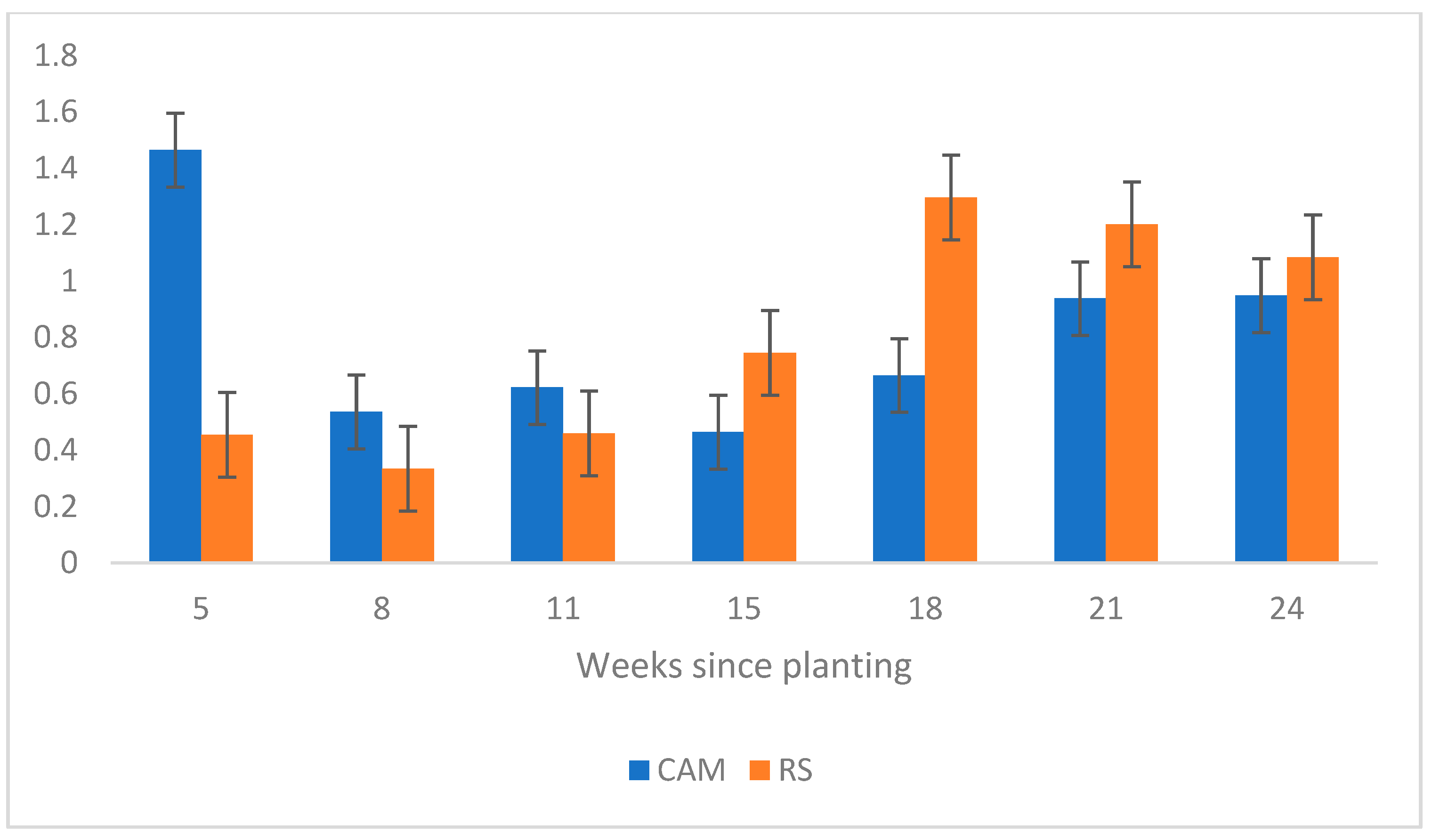
3.1. Question 1: Does Seed Source Matter?
3.2. Question 2: Is Site Preparation Treatment Necessary?
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Török, P.; Brudvig, L.A.; Kollmann, J.; Price, J.N.; Tóthmérész, B. The present and future of grassland restoration. Restor. Ecol. 2021, 29, e13378. [Google Scholar] [CrossRef]
- Camill, P.; McKone, M.J.; Sturges, S.T.; Severud, W.J.; Ellis, E.; Limmer, J.; Martin, C.B.; Navratil, R.T.; Purdie, A.J.; Sandel, B.S.; et al. Community and ecosystem-level changes in a species rich tallgrass prairie restoration. Ecol. Appl. 2004, 14, 1680–1694. [Google Scholar] [CrossRef]
- Kline, V.M.; Howell, E.A. Prairies. In Restoration Ecology: A Synthetic Approach to Ecological Research; Jordan, W.R., Gilpin, M.E., Aber, J.D., Eds.; Cambridge University Press: Cambridge, UK, 1999; pp. 75–84. [Google Scholar]
- Miller, C. The evolving understanding of grassland restoration seeding protocols. Ecol. Restor. 2013, 31, 127–129. [Google Scholar] [CrossRef]
- Packard, S.E.; Mutel, C.F. The Tallgrass Restoration Handbook: For Prairies, Savannahs, and Woodlands; Island Press: Washington, DC, USA, 1997. [Google Scholar]
- Rowe, H.I. Tricks of the trade: Techniques and opinions from 38 experts in tallgrass prairie restoration. Restor. Ecol. 2010, 18, 253–262. [Google Scholar] [CrossRef]
- Keyser, P.D.; Buehler, D.A.; Hedges, K.; Hodges, J.; Lituma, C.M.; Loncarich, F.; Martin, J.A. Eastern gasslands: Conservation challenges and opportunities on private lands. Wildl. Soc. Bull. 2019, 43, 382–390. [Google Scholar] [CrossRef]
- Hereford, J. A quantitative survey of local adaptation and fitness trade-offs. Am. Nat. 2009, 173, 579–588. [Google Scholar] [CrossRef]
- Leimu, R.; Fischer, M. A meta-analysis of local adaptation in plants. PLoS ONE 2008, 3, e4010. [Google Scholar] [CrossRef]
- Lowry, D. Ecotypes and the controversy over stages in the formation of new species. Biol. J. Linn. Soc. 2012, 106, 241–257. [Google Scholar] [CrossRef]
- Fedewa, C.A.; Stewart, J.R. Field establishment of little bluestem and prairie dropseed under managed conditions. Nativ. Plants J. 2011, 12, 111–117. [Google Scholar] [CrossRef]
- Radford, A.E.; Ahles, H.E.; Bell, C.R. Manual of the Vascular Flora of the Carolinas; University of North Carolina Press: Chapel Hill, NC, USA, 1968. [Google Scholar]
- Sorrie, B.A. A Field Guide to Wildflowers of the Sandhills Region; The University of North Carolina Press: Chapel Hill, NC, USA, 2011. [Google Scholar]
- Tober, D.; Jensen, N. Little Bluestem Schizachyrium Scoparium (Michx.); USDA Natural Resources Conservation Service: Washington, DC, USA, 2013.
- Huff, D.R.; Quinn, J.A.; Higgins, B.; Palazzo, A.J. Random amplified polymorphic DNA (RAPD) variation among native little bluestem [Schizachyrium scoparium (Michx.) Nash] populations from sites of high and low fertility in forest and grassland biomes. Mol. Ecol. 1998, 7, 1591–1597. [Google Scholar] [CrossRef]
- Bragg, L.H.; McMillan, C. Ecotypic Differentiation within Four North American Prairie Grasses. III. Chromatographic Variation. Am. J. Bot. 1966, 53, 893–901. [Google Scholar] [CrossRef]
- Gustafson, D.J.; Harris-Shultz, K.; Gustafson, P.E.; Giencke, L.M.; Denhof, R.C.; Kirkman, L.K. Seed sourcing for longleaf pine herbaceous understory restoration: Little bluestem (Schizachyrium scoparium) and hairy lespedeza (Lespedeza hirta) restoration genetics. Nat. Areas J. 2018, 38, 380–392. [Google Scholar] [CrossRef]
- Fu, Y.; Phan, A.T.; Coulman, B.; Richards, K.W. Genetic diversity in natural populations and corresponding seed collections of little bluestem as revealed by AFLP markers. Crop Sci. 2004, 44, 2254–2260. [Google Scholar] [CrossRef]
- Brockway, D.; Lewis, C. Long-term effects of dormant season prescribed fire on plant community diversity, structure, and productivity in a longleaf pine wiregrass ecosystem. For. Ecol. Manag. 1997, 96, 167–183. [Google Scholar] [CrossRef]
- Frost, C.C. Four centuries of changing landscape patterns in the longleaf pine ecosystem. In Proceedings of the Tall Timbers Fire Ecology Conference 1993, Tallahassee, FL, USA, 3–6 November 1993; Volume 18, pp. 17–43. [Google Scholar]
- Landers, L.; Van Lear, D.; Boyer, W. The longleaf pine forests of the Southeast: Requiem or renaissance. J. For. 1995, 93, 39–44. [Google Scholar]
- Peet, R.K. Ecological classification of the longleaf pine woodlands. In The Longleaf Pine Ecosystem: Ecology, Silviculture, and Restoration; Jose, S., Jokela, E.J., Miller, D.L., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; pp. 51–94. [Google Scholar]
- Sparks, J.; Masters, R.; Engle, D.; Palmer, M.; Bukenhofer, G. Effects of late growing-season and late-dormant season prescribed fire on herbaceous vegetation in restored pine-grassland communities. J. Veg. Sci. 1998, 9, 133–142. [Google Scholar] [CrossRef]
- Van Lear, D.; Carroll, W.; Kapeluck, P.; Johnson, R. History and restoration of the longleaf pine-grassland ecosystem: Implications for species at risk. For. Ecol. Manag. 2005, 211, 150–165. [Google Scholar] [CrossRef]
- Kush, J.S.; Meldahl, R.S.; Boyer, W.D. Understory plant community response to season of burn in natural longleaf pine forests. In Fire and Forest Ecology: Innovative Silviculture and Vegetation Management; Moser, W.K., Moser, C.F., Eds.; Tall Timbers Research Station: Tallahassee, FL, USA, 2000; pp. 32–39. [Google Scholar]
- Walker, J. Ground layer vegetation in longleaf pine landscapes: An overview for restoration and management. In Proceedings of the Longleaf Pine Ecosystem Restoration Symposium 1998, Charleston, SC, USA, 17–19 November 1998; Volume 9, pp. 2–13. [Google Scholar]
- Peet, R. A taxonomic study of Aristida stricta and A. beyrichiana. N. Engl. Botan. Club 1993, 95, 25–37. [Google Scholar]
- Walker, J.L.; Silletti, A.M. Restoring the ground layer of longleaf pine ecosystems. In The Longleaf Pine Ecosystem: Ecology, Silviculture, and Restoration; Jose, S., Jokela, E.J., Miller, D.L., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; pp. 297–325. [Google Scholar]
- Peet, R.; Allard, D. Longleaf pine vegetation of the southern atlantic and gulf coast regions: A preliminary classification. In Proceedings of the Annual Tall Timbers Fire Ecology Conference 1993, Tallahassee, FL, USA, May 1993; Volume 18, pp. 45–81. [Google Scholar]
- Way, A. Burned to be wild: Herbert Stoddard and the roots of ecological conservation in the southern longleaf pine forest. Environ. Hist. 2006, 11, 500–526. [Google Scholar] [CrossRef]
- Myers, F. Forest Management Plan for the Land of Historic Camden Foundation; Forest Resource Consultants & Real Estate Co.: Camden, SC, USA, 2017. [Google Scholar]
- Smith, S.; Legg, J.; Wilson, T. The Archeology of the Camden Battlefield: History, Private Collections, and Field Investigations; South Carolina Institute of Archeology and Anthropology: Columbia, SC, USA, 2009. [Google Scholar]
- Howell, E.; Kline, V. The role of competition in the successful establishment of selected prairie species. In Proceedings of the North American Prairie Conference 1992, Windsor, ON, Canada, 6–9 August 1992; Volume 13, pp. 193–198. [Google Scholar]
- Limb, R.; Fuhlendorf, S.; Engle, D.; Kerby, J. Growing-season disturbance in tallgrass prairie: Evaluating fire and grazing on Schzachyrium scoparium. Rangel. Ecol. Manag. 2011, 64, 28–36. [Google Scholar] [CrossRef]
- Johnson, E.E. Exploring Longleaf Pine Restoration in the Sandhills of South Carolina. Ph.D. Dissertation, Clemson University, Clemson, SC, USA, 2021. [Google Scholar]
- Mijnsbrugge, K.V.; Bischoff, A.; Smith, B. A question of origin: Where and how to collect seed ecological restoration. Basic Appl. Ecol. 2010, 11, 300–311. [Google Scholar] [CrossRef]
- Springer, T. Germination and early seedling growth of chaffy-seeded grasses at negative water potentials. Crop Sci. 2005, 45, 2075–2080. [Google Scholar] [CrossRef]
- NOAA National Centers for Environmental Information: Climate at a Glance. Available online: https://www.ncdc.noaa.gov/cag (accessed on 15 February 2021).
- National Gardening Association. NGA Tools & Apps: When to Plant Vegetables: The Garden Planting Calendar. Available online: https://garden.org/apps/calendar/?q=Kershaw%2C+SC (accessed on 20 March 2021).
- Meyer, M.; Gaynor, V. Effect of seeding date on establishment of native grasses. Nativ. Plants J. 2002, 3, 132–138. [Google Scholar]
- Dhillion, S.S.; Friese, C.F. The occurrence of mycorrhizas in prairies: Applications to ecological restoration. In Proceedings of the North American Prairie Conference 1992, Windsor, ON, Canada, 6–9 August 1992; Volume 13, pp. 103–114. [Google Scholar]
- Walker, E.A.; Hermann, M.; Kollmann, J. Grassland restoration by seeding: Seed source and growth form matter more than diversity. Appl. Veg. Sci. 2015, 18, 368–378. [Google Scholar] [CrossRef]
- Cox, A.; Gordon, D.; Slapcinsky, J.; Seamon, G. Understory restoration in longleaf pine sandhills. Nat. Areas J. 2004, 4, 14–24. [Google Scholar]
- Szitár, K.; Ónodi, G.; Somay, L.; Pándi, I.; Kucs, P.; Kröel-Dulay, G. Contrasting effects of land use legacies on grassland restoration in burnt pine plantations. Biol. Conserv. 2016, 201, 356–362. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Weeks | p-Value | CAM | RS | ||
|---|---|---|---|---|---|
| Average | Standard error | Average | Standard error | ||
| 7 | 0.0056 * | 7 | 1.871877 | 33 | 9.794654 |
| 9 | 0.0060 * | 10 | 2.95363 | 36 | 10.43949 |
| 11 | 0.0053 * | 15 | 3.692549 | 39 | 10.19393 |
| 13 | 0.2072 | 33 | 7.561725 | 53 | 15.52045 |
| 15 | 0.0699 | 81 | 16.82251 | 57 | 12.73483 |
| 18 | 0.0232 β | 77 | 11.62605 | 46 | 11.34509 |
| 21 | 0.0063 β | 72 | 10.24085 | 40 | 8.2786 |
| 25 | 0.0003 β | 68 | 9.194097 | 36 | 5.679928 |
| 29 | <0.0001 β | 63 | 7.901095 | 31 | 4.932561 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Johnson, E.; Hagan, A.H.; Hiesl, P. Seed Source for Restoration: Little Bluestem (Schizachyrium scoparium (Michx.) Nash) and the Carolina Sandhills. Ecologies 2023, 4, 344-354. https://doi.org/10.3390/ecologies4020022
Johnson E, Hagan AH, Hiesl P. Seed Source for Restoration: Little Bluestem (Schizachyrium scoparium (Michx.) Nash) and the Carolina Sandhills. Ecologies. 2023; 4(2):344-354. https://doi.org/10.3390/ecologies4020022
Chicago/Turabian StyleJohnson, Elizabeth, Althea Hotaling Hagan, and Patrick Hiesl. 2023. "Seed Source for Restoration: Little Bluestem (Schizachyrium scoparium (Michx.) Nash) and the Carolina Sandhills" Ecologies 4, no. 2: 344-354. https://doi.org/10.3390/ecologies4020022
APA StyleJohnson, E., Hagan, A. H., & Hiesl, P. (2023). Seed Source for Restoration: Little Bluestem (Schizachyrium scoparium (Michx.) Nash) and the Carolina Sandhills. Ecologies, 4(2), 344-354. https://doi.org/10.3390/ecologies4020022