
Behind the Scenes of PluriZyme Designs


Abstract
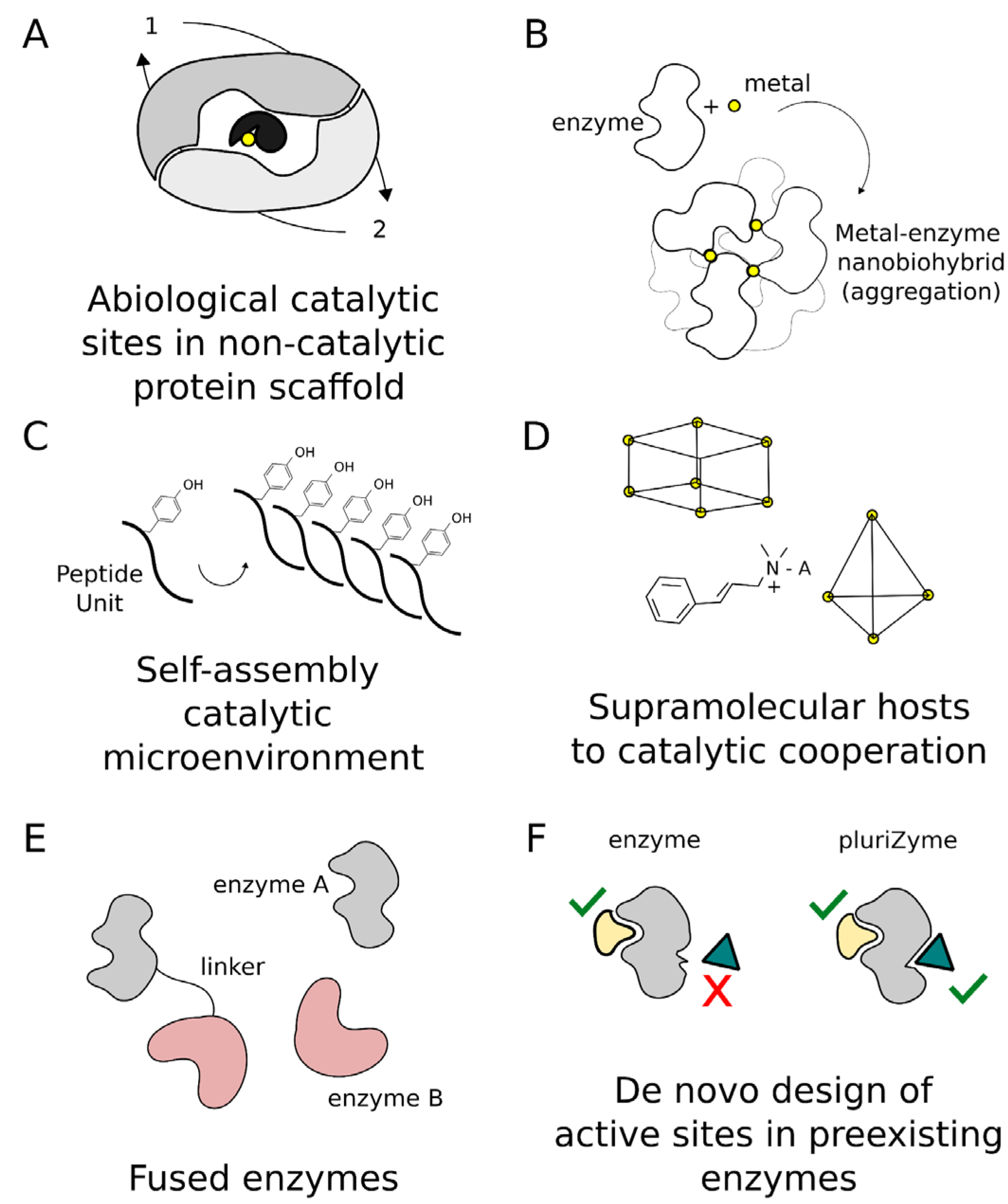
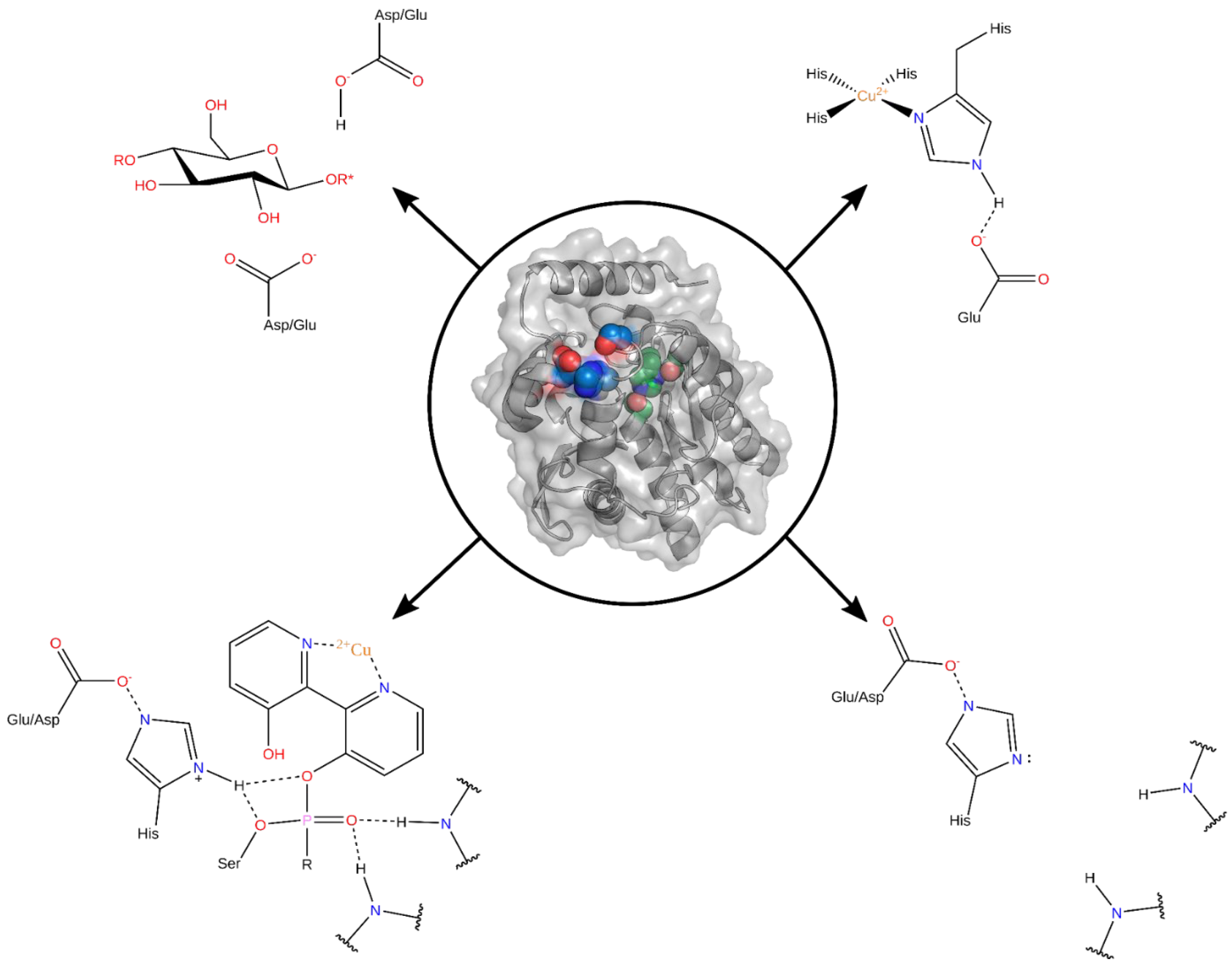
1. Introduction
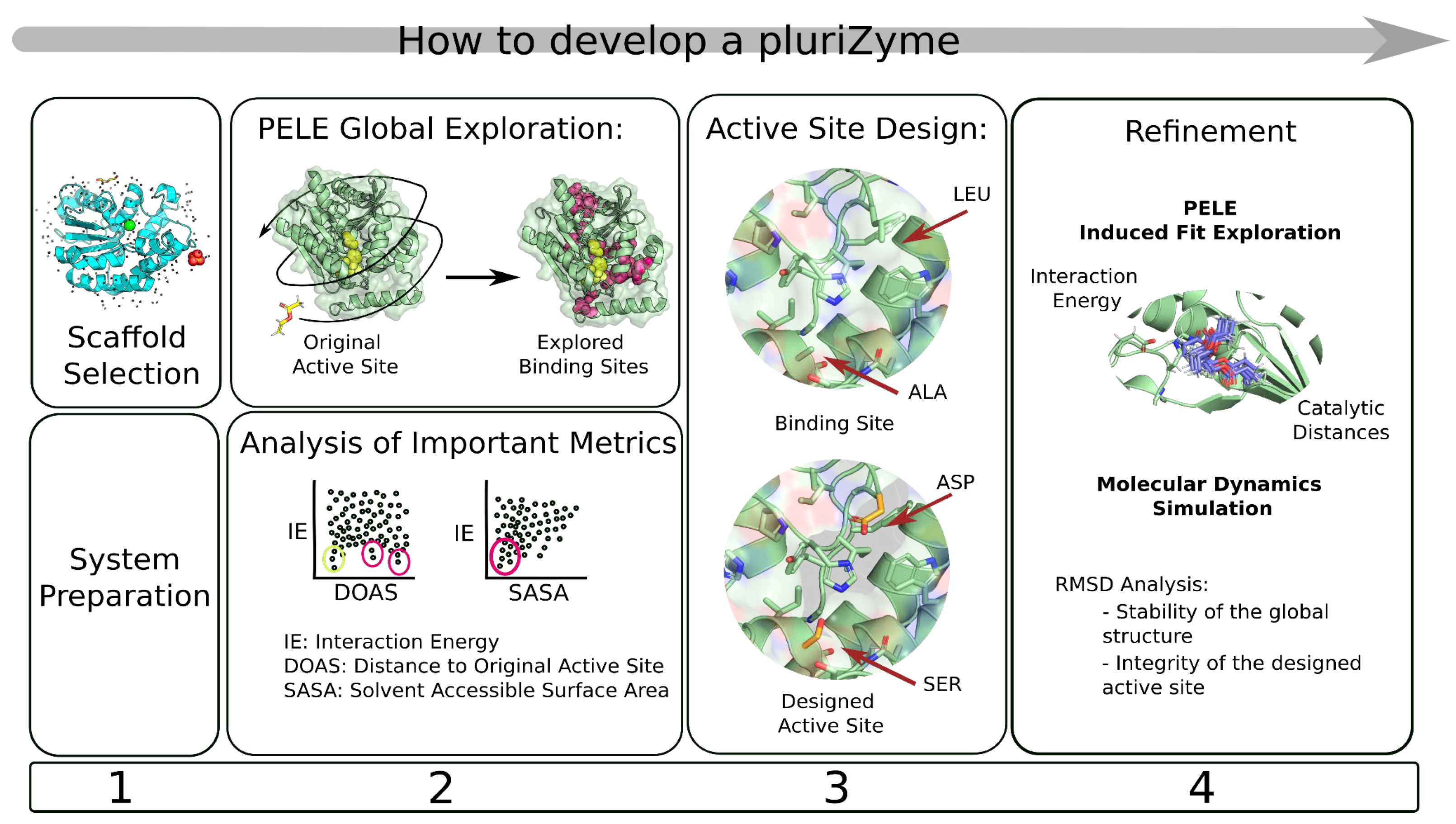
2. How to Develop a PluriZyme
2.1. System Preparation
2.2. Binding Site Search
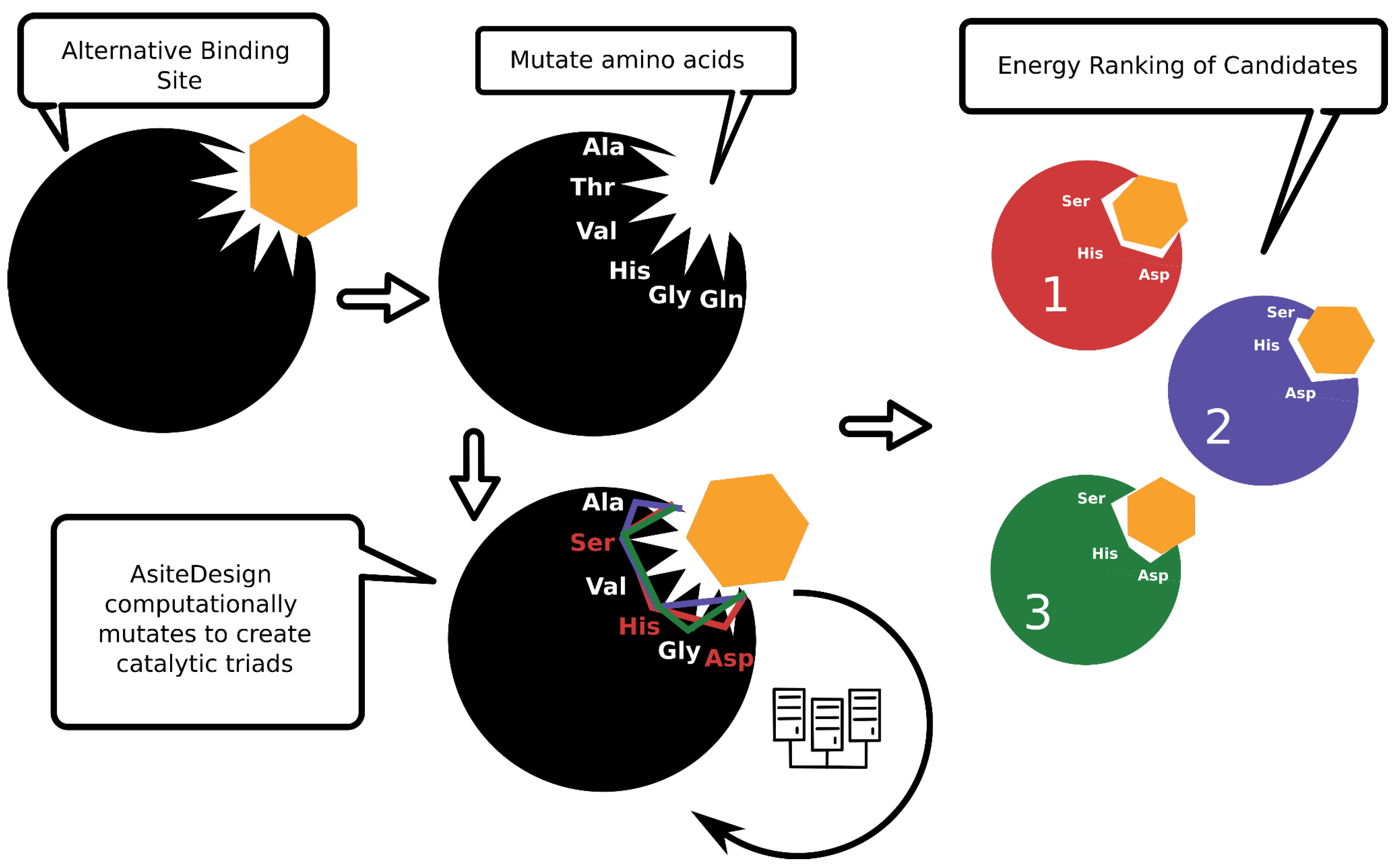
2.3. From Binding Site to Active Site
2.4. Refinement
2.5. Automation of PluriZyme Design
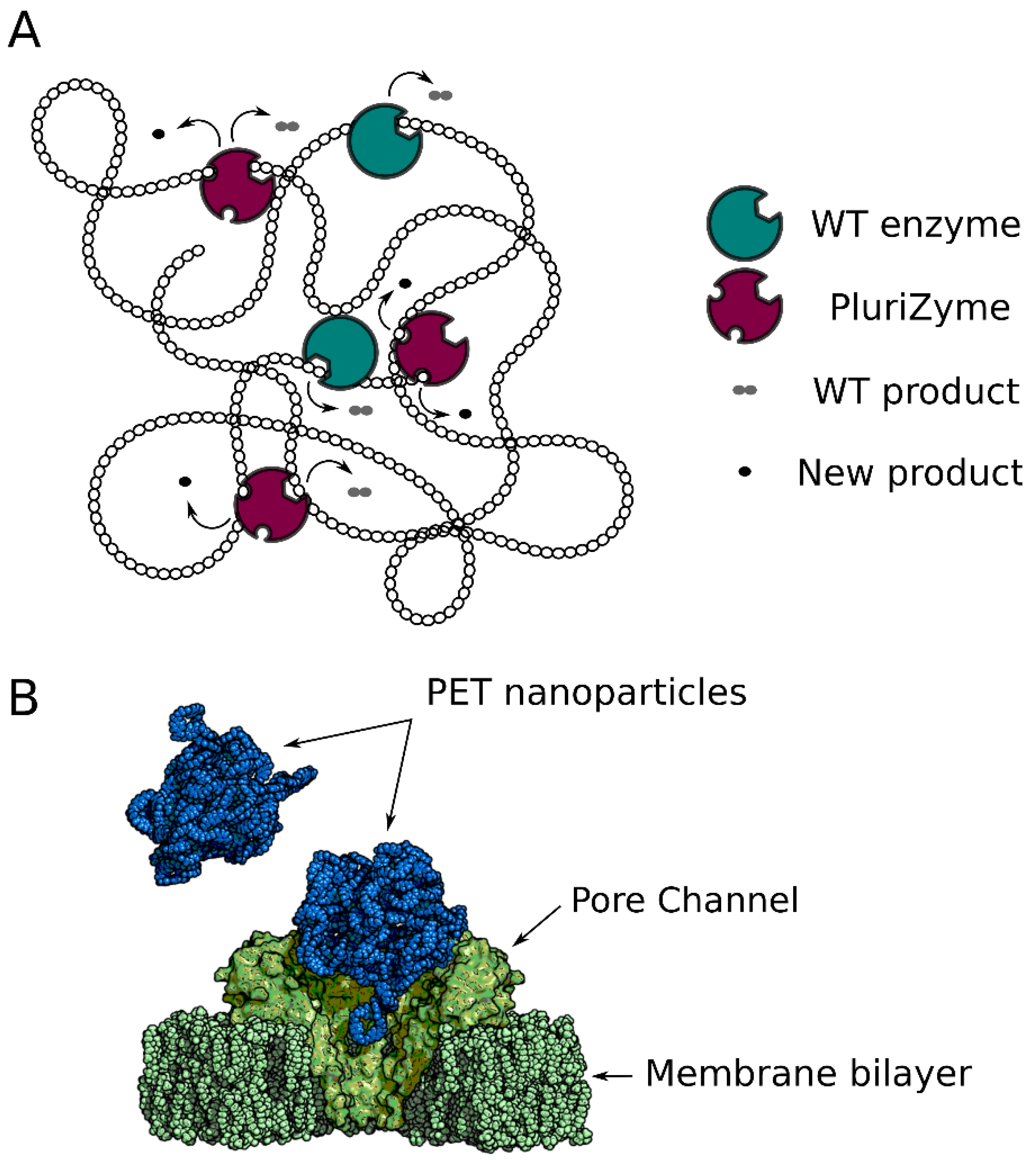
3. Successful Designs of PluriZymes

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| System (PDB ID) | Original Activity | Original Active Site(s) | Added Active Site | Improvement Fold 1 | Highest Activity 2 | ΔTm 3 (°C) | Topt 4 (°C) | PY 5 (mg/L) | Ref. |
|---|---|---|---|---|---|---|---|---|---|
| LAE6 (high-promiscuity esterase) [5JD4] | Esterase | S161/H286/D256 | S211/L214H/E25D | average/max kcat: ~3.4-fold/~74-fold | 57.8 s−1 (phenyl propionate) | - | ~35–45 | 12 | [21] |
| LAE5 (low-promiscuity esterase) [5JD3] | Esterase | S15/H195/D192 | L30S/H34/L57D | max kcat: ~11.5-fold | 6.28 U/mg (phenyl propionate) | - | ~25 | 45.5 | [21] |
| LAE6_pluriZyme [6I8F] | Esterase | S161/H286/D256 and S211/H214/D25 | L24C/H214 | - | 2.63 U/mg (casein) | - | 70–75 | ~10 | [43] |
| class III ω-transaminase [7QYG] | Transaminase | K289 | A172S/Q173H/E317 | - | 65.13 s−1 (phenyl propionate) | 5.62 | 50–55 (transaminase activity) 60–65 (hydrolase activity) | 32–45 | [22] |
| Actinia fragacea porin [4TSY] | - | - | K20H/T21S | - | 424 M−1 s−1 | 2.1 | 35–45 | 15 | [47] |
| Actinia fragacea porin [4TSY] | - | - | D38S/E173Q | - | 1362 M−1 s−1 | 6.3 | 35–45 | 30 | [47] |
| Feruloyl esterase [8BBI] | Xylanase | E144/E251 | * | - | 1.64 ± 10 U/mg) | - | - | - | [46] |
4. PluriZymes and Related Designs for Polymer Degradation
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lovelock, S.L.; Crawshaw, R.; Basler, S.; Levy, C.; Baker, D.; Hilvert, D.; Green, A.P. The road to fully programmable protein catalysis. Nature 2022, 606, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Snajdrova, R.; Moore, J.C.; Baldenius, K.; Bornscheuer, U.T. Biocatalysis: Enzymatic Synthesis for Industrial Applications. Angew. Chem. Int. Ed. Engl. 2021, 60, 88–119. [Google Scholar] [CrossRef] [PubMed]
- Arnal, G.; Anglade, J.; Gavalda, S.; Tournier, V.; Chabot, N.; Bornscheuer, U.T.; Weber, G.; Marty, A. Assessment of Four Engineered PET Degrading Enzymes Considering Large-Scale Industrial Applications. ACS Catal. 2023, 13, 13156–13166. [Google Scholar] [CrossRef] [PubMed]
- Roda, S.; Santiago, G.; Guallar, V. Mapping enzyme-substrate interactions: Its potential to study the mechanism of enzymes. Adv. Protein Chem. Struct. Biol. 2020, 122, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Dauparas, J.; Anishchenko, I.; Bennett, N.; Bai, H.; Ragotte, R.J.; Milles, L.F.; Wicky, B.I.M.; Courbet, A.; de Haas, R.J.; Bethel, N.; et al. Robust deep learning–based protein sequence design using ProteinMPNN. Science 2022, 378, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Pavelka, A.; Sebestova, E.; Kozlikova, B.; Brezovsky, J.; Sochor, J.; Damborsky, J. CAVER: Algorithms for Analyzing Dynamics of Tunnels in Macromolecules. IEEE/ACM Trans. Comput. Biol. Bioinform. 2016, 13, 505–517. [Google Scholar] [CrossRef]
- Richter, F.; Leaver-Fay, A.; Khare, S.D.; Bjelic, S.; Baker, D. De Novo Enzyme Design Using Rosetta3. PLoS ONE 2011, 6, e19230. [Google Scholar] [CrossRef]
- Labrou, N. Random Mutagenesis Methods for In Vitro Directed Enzyme Evolution. Curr. Protein Pept. Sci. 2010, 11, 91–100. [Google Scholar] [CrossRef]
- Agresti, J.J.; Antipov, E.; Abate, A.R.; Ahn, K.; Rowat, A.C.; Baret, J.-C.; Marquez, M.; Klibanov, A.M.; Griffiths, A.D.; Weitz, D.A. Ultrahigh-throughput screening in drop-based microfluidics for directed evolution. Proc. Natl. Acad. Sci. USA 2010, 107, 4004–4009. [Google Scholar] [CrossRef]
- Jakočiūnas, T.; Pedersen, L.E.; Lis, A.V.; Jensen, M.K.; Keasling, J.D. CasPER, a method for directed evolution in genomic contexts using mutagenesis and CRISPR/Cas9. Metab. Eng. 2018, 48, 288–296. [Google Scholar] [CrossRef]
- Anishchenko, I.; Pellock, S.J.; Chidyausiku, T.M.; Ramelot, T.A.; Ovchinnikov, S.; Hao, J.; Bafna, K.; Norn, C.; Kang, A.; Bera, A.K.; et al. De novo protein design by deep network hallucination. Nature 2021, 600, 547–552. [Google Scholar] [CrossRef] [PubMed]
- Burke, A.J.; Lovelock, S.L.; Frese, A.; Crawshaw, R.; Ortmayer, M.; Dunstan, M.; Levy, C.; Green, A.P. Design and evolution of an enzyme with a non-canonical organocatalytic mechanism. Nature 2019, 570, 219–223. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Diaz, D.J.; Czarnecki, N.J.; Zhu, C.; Kim, W.; Shroff, R.; Acosta, D.J.; Alexander, B.R.; Cole, H.O.; Zhang, Y.; et al. Machine learning-aided engineering of hydrolases for PET depolymerization. Nature 2022, 604, 662–667. [Google Scholar] [CrossRef] [PubMed]
- Mazurenko, S.; Prokop, Z.; Damborsky, J. Machine Learning in Enzyme Engineering. ACS Catal. 2020, 10, 1210–1223. [Google Scholar] [CrossRef]
- Amrein, B.A.; Bauer, P.; Duarte, F.; Carlsson, J.; Naworyta, A.; Mowbray, S.L.; Widersten, M.; Kamerlin, S.C.L. Expanding the Catalytic Triad in Epoxide Hydrolases and Related Enzymes. ACS Catal. 2015, 5, 5702–5713. [Google Scholar] [CrossRef]
- Hong, N.-S.; Petrović, D.; Lee, R.; Gryn’ova, G.; Purg, M.; Saunders, J.; Bauer, P.; Carr, P.D.; Lin, C.-Y.; Mabbitt, P.D.; et al. The Evolution of Multiple Active Site Configurations in a Designed Enzyme. Nat. Commun. 2018, 9, 3900. [Google Scholar] [CrossRef]
- Petrović, D.; Risso, V.A.; Kamerlin, S.C.L.; Sanchez-Ruiz, J.M. Conformational dynamics and enzyme evolution. J. R. Soc. Interface 2018, 15, 20180330. [Google Scholar] [CrossRef]
- Roda, S.; Robles-Martín, A.; Xiang, R.; Kazemi, M.; Guallar, V. Structural-Based Modeling in Protein Engineering. A Must Do. J. Phys. Chem. B 2021, 125, 6491–6500. [Google Scholar] [CrossRef]
- Santiago, G.; Martínez-Martínez, M.; Alonso, S.; Bargiela, R.; Coscolín, C.; Golyshin, P.N.; Guallar, V.; Ferrer, M. Rational Engineering of Multiple Active Sites in an Ester Hydrolase. Biochemistry 2018, 57, 2245–2255. [Google Scholar] [CrossRef]
- Carballares, D.; Morellon-Sterling, R.; Fernandez-Lafuente, R. Design of Artificial Enzymes Bearing Several Active Centers: New Trends, Opportunities and Problems. Int. J. Mol. Sci. 2022, 23, 5304. [Google Scholar] [CrossRef]
- Alonso, S.; Santiago, G.; Cea-Rama, I.; Fernandez-Lopez, L.; Coscolín, C.; Modregger, J.; Ressmann, A.K.; Martínez-Martínez, M.; Marrero, H.; Bargiela, R.; et al. Genetically engineered proteins with two active sites for enhanced biocatalysis and synergistic chemo- and biocatalysis. Nat. Catal. 2020, 3, 319–328. [Google Scholar] [CrossRef]
- Roda, S.; Fernandez-Lopez, L.; Benedens, M.; Bollinger, A.; Thies, S.; Schumacher, J.; Coscolín, C.; Kazemi, M.; Santiago, G.; Gertzen, C.G.W.; et al. A Plurizyme with Transaminase and Hydrolase Activity Catalyzes Cascade Reactions. Angew. Chem. Int. Ed. 2022, 61, e202207344. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Caballero, M.; Navarro, S.; Nuez-Martínez, M.; Peccati, F.; Rodríguez-Santiago, L.; Sodupe, M.; Teixidor, F.; Ventura, S. pH-Responsive Self-Assembly of Amyloid Fibrils for Dual Hydrolase-Oxidase Reactions. ACS Catal. 2021, 11, 595–607. [Google Scholar] [CrossRef]
- Bos, J.; Browne, W.R.; Driessen, A.J.M.; Roelfes, G. Supramolecular Assembly of Artificial Metalloenzymes Based on the Dimeric Protein LmrR as Promiscuous Scaffold. J. Am. Chem. Soc. 2015, 137, 9796–9799. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Roelfes, G. Synergistic catalysis in an artificial enzyme by simultaneous action of two abiological catalytic sites. Nat. Catal. 2020, 3, 289–294. [Google Scholar] [CrossRef]
- Palomo, J.M. Nanobiohybrids: A new concept for metal nanoparticles synthesis. Chem. Commun. 2019, 55, 9583–9589. [Google Scholar] [CrossRef] [PubMed]
- Filice, M.; Marciello, M.; Morales, M.D.P.; Palomo, J.M. Synthesis of heterogeneous enzyme–metal nanoparticle biohybrids in aqueous media and their applications in C–C bond formation and tandem catalysis. Chem. Commun. 2013, 49, 6876–6878. [Google Scholar] [CrossRef] [PubMed]
- Benavente, R.; Lopez-Tejedor, D.; Morales, M.D.P.; Perez-Rizquez, C.; Palomo, J.M. The enzyme-induced formation of iron hybrid nanostructures with different morphologies. Nanoscale 2020, 12, 12917–12927. [Google Scholar] [CrossRef]
- Benavente, R.; Lopez-Tejedor, D.; Palomo, J.M. Synthesis of a superparamagnetic ultrathin FeCO3 nanorods–enzyme bionanohybrid as a novel heterogeneous catalyst. Chem. Commun. 2018, 54, 6256–6259. [Google Scholar] [CrossRef]
- Zhao, L.; Cai, J.; Li, Y.; Wei, J.; Duan, C. A host–guest approach to combining enzymatic and artificial catalysis for catalyzing biomimetic monooxygenation. Nat. Commun. 2020, 11, 2903. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Sastry, G.M.; Adzhigirey, M.; Day, T.; Annabhimoju, R.; Sherman, W. Protein and ligand preparation: Parameters, protocols, and influence on virtual screening enrichments. J. Comput. Aided Mol. Des. 2013, 27, 221–234. [Google Scholar] [CrossRef] [PubMed]
- Bochevarov, A.D.; Harder, E.; Hughes, T.F.; Greenwood, J.R.; Braden, D.A.; Philipp, D.M.; Rinaldo, D.; Halls, M.D.; Zhang, J.; Friesner, R.A. Jaguar: A high-performance quantum chemistry software program with strengths in life and materials sciences. Int. J. Quantum Chem. 2013, 113, 2110–2142. [Google Scholar] [CrossRef]
- Acebes, S.; Fernandez-Fueyo, E.; Monza, E.; Lucas, M.F.; Almendral, D.; Ruiz-Dueñas, F.J.; Lund, H.; Martinez, A.T.; Guallar, V. Rational Enzyme Engineering Through Biophysical and Biochemical Modeling. ACS Catal. 2016, 6, 1624–1629. [Google Scholar] [CrossRef]
- Lecina, D.; Gilabert, J.F.; Guallar, V. Adaptive simulations, towards interactive protein-ligand modeling. Sci. Rep. 2017, 7, 8466. [Google Scholar] [CrossRef] [PubMed]
- Roda, S.; Fernandez-Lopez, L.; Cañadas, R.; Santiago, G.; Ferrer, M.; Guallar, V. Computationally Driven Rational Design of Substrate Promiscuity on Serine Ester Hydrolases. ACS Catal. 2021, 11, 3590–3601. [Google Scholar] [CrossRef]
- Campbell, S.J.; Gold, N.D.; Jackson, R.M.; Westhead, D.R. Ligand binding: Functional site location, similarity and docking. Curr. Opin. Struct. Biol. 2003, 13, 389–395. [Google Scholar] [CrossRef]
- Kua, J.; Zhang, Y.; McCammon, J.A. Studying Enzyme Binding Specificity in Acetylcholinesterase Using a Combined Molecular Dynamics and Multiple Docking Approach. J. Am. Chem. Soc. 2002, 124, 8260–8267. [Google Scholar] [CrossRef]
- Rauer, C.; Sen, N.; Waman, V.P.; Abbasian, M.; Orengo, C.A. Computational approaches to predict protein functional families and functional sites. Curr. Opin. Struct. Biol. 2021, 70, 108–122. [Google Scholar] [CrossRef]
- Pokkuluri, P.R.; Gu, M.; Cai, X.; Raffen, R.; Stevens, F.J.; Schiffer, M. Factors Contributing to Decreased Protein Stability When Aspartic Acid Residues Are in Beta-Sheet Regions. Protein Sci. 2002, 11, 1687–1694. [Google Scholar] [CrossRef]
- Roda, S.; Terholsen, H.; Meyer, J.-R.-H.; Guallar, V.; Bornscheuer, U.; Kazemi, M. AsiteDesign: A semi-rational algorithm for automated enzyme design. J. Phys. Chem. B 2023, 127, 2661–2670. [Google Scholar] [CrossRef] [PubMed]
- Chaudhury, S.; Lyskov, S.; Gray, J.J. PyRosetta: A script-based interface for implementing molecular modeling algorithms using Rosetta. Bioinformatics 2010, 26, 689–691. [Google Scholar] [CrossRef] [PubMed]
- Coscolín, C.; Katzke, N.; García-Moyano, A.; Navarro-Fernández, J.; Almendral, D.; Martínez-Martínez, M.; Bollinger, A.; Bargiela, R.; Gertler, C.; Chernikova, T.N.; et al. Bioprospecting Reveals Class III ω-Transaminases Converting Bulky Ketones and Environmentally Relevant Polyamines. Appl. Environ. Microbiol. 2019, 85, e02404-18. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Lopez, L.; Roda, S.; Gonzalez-Alfonso, J.L.; Plou, F.J.; Guallar, V.; Ferrer, M. Design and Characterization of In-One Protease-Esterase PluriZyme. Int. J. Mol. Sci. 2022, 23, 13337. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zheng, D.; Zhang, Z.; Ye, H.; Cao, Q.; Zhang, C.; Dong, Z.; Feng, D.; Zuo, J. Efficacy of combination of endo-xylanase and xylan-debranching enzymes in improving cereal bran utilization in piglet diet. Anim. Biosci. 2022, 35, 1733–1743. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, R.; Guallar, V. Rational Enzyme Engineering of Different Active Sites on a Xylanase; Barcelona Supercomputing Center: Barcelona, Spain, 2022; pp. 66–67. [Google Scholar]
- Robles-Martín, A.; Amigot-Sánchez, R.; Fernandez-Lopez, L.; Gonzalez-Alfonso, J.L.; Roda, S.; Alcolea-Rodriguez, V.; Heras-Márquez, D.; Almendral, D.; Coscolín, C.; Plou, F.J.; et al. Sub-Micro- and Nano-Sized Polyethylene Terephthalate Deconstruction with Engineered Protein Nanopores. Nat. Catal. 2023, 6, 1174–1185. [Google Scholar] [CrossRef]
- Tiso, T.; Narancic, T.; Wei, R.; Pollet, E.; Beagan, N.; Schröder, K.; Honak, A.; Jiang, M.; Kenny, S.T.; Wierckx, N.; et al. Towards bio-upcycling of polyethylene terephthalate. Metab. Eng. 2021, 66, 167–178. [Google Scholar] [CrossRef]
- Hu, Y.; Wang, Y.; Zhang, X.; Qian, J.; Xing, X.; Wang, X. Synthesis of poly(ethylene terephthalate) based on glycolysis of waste PET fiber. J. Macromol. Sci. Part A 2020, 57, 430–438. [Google Scholar] [CrossRef]
- Köpnick, H.; Schmidt, M.; Brügging, W.; Rüter, J.; Kaminsky, W. Polyesters. In Ullmann’s Encyclopedia of Industrial Chemistry; Wiley: Hoboken, NJ, USA, 2000; Available online: https://onlinelibrary.wiley.com/doi/10.1002/14356007.a21_227 (accessed on 12 October 2023).
- Ji, L.N. Study on Preparation Process and Properties of Polyethylene Terephthalate (PET). In Materials Science, Engineering, in Applied Mechanics and Materials; Zurich Tomo: Zürich, Switzerland, 2013; Volume 312, p. 406. [Google Scholar] [CrossRef]
- Shukla, S.; Harad, A.M.; Jawale, L.S. Recycling of waste PET into useful textile auxiliaries. Waste Manag. 2008, 28, 51–56. [Google Scholar] [CrossRef]
- Liu, P.; Zheng, Y.; Yuan, Y.; Zhang, T.; Li, Q.; Liang, Q.; Su, T.; Qi, Q. Valorization of Polyethylene Terephthalate to Muconic Acid by Engineering Pseudomonas putida. Int. J. Mol. Sci. 2022, 23, 10997. [Google Scholar] [CrossRef]
- Kim, H.T.; Kim, J.K.; Gil Cha, H.; Kang, M.J.; Lee, H.S.; Khang, T.U.; Yun, E.J.; Lee, D.-H.; Song, B.K.; Park, S.J.; et al. Biological Valorization of Poly(ethylene terephthalate) Monomers for Upcycling Waste PET. ACS Sustain. Chem. Eng. 2019, 7, 19396–19406. [Google Scholar] [CrossRef]
- Ghasemzadeh, M.A.; Mirhosseini-Eshkevari, B.; Abdollahi-Basir, M.H. MIL-53(Fe) Metal-Organic Frameworks (MOFs) as an Efficient and Reusable Catalyst for the One-Pot Four-Component Synthesis of Pyrano[2,3-c]-pyrazoles. Appl. Organomet. Chem. 2019, 33, e4679. [Google Scholar] [CrossRef]
- Zhan, Y.; Hou, W.; Li, G.; Shen, Y.; Zhang, Y.; Tang, Y. Oxidant-Free Transformation of Ethylene Glycol toward Glycolic Acid in Water. ACS Sustain. Chem. Eng. 2019, 7, 17559–17564. [Google Scholar] [CrossRef]
- Poshyvailo, L.; von Lieres, E.; Kondrat, S. Does Metabolite Channeling Accelerate Enzyme-Catalyzed Cascade Reactions? PLoS ONE 2017, 12, e0172673. [Google Scholar] [CrossRef] [PubMed]
- Gad, S.; Ayakar, S. Protein scaffolds: A tool for multi-enzyme assembly. Biotechnol. Rep. 2021, 32, e00670. [Google Scholar] [CrossRef] [PubMed]
- Wheeldon, I.; Minteer, S.D.; Banta, S.; Barton, S.C.; Atanassov, P.; Sigman, M. Substrate channelling as an approach to cascade reactions. Nat. Chem. 2016, 8, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Aalbers, F.S.; Fraaije, M.W. Enzyme Fusions in Biocatalysis: Coupling Reactions by Pairing Enzymes. Chembiochem 2019, 20, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Davis, H.J.; Ward, T.R. Artificial Metalloenzymes: Challenges and Opportunities. ACS Cent. Sci. 2019, 5, 1120–1136. [Google Scholar] [CrossRef]
- Schwizer, F.; Okamoto, Y.; Heinisch, T.; Gu, Y.; Pellizzoni, M.M.; Lebrun, V.; Reuter, R.; Köhler, V.; Lewis, J.C.; Ward, T.R. Artificial Metalloenzymes: Reaction Scope and Optimization Strategies. Chem. Rev. 2018, 118, 142–231. [Google Scholar] [CrossRef]
- Wang, J.; Lisanza, S.; Juergens, D.; Tischer, D.; Watson, J.L.; Castro, K.M.; Ragotte, R.; Saragovi, A.; Milles, L.F.; Baek, M.; et al. Scaffolding protein functional sites using deep learning. Science 2022, 377, 387–394. [Google Scholar] [CrossRef]
- Vornholt, T.; Christoffel, F.; Pellizzoni, M.M.; Panke, S.; Ward, T.R.; Jeschek, M. Systematic engineering of artificial metalloenzymes for new-to-nature reactions. Sci. Adv. 2021, 7, eabe4208. [Google Scholar] [CrossRef] [PubMed]
- CAZypedia Consortium. Ten Years of CAZypedia: A Living Encyclopedia of Carbohydrate-Active Enzymes. Glycobiology 2018, 28, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Branneby, C.; Carlqvist, P.; Magnusson, A.; Hult, K.; Brinck, T.; Berglund, P. Carbon−Carbon Bonds by Hydrolytic Enzymes. J. Am. Chem. Soc. 2003, 125, 874–875. [Google Scholar] [CrossRef] [PubMed]
- Rauwerdink, A.; Kazlauskas, R.J. How the Same Core Catalytic Machinery Catalyzes 17 Different Reactions: The Serine-Histidine-Aspartate Catalytic Triad of α/β-Hydrolase Fold Enzymes. ACS Catal. 2015, 5, 6153–6176. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Robles-Martín, A.; Roda, S.; Muñoz-Tafalla, R.; Guallar, V. Behind the Scenes of PluriZyme Designs. Eng 2024, 5, 91-103. https://doi.org/10.3390/eng5010006
Robles-Martín A, Roda S, Muñoz-Tafalla R, Guallar V. Behind the Scenes of PluriZyme Designs. Eng. 2024; 5(1):91-103. https://doi.org/10.3390/eng5010006
Chicago/Turabian StyleRobles-Martín, Ana, Sergi Roda, Rubén Muñoz-Tafalla, and Victor Guallar. 2024. "Behind the Scenes of PluriZyme Designs" Eng 5, no. 1: 91-103. https://doi.org/10.3390/eng5010006
APA StyleRobles-Martín, A., Roda, S., Muñoz-Tafalla, R., & Guallar, V. (2024). Behind the Scenes of PluriZyme Designs. Eng, 5(1), 91-103. https://doi.org/10.3390/eng5010006