
Different Expression Patterns of Metabolic Reprogramming Proteins in Testicular Germ Cell Cancer

 ,
,  ,
,  ,
,  ,
, 

Abstract
1. Introduction
2. Materials and Methods
2.1. Antibodies
2.2. Human Tissues
2.3. Histopathological Analysis
2.4. Immunohistochemistry
2.5. Scoring System
2.6. Statistical Analysis
3. Results
3.1. Morphological Analysis
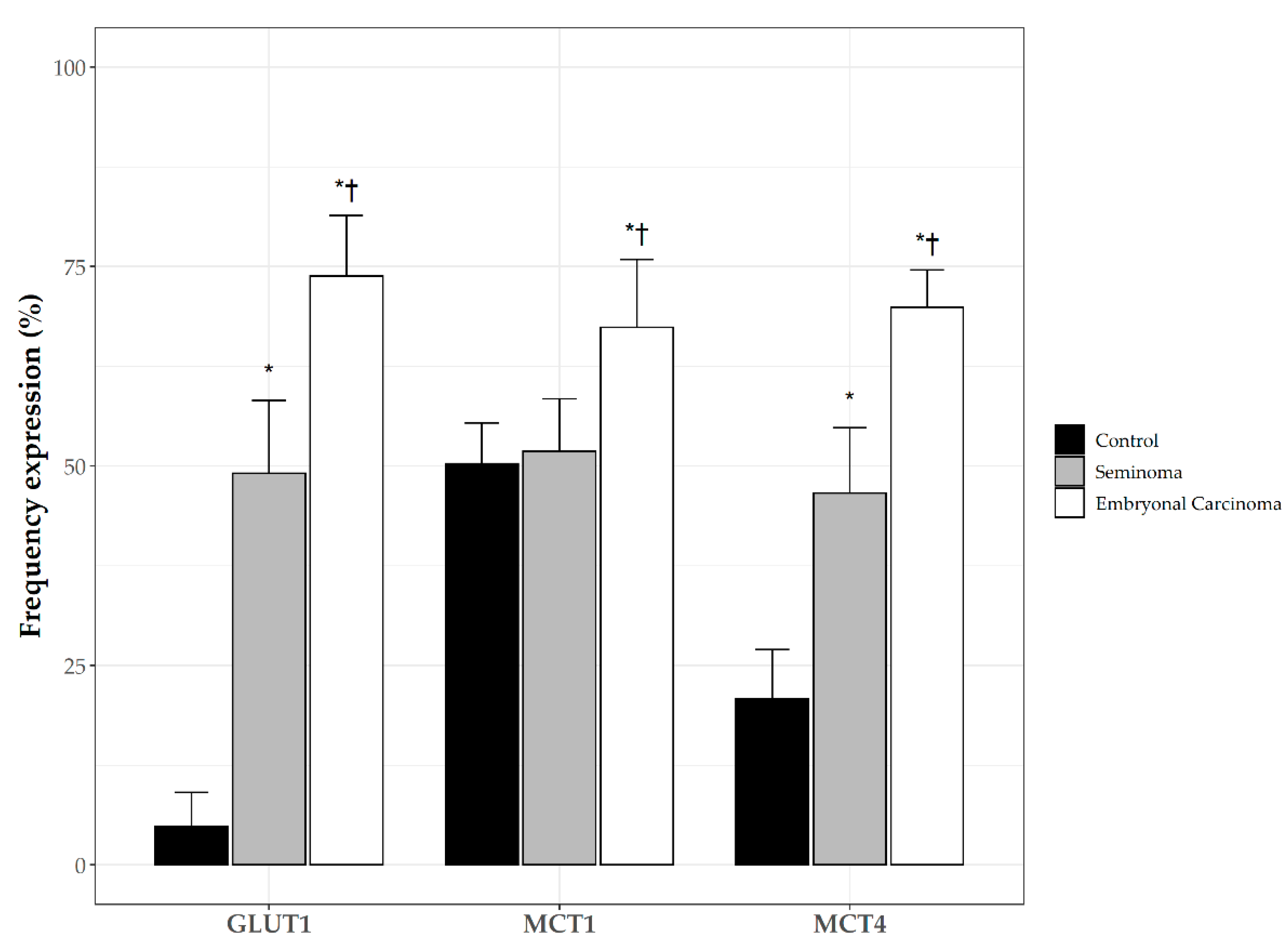
3.2. Immunohistochemical Localization Markers of Aerobic Glycolysis in Control Testis and TGCTs
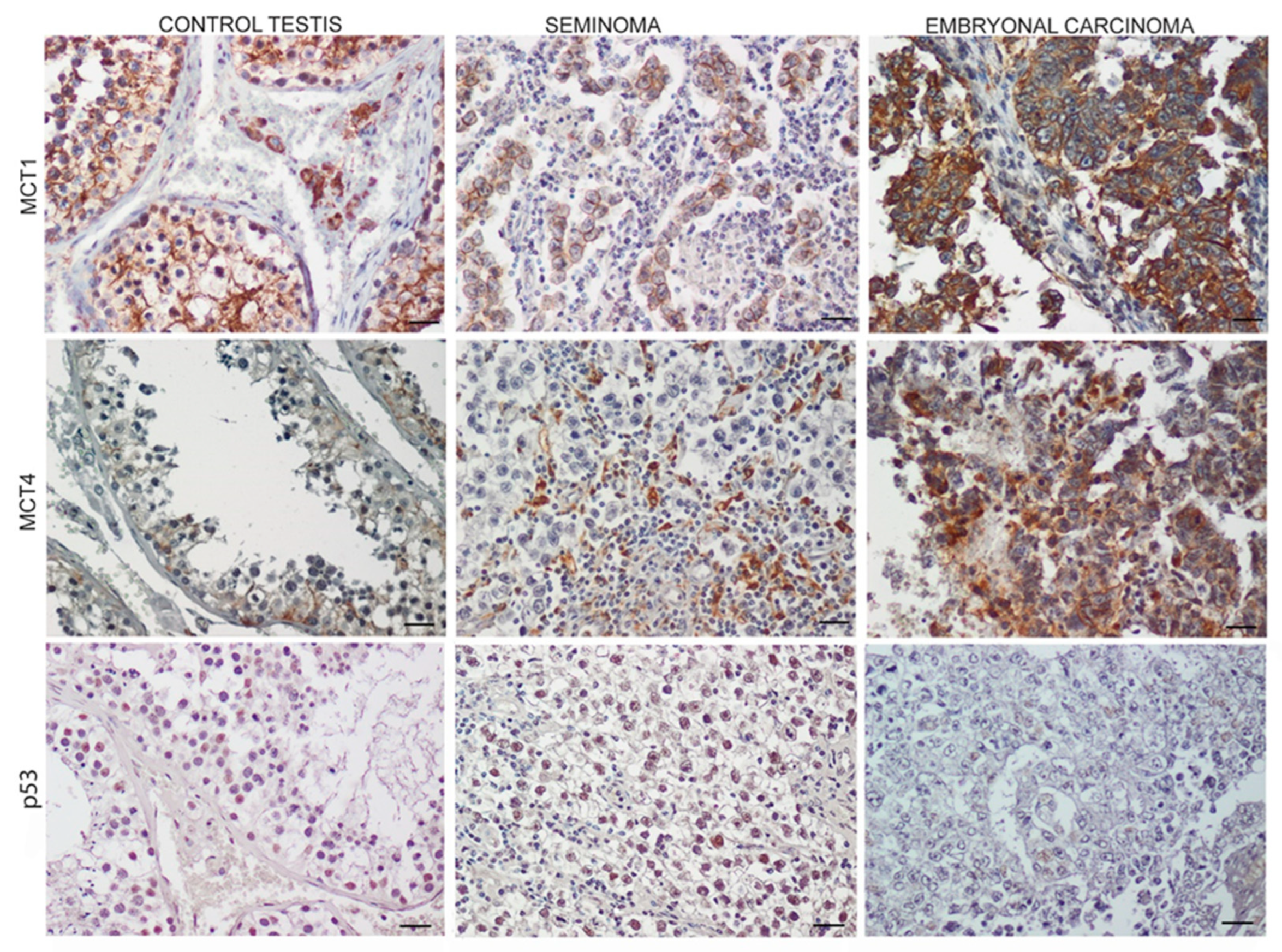
3.3. Monocarboxylate Transporters 1 (MCT1),4 (MCT4) and p53 Expression in TGCTs
3.4. LDH-A Pattern Expression in TGCTs
3.5. Oxidative Mitochondrial Protein Expression
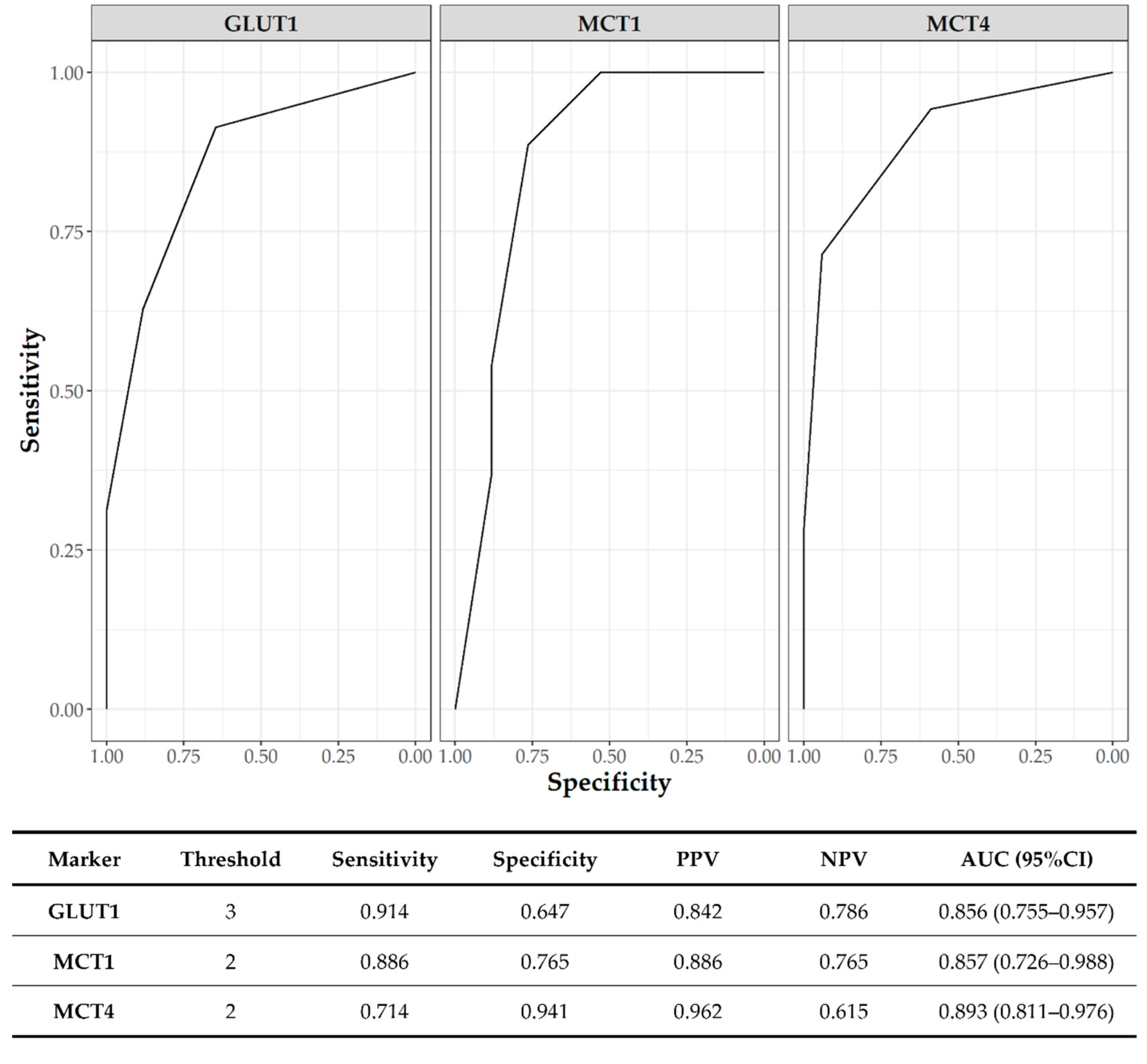
3.6. Clinicopathological Significance of GLUT1 and MCTs
4. Discussion
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Schneider, D.T.; Calaminus, G.; Koch, S.; Teske, C.; Schmidt, P.; Haas, R.J.; Harms, D.; Göbel, U. Epidemiologic Analysis of 1,442 Children and Adolescents Registered in the German Germ Cell Tumor Protocols. Pediatr. Blood Cancer 2004, 42, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Nallu, A.; Mannuel, H.D.; Hussain, A. Testicular Germ Cell Tumors: Biology and Clinical Update. Curr. Opin. Oncol. 2013, 25, 266–272. [Google Scholar] [CrossRef] [PubMed]
- Batool, A.; Liu, X.-M.; Zhang, C.-L.; Hao, C.-F.; Chen, S.-R.; Liu, Y.-X. Recent Advances in the Regulation of Testicular Germ Cell Tumors by MicroRNAs. Front. Biosci. Landmark Ed. 2019, 24, 765–776. [Google Scholar] [CrossRef] [PubMed]
- Ghazarian, A.A.; Rusner, C.; Trabert, B.; Braunlin, M.; McGlynn, K.A.; Stang, A. Testicular Cancer among US Men Aged 50 Years and Older. Cancer Epidemiol. 2018, 55, 68–72. [Google Scholar] [CrossRef]
- Berney, D.M.; Looijenga, L.H.J.; Idrees, M.; Oosterhuis, J.W.; Rajpert-De Meyts, E.; Ulbright, T.M.; Skakkebaek, N.E. Germ Cell Neoplasia in Situ (GCNIS): Evolution of the Current Nomenclature for Testicular Pre-Invasive Germ Cell Malignancy. Histopathology 2016, 69, 7–10. [Google Scholar] [CrossRef] [PubMed]
- Rajpert-De Meyts, E.; McGlynn, K.A.; Okamoto, K.; Jewett, M.A.S.; Bokemeyer, C. Testicular Germ Cell Tumours. Lancet Lond. Engl. 2016, 387, 1762–1774. [Google Scholar] [CrossRef]
- Piulats, J.M.; Jiménez, L.; García del Muro, X.; Villanueva, A.; Viñals, F.; Germà-Lluch, J.R. Molecular Mechanisms behind the Resistance of Cisplatin in Germ Cell Tumours. Clin. Transl. Oncol. Off. Publ. Fed. Span. Oncol. Soc. Natl. Cancer Inst. Mex. 2009, 11, 780–786. [Google Scholar] [CrossRef] [PubMed]
- Litchfield, K.; Levy, M.; Huddart, R.A.; Shipley, J.; Turnbull, C. The Genomic Landscape of Testicular Germ Cell Tumours: From Susceptibility to Treatment. Nat. Rev. Urol. 2016, 13, 409–419. [Google Scholar] [CrossRef]
- Pavlova, N.N.; Thompson, C.B. The Emerging Hallmarks of Cancer Metabolism. Cell Metab. 2016, 23, 27–47. [Google Scholar] [CrossRef] [PubMed]
- Warburg, O. On the Origin of Cancer Cells. Science 1956, 123, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- DeBerardinis, R.J.; Lum, J.J.; Hatzivassiliou, G.; Thompson, C.B. The Biology of Cancer: Metabolic Reprogramming Fuels Cell Growth and Proliferation. Cell Metab. 2008, 7, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, C.; Longatto-Filho, A.; Azevedo-Silva, J.; Casal, M.; Schmitt, F.C.; Baltazar, F. Role of Monocarboxylate Transporters in Human Cancers: State of the Art. J. Bioenerg. Biomembr. 2012, 44, 127–139. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Ren, P.; Liu, X.; Sun, X.; Zhang, C.; Du, X.; Xing, B. PPP1R26 Drives Hepatocellular Carcinoma Progression by Controlling Glycolysis and Epithelial-Mesenchymal Transition. J. Exp. Clin. Cancer Res. 2022, 41, 101. [Google Scholar] [CrossRef]
- Bonatelli, M.; Silva, E.C.A.; Cárcano, F.M.; Zaia, M.G.; Lopes, L.F.; Scapulatempo-Neto, C.; Pinheiro, C. The Warburg Effect Is Associated With Tumor Aggressiveness in Testicular Germ Cell Tumors. Front. Endocrinol. 2019, 10, 417. [Google Scholar] [CrossRef]
- Silva, E.C.A.; Cárcano, F.M.; Bonatelli, M.; Zaia, M.G.; Morais-Santos, F.; Baltazar, F.; Lopes, L.F.; Scapulatempo-Neto, C.; Pinheiro, C. The Clinicopathological Significance of Monocarboxylate Transporters in Testicular Germ Cell Tumors. Oncotarget 2018, 9, 20386–20398. [Google Scholar] [CrossRef]
- AIOM—Associazione Italiana di Oncologia Medica Tumore del Testicolo: Linee Guida 2021. Available online: https://www.aiom.it/wp-content/uploads/2021/11/2021_LGAIOM_TumoreTesticolo (accessed on 15 May 2022).
- Pavlides, S.; Whitaker-Menezes, D.; Castello-Cros, R.; Flomenberg, N.; Witkiewicz, A.K.; Frank, P.G.; Casimiro, M.C.; Wang, C.; Fortina, P.; Addya, S.; et al. The Reverse Warburg Effect: Aerobic Glycolysis in Cancer Associated Fibroblasts and the Tumor Stroma. Cell Cycle Georget. Tex 2009, 8, 3984–4001. [Google Scholar] [CrossRef]
- Sonveaux, P.; Végran, F.; Schroeder, T.; Wergin, M.C.; Verrax, J.; Rabbani, Z.N.; De Saedeleer, C.J.; Kennedy, K.M.; Diepart, C.; Jordan, B.F.; et al. Targeting Lactate-Fueled Respiration Selectively Kills Hypoxic Tumor Cells in Mice. J. Clin. Investig. 2008, 118, 3930–3942. [Google Scholar] [CrossRef]
- Romero-Garcia, S.; Moreno-Altamirano, M.M.B.; Prado-Garcia, H.; Sánchez-García, F.J. Lactate Contribution to the Tumor Microenvironment: Mechanisms, Effects on Immune Cells and Therapeutic Relevance. Front. Immunol. 2016, 7, 52. [Google Scholar] [CrossRef]
- Pertega-Gomes, N.; Felisbino, S.; Massie, C.E.; Vizcaino, J.R.; Coelho, R.; Sandi, C.; Simoes-Sousa, S.; Jurmeister, S.; Ramos-Montoya, A.; Asim, M.; et al. A Glycolytic Phenotype Is Associated with Prostate Cancer Progression and Aggressiveness: A Role for Monocarboxylate Transporters as Metabolic Targets for Therapy. J. Pathol. 2015, 236, 517–530. [Google Scholar] [CrossRef]
- García-Cañaveras, J.C.; Chen, L.; Rabinowitz, J.D. The Tumor Metabolic Microenvironment: Lessons from Lactate. Cancer Res. 2019, 79, 3155–3162. [Google Scholar] [CrossRef] [PubMed]
- Payen, V.L.; Mina, E.; Van Hée, V.F.; Porporato, P.E.; Sonveaux, P. Monocarboxylate Transporters in Cancer. Mol. Metab. 2020, 33, 48–66. [Google Scholar] [CrossRef]
- Pęcak, A.; Skalniak, Ł.; Pels, K.; Książek, M.; Madej, M.; Krzemień, D.; Malicki, S.; Władyka, B.; Dubin, A.; Holak, T.A.; et al. Anti-CD44 DNA Aptamers Selectively Target Cancer Cells. Nucleic Acid Ther. 2020, 30, 289–298. [Google Scholar] [CrossRef] [PubMed]
- Hao, J.; Chen, H.; Madigan, M.C.; Cozzi, P.J.; Beretov, J.; Xiao, W.; Delprado, W.J.; Russell, P.J.; Li, Y. Co-Expression of CD147 (EMMPRIN), CD44v3-10, MDR1 and Monocarboxylate Transporters Is Associated with Prostate Cancer Drug Resistance and Progression. Br. J. Cancer 2010, 103, 1008–1018. [Google Scholar] [CrossRef] [PubMed]
- Slomiany, M.G.; Grass, G.D.; Robertson, A.D.; Yang, X.Y.; Maria, B.L.; Beeson, C.; Toole, B.P. Hyaluronan, CD44, and Emmprin Regulate Lactate Efflux and Membrane Localization of Monocarboxylate Transporters in Human Breast Carcinoma Cells. Cancer Res. 2009, 69, 1293–1301. [Google Scholar] [CrossRef]
- Perri, A.; Rago, V.; Malivindi, R.; Maltese, L.; Lofaro, D.; Greco, E.A.; Tucci, L.; Bonofiglio, R.; Vergine, M.; Vignera, S.L.; et al. Overexpression of P75ntr in Human Seminoma: A New Biomarker? Life 2021, 11, 629. [Google Scholar] [CrossRef]
- Itahana, Y.; Itahana, K. Emerging Roles of P53 Family Members in Glucose Metabolism. Int. J. Mol. Sci. 2018, 19, 776. [Google Scholar] [CrossRef]
- Kotowski, K.; Rosik, J.; Machaj, F.; Supplitt, S.; Wiczew, D.; Jabłońska, K.; Wiechec, E.; Ghavami, S.; Dzięgiel, P. Role of PFKFB3 and PFKFB4 in Cancer: Genetic Basis, Impact on Disease Development/Progression, and Potential as Therapeutic Targets. Cancers 2021, 13, 909. [Google Scholar] [CrossRef]
- Zhou, S.; Min, Z.; Sun, K.; Qu, S.; Zhou, J.; Duan, H.; Liu, H.; Liu, X.; Gong, Z.; Li, D. MiR-199a-3p/Sp1/LDHA Axis Controls Aerobic Glycolysis in Testicular Tumor Cells. Int. J. Mol. Med. 2018, 42, 2163–2174. [Google Scholar] [CrossRef]
- Kennedy, K.M.; Dewhirst, M.W. Tumor Metabolism of Lactate: The Influence and Therapeutic Potential for MCT and CD147 Regulation. Future Oncol. Lond. Engl. 2010, 6, 127–148. [Google Scholar] [CrossRef]
- Brodsky, A.N.; Odenwelder, D.C.; Harcum, S.W. High Extracellular Lactate Causes Reductive Carboxylation in Breast Tissue Cell Lines Grown under Normoxic Conditions. PLoS ONE 2019, 14, e0213419. [Google Scholar] [CrossRef] [PubMed]
- Das, C.K.; Parekh, A.; Parida, P.K.; Bhutia, S.K.; Mandal, M. Lactate Dehydrogenase A Regulates Autophagy and Tamoxifen Resistance in Breast Cancer. Biochim. Biophys. Acta Mol. Cell Res. 2019, 1866, 1004–1018. [Google Scholar] [CrossRef] [PubMed]
- Jafary, F.; Ganjalikhany, M.R.; Moradi, A.; Hemati, M.; Jafari, S. Novel Peptide Inhibitors for Lactate Dehydrogenase A (LDHA): A Survey to Inhibit LDHA Activity via Disruption of Protein-Protein Interaction. Sci. Rep. 2019, 9, 4686. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, B.T.; Tamai, K.; He, X. Wnt/Beta-Catenin Signaling: Components, Mechanisms, and Diseases. Dev. Cell 2009, 17, 9–26. [Google Scholar] [CrossRef] [PubMed]
- Ding, Q.; He, X.; Xia, W.; Hsu, J.-M.; Chen, C.-T.; Li, L.-Y.; Lee, D.-F.; Yang, J.-Y.; Xie, X.; Liu, J.-C.; et al. Myeloid Cell Leukemia-1 Inversely Correlates with Glycogen Synthase Kinase-3beta Activity and Associates with Poor Prognosis in Human Breast Cancer. Cancer Res. 2007, 67, 4564–4571. [Google Scholar] [CrossRef]
- Farago, M.; Dominguez, I.; Landesman-Bollag, E.; Xu, X.; Rosner, A.; Cardiff, R.D.; Seldin, D.C. Kinase-Inactive Glycogen Synthase Kinase 3beta Promotes Wnt Signaling and Mammary Tumorigenesis. Cancer Res. 2005, 65, 5792–5801. [Google Scholar] [CrossRef]
- Kunnimalaiyaan, M.; Vaccaro, A.M.; Ndiaye, M.A.; Chen, H. Inactivation of Glycogen Synthase Kinase-3beta, a Downstream Target of the Raf-1 Pathway, Is Associated with Growth Suppression in Medullary Thyroid Cancer Cells. Mol. Cancer Ther. 2007, 6, 1151–1158. [Google Scholar] [CrossRef]
- Hay, N. Reprogramming Glucose Metabolism in Cancer: Can It Be Exploited for Cancer Therapy? Nat. Rev. Cancer 2016, 16, 635–649. [Google Scholar] [CrossRef]
- Whitaker-Menezes, D.; Martinez-Outschoorn, U.E.; Flomenberg, N.; Birbe, R.C.; Witkiewicz, A.K.; Howell, A.; Pavlides, S.; Tsirigos, A.; Ertel, A.; Pestell, R.G.; et al. Hyperactivation of Oxidative Mitochondrial Metabolism in Epithelial Cancer Cells in Situ: Visualizing the Therapeutic Effects of Metformin in Tumor Tissue. Cell Cycle Georget. Tex 2011, 10, 4047–4064. [Google Scholar] [CrossRef]
- Zhao, Z.; Han, F.; He, Y.; Yang, S.; Hua, L.; Wu, J.; Zhan, W. Stromal-Epithelial Metabolic Coupling in Gastric Cancer: Stromal MCT4 and Mitochondrial TOMM20 as Poor Prognostic Factors. Eur. J. Surg. Oncol. J. Eur. Soc. Surg. Oncol. Br. Assoc. Surg. Oncol. 2014, 40, 1361–1368. [Google Scholar] [CrossRef]
- Johnson, J.M.; Lai, S.Y.; Cotzia, P.; Cognetti, D.; Luginbuhl, A.; Pribitkin, E.A.; Zhan, T.; Mollaee, M.; Domingo-Vidal, M.; Chen, Y.; et al. Mitochondrial Metabolism as a Treatment Target in Anaplastic Thyroid Cancer. Semin. Oncol. 2015, 42, 915–922. [Google Scholar] [CrossRef] [PubMed]
- Hensley, C.T.; Faubert, B.; Yuan, Q.; Lev-Cohain, N.; Jin, E.; Kim, J.; Jiang, L.; Ko, B.; Skelton, R.; Loudat, L.; et al. Metabolic Heterogeneity in Human Lung Tumors. Cell 2016, 164, 681–694. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | SE | EC | |
|---|---|---|---|
| GLUT1 | 0 (0–0) | 2 (1–3) * | 4 (3–4) *† |
| CD44 | 0 (0–0) | 2 (1.5–2) * | 4 (3–4) *† |
| PFK-1 | 1 (1–1) | 1 (1–1) | 2 (2–2) *† |
| Control | SE | EC | |
|---|---|---|---|
| MCT1 | 2 (2–3) | 1 (0–2) | 4 (3–4) † |
| MCT4 | 1 (0–1) | 2 (0–3) | 4 (3–4) *† |
| P53 | 1 (1–1) | 3 (2–3) * | 0 (0–0) *† |
| Control | SE | EC | |
|---|---|---|---|
| LDH-A | 2 (2–2) | 3 (2–3) * | 4 (3–4) *† |
| pAKT | 1 (1–1) | 1 (1–1) | 2 (2–2) *† |
| pGSK3β | 0 (0–0) | 1 (0–1) | 2 (2–2) *† |
| Control | SE | EC | |
|---|---|---|---|
| TOM20 | 2 (2–2) | 3 (2.5–3) * | 0 (0–1) *† |
| PDH | 0 (0–0) | 1 (1–1) * | 3 (3–3) *† |
| PDK1 | 2 (2–2) | 0 (0–0) * | 1 (1–1) *† |
| Number | GLUT1 | MCT1 | MCT4 | |
|---|---|---|---|---|
| Positive (%) | Positive (%) | Positive (%) | ||
| Histology | ||||
| SE | 35 | 24 (68.6) * | 16 (45.7) * | 25 (71.4) * |
| EC | 17 | 17 (100.0) * | 15 (88.2) | 17 (100.0) * |
| T stage | ||||
| T1 | 31 | 27 (87.0) * | 24 (77.4) * | 25 (80.6) * |
| T2 | 12 | 11 (91.7) * | 8 (66.6) * | 10 (83.3) * |
| T3 | 6 | 5 (83.3) * | 4 (66.6) * | 5 (83.3) * |
| T4 | 3 | 2 (66.6) * | 1 (33.3) * | 2 (66.6) * |
| Vascular invasion | ||||
| No | 48 | 42 (87.5) * | 28 (79.0) * | 40 (83.3) * |
| Yes | 4 | 0 (0.0) | 0 (0.0) | 0 (0.0) * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perri, A.; Lofaro, D.; Bossio, S.; Maltese, L.; Casaburi, I.; Tucci, L.; La Vignera, S.; Aversa, A.; Aquila, S.; Rago, V. Different Expression Patterns of Metabolic Reprogramming Proteins in Testicular Germ Cell Cancer. Endocrines 2022, 3, 578-589. https://doi.org/10.3390/endocrines3040049
Perri A, Lofaro D, Bossio S, Maltese L, Casaburi I, Tucci L, La Vignera S, Aversa A, Aquila S, Rago V. Different Expression Patterns of Metabolic Reprogramming Proteins in Testicular Germ Cell Cancer. Endocrines. 2022; 3(4):578-589. https://doi.org/10.3390/endocrines3040049
Chicago/Turabian StylePerri, Anna, Danilo Lofaro, Sabrina Bossio, Lorenza Maltese, Ivan Casaburi, Luigi Tucci, Sandro La Vignera, Antonio Aversa, Saveria Aquila, and Vittoria Rago. 2022. "Different Expression Patterns of Metabolic Reprogramming Proteins in Testicular Germ Cell Cancer" Endocrines 3, no. 4: 578-589. https://doi.org/10.3390/endocrines3040049
APA StylePerri, A., Lofaro, D., Bossio, S., Maltese, L., Casaburi, I., Tucci, L., La Vignera, S., Aversa, A., Aquila, S., & Rago, V. (2022). Different Expression Patterns of Metabolic Reprogramming Proteins in Testicular Germ Cell Cancer. Endocrines, 3(4), 578-589. https://doi.org/10.3390/endocrines3040049