Abstract
Harmful cyanobacterial blooms of the toxin-producing Microcystis have become a growing problem for Southwest Florida freshwater bodies. Recently, a 2016 bloom in Lake Okeechobee and a 2018 bloom in the Caloosahatchee River both led to the declaration of a state of emergency for the state of Florida. Fast-acting suppression methods are needed to protect residents and wildlife. Hydrogen peroxide and L-lysine have shown promising results in selectively inhibiting the growth of Microcystis aeruginosa and are more ecologically friendly due to fast degradation in water or the biological enhancement of nontarget organisms, respectively. We further explored the use of hydrogen peroxide, L-lysine, and combined treatments of both chemicals, which have never been tested before, for the rapid suppression of Microcystis. We assessed the susceptibility of seven M. aeruginosa strains and six other phytoplankton (Cyanobium spp., Synechococcus sp., Dolichospermum planctonica, Mychonastes homosphaera, and Chromochloris zofingiensis) commonly found in Florida, and revealed that susceptibility was diverse. All three treatments were effective at inhibiting the growth of M. aeruginosa, mixed treatments (16.7 mg/L hydrogen peroxide: 8 mg/L L-lysine) were most effective with a median growth inhibition ratio of 94.2% on the last day of the experiment, while hydrogen peroxide (16.7 mg/L) (83.8%) and L-lysine (8 mg/L) (78.5%) were less so. We found axenic M. aeruginosa to be significantly more sensitive to hydrogen peroxide when compared with nonaxenic strains (p < 0.01, n = 18). L-lysine was found to be significantly more toxic to M. aeruginosa than other examined cyanobacteria and chlorophyte strains at the end of the experiment (p < 0.001, n = 33), demonstrating its specificity to this cyanobacterium, while hydrogen peroxide and mixed treatments had varying effects on the other tested phytoplankton.
1. Introduction
Microcystis is one of the most common and widely distributed toxic bloom-forming cyanobacteria in freshwater and brackish ecosystems [1]. These organisms can produce microcystin toxins, which threaten human health and wildlife in aquatic systems [2]. In recent years, cyanobacterial harmful algal blooms (cyanoHABs) have become a growing problem for Southwest Florida (SWFL). Lake Okeechobee, the largest lake in the Southern United States, and its connecting waterways (the Caloosahatchee River and Saint Lucie River) frequently experience Microcystis blooms [3,4]. Recent cyanoHABs in Lake Okeechobee and both waterways led to the declaration of a state of emergency in 2016 and 2018 [4,5,6,7]. Nutrient reduction is currently considered the best strategy to reduce bloom events; however, these strategies can take many years to show positive results [7]. The characteristically shallow water depth of these subtropical freshwater systems and reduced stratification periods also leave them more vulnerable to nutrient loading, making nutrient reduction strategies more difficult [8,9]. Many short-term reduction methods have focused on chemical spraying methods, such as copper sulfate and aluminum; however, these chemicals can become concentrated in the environment and often are nonselective in targeting cyanobacteria [10]. Rapid bloom-suppression methods are desperately needed to manage cyanoHABs to reduce the risk to human health and wildlife [7].
Hydrogen peroxide has gained popularity as an environmentally safe algaecide that targets cyanobacteria over other eukaryotic algae due to differential coping mechanisms when dealing with oxidative stress [11,12,13]. This algaecide is considered environmentally safe due to fast degradation rates in water and ecologically friendly due to positive succession from toxic cyanobacteria to nontoxic eukaryotic algae [11]. L-lysine, an amino acid, has been found to selectively inhibit the growth of Microcystis over other eukaryotic algae and cyanobacteria [14]. Mechanisms targeting Microcystis specifically are still not well understood [15,16]. Pond experiments have had success in the removal of Microcystis blooms leading to positive succession by eukaryotic phytoplankton [15,17]. This biologically derived substance is also considered environmentally friendly; as an essential amino acid for many fish and aquatic organisms, it can be easily metabolized in these systems and is also freely soluble [15].
In this study, we assessed the use of L-lysine, hydrogen peroxide, and the novel use of both together for inhibiting the growth of Microcystis. The novel use of both L-lysine and hydrogen peroxide for the treatment of Microcystis has never been examined to our knowledge, and we hypothesized that the synergetic effects of both algaecides acting on different growth inhibition mechanisms toward Microcystis will be highly effective in their removal. For this study, we assessed both axenic and unialgal Microcystis strains, with different morphological characteristics to gain an overall understanding of treatment sensitivity. We also used three Microcystis strains isolated from the Caloosahatchee River, Southwest Florida to better understand suitable treatment options for regional water bodies, as well as six other commonly found phytoplankton taxa isolated in local areas, to gain insight into potential phytoplankton community succession patterns that may occur after treatment.
2. Materials and Methods
2.1. Organisms and Cultured Conditions
Microcystis aeruginosa strains used in this study are listed in Table 1 along with isolation location, ecological characteristics, as well as axenic or nonaxenic conditions. Microcystis strains FD4, HC1, and AL2 were isolated from the Caloosahatchee River and its branching waterways in the 2018 bloom event [6]. Microcystis were identified based on microscopic observation using a compound microscope (OMAX MD82ES10) and genomic characterization. NIES-88, 90, 102, 843, and 4325 were obtained from the Microbial Culture Collection, National Institute for Environmental Studies (NIES), Tsukuba, Japan (Table 1). Genomes of five of the M. aeruginosa strains used in this study (FD4, NIES-843, NIES-88, NIES-4325, and NIES-102) have been sequenced and the reported genome size ranged from 3.88 Mb (NIES-4325) to 5.87 Mb (NIES-102) [6,18,19,20,21]. NIES-102 was originally reported as Microcystis viridis, one of five commonly reported morphospecies of Microcystis (M. aeruginosa, M. viridis, M. novacekii, M. ichthyoblabe, and M. wesenbergii) [22]. Morphospecies of Microcystis have been recognized primarily by colony morphology which is highly variable, and DNA-DNA hybridization reports have shown in the testing of multiple Microcystis strains, including NIES-102, the genus cannot be distinguished by species because the DNA reassociation values reach 70% [23,24,25]. Kondo et al. [24], for example, found NIES-102 had high DNA relatedness to M. aeruginosa strains NIES-87 (90.5%), NIES-89 (91.2%), and NIES-289 (78.5%). More recent genome comparisons have recognized NIES-102 as M. aeruginosa [20,21] and, therefore, in this study, it will be referred to as M. aeruginosa. M. aeruginosa strains NIES-102, NIES-843, NIES-88, AL2, and HC1 all produce microcystins, while M. aeruginosa strains FD4 and NIES-4325 are nontoxic, lacking complete microcystin synthetase gene clusters [6,25]. Other phytoplankton taxa assessed in this study are listed in Table 2. These phytoplankton strains were also locally isolated and identified using microscopy and either 16S rRNA or 23S rRNA gene sequences. Accession numbers for deposited 23S rRNA gene sequences are available in GenBank: FGCU3 (OL772022), FGCU52 (OL772026), FGCU54 (OL772027), FGCU15 (OL772003), and FGCU59 (OL772005).

Table 1.
Microcystis aeruginosa strains used in this study.
Table 2.
Cyanobacteria and Chlorophyta taxa examined in this study.
Prior to the treatment experiments, phytoplankton were precultured to exponential phase, harvested, and then transferred to fresh BG-11. For the initial examination using M. aeruginosa strain FD4 and Cyanobium sp. strain FGCU54, 3 mL of culture was transferred to 27 mL of diluted BG-11 (10%). For the proceeding treatment experiments, 1 mL of culture was transferred to 9 mL of diluted BG-11. Cultures were incubated at 25 °C in a Precision Plant Growth Chamber at an irradiance of 27.8 µmol/m2/s and a light/dark cycle of 12/12 h. All tests were carried out in triplicate, where the same exponential phase batch culture was used for each strain’s examination.
M. aeruginosa FD4 and Cyanobium sp. FGCU54 were initially used to find the optimal concentrations of each treatment for the further testing of other M. aeruginosa and phytoplankton strains. L-lysine was added to a final concentration of 3 mg/L and 8 mg/L and compared to a control. Hydrogen peroxide was added to a final concentration of 16.7 mg/L and 33.3 mg/L and compared to a control. The initial low- and high-dose concentrations of L-lysine and hydrogen peroxide treatments were then combined for the assessment of mixed treatments of both chemicals (low dose—3 mg/L L-lysine: 16.7 mg/L hydrogen peroxide; high dose—8 mg/L L-lysine: 33.3 mg/L hydrogen peroxide). Treatment experiments ran for an 8-day period. After the initial examination, it was found that a single application of high-dose hydrogen peroxide greatly suppressed the growth of Cyanobium sp. FGCU54, leading to the decision to use low-dose concentrations (16.7 mg/L) of hydrogen peroxide for further testing. The minimal impact of high-dose L-lysine concentration (8 mg/L) on Cyanobium sp. FGCU54 led to the decision to move forward with testing this concentration, and the final mixed treatment concentrations of 16.7 mg/L hydrogen peroxide: 8 mg/L L-lysine. The proceeding treatment experiments examining other M. aeruginosa strains and phytoplankton ran for a period of 7 days, with optical density (OD) measurements taken at the start of the experiment and on days 3 and 7.
2.2. Growth Measurements of Phytoplankton
Phytoplankton growth was examined spectrophotometrically at an OD of 750 nm using a HACH DR/2400 spectrophotometer. Growth was examined over a 7- to 8-day period. Prior to absorbance measurements, cultures were vortexed to disperse colonies and attain a homogeneous mixture. Three replicate readings were used for each treatment culture to average the optical density readings for each recording day. Relative growth inhibition (GI) ratios were calculated using the formula:
where ODc and ODt are the OD growth at 750 nm of the control and treated samples, respectively. The ODc readings for control samples were averaged (n = 3) for comparison to treatment replicates, which are displayed as ± standard deviation.
(GI)= [(ODc-ODt)/ODc] × 100
2.3. Fluorescence Microscopy
Formalin-fixed cells (20 µL) were diluted to 1 mL with ultrapure water and fluorescently stained with 1 µL of SYBR Green I (Lonza Bioscience, Wakersville, MD, USA) for 5 min in the dark. The stained cells were collected onto a 0.2 µm pore size Isopore membrane filter (25 mm, MilliporeSigma, Burlington, MA, USA) set in a vacuum filtration unit with a hand-held pump. After filtration, the filter was mounted on a microscope slide with an antibleaching agent (AF1: Citifluor, London, UK). The cells were viewed with an inverted fluorescence microscope IX51 (Olympus, Center Valley, PA, USA).
2.4. Statistical Analysis
The Shapiro–Wilk test was used to analyze the normality of growth inhibition ratio data for the tested phytoplankton groups. Since some data were nonparametric, a two-tailed Wilcoxon rank sum test was used to analyze the effects of hydrogen peroxide, L-lysine, and mixed treatments on the growth of examined phytoplankton in R [26]. The growth inhibition ratios from each replicate phytoplankton strain (n = 3) were grouped based on described characteristics (axenic, nonaxenic, M. aeruginosa, cyanobacteria, chlorophytes, etc.), for each treatment application on individual sampling days (3 and 7), to assess differences in treatment sensitivity among the tested phytoplankton groups.
3. Results and Discussion
3.1. Dose Sensitivity of Microcystis aeruginosa and Cyanobium sp.
M. aeruginosa strain FD4 and Cyanobium sp. FGCU54 were initially used to find the optimal concentrations of each treatment for further testing of the susceptibility of other algal strains to hydrogen peroxide, L-lysine, and mixed treatments of both chemicals. All treatment application concentrations showed an inhibitory effect on the growth of M. aeruginosa FD4 after two days; this was not found for Cyanobium sp. FGCU54 (Figure 1). The inhibitory effect on M. aeruginosa varied after this period by treatment application and concentration. Low-dose L-lysine (3 mg/L) and hydrogen peroxide (16.7 mg/L) treatments showed signs of recovery on the last day of the experiment, while mixed low-dose treatments of both chemicals (3 mg/L L-lysine: 16.7 mg/L hydrogen peroxide) showed synergetic effects in deterring recovery, with a growth inhibition ratio of 81.6 ± 0.7% (n = 3) on the last day (Figure 1). M. aeruginosa strain FD4 was found to be highly sensitive to 8 mg/L L-lysine treatment, with growth inhibition reaching 90% just 48 h after treatment application, while Cyanobium sp. FGCU54 did not show signs of sensitivity to any of the L-lysine treatments; however, it was found to be more sensitive to high-dose hydrogen peroxide (33.3 mg/L) (Figure 1). High-dose mixed treatments of both chemicals were the most effective at inhibiting the growth of M. aeruginosa.
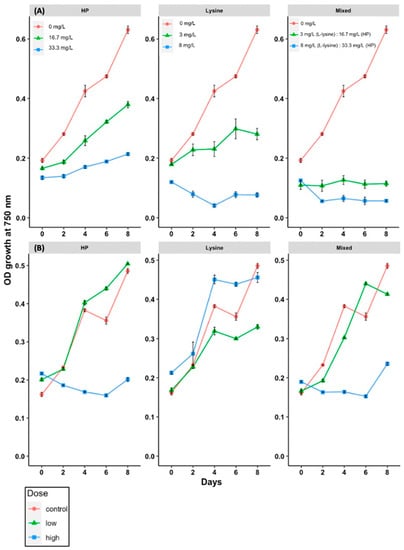
Figure 1.
Susceptibility of Microcystis aeruginosa FD4 (A) and Cyanobium sp. FGCU54 (B) to low- and high-dose hydrogen peroxide (16.7 mg/L, 33.3 mg/L), L-lysine (3 mg/L, 8 mg/L), and mixed treatments (3 mg/L L-lysine: 16.7 mg/L hydrogen peroxide, 8 mg/L L-lysine: 33.3 mg/L hydrogen peroxide). Growth was measured by optical density (OD) at 750 nm.
To better understand strain sensitivity and safe application for the natural environment, it was decided to proceed with the low-dose hydrogen peroxide (16.7 mg/L) for a single application and mixed treatments (16.7 mg/L hydrogen peroxide and 8 mg/L L-lysine), and high-dose L-lysine (8 mg/L).
3.2. Differences in Sensitivity of Microcystis aeruginosa Strains to Hydrogen Peroxide and L-Lysine Treatments
To examine the variable susceptibility of M. aeruginosa strains to hydrogen peroxide, L-lysine, and mixed treatments, three axenic strains (NIES-843, NIES-4325, and NIES-102) and three nonaxenic strains (NIES-88, HC1, AL2) were assessed in a 7-day treatment experiment (Table 1). M. aeruginosa strains were found to differ in sensitivity to treatments. Mixed treatments (16.7 mg/L hydrogen peroxide and 8 mg/L L-lysine) were found to be most effective at inhibiting the growth of M. aeruginosa (Figure 2). By day 3 of the experiment, growth at OD 750 nm for all the mixed treatment samples had dropped to below 50% of the measured OD 750 nm growth for the control samples. The mixed treatments resulted in the near-complete growth inhibition of the tested M. aeruginosa strains by day 7, with a median relative growth inhibition of 94.2% (interquartile range (IQR): 79.4–96.3%, n = 18). M. aeruginosa strains had variable sensitivity to hydrogen peroxide (16.7 mg/L) treatments, displaying near-total growth inhibition on M. aeruginosa strains NIES-102, NIES-4325, and NIES-88, ranging from 95 to 97% of growth inhibition on day 7 (Figure 3), while AL2 was found to grow under hydrogen peroxide treatments (Figure 2 and Figure 3). Hydrogen peroxide treatment application was found to have faster growth-inhibitory effects on M. aeruginosa than L-lysine applications, with a median growth inhibition of 76.4% (IQR: 57.6–88.6%) on day 3 of the experiment compared to 47.6% for L-lysine (IQR: 33.2–64.8%); by the end of the experiment, however, the median growth inhibition for L-lysine-treated M. aeruginosa reached 78.5% by day 7 (Figure 3).
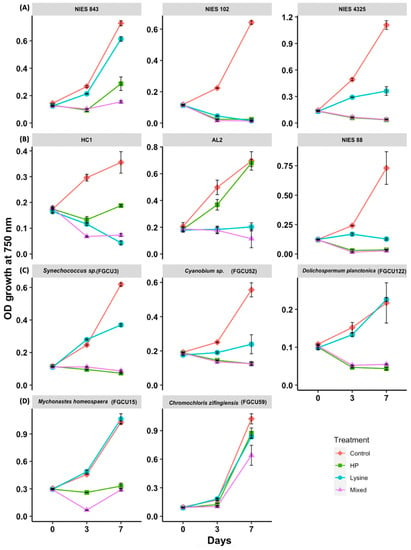
Figure 2.
Susceptibility of axenic Microcystis aeruginosa strains: NIES-843, NIES-102, and NIES-4325 (A), nonaxenic Microcystis strains: HC1, AL2, and NIES 88 (B), Southwest Florida isolated cyanobacterial strains: Synechococcus sp. FGCU3, Cyanobium sp. FGCU52, and Dolichospermum planctonica FGCU122 (C), and chlorophyte strains: Mychonastes homosphaera FGCU15 and Chromochloris zofingiensis FGCU59 (D), to hydrogen peroxide (16.7 mg/L), L-lysine (8 mg/L), and mixed treatments of both chemicals (16.7 mg/L hydrogen peroxide: 8 mg/L L-lysine).
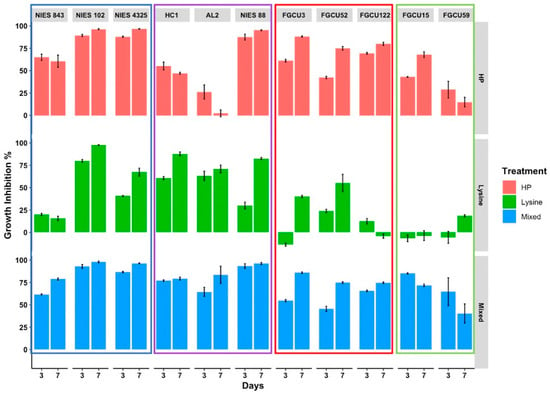
Figure 3.
Relative growth inhibition (%) for axenic Microcystis strains: NIES-843, NIES-102, and NIES-4325 (blue box), nonaxenic Microcystis strains: HC1, AL2, and NIES-88 (purple box), FGCU cyanobacteria strains: Synechococcus sp. FGCU3, Cyanobium sp. FGCU52, and Dolichospermum planctonica FGCU122 (red box), and FGCU chlorophyte strains: Mychonastes homosphaera FGCU15 and Chromochloris zofingiensis FGCU59 (green box) under hydrogen peroxide, L-lysine, and mixed treatments.
The growth of axenic M. aeruginosa (NIES-102, NIES-843, and NIES-4325) was found to be more inhibited by hydrogen peroxide application during the experiment; when compared with nonaxenic M. aeruginosa (AL2, HC1, and NIES-88) on day 7, growth was found to be significantly different (p < 0.01, n = 18). This finding was more significant when comparing the growth of only SWFL M. aeruginosa strains (HC1 and AL2) with the growth of axenic strains on day 7 (p < 0.001, n = 15); resilience to hydrogen peroxide treatments was also seen for the SWFL strain FD4 (Figure 1) and may be attributed to colony formation and the high bacterial presence found in these SWFL nonaxenic strains. M. aeruginosa lacks the genes for catalase which are used for reactive oxygen species (ROS) scavenging under high hydrogen peroxide conditions [27]. Kim et al. [28] found that M. aeruginosa cocultured with Rhizobium bacteria containing catalase genes had significantly higher growth rates than axenic M. aeruginosa cultures under 500 µM hydrogen peroxide treatments. Kim et al. [29] further confirmed this bacterial ROS remediation activity using additional high light stress along with hydrogen peroxide treatments, where M. aeruginosa cocultured with high-catalase-functioning bacterial groups were less sensitive to both ROS conditions compared to M. aeruginosa cocultured with low-catalase-functioning bacterial groups; hydrogen peroxide was also degraded significantly faster in high-catalase bacterial communities. This may have been demonstrated in our results, where axenic M. aeruginosa strains (NIES-843, NIES-102, and NIES-4325) were more sensitive to hydrogen peroxide than nonaxenic SWFL strains (FD4, HC1, and AL2) (Figure 1, Figure 2, Figure 3 and Figure 4). Another difference that may have lessened SWFL strain hydrogen peroxide sensitivity compared with unicellular axenic M. aeruginosa is these strains’ phenotypic characteristic of colony formation, which may be an adaptive advantage for M. aeruginosa species found in this subtropical region. Subtropical regions experience greater solar ultraviolet (UV) radiation, where algal cells are at an enhanced risk of UV damage and photoinhibition at the water surface [30]. M. aeruginosa colonies have been found to be better adapted to these high-light conditions compared with unicells due to self-shading mechanisms and increased extracellular polysaccharide content, which can facilitate the attachment of UV-B screening compounds [31,32]. Colony formation may also be favored due to environmental chemical stressors [33]. Ndungu et al. [5] found that along the Caloosahatchee River, where SWFL M. aeruginosa strains were isolated, hydrogen peroxide surface water concentrations were measured as high as 5.07 µM during bloom and rain events may increase the surface hydrogen peroxide concentrations, suggesting phytoplankton from this region may be exposed to high levels of environmental oxidative stress. Extracellular polymeric substances (EPS) are essential components of M. aeruginosa colony formation [34] and have been shown to have a strong scavenging ability toward hydrogen peroxide. Gao et al. [35] found in the hydrogen peroxide treatment of M. aeruginosa that 50% of the applied hydrogen peroxide could be consumed by EPS, for which M. aeruginosa cells with EPS were able to recover much quicker than those without, highlighting the potential reduced sensitivity of these colony-forming strains to hydrogen peroxide in comparison with unicellular forms. Colonies may also be better protected from these conditions due to the reduced surface area interaction of inner cells with chemical stressors [33] as well as increased antioxidative enzyme activity, such as superoxide dismutase (SOD), which has been reported in observations of colonies and unicellular cells under oxidative stress [36]. These unique conditions may promote the survival of M. aeruginosa morphospecies with phenotypic plasticity toward colony formation, allowing better protection from these localized environmental stressors, as well as heterotrophic bacterial communities capable of remediating this environmental ROS stress, together making them less sensitive to hydrogen peroxide algicidal treatments [28,35,36]. This idea is supported by the dense colony morphology and high bacterial presence found in the mucilage of SWFL strain AL2 (Figure 4), which was least sensitive to hydrogen peroxide of the tested M. aeruginosa strains (Figure 2 and Figure 3). Liu et al. [37] reported that a dosage of 20 mg/L was necessary to efficiently control Microcystis blooms whose colony size exceeds 25 μm, which the colony size of the examined M. aeruginosa AL2 far surpasses (>100 µm) (Figure 4). These findings may hold an important insight for biological controls of M. aeruginosa blooms in which higher concentrations of hydrogen peroxide are necessary to more effectively remove the bloom than the simplified laboratory experimental conditions.
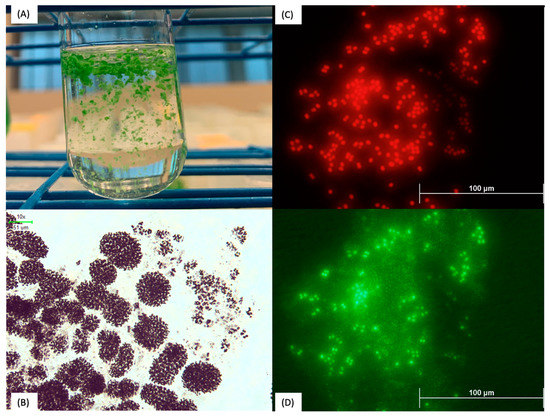
Figure 4.
Microcystis aeruginosa AL2 growing in BG-11 medium. (A) Surface scum, (B) colony, (C) autofluorescence of M. aeruginosa cells, and (D) M. aeruginosa and phycosphere bacteria. Large coccoid cells are M. aeruginosa and smaller cells are heterotrophs embedded in mucilage. It should be noted that some M. aeruginosa cells are not stained by SYBR Green I, possibly due to the incomplete penetration of the dye through mucilage.
As demonstrated, the M. aeruginosa strains were highly sensitive to L-lysine treatment compared with the other examined phytoplankton (Figure 2 and Figure 3). Our results support the previous finding that L-lysine can be selectively toxic toward M. aeruginosa over other phytoplankton taxa (Figure 2 and Figure 3) [14,38]. M. aeruginosa’s sensitivity to 8 mg/L L-lysine addition was diverse, with NIES-102 showing 79.1% growth inhibition 3 days after addition, and near-total growth inhibition at the end of the experiment, while NIES-843 was largely found to be nonsensitive (Figure 3). M. aeruginosa strains NIES-88, AL2, and HC1 were also found to be highly sensitive to L-lysine (growth inhibition: >74%) (Figure 3). Unlike hydrogen-peroxide-treated M. aeruginosa, significant differences between the growth of axenic and nonaxenic strains were not found under L-lysine application. L-lysine toxicity mechanisms on M. aeruginosa, over other phytoplankton and cyanobacterial genera, are still not well understood [14]. Takamura et al. [15] suggested meso-diaminopimelic acid may be replaced by L-lysine in cell wall peptidoglycan, and differentiation in cyanobacterial sensitivity may be the result of carbon sources used for growth or varied metabolization rates of amino acids. Zimba et al. [38] examined M. aeruginosa sensitivity to L-arginine and L-leucine which had more and fewer nitrogen groups than L-lysine, respectively, and found these basic amino acids were not inhibitory to growth, postulating that L-lysine may increase enzyme active sites, causing feedback inhibition relating to Chl-a accumulation. Tian et al. [16] found that M. aeruginosa cells treated with L-lysine had significant increases in malondialdehyde (MDA) content, likely causing lipid peroxidation cell membrane damage, and oxidative stress relief mechanisms (e.g., SOD) were induced. These mechanisms inhibiting M. aeruginosa growth warrant further study as L-lysine could act as an environmentally safe M. aeruginosa-specific bloom mitigation method, where the essential amino acids can be easily metabolized and degraded in freshwater ecosystems [15,16]. The two most recent high-resolution studies also supported these ideas and concluded that L-lysine incorporated into the leaky peptide glycan structure caused irreversible damage to the photosynthetic system and membrane integrity [39], and affected arginine metabolism and the ornithine–ammonia cycle to inhibit the growth of M. aeruginosa [40]. Although our results support previous findings of M. aeruginosa sensitivity to L-lysine, longer duration laboratory and field application studies are needed to fully evaluate the effective use of L-lysine for the removal of nuisance M. aeruginosa blooms. For example, Lürling and Van Oosterhout [41] found L-lysine additions above 4.3 mg/L were sufficient for a complete growth inhibition of M. aeruginosa; however, photosystem II efficiency showed recovery after 6 days.
The novel use of combining both hydrogen peroxide and L-lysine for cyanoHAB treatment was found to be highly successful at inhibiting the growth of the examined M. aeruginosa strains (Figure 2 and Figure 3). Mixed treatments resulted in the near-complete growth inhibition of all the tested M. aeruginosa strains, showing no variation in toxicity based on the presence of bacteria, or unicellular and colony morphological characteristics (Figure 2 and Figure 3). Past algaecide combination treatments have largely focused on combinations of chemical and coagulant algaecides; however, coagulants are not suitable for shallow lakes where resuspension is prevalent and have been noted to cause potential ecological harm through the nonselective targeting of phytoplankton [42], growth reductions of aquatic plants [43], acute toxicity to fish and benthic invertebrates [44], and an accumulation of coagulant in sediments [45]. Given the efficacy of the fast degradation rates found for both L-lysine and hydrogen peroxide, while also providing more selective targeting of cyanobacteria over eukaryotic algae [11,15], we suggest this chemical and biological combination warrants further exploration for the water management of M. aeruginosa where synergetic effects are achieved, similar to other algicidal combinations, without causing enhanced ecological harm. Although hydrogen peroxide is recognized as being a safe and effective cyanoHAB mitigation method due to its fast degradation rates and reduced sensitivity to beneficial eukaryotic plankton, low-dose application to achieve these benefits may not be sufficient to eliminate dense M. aeruginosa blooms with the environmental and physiological characteristics discussed previously [11,28,35,46]. This can lead to the need for the costly reapplication of chemicals, where residents and wildlife have prolonged risks of exposure to cyanotoxins. Our results demonstrate that mixed treatments of hydrogen peroxide and L-lysine chemicals were highly effective at inhibiting the growth of M. aeruginosa in just 3 days (median growth inhibition: 82.1%) (Figure 3).
3.3. Differences in Sensitivity of Other Phytoplankton to Hydrogen Peroxide and L-Lysine Treatments
Other Cyanophyta and Chlorophyta isolated from SWFL water bodies were used to assess sensitivity and potential succession that may occur if hydrogen peroxide, L-lysine, and mixed treatments were applied to treat local M. aeruginosa blooms. Similar to Cyanobium sp. FGCU54 (Figure 1), two other picocyanobacterial strains (FGCU3 and FGCU52) were found to be less sensitive to L-lysine than the examined M. aeruginosa strains on the last day of the experiment (p < 0.05, n = 24) (Figure 2 and Figure 3). These strains were found, however, to be sensitive to hydrogen peroxide, with the average growth inhibition ratios on the last day of the experiment being 88.1% for Synechococcus sp. FGCU3 and 74.9% for Cyanobium sp. FGCU52 (Figure 3). The growth of Dolichospermum planctonica strain FGCU122 was also not susceptible to L-lysine addition (Figure 2 and Figure 3). This finding agrees with past studies showing Anabaena, also a member of the order Nostocales, was not sensitive to L-lysine concentrations up to 10 mg/L [14]. FGCU122 was highly sensitive to hydrogen peroxide and the growth never recovered after the initial crash on day 3 (Figure 2), supporting previous findings of a 2 mg/L hydrogen peroxide dose being required to completely remove Dolichospermum circinale [47]. Dolichospermum are another toxin-producing cyanobacterium known to frequently form nuisance blooms in SWFL [48], and this result highlights the effectiveness of hydrogen peroxide on Dolichospermum blooms [49]. All the aforementioned cyanobacterial strains (FGCU3, FGCU52, and FGCU122) were found to be highly sensitive to mixed treatments, with the growth inhibition ranging from 73.2 to 87.5% (n = 9) on the last day of the experiment (Figure 3). The examined SWFL chlorophytes had varying sensitivity to hydrogen peroxide, L-lysine, and mixed treatments (Figure 2). Both chlorophyte Chromochloris zofingiensis strain FGCU59 and Mychonastes homosphaera strain FGCU15 were not sensitive to L-lysine (Figure 3). This result is supported by previous findings indicating eukaryotes are less susceptible to L-lysine; Zimba et al. [38] found the Chl-a accumulation for the chlorophyte Scenedesmus dimorphus was not inhibited by L-lysine concentrations < 400 mg/L, and the diatom Cyclotella meneghiniana was not sensitive up to concentrations of 500 mg/L. C. zofingiensis growth was not found to be inhibited by hydrogen peroxide and was the least susceptible of the tested taxa to mixed-treatment applications (Figure 3 and Figure 4). M. homosphaera strain FGCU15, a small picoeukaryote, was susceptible to these treatments; however, it showed signs of recovery on the last day (Figure 2).
The specificity of L-lysine toxicity to M. aeruginosa was clearly demonstrated in this study, where M. aeruginosa growth was found to be significantly more inhibited when compared with other examined cyanobacteria and chlorophytes (FGCU3, FGCU15, FGCU52, FGCU59, and FGCU122) on days 3 (p < 0.001, n = 33) and 7 (p < 0.001, n = 33) (Figure 3). These results demonstrate L-lysine may be a viable option for targeting M. aeruginosa blooms specifically, leading potentially to succession by eukaryotic algae. Takamura et al. [15] found in a pond experiment that L-lysine application of 50 μM was able to completely remove M. aeruginosa colonies after 3 days, where the eukaryote Euglena then became dominant and M. aeruginosa remained absent for the proceeding 11 days of the experiment. Kaya et al. [17] had different findings in pond experiments, where, after L-lysine application, M. aeruginosa dominance was succeeded by diatoms, but only lasted 7 days before M. aeruginosa returned. Our laboratory results, therefore, are cautionary findings, and the further investigation of L-lysine toxicity on M. aeruginosa growth suppression should be assessed for further field applications as well as other potential environmental impacts. Given the high nitrogen content in L-lysine, nutrient regimes and further eutrophication should be evaluated when potentially using this algaecide. Our concentration of 8 mg/L would be equivalent to 1.5 mg/L as nitrogen when applied [15,50], which is negligible because the expected dose could be extremely small in comparison with the size of a water body. The severe cell lysing of M. aeruginosa cells could also influence dissolved oxygen (DO); 6.7 mg/L application in a pond experiment led to a 70% decline in DO after application. However, levels eventually recovered to those of controls, but low DO periods could impact other biota [17]. In the aforementioned experiment, cladoceran zooplankton as well as two macrophytes were found to increase in comparison with controls. Being an essential amino acid, this algaecide likely will not cause direct harm to nontarget biota; however, these benefits should be balanced with the potential impacts to ecosystem functioning also described. The sensitivity of other cyanobacteria (FGCU3, FGCU52, and FGCU122) to L-lysine was significantly less than M. aeruginosa on the last day of the experiment (p < 0.01, n = 27) (Figure 3), showing this treatment option does not have broad application potential for the removal of other CyanoHAB-forming taxa as is found with hydrogen-peroxide-based algaecides [14]. From our understanding, picocyanobacterial sensitivity to L-lysine application is limited, and our finding that the tested strains (FGCU3, FGCU52, and FGCU54) were less sensitive than most examined M. aeruginosa strains is beneficial as these organisms are found to have high annual abundance in this subtropical region [48,51], where they contribute significantly to carbon fixation and biogeochemical cycling [52].
Although highlighted for selective toxicity toward cyanobacteria, at high concentrations, hydrogen peroxide can be harmful to nontarget phytoplankton and biota [11,53]. Growth inhibition toward the chlorophyte M. homosphaera strain FGCU15 was observed with hydrogen peroxide treatments (16.7 mg/L) (Figure 2 and Figure 3). However, its small cell size (1.5–4.5 µm) may be why it was found to be more sensitive than chlorophyte C. zofingiensis strain FGCU59. The growth inhibition of hydrogen-peroxide-treated FGCU59 (14.8% ± 9.3%) was significantly less than that of the tested cyanobacteria taxa (Table 1 and Table 2) on the last day of the experiment (p < 0.02, n = 30), which had a median growth inhibition of 79.6% (n = 27). This finding reinforces the commonly reported characteristic of hydrogen-peroxide-based algaecide’s specific targeting of cyanobacteria while having a lesser impact on eukaryotic algae [11,12,46]. For instance, in a study by Weenink et al. [54], it was observed that the photosynthetic yield of Chlorophyte strains, including Chlorella, Desmodesmus, Kirchneriella, Ankistrodesmus, and Monophoridium, was significantly inhibited only when the concentration of hydrogen peroxide exceeded 35 mg/L. On the other hand, Chlamydomonas showed no sensitivity until the dosage reached 75 mg/L. Although our 16.7 mg/L exceeds the recommended environmental dosage of >5 mg/L [11], the high concentration may be necessary for the removal of M. aeruginosa. Yang et al. [12] found the EC50 concentration of hydrogen peroxide necessary for Microcystis to be ten times higher (5.06 mg/L) than that of Anabaena (0.50 mg/L), Cylindrospermopsis (0.32 mg/L), and Planktothrix (0.42 mg/L); this, along with the previously described resilience of colony-forming M. aeruginosa [35,37], was the reason for our comparatively high hydrogen peroxide dosage selection. This dosage could impact nontarget zooplankton and macroinvertebrates and, therefore, the consideration of nontarget impacts should be weighed with ecological benefits of cyanoHAB removal. Yang et al. [12] found little impact on zooplankton at a hydrogen peroxide concentration of 6.7 mg/L in mesocosm experiments, while 20 mg/L dosage caused significant declines in comparison with controls. In an aquaculture pond experiment comparing algaecides, liquid 10.2 mg/L hydrogen peroxide application was found to significantly reduce zooplankton one day after application; however, biomass was found to steadily increase over 35 days, where at the termination of the experiment, the zooplankton biomass was similar to that of controls [55]. The two copper-based products were found to have the lowest zooplankton biomass at the end of the experiment, while another sodium carbonate peroxyhydrate product was also found to have significantly lower zooplankton biomass. Concerns over nontarget organism impacts in using high-dose hydrogen peroxide, therefore, are valid. However, a shortfall in mesocosm studies as well as laboratory toxicity trials is the removal of active avoidance behaviors these organisms may exhibit in the natural environment when exposed to surface-layer hydrogen peroxide [56], affecting exposure time. Exposure is also impacted by the residence time and degradation rates of hydrogen peroxide in water bodies of varying physiochemical and biological characteristics [46,57]. The removal of nontarget organisms is of critical concern and should be assessed based on distinct water body characteristics, where the potential for the removal of nontarget organisms should be avoided, or weighed with the ecological risk of continued persistence of M. aeruginosa.
The other SWFL isolated cyanobacteria and chlorophyte strains were also found to be highly sensitive to mixed-treatment applications. Chlorophyte strain FGCU59 was again found to have reduced sensitivity, with the growth inhibition (40.3% ± 18.6%) on the last day of the experiment being significantly less than that of all examined cyanobacteria strains (Table 1 and Table 2) (p < 0.001, n = 30), which had a median growth inhibition of 84.5% (Figure 3). These results suggest the combination of hydrogen peroxide and L-lysine treatments may be a viable option for targeting M. aeruginosa blooms which could lead to positive succession by eukaryotic algae. Residence time and other environmental factors could greatly influence the degradation of hydrogen peroxide as well as L-lysine in the natural environment and, therefore, further examination through field studies is needed to assess the appropriate dosage for safe and effective application [46,50]. Our result of M. aeruginosa FD4 as sensitive to mixed combinations of low-dose hydrogen peroxide (16.7 mg/L) and L-lysine (3 mg/L) suggests that lowered concentrations to achieve ecologically beneficial results may still be effective for the removal of M. aeruginosa blooms. Although not examined in this study, L-lysine combinations with lower-dose hydrogen peroxide could be used for the improved protection of nontarget organisms while still attaining the benefits of M. aeruginosa removal, and should be explored further in laboratory and field studies. Wang et al. [46] found that a 10.2 mg/L single-application hydrogen peroxide dose was sufficient to completely remove Microcystis, and conceivably, given the examined M. aeruginosa sensitivity to L-lysine, a lower hydrogen peroxide dose application along with L-lysine may be sufficient to remove Microcystis.
3.4. Other Application Considerations
Although not examined in the present study, the potential for microcystin exposure after the application of the described algaecides and M. aeruginosa cell lysis, releasing intracellular microcystin, is an important consideration in the application of any algaecide. From our understanding, extracellular measurements of microcystin after Microcystis exposure to L-lysine have not been examined. In a pond treatment of 9.1 mg/L L-lysine, Kaya et al. [17] found that the total microcystin concentration dropped by half two days after application in the treated pond and further decreased in concentration up to day 7, until reaching the starting value on day 28 after Microcystis had reappeared. The total microcystin was significantly less than the controls, however, throughout the 28-day experiment. In another experiment, the exposure of Microcystis to 100 µmol/L (14.62 mg/L) L-lysine after no nitrogen incubation for 7 days was reported to result in continually decreasing microcystin production; even when compared with controls that continued N-starvation, microcystin production was always less in L-lysine treated Microcystis [58]. These results suggest L-lysine can reduce intracellular microcystin content, and application may result in an overall reduction in microcystin.
The release of intracellular microcystin after hydrogen peroxide application has been reported. Yang et al. [12] observed an increase in measured extracellular microcystin 24 h after hydrogen peroxide applications (1.3 mg/L, 6.7 mg/L, and 20 mg/L); however, after 7 days, these levels returned to those seen in the control mesocosms, and the intracellular levels for higher doses (6.7 mg/L and 20 mg/L) were less than that in the controls. Chen et al. [59] similarly observed a sharp increase in extracellular microcystin 12 h after 10 and 20 mg/L hydrogen peroxide application; however, microcystin concentration returned to similar starting values one day later. The risk of microcystin exposure after hydrogen peroxide application, therefore, is warranted; however, other beneficial findings have also shown reduced transcription of microcystin synthetase genes after hydrogen peroxide exposure [60], as well as the potential capabilities of oxidized hydrogen peroxide to degrade extracellular microcystin in the water column [59,61], which—in comparison with other algaecides—could also reduce the potential risk of toxin exposure. The intracellular release of microcystin under combined hydrogen peroxide and L-lysine application, therefore, is a concern for the use of this treatment and should be further examined in laboratory studies to assess the risk.
The estimated market price of L-lysine per ton has been previously calculated to range from USD 1000–2000 [41,62], which is similar to the described cost of hydrogen peroxide per ton (USD 500–2000) [59]. Without doubt, the combined use of both algaecides would be costly; however, given the extreme sensitivity of the examined M. aeruginosa to this combined treatment, a one-time application to completely remove M. aeruginosa may be more achievable, reducing the potential costs of reapplication in comparison with singular chemical treatments. Although more expensive than some metal-based algaecides and coagulants [62], many of the ecological benefits, such as fast degradation rates and low toxicity for nontarget organisms, may outweigh these costs. Copper sulfate, for instance, is a cheap, widely used algaecide; however, it has proven to be nonselectively toxic to other organisms such as fish as well as accumulate in sediments, further impairing ecosystem functioning and biota [10,42,63].
4. Conclusions
In summary, our laboratory-scale experiment provided valuable information on the application concentrations and comparative toxicity of M. aeruginosa and other phytoplankton to L-lysine, hydrogen peroxide, and mixed treatments for future implementation at larger scales. We found that M. aeruginosa sensitivity to L-lysine and hydrogen peroxide treatments varied by strain; axenic M. aeruginosa strains were significantly more sensitive to hydrogen peroxide than nonaxenic strains, while the growth of axenic and nonaxenic M. aeruginosa did not differ for L-lysine. All the tested M. aeruginosa strains were highly sensitive to mixed treatments of L-lysine and hydrogen peroxide, showing this combination can be an effective mitigation method. L-lysine was found to selectively inhibit the growth of M. aeruginosa, while the other tested phytoplankton had reduced or no sensitivity. Some of these strains were sensitive to hydrogen peroxide and mixed treatments; however, chlorophyte C. zofingiensis was found to be more resilient to both treatments. Given the growing concern over increased eutrophication and incidences of M. aeruginosa blooms worldwide, information such as this on environmentally friendly algaecide options can greatly assist water managers, taking into consideration the environmental and ecological characteristics of applied water bodies.
Author Contributions
Conceptualization, E.K.D. and H.U.; methodology, E.K.D. and H.U.; validation, E.K.D.; formal analysis, E.K.D.; investigation, E.K.D.; data curation, E.K.D. and H.U.; writing—original draft, E.K.D.; writing—review and editing, E.K.D. and H.U.; supervision, H.U.; project administration, H.U.; funding acquisition, H.U. All authors have read and agreed to the published version of the manuscript.
Funding
This research was partially supported by the Florida Department of Environmental Protection Innovative Technology Grant INV_007, and the NSF Division of Environmental Biology grant DEB-1664052. Elizabeth Dahedl and Hidetoshi Urakawa were also supported by Seidler Student/Faculty Scholarly Collaboration Fellowship (2020 summer research) at Florida Gulf Coast University.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Acknowledgments
We thank Julia Davis, Michael Kratz, and Taylor Hancock, Haruka Urakawa at Florida Gulf Coast University for their assistance.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Falconer, I.R. An overview of problems caused by toxic blue–green algae (cyanobacteria) in drinking and recreational water. Environ. Toxicol. 1999, 14, 5–12. [Google Scholar] [CrossRef]
- Guedes, I.A.; da Costa Leite, D.M.; Manhães, L.A.; Bisch, P.M.; Azevedo, S.M.; Pacheco, A.B.F. Fluctuations in microcystin concentrations, potentially toxic Microcystis and genotype diversity in a cyanobacterial community from a tropical reservoir. Harmful Algae 2014, 39, 303–309. [Google Scholar] [CrossRef]
- Oehrle, S.; Rodriguez-Matos, M.; Cartamil, M.; Zavala, C.; Rein, K.S. Toxin composition of the 2016 Microcystis aeruginosa bloom in the St. Lucie Estuary, Florida. Toxicon 2017, 138, 169–172. [Google Scholar] [CrossRef] [PubMed]
- Kramer, B.J.; Davis, T.W.; Meyer, K.A.; Rosen, B.H.; Goleski, J.A.; Dick, G.J.; Oh, G.; Gobler, C.J. Nitrogen limitation, toxin synthesis potential, and toxicity of cyanobacterial populations in Lake Okeechobee and the St. Lucie River Estuary, Florida, during the 2016 state of emergency event. PLoS ONE 2018, 13, e0196278. [Google Scholar] [CrossRef] [PubMed]
- Ndungu, L.K.; Steele, J.H.; Hancock, T.L.; Bartleson, R.D.; Milbrandt, E.C.; Parsons, M.L.; Urakawa, H. Hydrogen peroxide measurements in subtropical aquatic systems and their implications for cyanobacterial blooms. Ecol. Eng. 2019, 138, 444–453. [Google Scholar] [CrossRef]
- Urakawa, H.; Hancock, T.L.; Steele, J.H.; Dahedl, E.K.; Urakawa, H.E.; Ndungu, L.K.; Krausfeldt, L.E.; Rosen, B.H.; Lopez, J.V. Complete genome sequence of Microcystis aeruginosa FD4, isolated from a subtropical river in southwest Florida. Microbiol. Resour. Announc. 2020, 9, e00813-20. [Google Scholar] [CrossRef] [PubMed]
- Kinley-Baird, C.; Calomeni, A.; Berthold, D.E.; Lefler, F.W.; Barbosa, M.; Rodgers, J.H.; Laughinghouse IV, H.D. Laboratory-scale evaluation of algaecide effectiveness for control of microcystin-producing cyanobacteria from Lake Okeechobee, Florida (USA). Ecotoxicol. Environ. Saf. 2021, 207, 111233. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.A.; Rollwagen-Bollens, G.; Bollens, S.M.; Faber-Hammond, J.J. Environmental influence on cyanobacteria abundance and microcystin toxin production in a shallow temperate lake. Ecotoxicol. Environ. Saf. 2015, 114, 318–325. [Google Scholar] [CrossRef]
- Havens, K.E.; Ji, G.; Beaver, J.R.; Fulton, R.S.; Teacher, C.E. Dynamics of cyanobacteria blooms are linked to the hydrology of shallow Florida lakes and provide insight into possible impacts of climate change. Hydrobiologia 2019, 829, 43–59. [Google Scholar] [CrossRef]
- Hanson, M.J.; Stefan, H.G. Side effects of 58 years of copper sulfate treatment of the Fairmont lakes, Minnesota 1. JAWRA J. Am. Water Resour. Assoc. 1984, 20, 889–900. [Google Scholar] [CrossRef]
- Weenink, E.F.; Luimstra, V.M.; Schuurmans, J.M.; Van Herk, M.J.; Visser, P.M.; Matthijs, H.C. Combatting cyanobacteria with hydrogen peroxide: A laboratory study on the consequences for phytoplankton community and diversity. Front. Microbiol. 2015, 6, 714. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Buley, R.P.; Fernandez-Figueroa, E.G.; Barros, M.U.; Rajendran, S.; Wilson, A.E. Hydrogen peroxide treatment promotes chlorophytes over toxic cyanobacteria in a hyper-eutrophic aquaculture pond. Environ. Pollut. 2018, 240, 590–598. [Google Scholar] [CrossRef] [PubMed]
- Pokrzywinski, K.L.; Bishop, W.M.; Grasso, C.R.; Fernando, B.M.; Sperry, B.P.; Berthold, D.E.; Laughinghouse, H.D., IV; Van Goethem, E.M.; Volk, K.; Heilman, M.; et al. Evaluation of a peroxide-based algaecide for cyanobacteria control: A mesocosm trial in Lake Okeechobee, FL, USA. Water 2022, 14, 169. [Google Scholar] [CrossRef]
- Hehmann, A.; Kaya, K.; Watanabe, M.M. Selective control of Microcystis using an amino acid–a laboratory assay. J. Appl. Phycol. 2002, 14, 85–89. [Google Scholar] [CrossRef]
- Takamura, Y.; Yamada, T.; Kimoto, A.; Kanehama, N.; Tanaka, T.; Nakadaira, S.; Yagi, O. Growth inhibition of Microcystis cyanobacteria by L-lysine and disappearance of natural Microcystis blooms with spraying. Microbes Environ. 2004, 19, 31–39. [Google Scholar] [CrossRef]
- Tian, L.; Chen, M.; Ren, C.; Wang, Y.; Li, L. Anticyanobacterial effect of l-lysine on Microcystis aeruginosa. RSC Adv. 2018, 8, 21606–21612. [Google Scholar] [CrossRef]
- Kaya, K.; Liu, Y.D.; Shen, Y.W.; Xiao, B.D.; Sano, T. Selective control of toxic Microcystis water blooms using lysine and malonic acid: An enclosure experiment. Environ. Toxicol. Int. J. 2005, 20, 170–178. [Google Scholar] [CrossRef]
- Kaneko, T.; Nakajima, N.; Okamoto, S.; Suzuki, I.; Tanabe, Y.; Tamaoki, M.; Nakamura, Y.; Kasai, F.; Watanabe, M.M. Complete genomic structure of the bloom-forming toxic cyanobacterium Microcystis aeruginosa NIES-843. DNA Res. 2007, 14, 247–256. [Google Scholar] [CrossRef]
- Parajuli, A.; Kwak, D.H.; Dalponte, L.; Leikoski, N.; Galica, T.; Umeobika, U.; Trembleau, L.; Bent, A.; Sivonen, K.; Wahlsten, M.; et al. A unique tryptophan C-prenyltransferase from the Kawaguchipeptin biosynthetic pathway. Angew. Chem. Int. Ed. 2016, 55, 3596–3599. [Google Scholar] [CrossRef]
- Tanabe, Y.; Yamaguchi, H.; Sano, T.; Kawachi, M. A novel salt-tolerant genotype illuminates the sucrose gene evolution in freshwater bloom-forming cyanobacterium Microcystis aeruginosa. FEMS Microbiol. Lett. 2019, 366, fnz190. [Google Scholar] [CrossRef]
- Yamaguchi, H.; Suzuki, S.; Osana, Y.; Kawachi, M. Genomic characteristics of the toxic bloom-forming cyanobacterium Microcystis aeruginosa NIES-102. J. Genom. 2020, 8, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M. Isolation, cultivation and classification of bloom-forming Microcystis in Japan. In Toxic Microcystis; Watanabe, M.F., Harada, K., Carmichael, W.W., Fujiki, H., Eds.; CRC Press: Boca Raton, FL, USA, 1996; pp. 13–34. [Google Scholar]
- Otsuka, S.; Suda, S.; Li, R.; Matsumoto, S.; Watanabe, M.M. Morphological variability of colonies of Microcystis morphospecies in culture. J. Gen. Appl. Microbiol. 2000, 46, 39–50. [Google Scholar] [CrossRef] [PubMed]
- Kondo, R.; Yoshida, T.; Yuki, Y.; Hiroishi, S. DNA-DNA reassociation among a bloom-forming cyanobacterial genus, Microcystis. Int. J. Syst. Evol. Microbiol. 2000, 50, 767–770. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Xu, D.; Zhang, T.; Ren, Q.; Xiang, L.; Ning, C.; Zhang, Y.; Gao, R. Comprehensive and functional analyses reveal the genomic diversity and potential toxicity of Microcystis. Harmful Algae 2022, 113, 102186. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014. [Google Scholar]
- Hishinuma, S.; Yuki, M.; Fujimura, M.; Fukumori, F. OxyR regulated the expression of two major catalases, KatA and KatB, along with peroxiredoxin, AhpC in Pseudomonas putida. Environ. Microbiol. 2006, 8, 2115–2124. [Google Scholar] [CrossRef]
- Kim, M.; Shin, B.; Lee, J.; Park, H.Y.; Park, W. Culture-independent and culture-dependent analyses of the bacterial community in the phycosphere of cyanobloom-forming Microcystis aeruginosa. Sci. Rep. 2019, 9, 20416. [Google Scholar] [CrossRef]
- Kim, M.; Kim, W.; Lee, Y.; Park, W. Linkage between bacterial community-mediated hydrogen peroxide detoxification and the growth of Microcystis aeruginosa. Water Res. 2021, 207, 117784. [Google Scholar] [CrossRef]
- Lee, T.M.; Shiu, C.T. Implications of mycosporine-like amino acid and antioxidant defenses in UV-B radiation tolerance for the algae species Ptercladiella capillacea and Gelidium amansii. Mar. Environ. Res. 2009, 67, 8–16. [Google Scholar] [CrossRef]
- Reynolds, C.S. The Ecology of Phytoplankton; Cambridge University Press: Cambridge, UK, 2006. [Google Scholar]
- Beardall, J.; Allen, D.; Bragg, J.; Finkel, Z.V.; Flynn, K.J.; Quigg, A.; Rees, T.A.V.; Richardson, A.J.; Raven, J.A. Allometry and stoichiometry of unicellular, colonial and multicellular phytoplankton. New Phytologist. 2009, 181, 295–309. [Google Scholar] [CrossRef]
- Xiao, M.; Li, M.; Reynolds, C.S. Colony formation in the cyanobacterium Microcystis. Biol. Rev. 2018, 93, 1399–1420. [Google Scholar] [CrossRef]
- Van Le, V.; Srivastava, A.; Ko, S.R.; Ahn, C.Y.; Oh, H.M. Microcystis colony formation: Extracellular polymeric substance, associated microorganisms, and its application. Bioresour. Technol. 2022, 360, 127610. [Google Scholar]
- Gao, L.; Pan, X.; Zhang, D.; Mu, S.; Lee, D.J.; Halik, U. Extracellular polymeric substances buffer against the biocidal effect of H2O2 on the bloom-forming cyanobacterium Microcystis aeruginosa. Water Res. 2015, 69, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.X.; Gan, N.Q.; Huang, Q.; Song, L.R. Response of Microcystis to copper stress–do phenotypes of Microcystis make a difference in stress tolerance? Environ. Pollut. 2007, 147, 324–330. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Shi, X.; Chen, C.; Yu, L.; Sun, C. Responses of Microcystis colonies of different sizes to hydrogen peroxide stress. Toxins 2017, 9, 306. [Google Scholar] [CrossRef]
- Zimba, P.V.; Dionigi, C.P.; Brashear, S.S. Selective toxicity of exogenous l-lysine to cyanobacteria, relative to a chlorophyte and a diatom. Phycologia 2001, 40, 483–486. [Google Scholar] [CrossRef]
- Kim, W.; Kim, M.; Park, W. Unlocking the mystery of lysine toxicity on Microcystis aeruginosa. J. Hazard. Mater. 2023, 448, 130932. [Google Scholar] [CrossRef]
- Yan, X.; Zhao, F.; Wang, G.; Wang, Z.; Zhou, M.; Zhang, L.; Wang, G.; Chen, Y. Metabolomic analysis of Microcystis aeruginosa after exposure to the algicide L-lysine. Bull. Environ. Contam. Toxicol. 2023, 110, 12. [Google Scholar] [CrossRef]
- Lürling, M.; Van Oosterhout, F. Effect of selected plant extracts and D-and L-lysine on the cyanobacterium Microcystis aeruginosa. Water 2014, 6, 1807–1825. [Google Scholar] [CrossRef]
- Jančula, D.; Maršálek, B. Seven years from the first application of polyaluminium chloride in the Czech Republic–effects on phytoplankton communities in three water bodies. Chem. Ecol. 2012, 28, 535–544. [Google Scholar] [CrossRef]
- Gensemer, R.W.; Playle, R.C. The bioavailability and toxicity of aluminum in aquatic environments. Crit. Rev. Environ. Sci. Technol. 1999, 29, 315–450. [Google Scholar] [CrossRef]
- Smeltzer, E. A successful alum/aluminate treatment of Lake Morey, Vermont. Lake Reserv. Manag. 1990, 6, 9–19. [Google Scholar] [CrossRef]
- Reitzel, K.; Jensen, H.S.; Egemose, S. pH dependent dissolution of sediment aluminum in six Danish lakes treated with aluminum. Water Res. 2013, 47, 1409–1420. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Song, Q.; Long, J.; Song, G.; Mi, W.; Bi, Y. Optimization method for Microcystis bloom mitigation by hydrogen peroxide and its stimulative effects on growth of chlorophytes. Chemosphere 2019, 228, 503–512. [Google Scholar] [CrossRef] [PubMed]
- Aragão, M.C.; Dos Reis, K.C.; Rocha, M.A.M.; de Oliveira Guedes, D.; Dos Santos, E.C.; Capelo-Neto, J. Removal of Dolichospermum circinale, Microcystis aeruginosa, and their metabolites using hydrogen peroxide and visible light. Aquat. Toxicol. 2021, 232, 105735. [Google Scholar] [CrossRef] [PubMed]
- Urakawa, H.; Steele, J.H.; Hancock, T.L.; Dahedl, E.K.; Schroeder, E.R.; Sereda, J.V.; Kratz, M.A.; García, P.E.; Armstrong, R.A. Interaction among spring phytoplankton succession, water discharge patterns, and hydrogen peroxide dynamics in the Caloosahatchee River in southwest Florida. Harmful Algae 2023, 126, 102434. [Google Scholar] [CrossRef]
- Lusty, M.W.; Gobler, C.J. The efficacy of hydrogen peroxide in mitigating cyanobacterial blooms and altering microbial communities across four lakes in NY, USA. Toxins 2020, 12, 428. [Google Scholar] [CrossRef]
- Shao, J.; Li, R.; Lepo, J.E.; Gu, J.D. Potential for control of harmful cyanobacterial blooms using biologically derived substances: Problems and prospects. J. Environ. Manag. 2013, 125, 149–155. [Google Scholar] [CrossRef]
- Murrell, M.C.; Lores, E.M. Phytoplankton and zooplankton seasonal dynamics in a subtropical estuary: Importance of cyanobacteria. J. Plankton Res. 2004, 26, 371–382. [Google Scholar] [CrossRef]
- Jardillier, L.; Zubkov, M.V.; Pearman, J.; Scanlan, D.J. Significant CO2 fixation by small prymnesiophytes in the subtropical and tropical northeast Atlantic Ocean. ISME J. 2010, 4, 1180–1192. [Google Scholar] [CrossRef]
- Matthijs, H.C.; Visser, P.M.; Reeze, B.; Meeuse, J.; Slot, P.C.; Wijn, G.; Talens, R.; Huisman, J. Selective suppression of harmful cyanobacteria in an entire lake with hydrogen peroxide. Water Res. 2012, 46, 1460–1472. [Google Scholar] [CrossRef]
- Weenink, E.F.; Matthijs, H.C.; Schuurmans, J.M.; Piel, T.; van Herk, M.J.; Sigon, C.A.; Visser, P.M.; Huisman, J. Interspecific protection against oxidative stress: Green algae protect harmful cyanobacteria against hydrogen peroxide. Environ. Microbiol. 2021, 23, 2404–2419. [Google Scholar] [CrossRef] [PubMed]
- Buley, R.P.; Adams, C.; Belfiore, A.P.; Fernandez-Figueroa, E.G.; Gladfelter, M.F.; Garner, B.; Wilson, A.E. Field evaluation of seven products to control cyanobacterial blooms in aquaculture. Environ. Sci. Pollut. Res. 2021, 28, 29971–29983. [Google Scholar] [CrossRef] [PubMed]
- Barrington, D.J.; Reichwaldt, E.S.; Ghadouani, A. The use of hydrogen peroxide to remove cyanobacteria and microcystins from waste stabilization ponds and hypereutrophic systems. Ecol. Eng. 2013, 50, 86–94. [Google Scholar] [CrossRef]
- Barrington, D.J.; Ghadouani, A.; Ivey, G.N. Environmental factors and the application of hydrogen peroxide for the removal of toxic cyanobacteria from waste stabilization ponds. J. Environ. Eng. 2011, 137, 952–960. [Google Scholar] [CrossRef]
- Dai, R.; Liu, H.; Qu, J.; Zhao, X.; Hou, Y. Effects of amino acids on microcystin production of the Microcystis aeruginosa. J. Hazard. Mater. 2009, 161, 730–736. [Google Scholar] [CrossRef]
- Chen, C.; Wang, Y.; Chen, K.; Shi, X.; Yang, G. Using hydrogen peroxide to control cyanobacterial blooms: A mesocosm study focused on the effects of algal density in Lake Chaohu, China. Environ. Pollut. 2021, 272, 115923. [Google Scholar] [CrossRef]
- Qian, H.; Yu, S.; Sun, Z.; Xie, X.; Liu, W.; Fu, Z. Effects of copper sulfate, hydrogen peroxide and N-phenyl-2-naphthylamine on oxidative stress and the expression of genes involved photosynthesis and microcystin disposition in Microcystis aeruginosa. Aquat. Toxicol. 2010, 99, 405–412. [Google Scholar] [CrossRef]
- Huo, X.; Chang, D.W.; Tseng, J.H.; Burch, M.D.; Lin, T.F. Exposure of Microcystis aeruginosa to hydrogen peroxide under light: Kinetic modeling of cell rupture and simultaneous microcystin degradation. Environ. Sci. Technol. 2015, 49, 5502–5510. [Google Scholar] [CrossRef]
- Matthijs, H.C.; Jančula, D.; Visser, P.M.; Maršálek, B. Existing and emerging cyanocidal compounds: New perspectives for cyanobacterial bloom mitigation. Aquat. Ecol. 2016, 50, 443–460. [Google Scholar] [CrossRef]
- Clearwater, S.J.; Jellyman, P.G.; Biggs, B.J.; Hickey, C.W.; Blair, N.; Clayton, J.S. Pulse-dose application of chelated copper to a river for Didymosphenia geminata control: Effects on macroinvertebrates and fish. Environ. Toxicol. Chem. 2011, 30, 181–195. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).