

Widespread Coral Bleaching and Mass Mortality of Reef-Building Corals in Southern Mexican Pacific Reefs Due to 2023 El Niño Warming

 ,
,  , and
, and
Abstract
1. Introduction
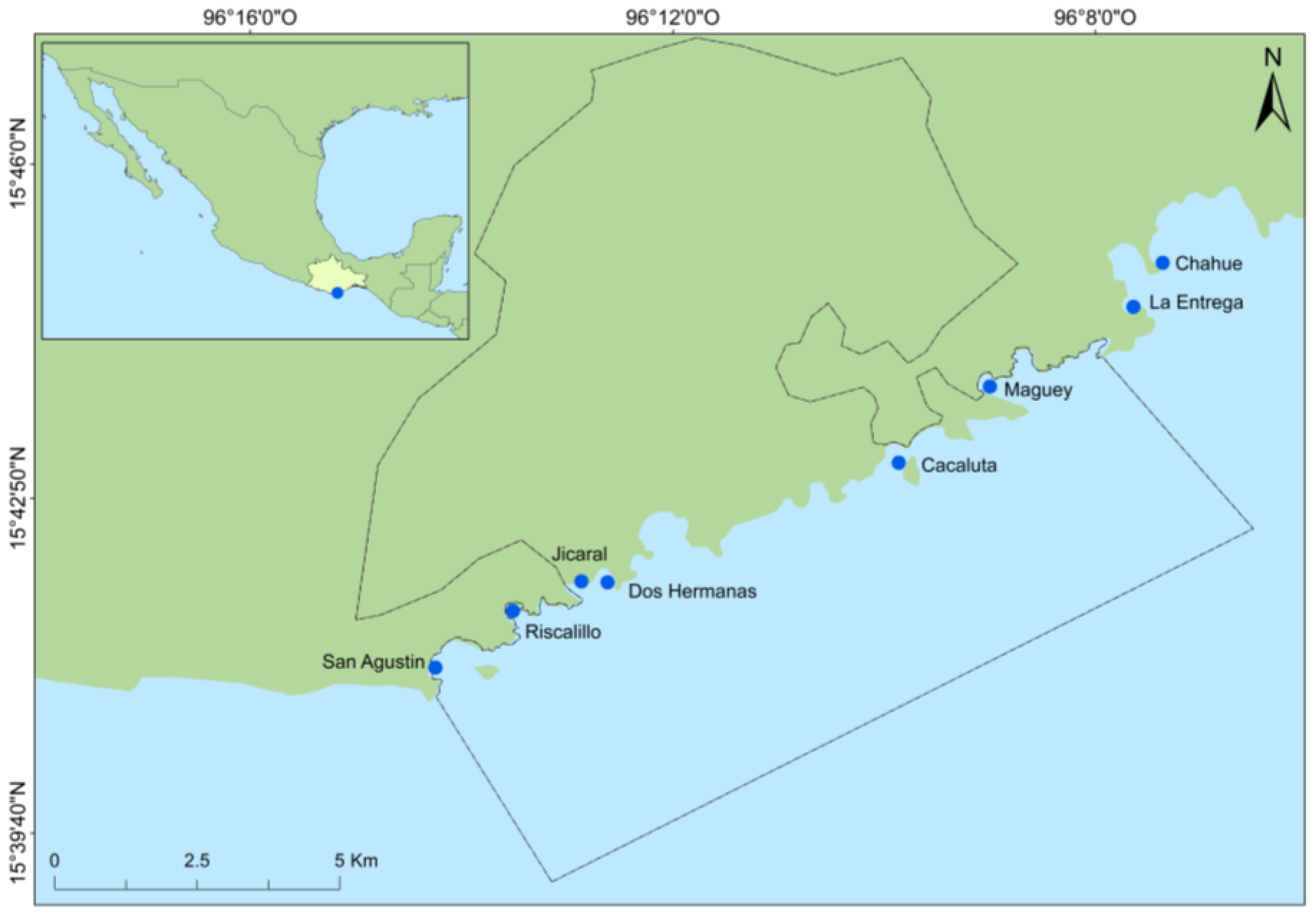
2. Materials and Method
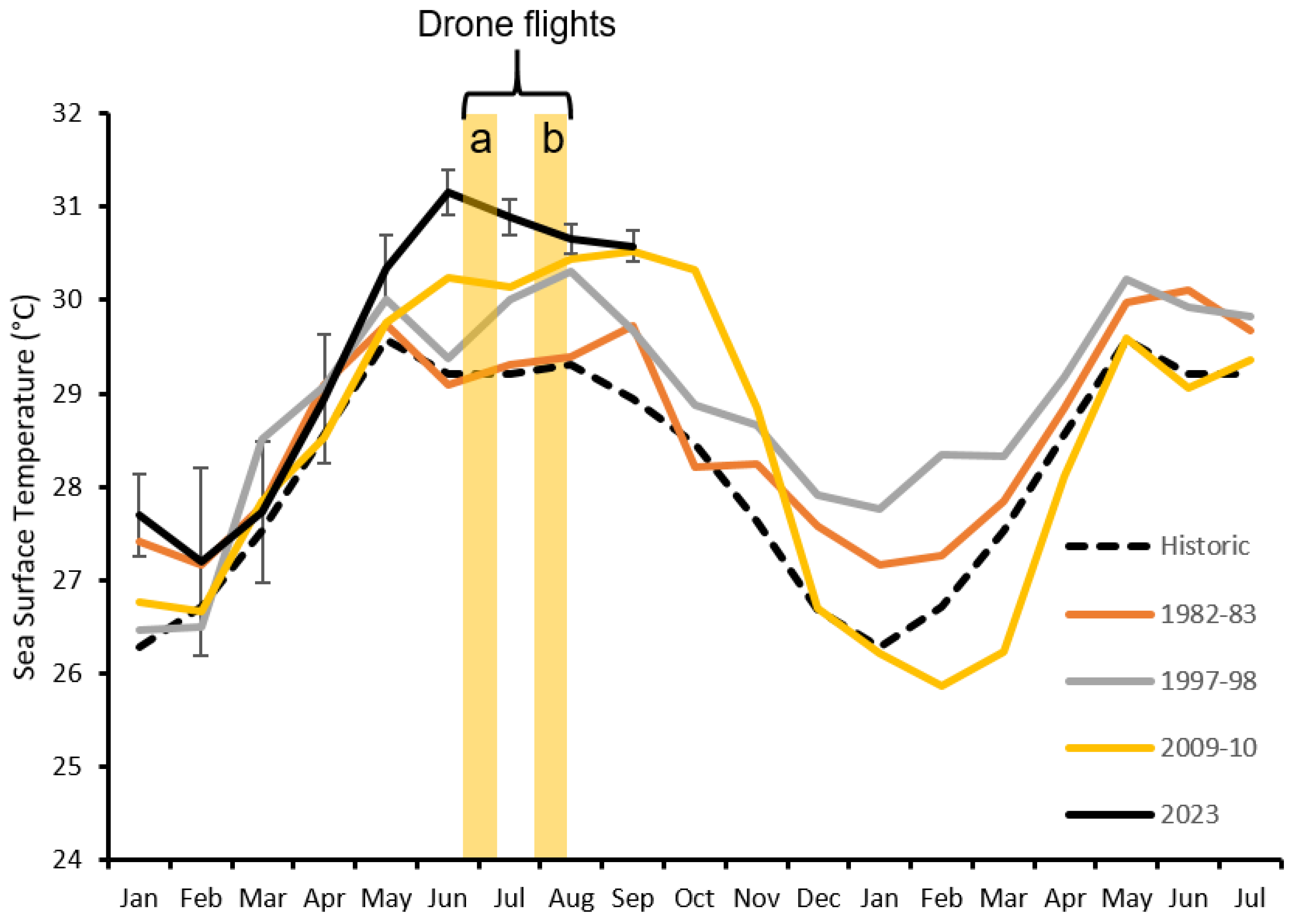
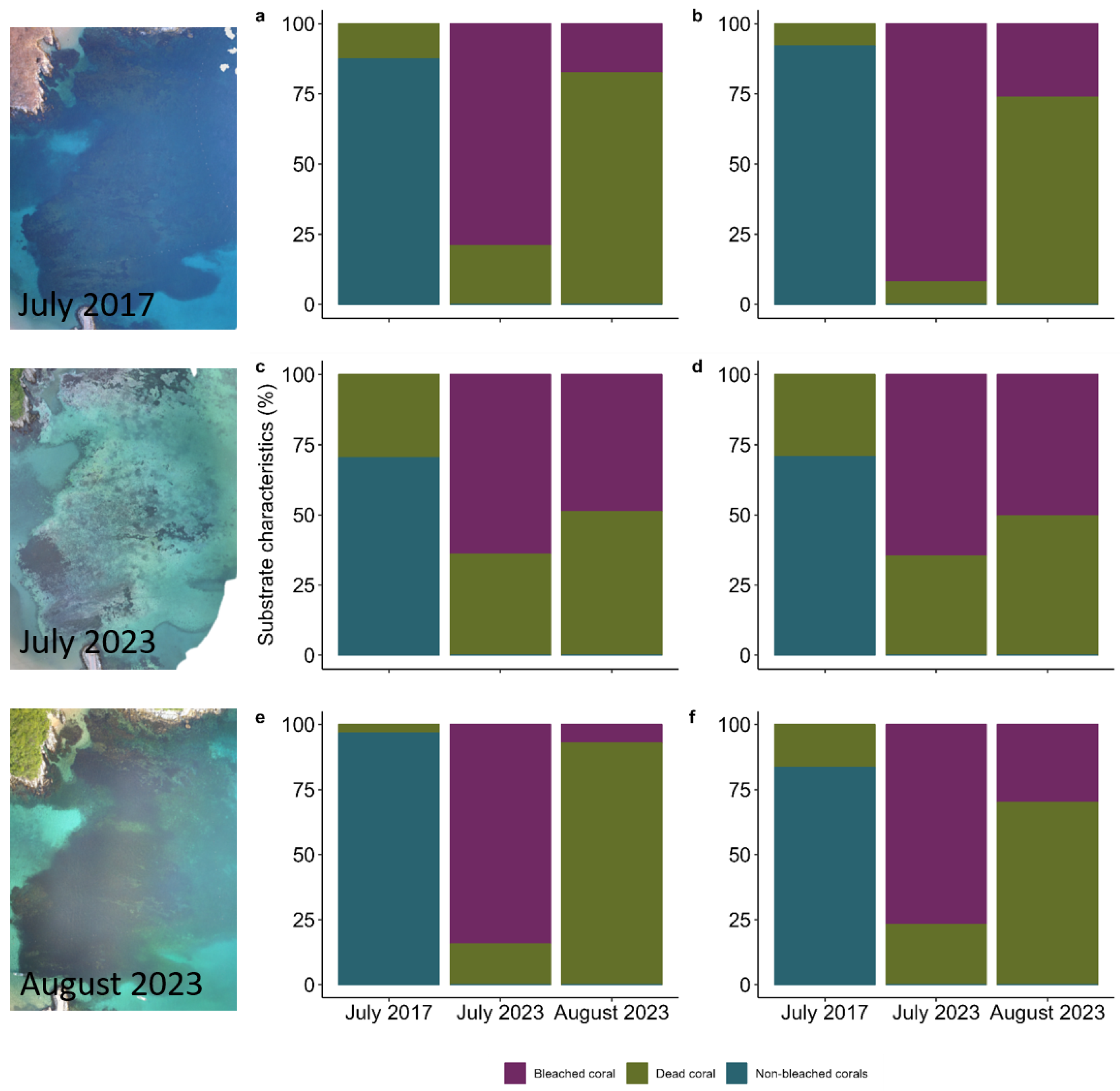
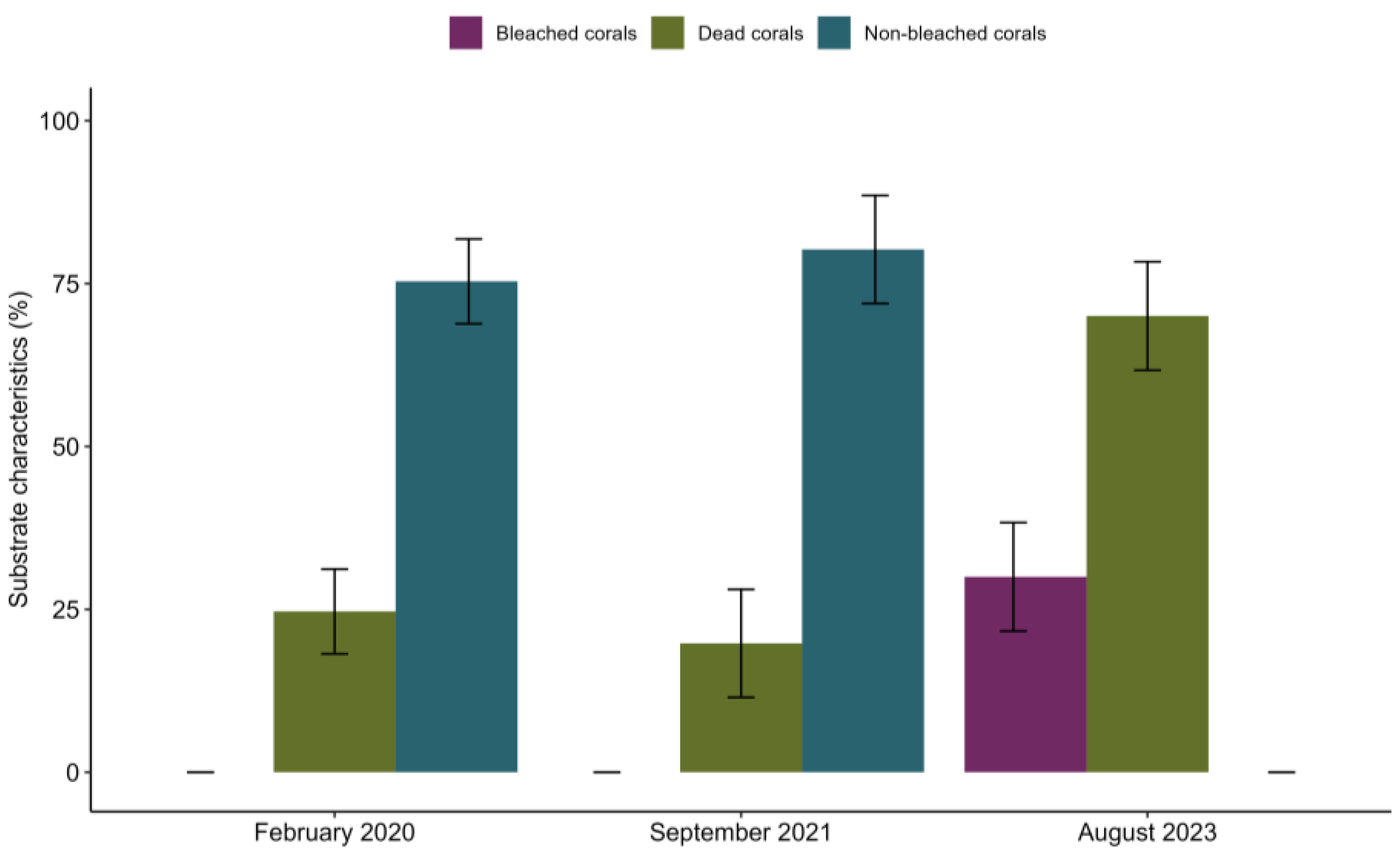
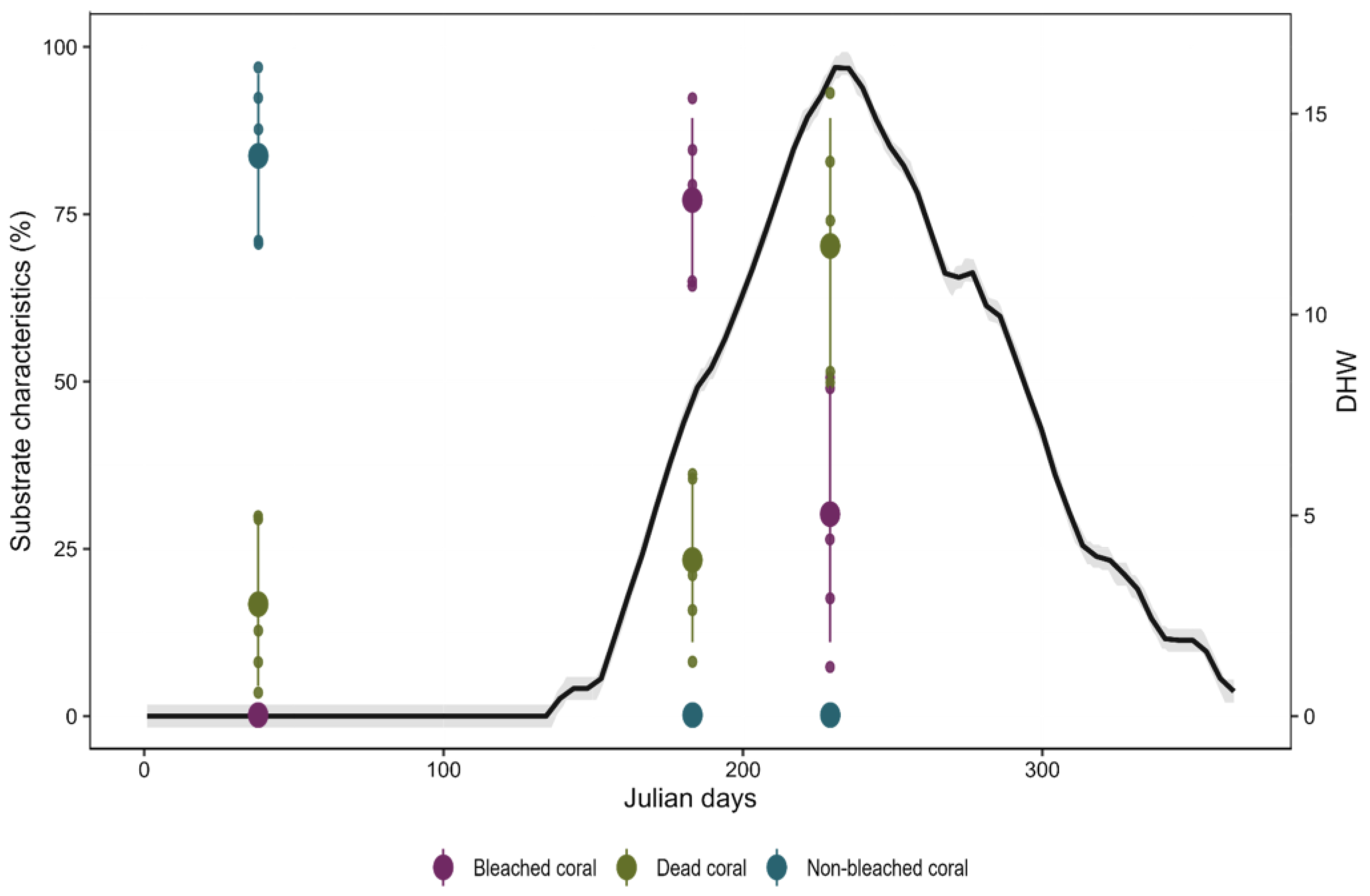
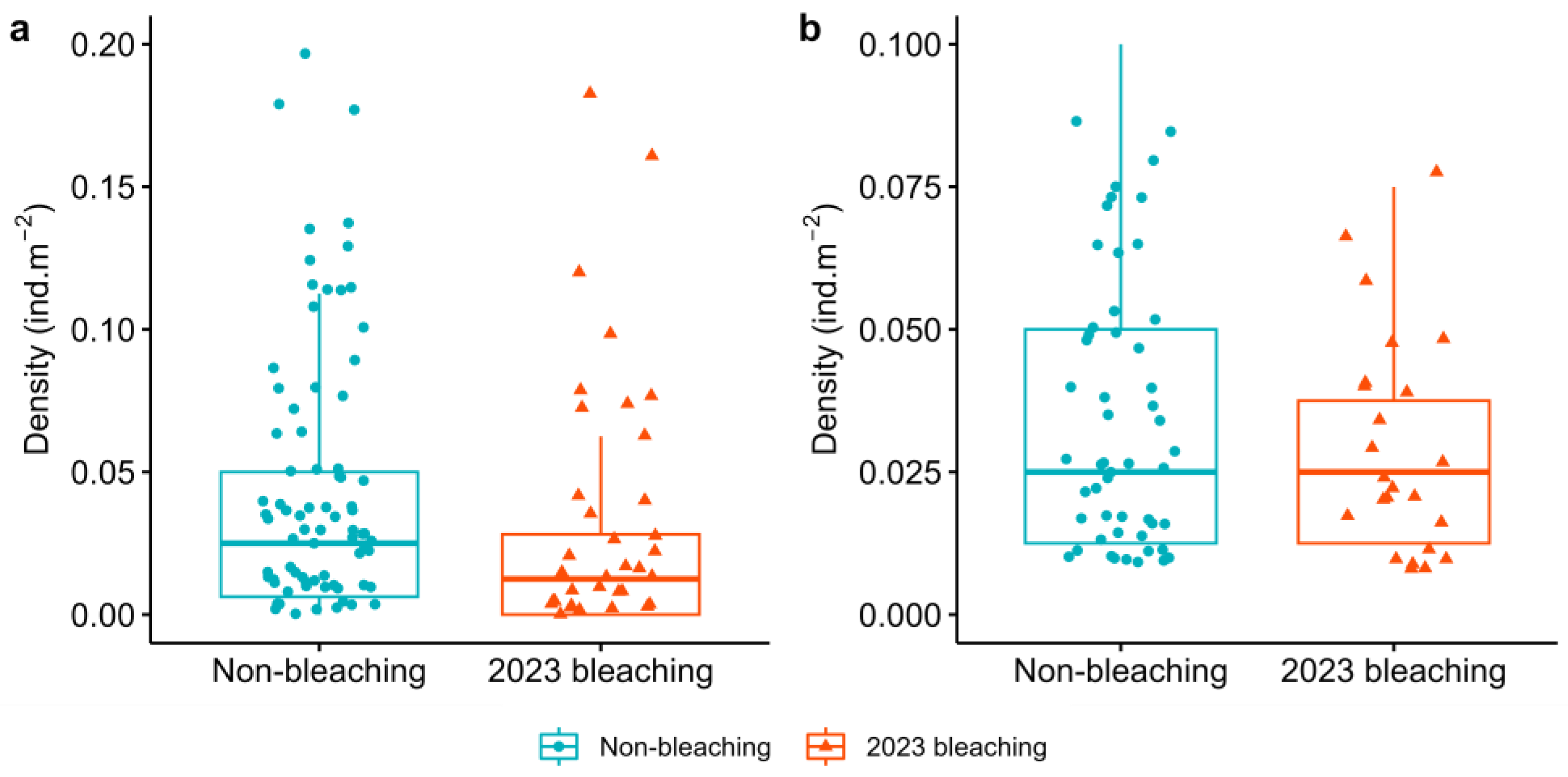
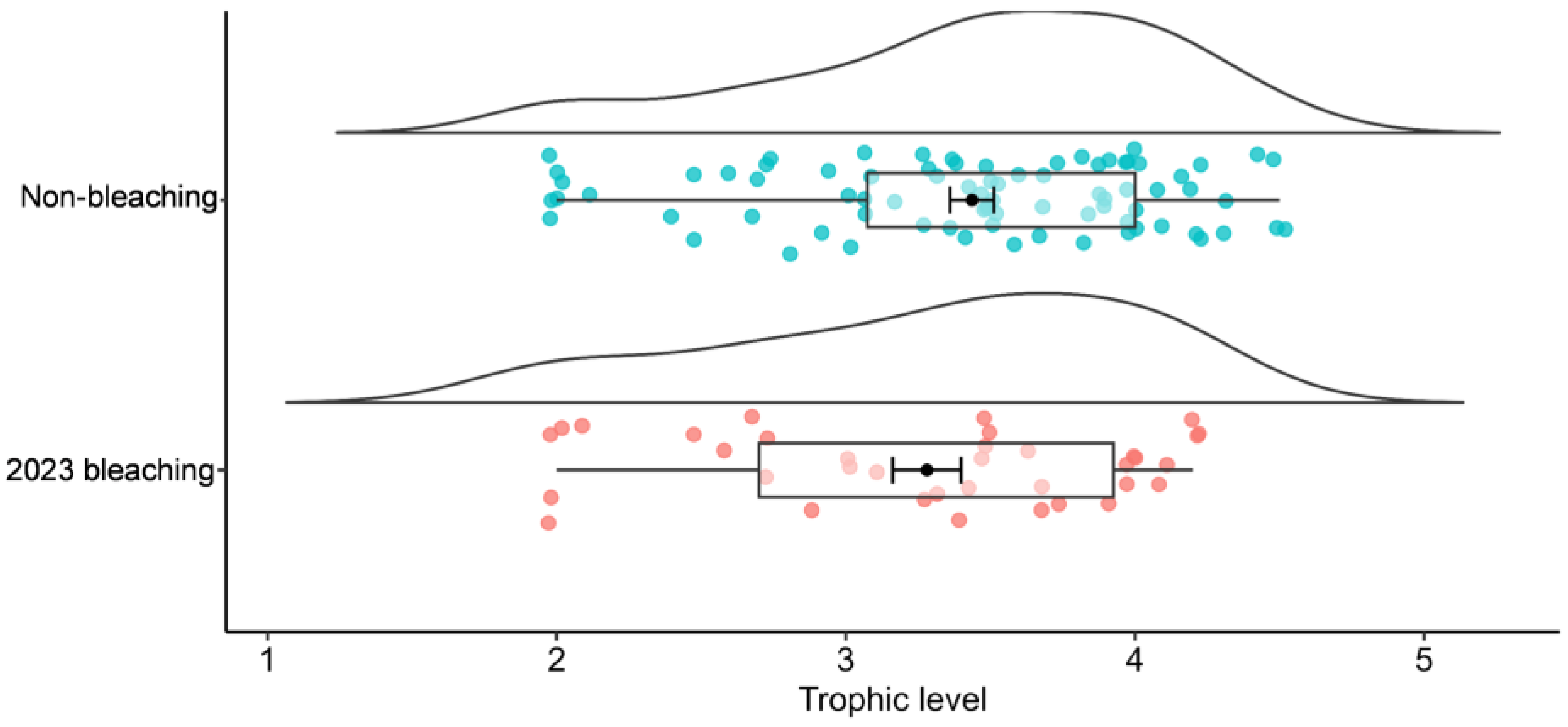
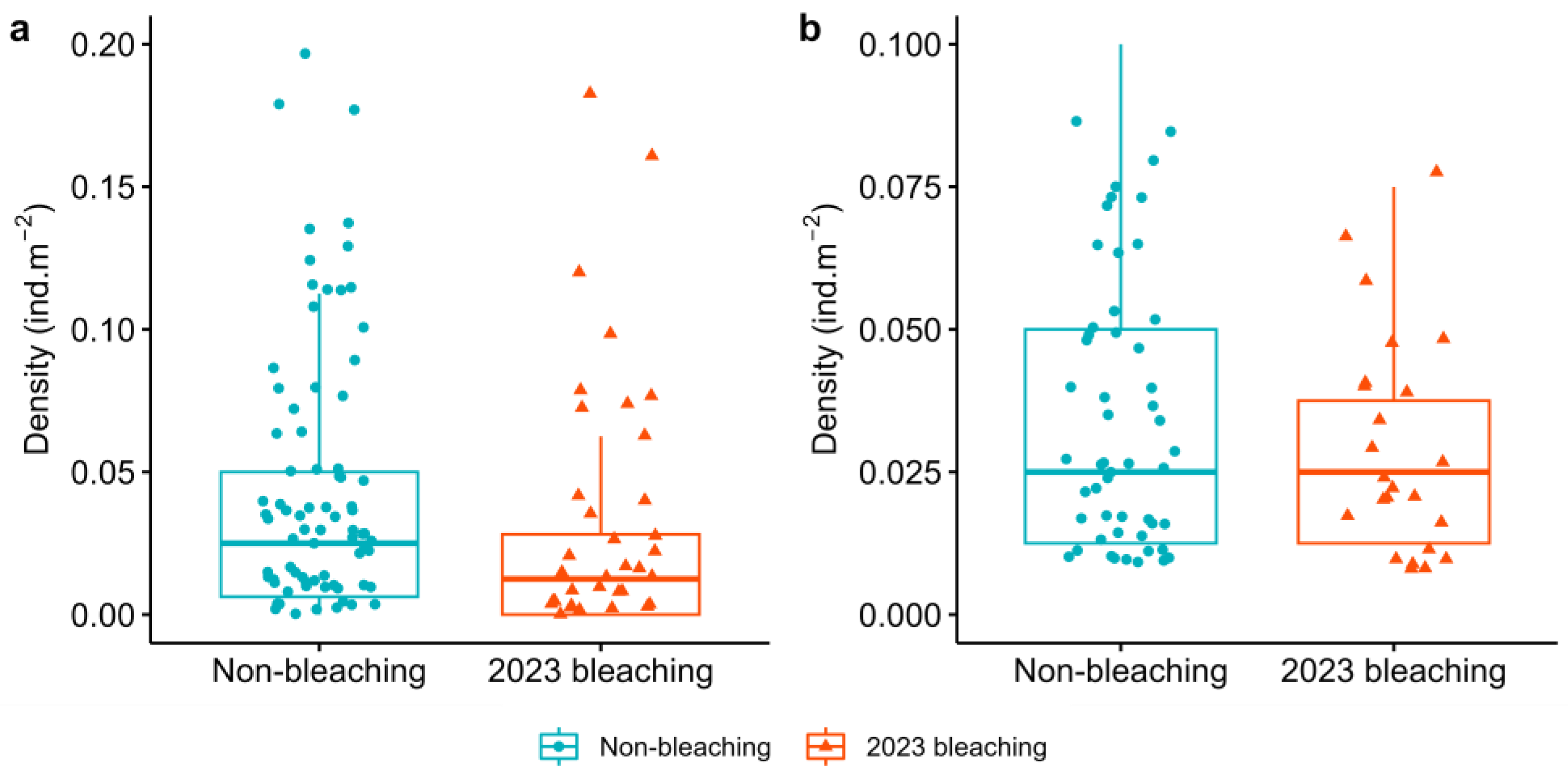
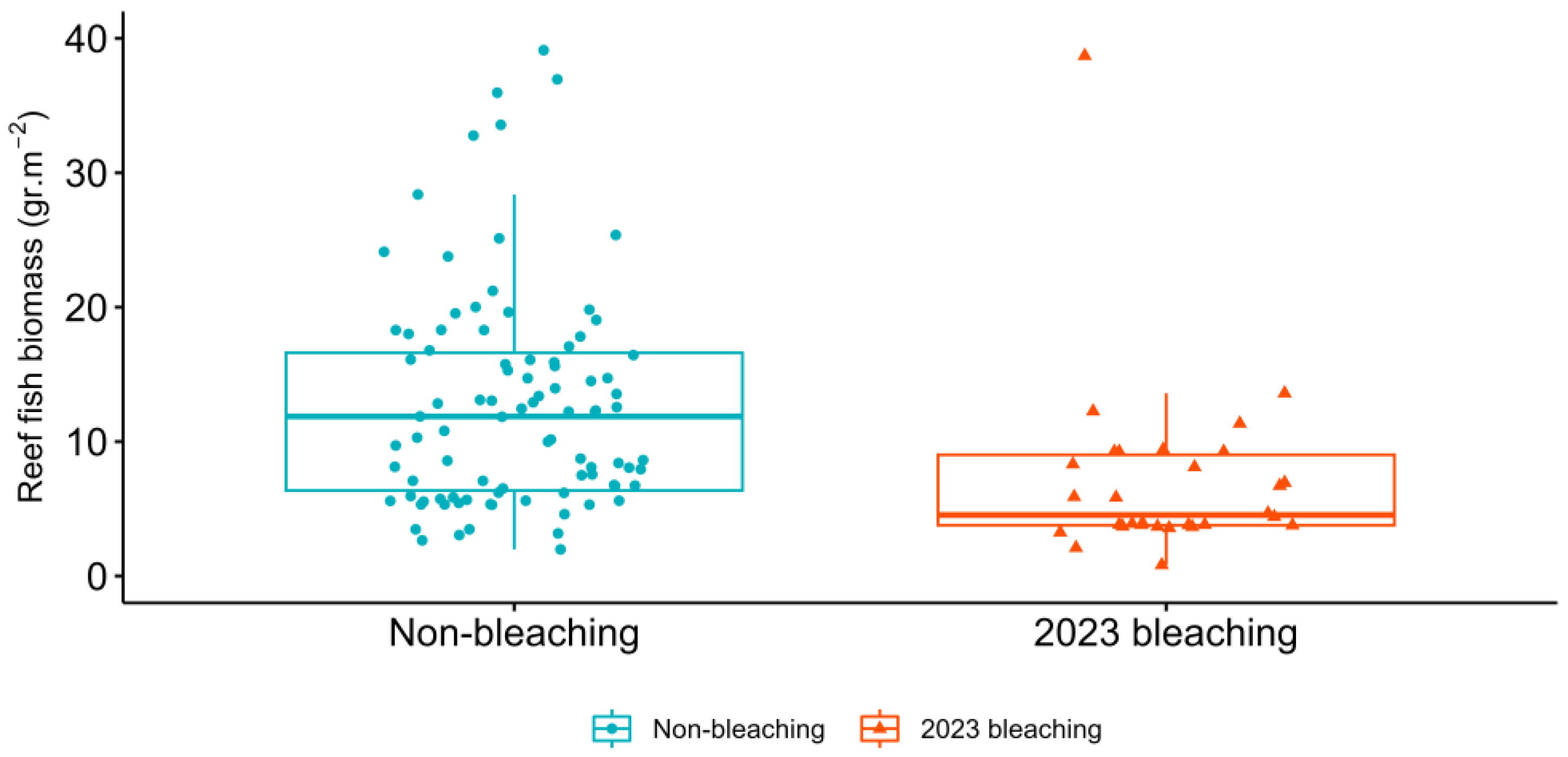
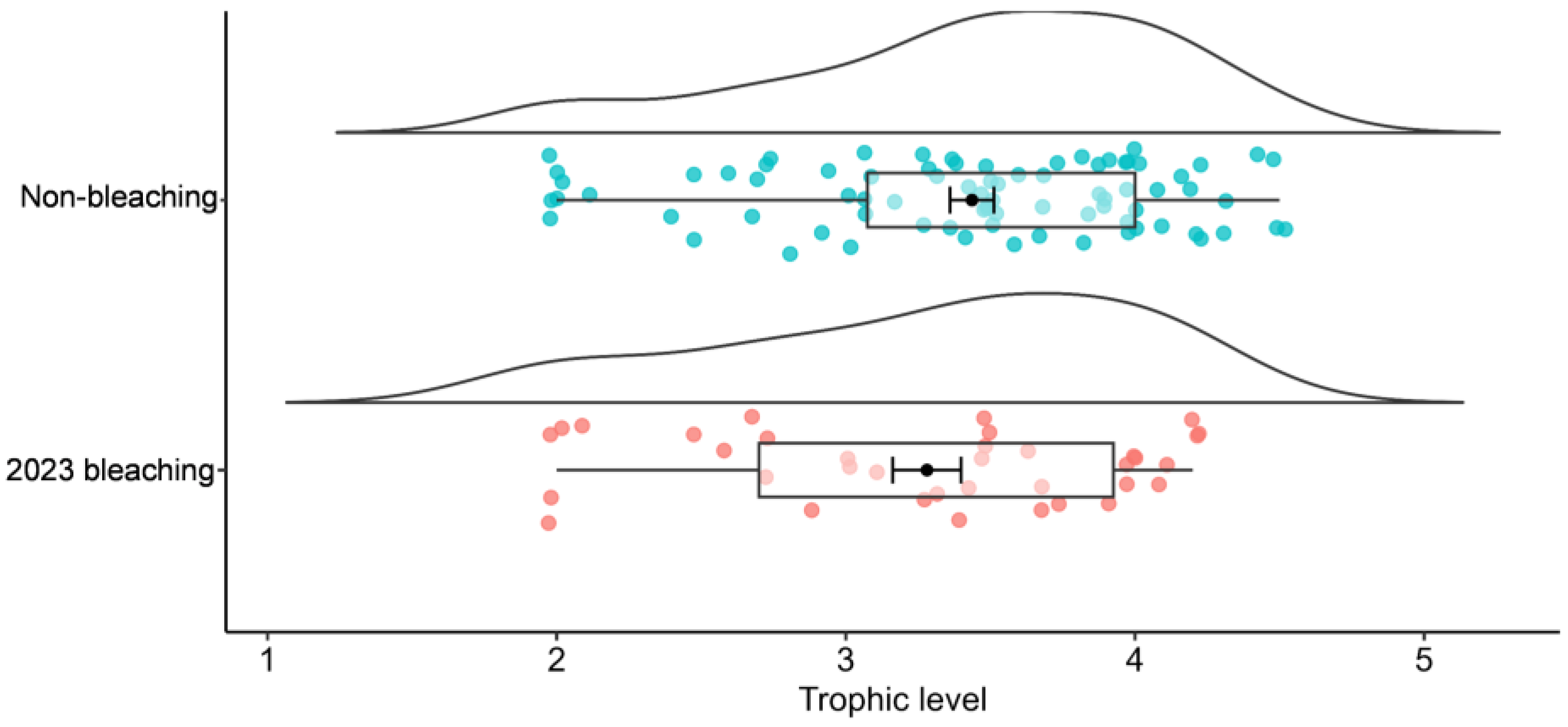
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Carré, M.; Sachs, J.P.; Purca, S.; Schauer, A.J.; Braconnot, P.; Falcón, R.A.; Julien, M.; Lavallée, D. Holocene history of ENSO variance and asymmetry in the eastern tropical Pacific. Science 2014, 345, 1045–1048. [Google Scholar] [CrossRef]
- Clarke, A.J. El Niño physics and El Niño predictability. Annu. Rev. Mar. Sci. 2014, 6, 79–99. [Google Scholar] [CrossRef]
- Mumby, P.J.; Chisholm, J.R.; Edwards, A.J.; Andrefouet, S.; Jaubert, J. Cloudy weather may have saved Society Island reef corals during the 1998 ENSO event. Mar. Ecol. Prog. Ser. 2001, 222, 209–216. [Google Scholar] [CrossRef]
- Zhang, N.; Feng, M.; Hendon, H.H.; Hobday, A.J.; Zinke, J. Opposite polarities of ENSO drive distinct patterns of coral bleaching potentials in the southeast Indian Ocean. Sci. Rep. 2017, 7, 2443. [Google Scholar] [CrossRef]
- Allemand, D.; Tambutté, É.; Zoccola, D.; Tambutté, S. Coral calcification, cells to reefs. In Coral Reefs: An Ecosystem in Transition; Dubinsky, Z., Stambler, N., Eds.; Springer: Dordrecht, The Netherlands, 2011; pp. 119–150. [Google Scholar] [CrossRef]
- Fitt, W.K.; McFarland, F.K.; Warner, M.E.; Chilcoat, G.C. Seasonal patterns of tissue biomass and densities of symbiotic dinoflagellates in reef corals and relation to coral bleaching. Limnol. Oceanogr. 2000, 45, 677–685. [Google Scholar] [CrossRef]
- Douglas, A.E. Coral bleaching––how and why? Mar. Poll. Bull. 2003, 46, 385–392. [Google Scholar] [CrossRef] [PubMed]
- Glynn, P.W.; Mones, A.B.; Podestá, G.P.; Colbert, A.; Colgan, M.W. El Niño-Southern Oscillation: Effects on Eastern Pacific coral reefs and associated biota. In Coral Reefs of the Eastern Tropical Pacific: Persistence and Loss in a Dynamic Environment; Glynn, P.W., Manzello, D.P., Enoch, I.C., Eds.; Springer: Dordrecht, The Netherlands, 2017; pp. 251–290. [Google Scholar] [CrossRef]
- Glynn, P.W. Widespread coral mortality and the 1982–83 El Niño warming event. Environ. Conserv. 1984, 11, 133–146. [Google Scholar] [CrossRef]
- Reyes-Bonilla, H. 1987 coral reef bleaching at Cabo Pulmo reef, Gulf of California, Mexico. Bull. Mar. Sci. 1993, 52, 832–837. [Google Scholar]
- Reyes-Bonilla, H. Effects of the 1997–1998 El Niño–Southern Oscillation on coral communities of the Gulf of California, Mexico. Bull. Mar. Sci. 2001, 69, 251–266. [Google Scholar]
- Iglesias-Prieto, R.; Reyes-Bonilla, H.; Rodríguez, R.R. Effects of 1997-1998 ENSO on coral reef communities in the Gulf of California, Mexico. Geofís. Int. 2003, 42, 467–471. [Google Scholar]
- LaJeunesse, T.C.; Reyes-Bonilla, H.; Warner, M.E. Spring “bleaching” among Pocillopora in the Sea of Cortez, eastern Pacific. Coral Reefs 2007, 26, 265–270. [Google Scholar] [CrossRef]
- Carriquiry, J.D.; Cupul-Magaña, A.L.; Rodríguez-Zaragoza, F.; Medina-Rosas, P. Coral bleaching and mortality in the Mexican Pacific during the 1997–98 El Niño and prediction from a remote sensing approach. Bull. Mar. Sci. 2001, 69, 237–249. [Google Scholar]
- Cruz-García, R.; Rodríguez-Troncoso, A.P.; Rodríguez-Zaragoza, F.A.; Mayfield, A.; Cupul-Magaña, A.L. Ephemeral effects of El Niño–Southern Oscillation events on an eastern tropical Pacific coral community. Mar. Freshw. Res. 2020, 71, 1259–1268. [Google Scholar] [CrossRef]
- López-Pérez, A.; Guendulain-García, S.; Granja-Fernández, R.; Hernández-Urraca, V.; Galván-Rowland, L.; Zepeta-Vilchis, R.; López-López, D. Reef community changes associated with the 2009–2010 El Niño in the southern Mexican Pacific. Pac. Sci. 2016, 70, 175–190. [Google Scholar] [CrossRef]
- Nava, H.; López, N.; Ramírez-García, P.; Garibay-Valladolid, E. Contrasting effects of the El Niño 2015–16 event on coral reefs from the central pacific coast of Mexico. Mar. Ecol. 2021, 42, e12630. [Google Scholar] [CrossRef]
- Manzello, D.P.; Enochs, I.C.; Bruckner, A.; Renaud, P.G.; Kolodziej, G.; Budd, D.A.; Carlton, R.; Glynn, P.W. Galápagos coral reef persistence after ENSO warming across an acidification gradient. Geophys. Res. Lett. 2014, 41, 9001–9008. [Google Scholar] [CrossRef]
- Alvarado, J.J.; Sánchez-Noguera, C.; Arias-Godínez, G.; Araya, T.; Fernández-García, C.; Guzmán, A.G. Impact of El Niño 2015-2016 on the coral reefs of the Pacific of Costa Rica: The potential role of marine protection. Rev. Biol. Trop. 2020, 68, 271–282. [Google Scholar] [CrossRef]
- Glynn, P.W.; Leyte-Morales, G.E. Coral reefs of Huatulco, West Mexico: Reef development in upwelling Gulf of Tehuantepec. Rev. Biol. Trop. 1997, 45, 1033–1047. Available online: https://revistas.ucr.ac.cr/index.php/rbt/article/view/21114 (accessed on 20 September 2023).
- Reyes-Bonilla, H.; Carriquiry, J.; Leyte-Morales, G.; Cupul-Magaña, A. Effects of the El Niño-Southern Oscillation and the anti-El Niño event (1997–1999) on coral reefs of the western coast of México. Coral Reefs 2002, 21, 368–372. [Google Scholar] [CrossRef]
- Horta-Puga, G.; Gutiérrez-Martínez, J.; Morales-Aranda, A.A.; Victoria-Muguira, M. Mass bleaching in SW Gulf of Mexico. Reef Encount. 2023, 38, 59–63. [Google Scholar]
- Fiedler, P.C.; Talley, L.D. Hydrography of the eastern tropical Pacific: A review. Prog. Oceanogr 2006, 69, 143–180. [Google Scholar] [CrossRef]
- Trasviña, A.; Barton, E.D.; Brown, J.; Velez, H.S.; Kosro, P.M.; Smith, R.L. Offshore wind forcing in the Gulf of Tehuantepec, Mexico: The asymmetric circulation. J. Geophys. Res. Oceans 1995, 100, 20649–20663. [Google Scholar] [CrossRef]
- Skirving, W.; Marsh, B.; De La Cour, J.; Liu, G.; Harris, A.; Maturi, E.; Geiger, E.; Eakin, C.M. CoralTemp and the Coral Reef Watch Coral Bleaching Heat Stress Product Suite Version 3.1. Remote Sens. 2020, 12, 3856. [Google Scholar] [CrossRef]
- Westoby, M.J.; Brasington, J.; Glasser, N.F.; Hambrey, M.J.; Reynolds, J.M. ‘Structure-from-Motion’photogrammetry: A low-cost, effective tool for geoscience applications. Geomorphology 2012, 179, 300–314. [Google Scholar] [CrossRef]
- Froese, R.; Pauly, D. FishBase. 2019. Available online: http://www.fishbase.org (accessed on 20 September 2023).
- Lin, H.Y.; Costello, M.J. Body size and trophic level increase with latitude, and decrease in the deep-sea and Antarctica, for marine fish species. PeerJ 2023, 11, e15880. [Google Scholar] [CrossRef] [PubMed]
- Zar, J.H. Biostatistical Analysis, 5th ed.; Prentice Hall: Upper Saddle River, NJ, USA, 2010; p. 944. [Google Scholar]
- RStudio Team. RStudio: Integrated Development for R. RStudio; PBC: Boston, MA, USA, 2020; Available online: http://www.rstudio.com/ (accessed on 20 September 2023).
- Jones, R.J.; Hoegh-Guldberg, O.; Larkum, A.W.D.; Schreiber, U. Temperature-induced bleaching of corals begins with impairment of the CO2 fixation mechanism in zooxanthellae. Plant Cell Environ. 1998, 21, 1219–1230. [Google Scholar] [CrossRef]
- Warner, M.E.; Fitt, W.K.; Schmidt, G.W. Damage to photosystem II in symbiotic dinoflagellates: A determinant of coral bleaching. Proc. Natl. Acad. Sci. USA 1999, 96, 8007–8012. [Google Scholar] [CrossRef] [PubMed]
- Lesser, M.P.; Farrell, J.H. Exposure to solar radiation increases damage to both host tissues and algal symbionts of corals during thermal stress. Coral Reefs 2004, 23, 367–377. [Google Scholar] [CrossRef]
- Palacio-Castro, A.; Smith, T.; Brandtneris, V.; Snyder, G.; van Hooidonk, R.; Mate, J.; Manzello, D.P.; Glynn, P.; Fong, P.; Baker, A. Increased dominance of heat-tolerant symbionts create resilient coral reefs in nearterm ocean warming. Proc. Natl. Acad. Sci. USA 2023, 120, e2202388120. [Google Scholar] [CrossRef]
- LaJeunesse, T.C.; Smith, R.; Walther, M.; Pinzón, J.; Pettay, D.T.; McGinley, M.; Aschaffenburg, M.; Medina-Rosas, P.; Cupul-Magaña, A.L.; Pérez, A.L.; et al. Host–symbiont recombination versus natural selection in the response of coral–dinoflagellate symbioses to environmental disturbance. Proc. R. Soc. B Biol. Sci. 2010, 277, 2925–2934. [Google Scholar] [CrossRef]
- Alvarado, J.J.; Reyes-Bonilla, H.; Benítez-Villalobos, F. Diadema mexicanum, erizo de mar clave en los arrecifes coralinos del Pacífico Tropical Oriental: Lo que sabemos y perspectivas futuras (Diadematoida: Diadematidae). Rev. Biol. Trop. 2015, 63, 135–157. [Google Scholar]
- Zamorano, P.; Leyte-Morales, G. Densidad poblacional de cuatro especies de erizo (Echinodermata: Echinoidea) en el arrecife de La Entrega, Oaxaca. CICIMAR Ocenánides 2005, 20, 65–72. [Google Scholar] [CrossRef]
- Alvarado, J.J.; Cortés, J.; Guzman, H.; Reyes-Bonilla, H. Density, size, and biomass of Diadema mexicanum (Echinoidea) in Eastern Tropical Pacific coral reefs. Aquat. Biol. 2016, 24, 151–161. [Google Scholar] [CrossRef]
- Herrera-Escalante, T.; López-Pérez, R.A.; Leyte-Morales, G.E. Bioerosion caused by the sea urchin Diadema mexicanum (Echinodermata: Echinoidea) at Bahías de Huatulco, Western Mexico. Rev. Biol. Trop. 2005, 53, 263–273. [Google Scholar] [PubMed]
- Glynn, P.W. El Niño warming, coral mortality and reef framework destruction by echinoid bioerosion in the eastern Pacific. Galaxea 1988, 7, 129–160. [Google Scholar]
- Benítez-Villalobos, F.; Díaz-Martínez, J.P.; Martínez-García, M. Mass mortality of the sea urchin Diadema mexicanum in La Entrega at Bahias de Huatulco, Western Mexico. Coral Reefs 2009, 28, 1017. [Google Scholar] [CrossRef]
- Hickman, C.P., Jr. A Field Guide to Sea Stars and Other Echinoderms of Galápagos; Sugar Spring Press: Lexington, KY, USA, 1998; p. 83. [Google Scholar]
- Glynn, P.W.; Wellington, G.M.; Birkeland, C. Coral reef growth in the Galapagos: Limitation by sea urchins. Science 1979, 203, 47–49. [Google Scholar] [CrossRef] [PubMed]
- Hoegh-Guldberg, O.; Mumby, P.J.; Hooten, A.J.; Steneck, R.S.; Greenfield, P.; Gomez, E.; Harvell, C.D.; Sale, P.F.; Edwards, A.J.; Caldeira, K.; et al. Coral reefs under rapid climate change and ocean acidification. Science 2007, 318, 1737–1742. [Google Scholar] [CrossRef] [PubMed]
- Hempson, T.N.; Graham, N.A.; MacNeil, M.A.; Hoey, A.S.; Wilson, S.K. Ecosystem regime shifts disrupt trophic structure. Ecol. Appl. 2018, 28, 191–200. [Google Scholar] [CrossRef]
- Glynn, P.W.; De Weerdt, W.H. Elimination of two reef-building hydrocorals following the 1982-83 El Niño warming event. Science 1991, 253, 69–71. [Google Scholar] [CrossRef]
- Lluch-Belda, D.; Lluch-Belda, D.B.; Lluch-Cota, S.E. Changes in marine faunal distributions and ENSO events in the California Current. Fish. Oceanogr. 2005, 14, 458–467. [Google Scholar] [CrossRef]
- Leyte-Morales, G.E.; Reyes-Bonilla, H.; Cintra-Buenrostro, C.E.; Glynn, P.W. Range extension of Leptoseris papyracea (Dana, 1846) to the west coast of Mexico. Bull. Mar. Sci. 2001, 69, 1233–1237. [Google Scholar]
- Osland, M.J.; Stevens, P.W.; Lamont, M.M.; Brusca, R.C.; Hart, K.M.; Waddle, J.H.; Langtimm, C.A.; Williams, C.M.; Keim, B.D.; Terando, A.J.; et al. Tropicalization of temperate ecosystems in North America: The northward range expansion of tropical organisms in response to warming winter temperatures. Glob. Chang. Biol. 2021, 27, 3009–3034. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Velasco, L.; Beier, E.; Godínez, V.M.; Barton, E.D.; Santamaría-del-Angel, E.; Jiménez-Rosemberg, S.P.A.; Marinone, S.G. Hydrographic and fish larvae distribution during the “Godzilla El Niño 2015–2016” in the northern end of the shallow oxygen minimum zone of the Eastern Tropical Pacific Ocean. J. Geophys. Res. Oceans 2017, 122, 2156–2170. [Google Scholar] [CrossRef]
- Engle, J.M.; Richards, D.V. New and unusual marine invertebrates discovered at the California Channel Islands during the 1997–1998 El Niño. Bull. South. Calif. Acad. Sci. 2001, 100, 186–198. [Google Scholar]
- Medellín-Maldonado, F.; Cruz-Ortega, I.; Pérez-Cervantes, E.; Norzogaray-López, O.; Carricart-Ganivet, J.P.; López-Pérez, A.; Alvarez-Filip, L. Newly deceased Caribbean reef-building corals experience rapid carbonate loss and colonization by endolithic organisms. Commun. Biol. 2023, 6, 934. [Google Scholar] [CrossRef]
- Alvarez-Filip, L.; Dulvy, N.K.; Gill, J.A.; Côté, I.M.; Watkinson, A.R. Flattening of Caribbean coral reefs: Region-wide declines in architectural complexity. Proc. R. Soc. B Biol. Sci. 2009, 276, 3019–3025. [Google Scholar] [CrossRef]
- Perry, C.T.; Alvarez-Filip, L. Changing geo-ecological functions of coral reefs in the Anthropocene. Funct. Ecol. 2019, 33, 976–988. [Google Scholar] [CrossRef]
- Williams, G.J.; Graham, N.A. Rethinking coral reef functional futures. Funct. Ecol. 2019, 33, 942–947. [Google Scholar] [CrossRef]
- González-Barrios, F.J.; Álvarez-Filip, L. A framework for measuring coral species-specific contribution to reef functioning in the Caribbean. Ecol. Indic. 2018, 95, 877–886. [Google Scholar] [CrossRef]
- Lequeux, B.D.; Ahumada-Sempoal, M.A.; López-Pérez, A.; Reyes-Hernández, C. Coral connectivity between equatorial eastern Pacific marine protected areas: A biophysical modeling approach. PLoS ONE 2018, 13, e0202995. [Google Scholar] [CrossRef] [PubMed]
- Suggett, D.J.; Smith, D.J. Coral bleaching patterns are the outcome of complex biological and environmental networking. Glob. Chang. Biol. 2020, 26, 68–79. [Google Scholar] [CrossRef] [PubMed]
- Guzman, H.M.; Cortés, J. Reef recovery 20 years after the 1982–1983 El Niño massive mortality. Mar. Biol. 2007, 151, 401–411. [Google Scholar] [CrossRef]
- Valdez-Rojas, C.; Beas-Luna, R.; Lorda, J.; Zepeda-Domínguez, J.A.; Montáño-Moctezuma, G.; Medellín-Ortíz, A.; Torre, J.; Micheli, F. Using a social-ecological systems perspective to identify context specific actions to build resilience in small scale fisheries in Mexico. Front. Mar. Sci. 2022, 9, 904859. [Google Scholar] [CrossRef]



{kind=link}
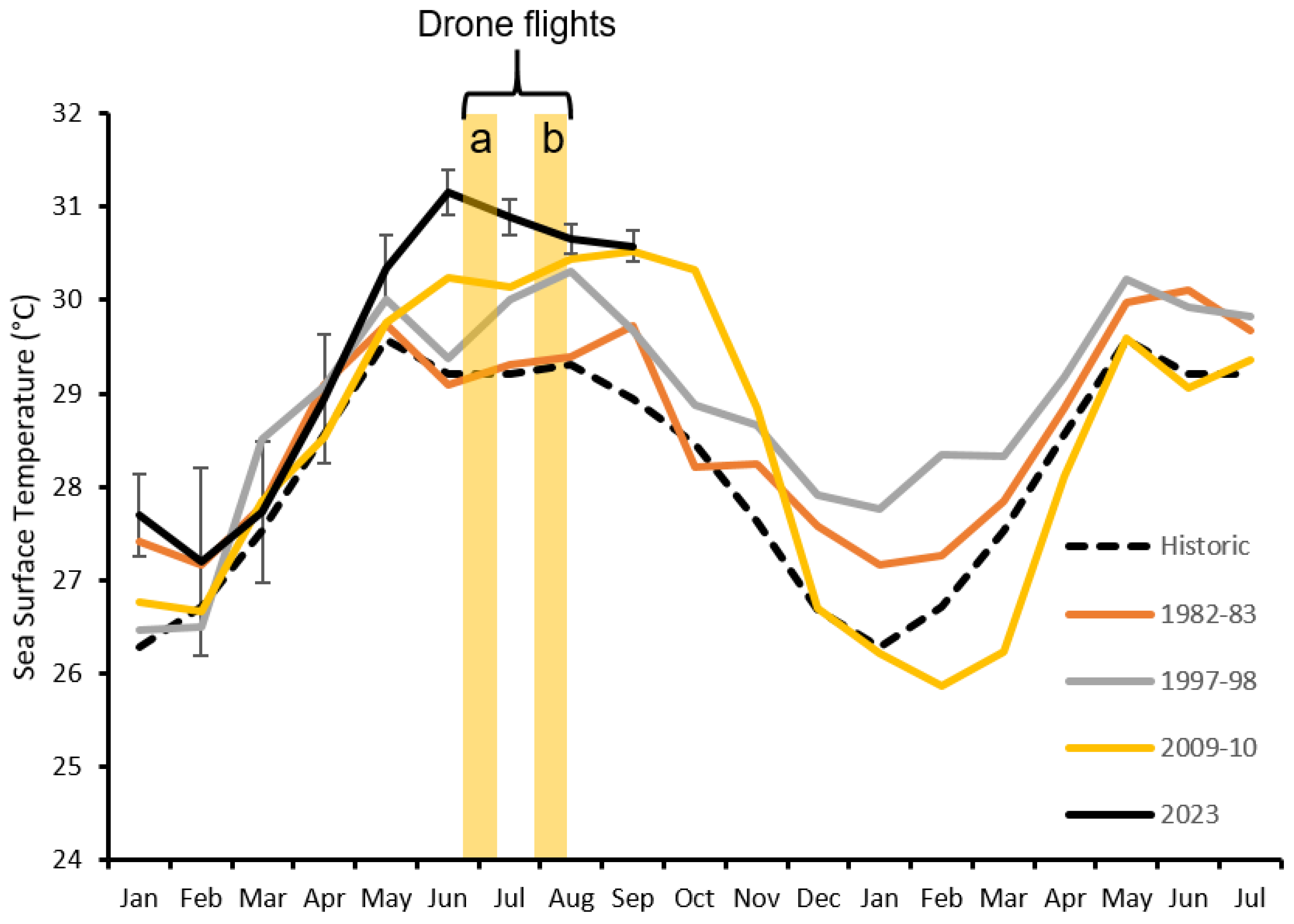{kind=link}
{kind=link}
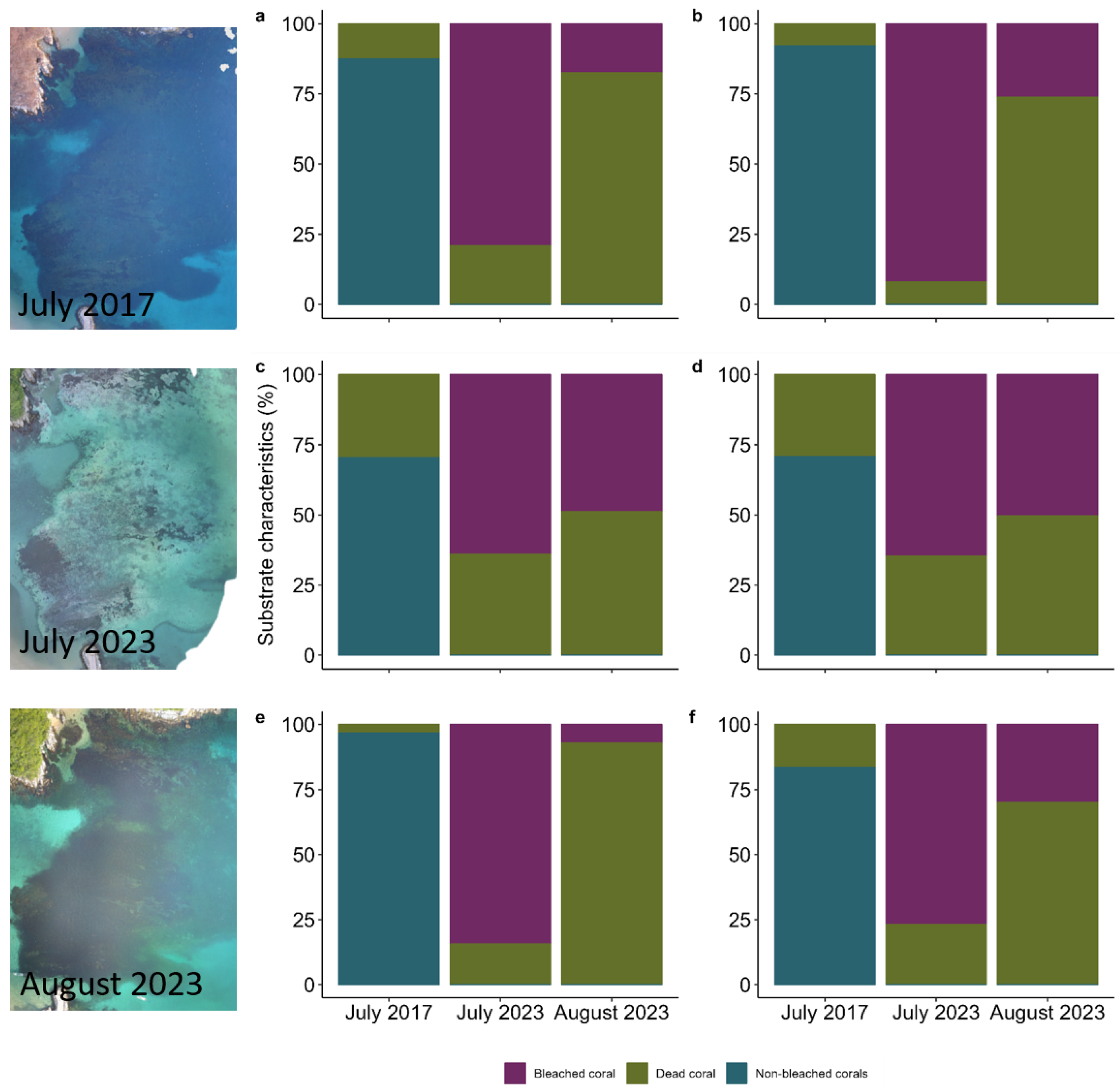{kind=link}
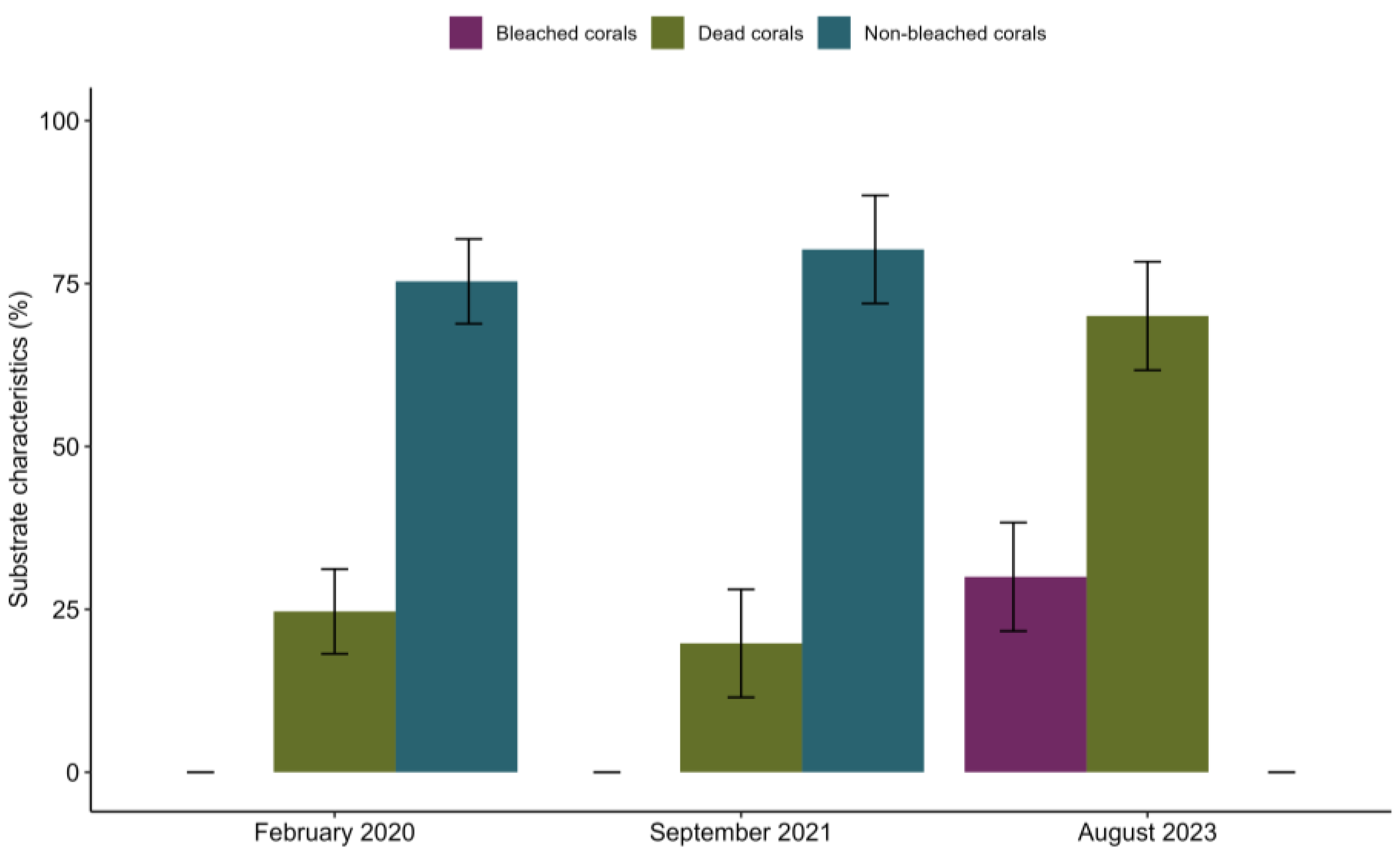{kind=link}
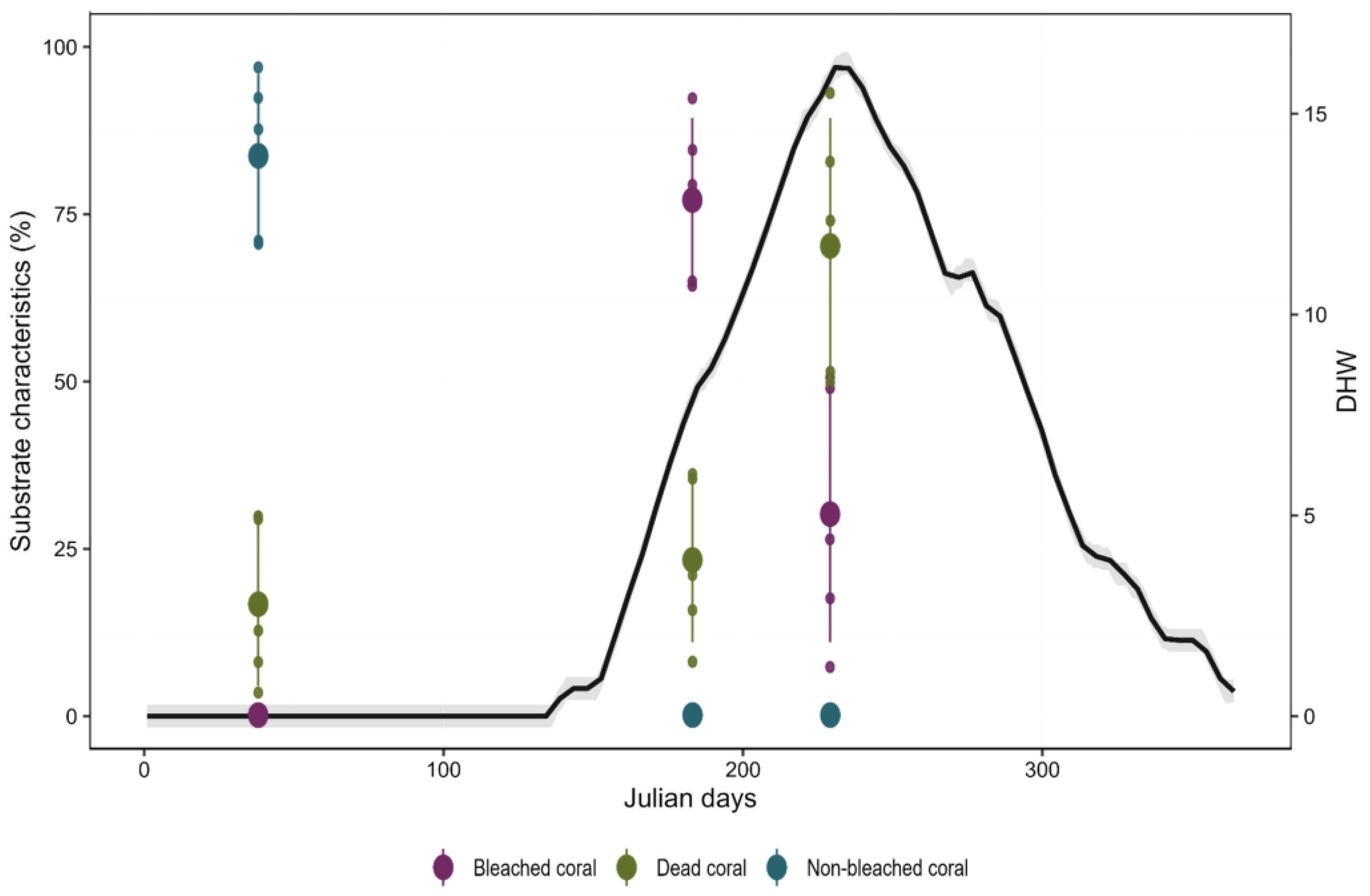{kind=link}
{kind=link}
{kind=link}
{kind=link}
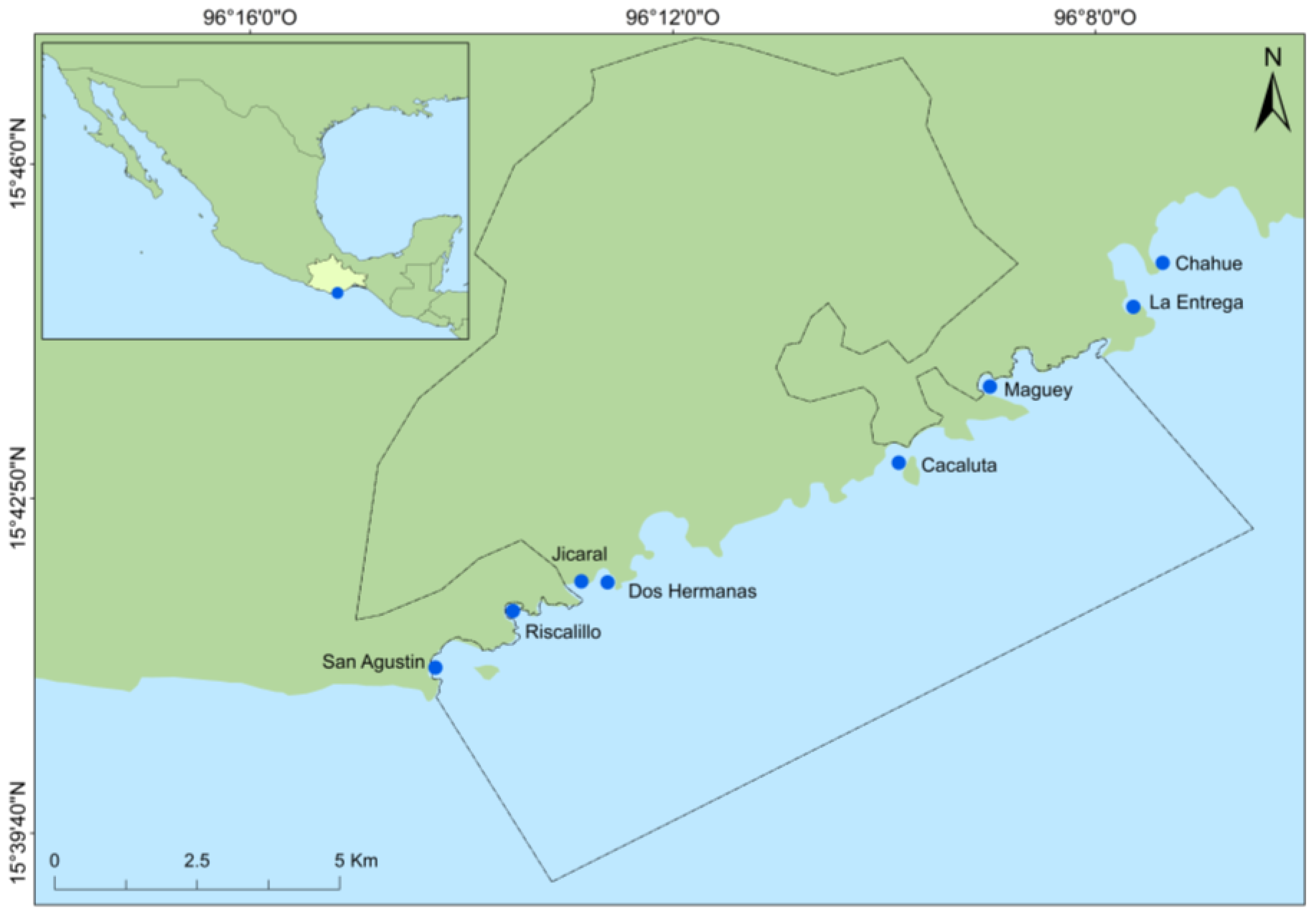
| Site | Drone | Transect |
|---|---|---|
| San Agustín | June–July 2017, 27 June–2 July, 14–18 August | 27 June–2 July, 17–22 August |
| Riscalillo | June–July 2017, 27 June–2 July, 14–18 August | 27 June–2 July, 17–22 August |
| Jicaral | June–July 2017, 27 June–2 July, 14–18 August | 27 June–2 July, 17–22 August |
| Dos Hermanas | June–July 2017, 27 June–2 July, 14–18 August | 27 June–2 July, 17–22 August |
| Cacaluta | June–July 2017, 27 June–2 July, 14–18 August | 27 June–2 July, 17–22 August |
| Maguey | June–July 2017, 27 June–2 July, 14–18 August | 27 June–2 July, 17–22 August |
| La Entrega | June–July 2017, 27 June–2 July, 14–18 August | 27 June–2 July, 17–22 August |
| Chahué | June–July 2017, 27 June–2 July, 14–18 August | 27 June–2 July, 17–22 August |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
López-Pérez, A.; Granja-Fernández, R.; Ramírez-Chávez, E.; Valencia-Méndez, O.; Rodríguez-Zaragoza, F.A.; González-Mendoza, T.; Martínez-Castro, A. Widespread Coral Bleaching and Mass Mortality of Reef-Building Corals in Southern Mexican Pacific Reefs Due to 2023 El Niño Warming. Oceans 2024, 5, 196-209. https://doi.org/10.3390/oceans5020012
López-Pérez A, Granja-Fernández R, Ramírez-Chávez E, Valencia-Méndez O, Rodríguez-Zaragoza FA, González-Mendoza T, Martínez-Castro A. Widespread Coral Bleaching and Mass Mortality of Reef-Building Corals in Southern Mexican Pacific Reefs Due to 2023 El Niño Warming. Oceans. 2024; 5(2):196-209. https://doi.org/10.3390/oceans5020012
Chicago/Turabian StyleLópez-Pérez, Andrés, Rebeca Granja-Fernández, Eduardo Ramírez-Chávez, Omar Valencia-Méndez, Fabián A. Rodríguez-Zaragoza, Tania González-Mendoza, and Armando Martínez-Castro. 2024. "Widespread Coral Bleaching and Mass Mortality of Reef-Building Corals in Southern Mexican Pacific Reefs Due to 2023 El Niño Warming" Oceans 5, no. 2: 196-209. https://doi.org/10.3390/oceans5020012
APA StyleLópez-Pérez, A., Granja-Fernández, R., Ramírez-Chávez, E., Valencia-Méndez, O., Rodríguez-Zaragoza, F. A., González-Mendoza, T., & Martínez-Castro, A. (2024). Widespread Coral Bleaching and Mass Mortality of Reef-Building Corals in Southern Mexican Pacific Reefs Due to 2023 El Niño Warming. Oceans, 5(2), 196-209. https://doi.org/10.3390/oceans5020012