
The Effects of Roundup™ on Benthic Microbial Assemblages


Abstract
1. Introduction
2. Materials and Methods
2.1. Winogradsky Column Construction
2.2. Microbe and Water Chemistry Sampling
2.3. Statistical Analyses
3. Results
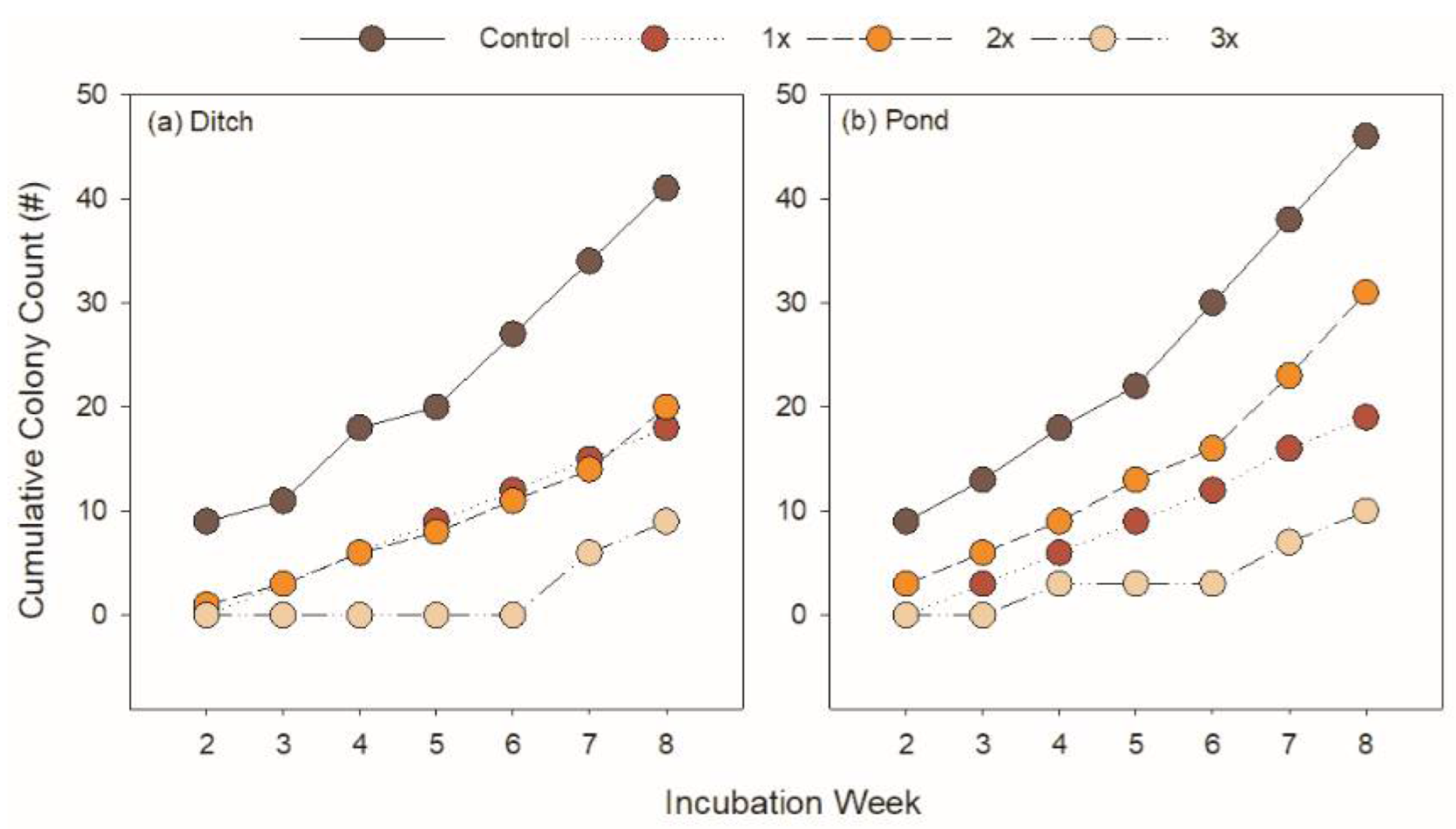
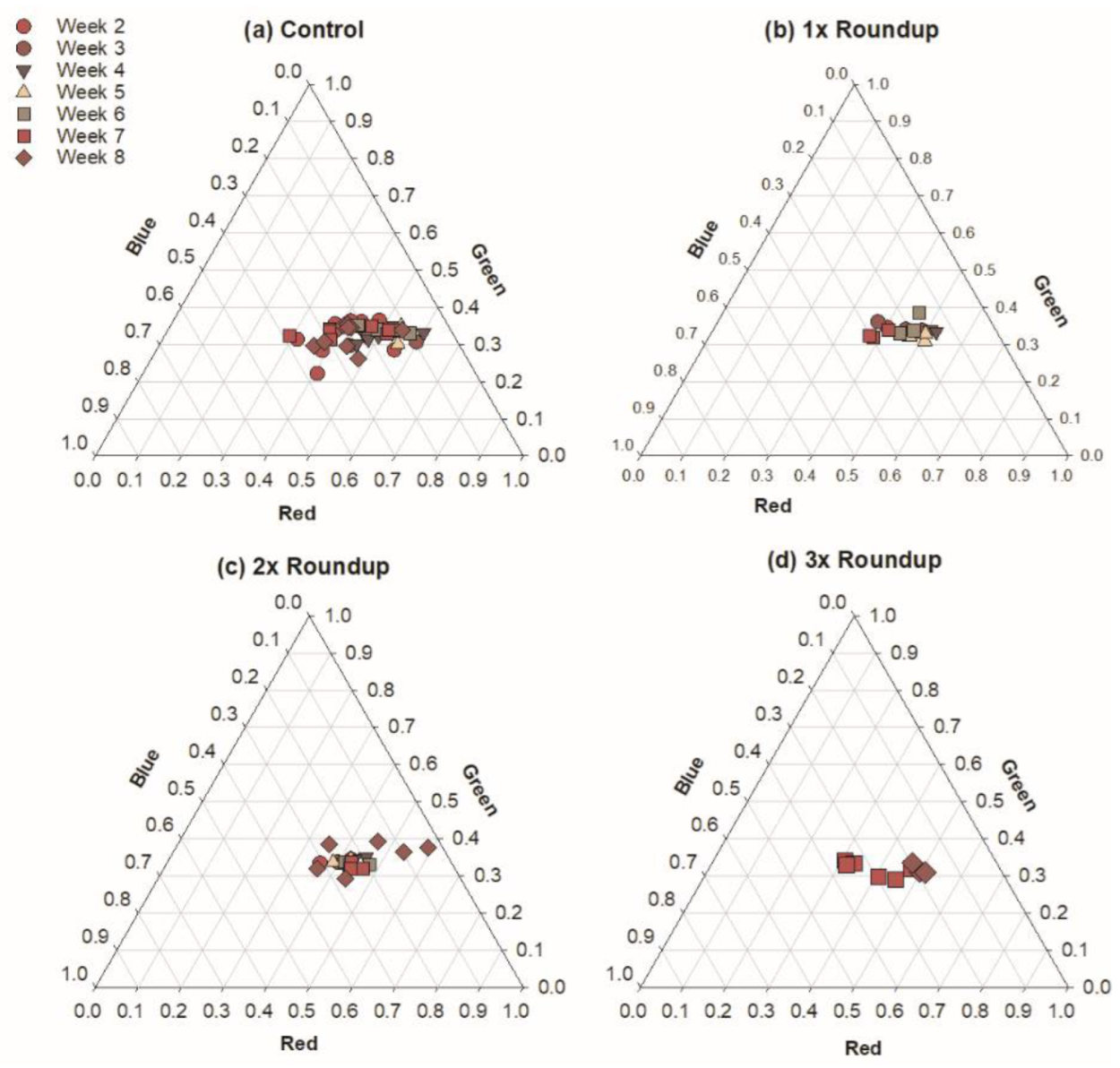
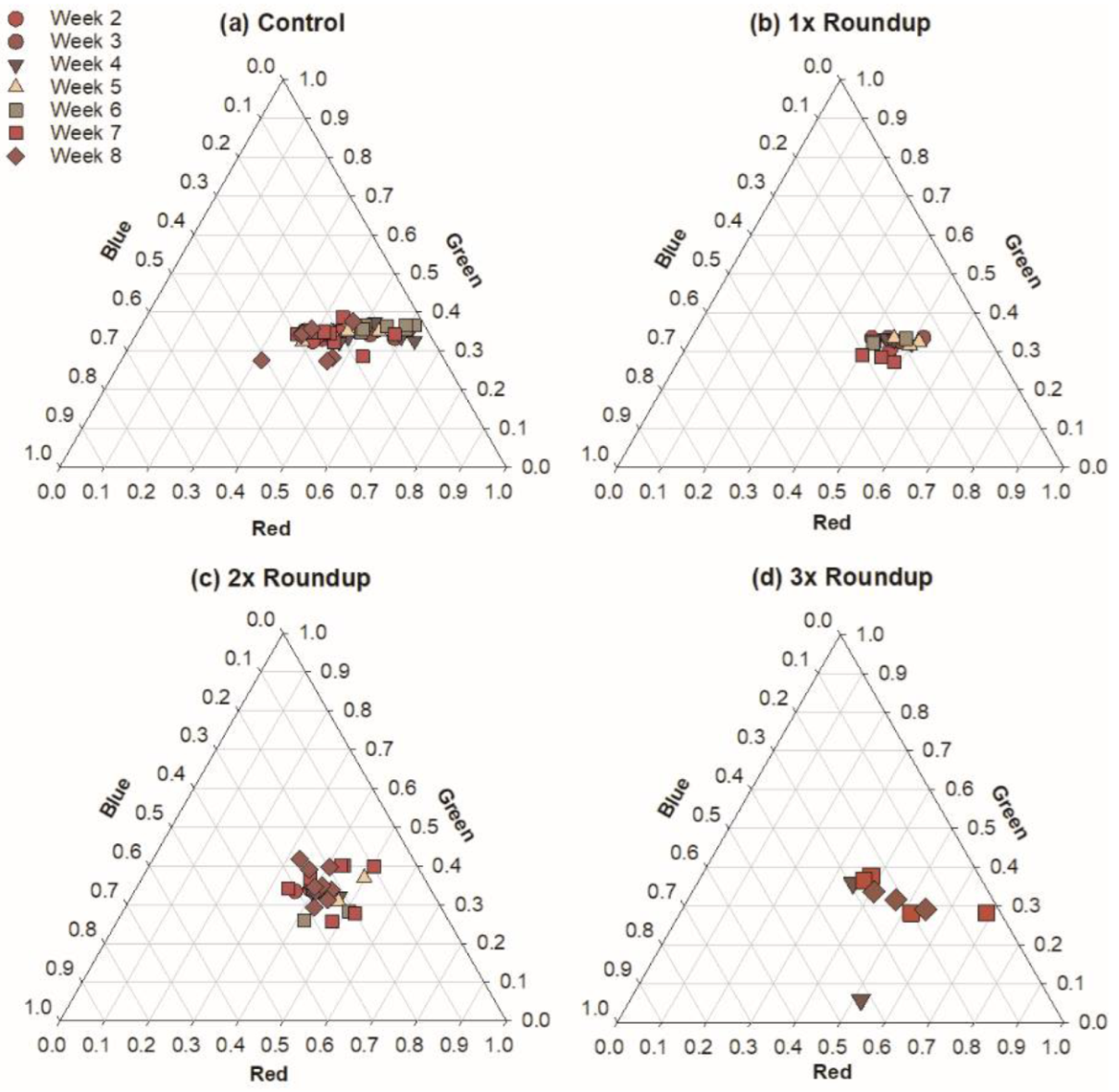
3.1. Roundup Reduces Microbial Productivity
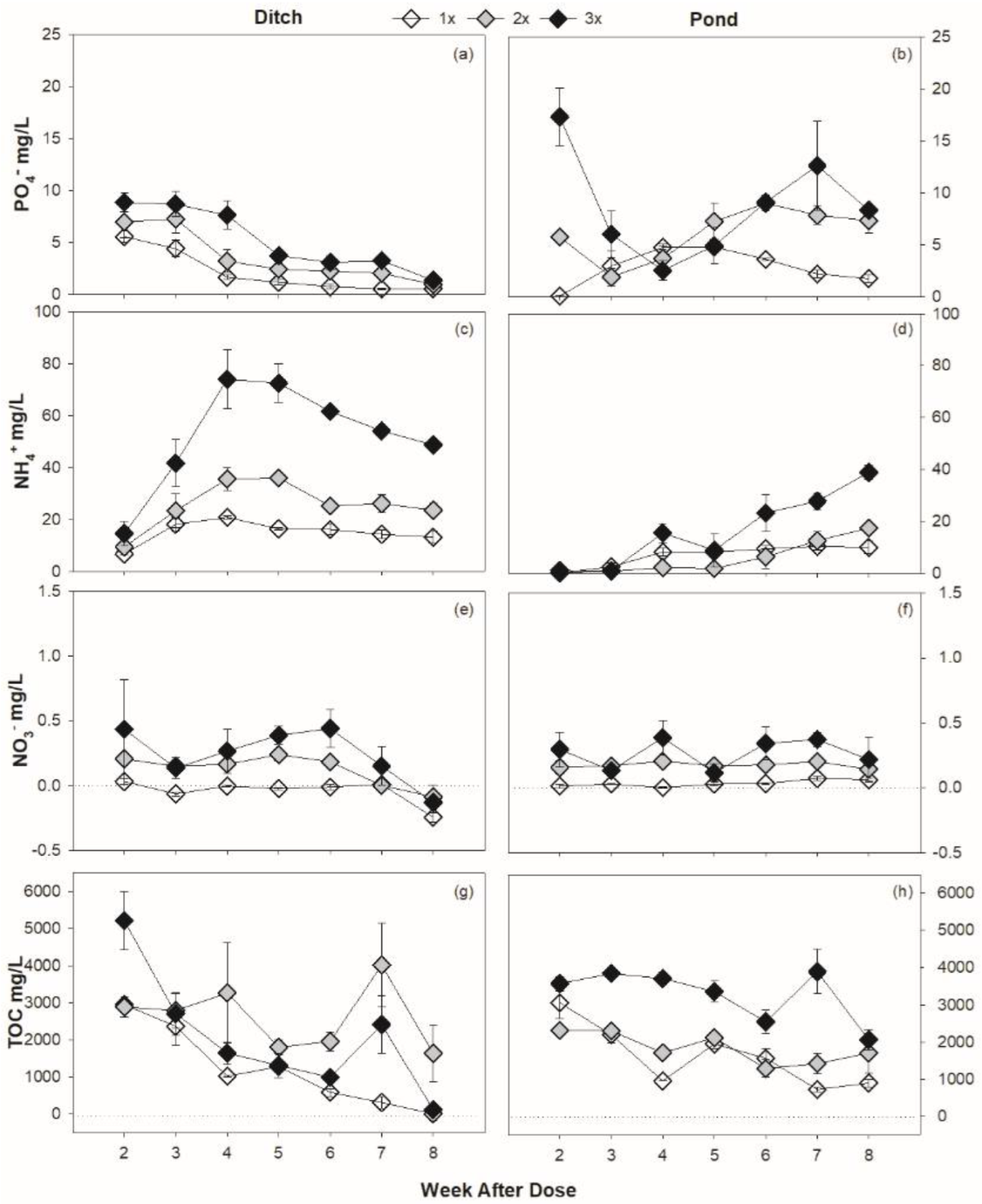
3.2. Roundup Serves as Nutrient Source
3.3. DOC Trends Vary by Habitat
4. Discussion
4.1. Roundup Harms Photosynthetic Microbes
4.2. Roundup Is a Source of Nutrients
4.3. Roundup Alters Carbon Availability
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Vanni, M.J. Nutrient cycling by animals in freshwater ecosystems. Annu. Rev. Ecol. Syst. 2002, 33, 341–370. [Google Scholar] [CrossRef]
- Bormann, F.H.; Likens, G.E. The Nutrient Cycles of an Ecosystem. Sci. Am. 1970, 223, 92–101. [Google Scholar] [CrossRef]
- Arrigo, K.R. Marine microorganisms and global nutrient cycles. Nature 2005, 437, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Wetzel, R.G. Limnology: Lake and River Ecosystems, 3rd ed.; Academic Press: San Diego, CA, USA, 2001. [Google Scholar]
- Tian, J.; Ge, F.; Zhang, D.; Deng, S.; Liu, X. Roles of phosphate solubilizing microorganisms from managing soil phosphorus deficiency to mediating biogeochemical P cycle. Biology 2021, 10, 158. [Google Scholar] [CrossRef] [PubMed]
- Ricklefs, R.E.; Miller, G.L. Ecology, 4th ed.; W. H. Freeman and Company: New York, NY, USA, 2000. [Google Scholar]
- Sim, J.X.; Drigo, B.; Doolette, C.L.; Vasileiadis, S.; Karpouzas, D.G.; Lombi, E. Impact of twenty pesticides on soil carbon microbial functions and community composition. Chemosphere 2022, 307, 135820. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Wu, L.; Liu, Y.; Zhang, P.; Li, Q.; Zhou, J.; Hess, N.J.; Hazen, T.C.; Yang, W.; Chakraborty, R. Microbial interactions with dissolved organic matter drive carbon dynamics and community succession. Front. Microbiol. 2018, 9, 1234. [Google Scholar] [CrossRef]
- Berggren, M.; Laudon, H.; Jansson, M. Aging of allochthonous organic carbon regulates bacterial production in unproductive boreal lakes. Limnol. Oceanogr. 2009, 54, 1333–1342. [Google Scholar] [CrossRef]
- Sillanpää, M.; Matilanien, A.; Lahtinen, T. Characterization of NOM. In Natural Organic Matter in Water, 1st ed.; IWA Publishing: London, UK, 2014. [Google Scholar]
- Grasset, C.; Rodriguez, C.; Delolme, C.; Marmonier, P.; Bornette, G. Can soil organic carbon fractions be used as functional indicators of wetlands? Wetlands 2017, 37, 1195–1205. [Google Scholar] [CrossRef]
- Babcsányi, I.; Meite, F.; Imfeld, G. Biogeochemical gradients and microbial communities in Winogradsky columns established with polluted wetland sediments. FEMS Microbiol. 2017, 93, fix089. [Google Scholar] [CrossRef]
- Rundell, E.A.; Banta, L.M.; Ward, D.V.; Watts, C.D.; Birren, B.; Esteban, D.J. 16S rRNA gene survey of microbial communities in Winogradsky columns. PLoS ONE 2014, 9, e104134. [Google Scholar] [CrossRef]
- Lu, T.; Xu, N.; Zhang, Q.; Zhang, Z.; Debognies, A.; Zhou, Z.; Sun, L.; Qian, H. Understanding the influence of glyphosate on the structure and function of freshwater microbial community in a microcosm. Environ. Pollut. 2020, 260, 114012. [Google Scholar] [CrossRef] [PubMed]
- Perez, G.L.; Vera, M.S.; Miranda, L.A. Effects of herbicide glyphosate and glyphosate-based formulations on aquatic ecosystems. In Herbicides and Environment, 1st ed.; Kortekamp, A., Ed.; InTech: New York, NY, USA; pp. 343–368.
- Sang, Y.; Mejuto, J.C.; Xiao, J.; Simal-Gandara, J. Assessment of glyphosate impact on the agrifood ecosystem. Plants 2021, 10, 405. [Google Scholar] [CrossRef] [PubMed]
- Helander, M.; Saloniemi, I.; Saikkonen, K. Glyphosate in northern ecosystems. Trends Plant Sci. 2012, 10, 569–574. [Google Scholar] [CrossRef] [PubMed]
- Kanissery, R.; Gairhe, B.; Kadyampakeni, D.; Batuman, O.; Alferez, F. Glyphosate: Its environmental persistence and impact on crop health and nutrition. Plants 2019, 8, 499. [Google Scholar] [CrossRef] [PubMed]
- Estok, D.; Freedman, B.; Boyle, D. Effects of the herbicides 2,4-D, glyphosate, hexazinone, and triclopyr on the growth of three species of ectomycorrhizal fungi. Bull. Environ. Contam. Toxicol. 1989, 42, 835–839. [Google Scholar] [CrossRef] [PubMed]
- Newman, M.M.; Hoilett, N.; Lorenz, N.; Dick, R.P.; Liles, M.R.; Ramsier, C.; Kloepper, J.W. Glyphosate effects on soil rhizosphere-associated bacterial communities. Sci. Total Environ. 2016, 543, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Shehata, A.A.; Schrödl, W.; Aldin, A.A.; Hafez, H.M.; Krüger, M. The effect of glyphosate on potential pathogens and beneficial members of poultry microbiota in vitro. Curr. Microbiol. 2013, 66, 350–358. [Google Scholar] [CrossRef]
- Kittle, R.P.; McDermid, K.J. Glyphosate herbicide toxicity to native Hawaiian macroalgal and seagrass species. J. Appl. Phycol. 2016, 28, 2597–2604. [Google Scholar] [CrossRef]
- Correia, F.V.; Moreira, J.C. Effects of glyphosate and 2, 4-D on earthworms (Eisenia foetida) in laboratory tests. Bull. Environ. Contam. Toxicol. 2010, 85, 264–268. [Google Scholar] [CrossRef]
- Zaller, J.G.; Heigl, F.; Ruess, L.; Grabmaier, A. Glyphosate herbicide affects belowground interactions between earthworms and symbiotic mycorrhizal fungi in a model ecosystem. Sci. Rep. 2014, 4, 5634. [Google Scholar] [CrossRef]
- Piola, L.; Fuchs, J.; Oneto, M.L.; Basack, S.; Kesten, E.; Casabe, N. Comparative toxicity of two glyphosate-based formulations to Eisenia andrei under laboratory conditions. Chemosphere 2013, 91, 545–551. [Google Scholar] [CrossRef] [PubMed]
- Richard, S.; Moslemi, S.; Sipahutar, H.; Benachour, N.; Seralini, G.E. Differential effects of glyphosate and roundup on human placental cells and aromatase. Environ. Health Perspect. 2005, 113, 716. [Google Scholar] [CrossRef] [PubMed]
- Koller, V.J.; Fürhacker, M.; Nersesyan, A.; Misik, M.; Eisenbauer, M.; Knasmueller, S. Cytotoxic and DNA-damaging properties of glyphosate and Roundup in human-derived buccal epithelial cells. Arch. Toxicol. 2012, 86, 805–813. [Google Scholar] [CrossRef] [PubMed]
- Martini, C.N.; Gabrielli, M.; Codesido, M.M.; Del Vila, M.C. Glyphosate-based herbicides with different adjuvants are more potent inhibitors of 3T3-L1 fibroblast proliferation and differentiation to adipocytes than glyphosate alone. Comp. Clin. Path. 2016, 25, 607–613. [Google Scholar] [CrossRef]
- Gill, J.P.K.; Sethi, N.; Mohan, A.; Datta, S.; Girdhar, M. Glyphosate toxicity for animals. Environ. Chem. Lett. 2018, 16, 401–426. [Google Scholar] [CrossRef]
- Glyphosate | Ingredients Used in Pesticide Products | US EPA. Available online: https://www.epa.gov/ingredients-used-pesticide-products/glyphosate (accessed on 4 November 2020).
- Duke, S.O.; Powles, S.B. Glyphosate: A once-in-a-century herbicide. Pest. Manag. Sci. 2008, 64, 319–325. [Google Scholar] [CrossRef]
- Judd, K.E.; Francoeur, S.N. Short-term impacts of Phragmites management on nutrient budgets and plant communities in Great Lakes coastal freshwater marshes. Wetl. Ecol. Manag. 2019, 27, 55–74. [Google Scholar] [CrossRef]
- Native-Land. 2020. Available online: https://native-land.ca/ (accessed on 12 December 2020).
- Higgins, B.A.; Puglia, P.S.; Leonard, R.P.; Yaokum, T.D.; Wirtz, W.A. Soil Survey of Niagara County, New York; United States Department of Agriculture: Washington, DC, USA, 1972; p. 208. [Google Scholar]
- Hansen, T.E. The Winogradsky Column: An Enclosed Self Sustaining Microbial Ecosystem. Available online: https://sites.udel.edu/winogradsky/the-columns/build-a-column/ (accessed on 20 October 2020).
- Soil Science: Make a Winogradsky Column. Available online: https://www.scientificamerican.com/article/bring-science-home-soil-column/?gclid=Cj0KCQjwkOqZBhDNARIsAACsbfJW4W7_anh5eKgy2Awh0SIlqGM9Imcu__4jO-zXnkEuLALLnrVQjZ0aAvAxEALw_wcB (accessed on 20 October 2020).
- Tsui, M.T.K.; Chu, L.M. Environmental fate and non-target impact of glyphosate-based herbicide (Roundup®) in a subtropical wetland. Chemosphere 2008, 71, 439–446. [Google Scholar] [CrossRef]
- Baird, R.; Eaton, A.D.; Rice, E. Standard Methods for The Examination of Water and Wastewater, 21st ed.; Amer Public Health Assn: Washington, DC, USA, 2005. [Google Scholar]
- Albrektiene, R.; Rimeika, M.; Salieckiene, E.; Šaulys, V.; Zagorskis, A. Determination of organic matter by UV absorption in the groundwater. J. Environ. Eng. Landsc. 2012, 20, 163–167. [Google Scholar]
- Saxton, M.; Morrow, E.A.; Bouronniere, R.A.; Wilhelm, S.W. Glyphosate influence on phytoplankton community structure in Lake Erie. J. Great Lakes Res. 2011, 37, 683–690. [Google Scholar] [CrossRef]
- Bonansea, R.I.; Filippi, I.; Wunderlin, D.A.; Marino, D.J.G.; Ame, M.V. The fate of glyphosate and AMPA in a freshwater endorheic basin: An ecotoxicological risk assessment. Toxics 2018, 6, 3. [Google Scholar] [CrossRef] [PubMed]
- Imparato, V.; Santos, S.S.; Johansen, A.; Geisen, S. Simulation of bacteria and protists in rhizosphere of glyphosate-treated barley. Appl. Soil Ecol. 2016, 98, 47–55. [Google Scholar] [CrossRef]
- Matiwalage, I.N.; Rajapaksha, R.M.C.P. Toxic effects of paraquat and glyphosate on bacteria in wetland rice soil. J. Soil Sci. Soc. Sri Lanka 2008, 20, 9–16. [Google Scholar]
- Tahir, S.; Marschner, P. Clay addition to sandy soil-effect of clay concentration and ped size on microbial biomass and nutrient dynamics after addition of low C/N ratio residue. J. Plant Nutr. Soil Sci. 2016, 16, 864–875. [Google Scholar] [CrossRef]
- Pal, S.; Marschner, P. Influence of clay concentration, residue C/N and particle size on microbial activity and nutrient availability in clay amended sandy soil. J. Soil Sci. Plant Nutr. 2016, 16, 350–361. [Google Scholar] [CrossRef]
- Don, A.; Schulze, E.D. Controls on fluxes and export of dissolved organic carbon in grasslands with contrasting soil types. Biogeochemistry 2008, 91, 117–131. [Google Scholar] [CrossRef]
- Kaiser, K.; Zech, W. Dissolved organic matter absorption by mineral constituents of subsoil clay fractions. J. Plant Nutr. Soil Sci. 2000, 163, 531–535. [Google Scholar] [CrossRef]
- Ren, L.X.; Wang, P.F.; Wang, C.; Chen, J.; Hou, J.; Qian, J. Algal growth and utilization of phosphorus studied by combined mono-culture and co-culture experiments. Environ. Pollut. 2017, 220, 274–285. [Google Scholar] [CrossRef]
- Alshameri, A.; He, H.; Zhu, J.; Xi, Y.; Zhu, R.; Ma, L.; Tao, Q. Adsorption of ammonium by different natural clay minerals: Characterization, kinetics, and adsorption isotherms. Appl. Clay Sci. 2018, 159, 83–93. [Google Scholar] [CrossRef]
- Yu, W.H.; Li, N.; Tong, D.S.; Zhou, C.H.; Lin, C.W.; Xu, C.Y. Adsorption of proteins and nucleic acids on clay minerals and their interactions: A review. Appl. Clay Sci. 2013, 80, 443–452. [Google Scholar] [CrossRef]
- Grim, R.; Asllaway, W.; Cuthbert, F. Reaction of different clay minerals with some organic cations. J. Am. Ceram. Soc. 1947, 30, 137–142. [Google Scholar] [CrossRef]
- Spohn, M.; Kuzyakov, Y. Phosphorus mineralization can be driven by microbial need for carbon. Soil Biol. Biochem. 2013, 63, 69–75. [Google Scholar] [CrossRef]
- Wang, G.; Liu, J.; Tang, J. The long-term nutrient accumulation with respect to anthropogenic impacts in the sediments from two freshwater marshes (Xianghai Wetlands, Northeast China). Water Res. 2004, 38, 4462–4474. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Habitat | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Treatment | Week | Ditch | Pond | ||||||
| PO43− mg/L | NH4+ mg/L | NO3− mg/L | DOC mg/L | PO43− mg/L | NH4+ mg/L | NO3− mg/L | DOC mg/L | ||
| Control | 2 | 0.01 (0.005) | 0.91 (0.11) | 0.02 (0.002) | 501.5 (23.0) | 0.02 (0.001) | 0.06 (0.02) | 0.04 (0.01) | 304.4 (16.0) |
| 3 | 0.03 (0.008) | 0.82 (0.23) | 0.09 (0.03) | 758.1 (33.5) | 0.01 (0.001) | 0.08 (0.01) | 0.04 (0.01) | 388.1 (16.6) | |
| 4 | 0.03 (0.004) | 1.14 (0.36) | 0.05 (0.02) | 786.3 (63.6) | 0.01 (0.001) | 0.20 (0.04) | 0.03 (0.01) | 454.1 (37.0) | |
| 5 | 0.03 (0.003) | 1.61 (0.31) | 0.04 (0.01) | 1100.7 (41.8) | 0.01 (0.001) | 0.26 (0.07) | 0.04 (0.01) | 411.9 (61.9) | |
| 6 | 0.03 (0.004) | 1.67 (0.52) | 0.07 (0.03) | 1147.4 (24.6) | 0.01 (0.001) | 0.19 (0.08) | 0.04 (0.01) | 450.4 (32.6) | |
| 7 | 0.04 (0.005) | 1.60 (0.66) | 0.16 (0.07) | 1309.6 (52.7) | 0.01 (0.001) | 0.13 (0.05) | 0.04 (0.004) | 500.0 (39.0) | |
| 8 | 0.10 (0.020) | 0.64 (0.13) | 0.32 (0.05) | 1399.3 (59.3) | 0.01 (0.001) | 0.13 (0.04) | 0.05 (0.01) | 424.4 (10.6) | |
| 1x | 2 | 5.58 (0.55) | 7.60 (0.45) | 0.06 (0.02) | 3455.6 (61.2) | 0.07 (0.06) | 0.32 (0.08) | 0.05 (0.01) | 3363.0 (431.4) |
| 3 | 4.45 (0.79) | 19.01 (2.68) | 0.03 (0.01) | 3129.6 (524.7) | 2.96 (1.40) | 2.71 (1.34) | 0.07 (0.01) | 2588.9 (222.3) | |
| 4 | 1.69 (0.25) | 21.98 (0.63) | 0.05 (0.01) | 1811.1 (17.0) | 4.75 (0.18) | 8.48 (1.59) | 0.03 (0.01) | 1407.8 (6.2) | |
| 5 | 1.22 (0.28) | 18.10 (0.51) | 0.02 (0.01) | 2381.5 (91.0) | 4.79 (0.27) | 8.57 (0.54) | 0.07 (0.01) | 2360.0 (123.6) | |
| 6 | 0.80 (0.21) | 17.88 (2.09) | 0.06 (0.02) | 1737.0 (106.3) | 3.60 (0.10) | 9.66 (1.16) | 0.07 (0.01) | 2014.4 (261.0) | |
| 7 | 0.58 (0.10) | 15.93 (1.55) | 0.17 (0.03) | 1625.9 (163.8) | 2.20 (0.40) | 10.63 (1.43) | 0.12 (0.02) | 1224.4 (67.7) | |
| 8 | 0.66 (0.10) | 13.87 (2.40) | 0.08 (0.04) | 1411.1 (203.8) | 1.74 (0.37) | 10.21 (2.36) | 0.11 (0.01) | 1315.6 (103.3) | |
| 2x | 2 | 7.01 (1.51) | 10.40 (1.08) | 0.23 (0.02) | 3392.6 (268.5) | 5.75 (0.13) | 0.38 (0.13) | 0.19 (0.05) | 2622.2 (130.2) |
| 3 | 7.31 (1.37) | 24.26 (6.52) | 0.24 (0.02) | 3551.9 (463.4) | 1.90 (0.85) | 0.91 (0.48) | 0.21 (0.01) | 2688.9 (52.5) | |
| 4 | 3.24 (1.10) | 36.80 (4.44) | 0.22 (0.04) | 4055.6 (1356.1) | 3.67 (1.40) | 2.55 (1.23) | 0.23 (0.01) | 2163.0 (129.9) | |
| 5 | 2.47 (0.53) | 37.63 (1.09) | 0.28 (0.05) | 2907.4 (198.8) | 7.25 (1.74) | 2.17 (1.56) | 0.21 (0.02) | 2528.9 (29.4) | |
| 6 | 2.27 (0.58) | 26.96 (1.60) | 0.26 (0.01) | 3111.1 (258.8) | 8.99 (0.76) | 6.62 (4.43) | 0.22 (0.03) | 1735.6 (235.2) | |
| 7 | 2.11 (0.45) | 27.79 (3.38) | 0.16 (0.01) | 5329.6 (1118.4) | 7.86 (0.92) | 12.79 (3.62) | 0.25 (0.03) | 1917.8 (277.7) | |
| 8 | 1.08 (0.18) | 24.27 (2.58) | 0.23 (0.09) | 3040.7 (773.0) | 7.33 (1.16) | 17.64 (2.36) | 0.19 (0.05) | 2126.7 (527.0) | |
| 3x | 2 | 8.90 (0.88) | 15.53 (4.82) | 0.46 (0.38) | 5707.4 (787.2) | 17.32 (2.78) | 1.26 (0.24) | 0.33 (0.13) | 3877.8 (192.8) |
| 3 | 8.74 (1.21) | 42.54 (9.15) | 0.22 (0.08) | 3470.4 (572.5) | 6.01 (2.25) | 1.11 (0.78) | 0.17 (0.02) | 4237.0 (93.5) | |
| 4 | 7.67 (1.36) | 75.21 (11.32) | 0.32 (0.17) | 2437.0 (296.6) | 2.52 (0.95) | 15.79 (3.74) | 0.42 (0.13) | 4166.7 (161.1) | |
| 5 | 3.78 (0.33) | 74.15 (7.44) | 0.43 (0.07) | 2414.8 (326.5) | 4.92 (1.78) | 9.18 (6.27) | 0.16 (0.07) | 3774.1 (293.8) | |
| 6 | 3.14 (0.66) | 63.35 (0.83) | 0.51 (0.14) | 2144.4 (142.9) | 9.09 (0.43) | 23.53 (7.15) | 0.38 (0.13) | 2996.3 (309.8) | |
| 7 | 3.31 (0.26) | 55.75 (1.89) | 0.31 (0.15) | 3725.9 (768.2) | 12.62 (4.32) | 28.00 (3.07) | 0.42 (0.06) | 4396.3 (595.4) | |
| 8 | 1.47 (0.19) | 49.39 (1.69) | 0.19 (0.03) | 1522.2 (17.0) | 8.33 (0.45) | 39.08 (2.78) | 0.27 (0.17) | 2485.2 (276.4) | |
| Comparison | |||||||
|---|---|---|---|---|---|---|---|
| T | H | T × H | W | W × T | W × H | W × T × H | |
| df | 3.16 | 1.16 | 3.16 | 6.96 | 18.96 | 6.96 | 18.96 |
| PO43− mg/L | 48.58, <0.01 * | 14.54, 0.002 * | 3.31, 0.047 * | 9.76, <0.01 * | 5.87, <0.01 * | 15.82, <0.01 * | 8.26, <0.01 * |
| NH4+ mg/L | 171.65, <0.01 * | 236.48, <0.01 * | 49.79, <0.01 * | 29.68, <0.01 * | 8.21, <0.01 * | 16.75, <0.01 * | 4.86, <0.01 * |
| NO3− mg/L | 27.24, <0.01 * | 1.53, 0.23 | 0.49, 0.69 | 0.80, 0.57 | 1.13, 0.33 | 0.31, 0.93 | 0.83, 0.67 |
| DOC mg/L | 158.07, <0.01 * | 15.93, 0.001 * | 20.17, <0.01 * | 9.61, <0.01 * | 4.87, <0.01 * | 1.82, 0.10 | 2.41, 0.003 * |
| Ditch | Pond | |||||
|---|---|---|---|---|---|---|
| Treatment | PO43− mg/L | NH4+ mg/L | NO3− mg/L | PO43− mg/L | NH4+ mg/L | NO3− mg/L |
| Overall | 0.61, <0.01 * | 0.23, 0.11 | 0.30, 0.02 * | 0.57, <0.01 * | 0.30, 0.02 * | 0.55, <0.01 * |
| Control | 0.70, <0.01 * | 0.24, 0.83 | 0.63, 0.01 * | −0.62, 0.01 * | 0.36, 0.33 | 0.01, 1.00 |
| 1x | 0.88, <0.01 * | −0.43, 0.16 | −0.34, 0.38 | −0.36, 0.34 | −0.75, <0.01 * | −0.36, 0.32 |
| 2x | −0.02, 1.00 | −0.18, 1.00 | −0.37, 0.29 | −0.24, 0.90 | −0.61, 0.01 * | −0.51, 0.06 |
| 3x | 0.58, 0.02 * | −0.70, <0.01 * | 0.19, 1.00 | 0.01, 1.00 | −0.54, 0.03 * | −0.01, 1.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weatherley, S.P.; Laird, H.K.; Gatley-Montross, C.M.; Whorley, S.B. The Effects of Roundup™ on Benthic Microbial Assemblages. Ecologies 2022, 3, 557-569. https://doi.org/10.3390/ecologies3040041
Weatherley SP, Laird HK, Gatley-Montross CM, Whorley SB. The Effects of Roundup™ on Benthic Microbial Assemblages. Ecologies. 2022; 3(4):557-569. https://doi.org/10.3390/ecologies3040041
Chicago/Turabian StyleWeatherley, Shannon P., Hannah K. Laird, Caitlyn M. Gatley-Montross, and Sarah B. Whorley. 2022. "The Effects of Roundup™ on Benthic Microbial Assemblages" Ecologies 3, no. 4: 557-569. https://doi.org/10.3390/ecologies3040041
APA StyleWeatherley, S. P., Laird, H. K., Gatley-Montross, C. M., & Whorley, S. B. (2022). The Effects of Roundup™ on Benthic Microbial Assemblages. Ecologies, 3(4), 557-569. https://doi.org/10.3390/ecologies3040041