

Evidence of Sexual Reproduction in Out-Planted Coral Colonies




Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
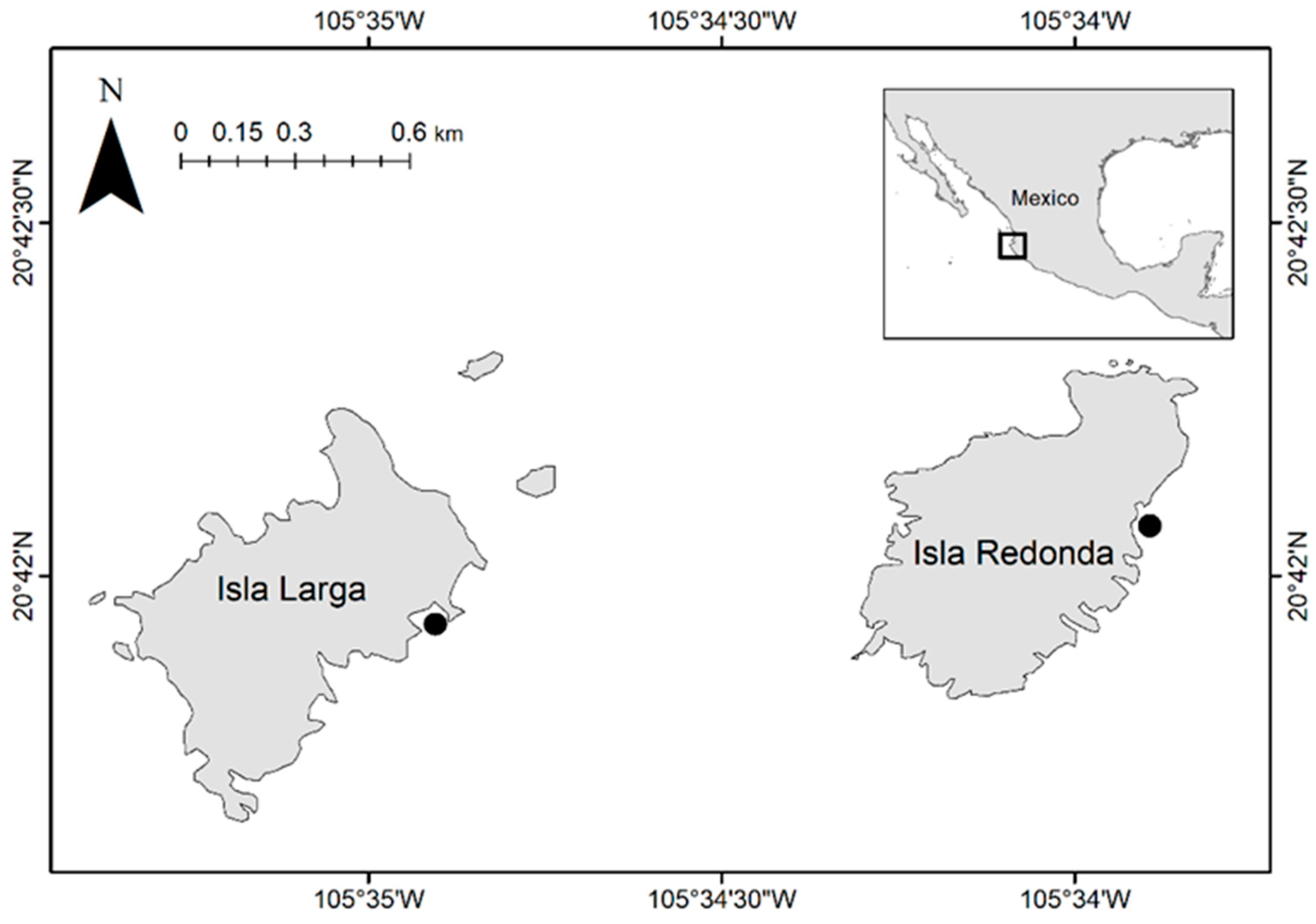
2. Materials and Method
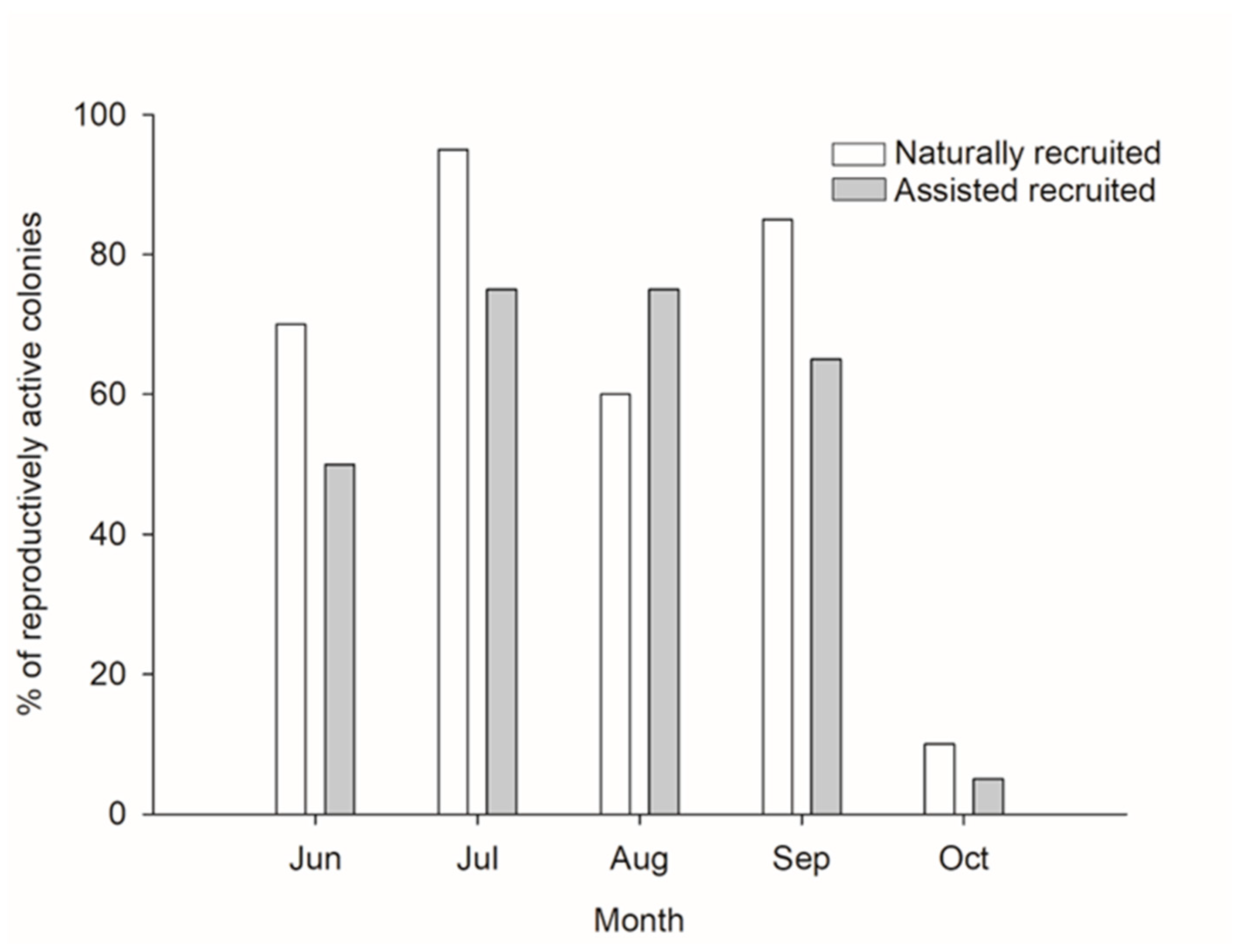
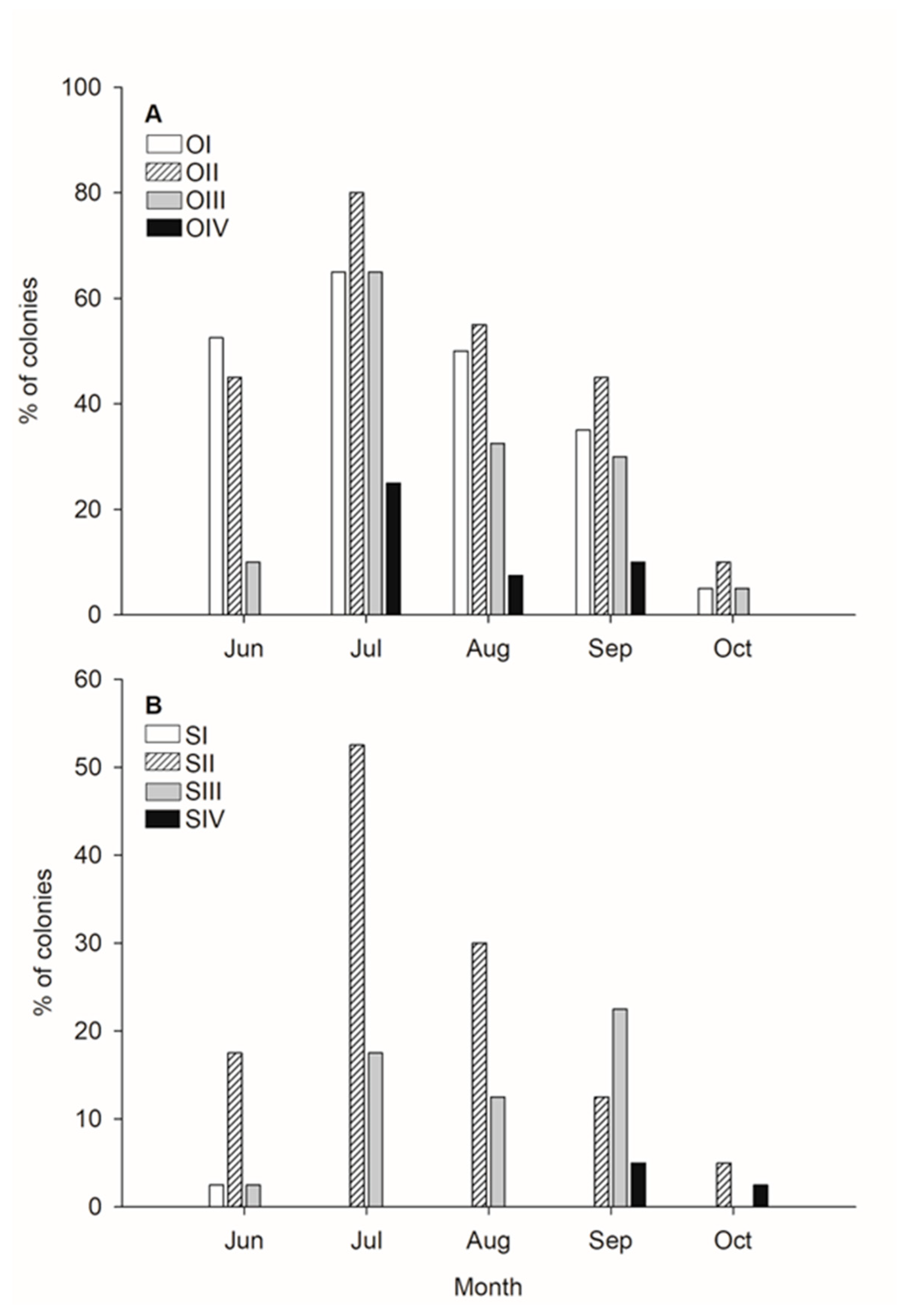
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hoegh-Guldberg, O. Coral reef ecosystems and anthropogenic climate change. Reg. Environ. Chang. 2011, 11, S215–S227. [Google Scholar] [CrossRef]
- Graham, N.A.J.; Bellwood, D.R.; Cinner, J.E.; Hughes, T.P.; Norström, A.V.; Nyström, M. Managing resilience to reverse phase shifts in coral reefs. Front. Ecol. Environ. 2013, 11, 541–548. [Google Scholar] [CrossRef]
- Hughes, T.P.; Anderson, K.D.; Connolly, S.R.; Heron, S.F.; Kerry, J.T.; Lough, J.M.; Baird, A.H.; Baum, J.K.; Berumen, M.L.; Bridge, T.C.; et al. Spatial and temporal patterns of mass bleaching of corals in the Anthropocene. Science 2018, 359, 80–83. [Google Scholar] [CrossRef] [PubMed]
- Cesar, H.S. Coral reefs: Their functions, threats and economic value. In Collected Essays on the Economics of Coral Reefs; Cesar, H.S.J., Ed.; CORDIO, Department of Biology and Environmental Sciences, Kalmar University: Kalmar, Sweden, 2002; pp. 14–39. [Google Scholar]
- Halpern, B.S.; Selkoe, K.A.; Micheli, F.; Kappel, C.V. Evaluating and ranking the vulnerability of global marine ecosystems to anthropogenic threats. Conserv. Biol. 2006, 21, 1301–1315. [Google Scholar] [CrossRef] [PubMed]
- van Oppen, M.J.H.; Gates, R.D.; Blackall, L.L.; Cantin, N.; Chakravarti, L.J.; Chan, W.Y.; Cormick, C.; Crean, A.; Damjanovic, K.; Epstein, H.; et al. Shifting paradigms in restoration of the world’s coral reefs. Glob. Chang. Biol. 2017, 23, 3437–3448. [Google Scholar] [CrossRef]
- Boström-Einarsson, L.; Babcock, R.C.; Bayraktarov, E.; Ceccarelli, D.; Cook, N.; Ferse, S.C.A.; Hancock, B.; Harrison, P.; Hein, M.; Shaver, E. Coral restoration—A systematic review of current methods, successes, failures and future directions. PLoS ONE 2020, 15, e0226631. [Google Scholar] [CrossRef]
- Barton, J.A.; Willis, B.L.; Hutson, K. Coral propagation: A review of techniques for ornamental trade and reef restoration. Rev. Aquac. 2017, 9, 238–256. [Google Scholar] [CrossRef]
- van Oppen, M.J.H.; Oliver, J.K.; Putnam, H.M.; Gates, R.D. Building coral reef resilience through assisted evolution. Proc. Natl. Acad. Sci. USA 2015, 112, 2307–2313. [Google Scholar] [CrossRef]
- Westoby, R.; Becken, S.; Laria, A.P. Perspectives on the human dimensions of coral restoration. Reg. Environ. Chang. 2020, 20, 109. [Google Scholar] [CrossRef]
- Viehman, T.S.; Reguero, B.G.; Lenihan, H.S.; Rosman, J.H.; Storlazzi, C.D.; Goergen, E.A.; Canals Silander, M.F.; Groves, S.H.; Holstein, D.M.; Bruckner, A.W.; et al. Coral restoration for coastal resilience: Integrating ecology, hydrodynamics, and engineering at multiple scales. Ecosphere 2023, 14, e4517. [Google Scholar] [CrossRef]
- Suggett, D.J.; van Oppen, M.J. Horizon scan of rapidly advancing coral restoration approaches for 21st century reef management. Emerg. Top. Life Sci. 2022, 14, 125–136. [Google Scholar] [CrossRef] [PubMed]
- Omori, M. Coral restoration research and technical developments: What we have learned so far. Mar. Biol. Res. 2019, 15, 377–409. [Google Scholar] [CrossRef]
- Tortolero-Langarica, J.J.A.; Rodríguez-Troncoso, A.P.; Cupul-Magaña, A.L.; Alarcón-Ortega, L.C.; Santiago-Valentín, J.D. Accelerated recovery of calcium carbonate production in coral reefs using low-tech ecological restoration. Ecol. Eng. 2019, 128, 89–97. [Google Scholar] [CrossRef]
- Rodríguez-Troncoso, A.P.; Tortolero-Langarica, J.J.A.; Padilla-Guzmán, R.; Kelly-Gutiérrez, L.D.; Cupul-Magaña, A.L. Spatio-temporal variation in the growth of coral fragments of opportunity in the Eastern Tropical Pacific: Implications for coral reef restoration. Rev. Biol. Trop. 2023, 71, e54850. [Google Scholar] [CrossRef]
- Tortolero-Langarica, J.J.A.; Rodríguez-Troncoso, A.P.; Cupul-Magaña, A.L.; Carricart-Ganivet, J.P. Calcification and growth rate recovery of the reef-building Pocillopora species in the northeast tropical Pacific following an ENSO disturbance. PeerJ 2017, 5, e3191. [Google Scholar] [CrossRef]
- Martínez-Castillo, V.; Rodríguez-Troncoso, A.P.; Cupul-Magaña, A.L. Active restoration efforts in the Central Mexican Pacific as a strategy for coral reef recovery. Rev. Biol. Trop. 2023, 71, e54795. [Google Scholar] [CrossRef]
- Pantoja, D.A.; Marinone, S.G.; Parés-Sierra, A.; Gómez-Valdivia, F. Numerical modeling of seasonal and mesoscale hydrography and circulation in the Mexican Central Pacific. Cienc. Mar. 2012, 38, 363–379. [Google Scholar] [CrossRef]
- Appendini, C.M.; Rosengaus, M.; Meza-Padilla, R.; Camacho-Magaña, V. Operational Hazard assessment of waves and storm surges from tropical cyclones in Mexico. Bull. Am. Meteorol. Soc. 2017, 98, 503–515. [Google Scholar] [CrossRef]
- Comisión Nacional de Áreas Naturales Protegidas (CONANP). Programa de Conservación y Manejo del Parque Nacional Islas Marietas, 1st ed.; Secretaria de Medio Ambiente y Recursos Naturales: Mexico City, México, 2007. [Google Scholar]
- Cupul-Magaña, A.L.; Aranda-Mena, O.S.; Medina-Rosas, P.; Vizcaíno-Ochoa, V. Comunidades coralinas de las Islas Marietas, Bahía de Banderas, Jalisco-Nayarit, México. Mexicoa 2000, 2, 15–22. [Google Scholar]
- Romero-Torres, M.; Acosta, A.; Palacio-Castro, A.M.; Treml, E.A.; Zapata, F.A.; Paz-García, D.A.; Porter, J.W. Coral reef resilience to thermal stress in the Eastern Tropical Pacific. Glob. Chang. Biol. 2020, 26, 3880–3890. [Google Scholar] [CrossRef]
- Glynn, P.W.; Gassman, N.J.; Eakin, C.M.; Cortes, J.; Smith, D.B.; Guzman, H.M. Reef coral reproduction in the eastern Pacific: Costa Rica, Panama, and Galápagos Islands (Ecuador). I. Pocilloporidae. Mar. Biol. 1991, 109, 355–368. [Google Scholar] [CrossRef]
- Carpizo-Ituarte, E.; Vizcaíno-Ochoa, V.; Chi-Barragán, G.; Tapia-Vázquez, O.; Cupul-Magaña, A.L.; Medina-Rosas, P. Evidencia de reproducción sexual en los corales hermatípicos Pocillopora damicornis, Porites panamensis y Pavona gigantea en Bahía de Banderas, Pacífico mexicano. Cienc. Mar. 2011, 37, 97–112. [Google Scholar]
- Santiago-Valentín, J.D.; Colley, S.B.; Glynn, P.W.; Cupul-Magaña, A.L.; López-Pérez, R.A.; Rodríguez-Zaragoza, F.A.; Benítez-Villalobos, F.; Bautista-Guerrero, G.; Zavala-Casas, D.A.; Rodríguez-Troncoso, A.P. Regional and species specific sexual reproductive patterns of three zooxanthellate Scleractinian corals across the Eastern Tropical Pacific. Mar. Ecol. 2018, 39, e12497. [Google Scholar] [CrossRef]
- Humason, G.L. Animal Tissue Technique; W.H. Freeman and Company: New York, NY, USA, 1979. [Google Scholar]
- Prophet, E.B.; Mills, B.; Arrington, J.B.; Sobin, L.H. Laboratory Methods in Histotechnology: Hematoxylin and Eosin; American Registry of Pathology and Armed Forces Institute of Pathology: Washington, DC, USA, 1994.
- Systat Software, Inc. SigmaPlot for Windows, Ver. 11.0; Systat Software, Inc.: San Jose CA, USA, 2008.
- Spalding, M.; Burke, L.; Wood, S.A.; Ashpole, J.; Hutchinson, J.; Zu Ermgassen, P. Mapping the global value and distribution of coral reef tourism. Mar. Policy 2017, 82, 104–113. [Google Scholar] [CrossRef]
- Bayraktaroc, E.; Banaszak, A.T.; Montoya-Maya, P.; Kleypas, J.; Arias-González, J.E.; Blanco, M.; Calle-Triviño, J.; Charuvi, N.; Cortés-Useche, C.; Galván, V.; et al. Coral reef restoration efforts in Latin American countries and territories. PLoS ONE 2020, 15, e0228477. [Google Scholar]
- Rodríguez-Troncoso, A.P.; Tortolero-Langarica, J.J.A.; Contreras Medellín-López, P.; Canizales-Flores, H.M.; Godínez-Domínguez, E.; Cupul-Magaña, A.L. Evaluación de los indicadores demográficos de Pocillopora (Scleractinia: Pocilloporidae) en sitios insulares vs. Costeros: Implicaciones para un programa de restauración regional. Rev. Biol. Trop. 2023, 71, e54790. [Google Scholar] [CrossRef]
- Parkinson, J.E.; Baker, A.C.; Baums, I.B.; Davies, S.W.; Grottoli, A.G.; Kitchen, S.A.; Matz, M.V.; Miller, M.W.; Shantz, A.A.; Kenkel, C.D. Molecular tools for coral reef restoration: Beyond biomarker discovery. Conserv. Lett. 2020, 13, e12687. [Google Scholar] [CrossRef]
- Martínez-Castillo, V.; Rodríguez-Troncoso, A.P.; Mayfield, A.B.; Rodríguez-Zaragoza, F.A.; Cupul-Magaña, A.L. Coral Recovery in the Central Mexican Pacific 20 Years after the 1997–1998 El Niño Event. Oceans 2022, 3, 48–59. [Google Scholar] [CrossRef]
- Fabregat-Malé, S.; Mena, S.; Alvarado, J.J. Nursery-reared coral outplanting success in an upwelling-influenced area in Costa Rica. Rev. Biol. Trop. 2023, 71, e54879. [Google Scholar] [CrossRef]
- Sellares-Blasco, R.I.; Villalpando, M.F.; Guendulain-García, S.D.; Croquer, A. Assisted coral reproduction in the Dominican Republic: A successful story to replicate in the Caribbean. Front. Mar. Sci. 2021, 8, 669505. [Google Scholar] [CrossRef]
- Rodríguez-Troncoso, A.P.; Carpizo-Ituarte, E.; Cupul-Magaña, A.L. Physiological response to high temperature in the Eastern Tropical Pacific coral Pocillopora verrucosa. Mar. Ecol. 2016, 37, 1168–1175. [Google Scholar] [CrossRef]
- Martínez-Castillo, V.; Rodríguez-Troncoso, A.P.; Bautista-Guerrero, E.; Cupul-Magaña, A.L. Symbiont-coral relationship in the main reef building Scleractinians of the Central Mexican Pacific. Symbiosis 2022, 86, 315–323. [Google Scholar] [CrossRef]
- LaJeunesse, T.C.; Parkinson, J.E.; Gabrielson, P.W.; Jeong, H.J.; Reimer, J.D.; Voolstra, C.R.; Santos, S.R. Systematic revision of Symbiodiniaceae highlights the antiquity and diversity of coral endosymbionts. Curr. Biol. 2018, 28, 2570–2580. [Google Scholar] [CrossRef] [PubMed]
- Harrison, P.L. Sexual reproduction of Scleractinian corals. In Coral Reefs: An Ecosystem in Transition; Dubinsky, Z., Stambler, N., Eds.; Springer: Dordrecht, The Netherlands, 2011; pp. 59–85. [Google Scholar]
- Baums, I.B.; Baker, A.C.; Davies, S.W.; Grottoli, A.G.; Kenkel, C.D.; Kitchen, S.A.; Shantz, A.A. Considerations for maximizing the adaptive potential of restored coral populations in the western Atlantic. Ecol. App. 2019, 29, e01978. [Google Scholar] [CrossRef] [PubMed]
- Whitaker, K. Genetic evidence for mixed modes of reproduction in the coral Pocillopora damicornis and its effect on population structure. Mar. Ecol. Prog. Ser. 2006, 306, 115–124. [Google Scholar] [CrossRef]
- Pinzón, J.H.; Reyes-Bonilla, H.; Baums, I.B.; LeJaunesse, T.C. Contrasting clonal structure among Pocillopora (Scleractinia) communities at two environmentally distinct sites in the Gulf of California. Coral Reefs 2012, 31, 765–777. [Google Scholar] [CrossRef]
- Pinzón, J.H.; LaJeunesse, T.C. Species delimitation of common reef corals in the genus Pocillopora using nucleotide sequence phylogenies, population genetics and symbiosis ecology. Mol. Ecol. 2011, 20, 311–325. [Google Scholar] [CrossRef]
- Santiago-Valentín, J.D.; Rodríguez-Troncoso, A.P.; Bautista-Guerrero, E.; López-Pérez, A.; Cupul-Magaña, A.L. Settlement ecology of Scleractinian corals of the Northeastern Tropical Pacific. Coral Reefs 2020, 39, 133–146. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martínez-Castillo, V.; Rodríguez-Troncoso, A.P.; Cupul-Magaña, A.L. Evidence of Sexual Reproduction in Out-Planted Coral Colonies. Oceans 2023, 4, 350-359. https://doi.org/10.3390/oceans4040024
Martínez-Castillo V, Rodríguez-Troncoso AP, Cupul-Magaña AL. Evidence of Sexual Reproduction in Out-Planted Coral Colonies. Oceans. 2023; 4(4):350-359. https://doi.org/10.3390/oceans4040024
Chicago/Turabian StyleMartínez-Castillo, Violeta, Alma Paola Rodríguez-Troncoso, and Amílcar Leví Cupul-Magaña. 2023. "Evidence of Sexual Reproduction in Out-Planted Coral Colonies" Oceans 4, no. 4: 350-359. https://doi.org/10.3390/oceans4040024
APA StyleMartínez-Castillo, V., Rodríguez-Troncoso, A. P., & Cupul-Magaña, A. L. (2023). Evidence of Sexual Reproduction in Out-Planted Coral Colonies. Oceans, 4(4), 350-359. https://doi.org/10.3390/oceans4040024