
Hurricane-Induced Oceanic Carbon Changes in the Upper Ocean



Abstract
1. Introduction
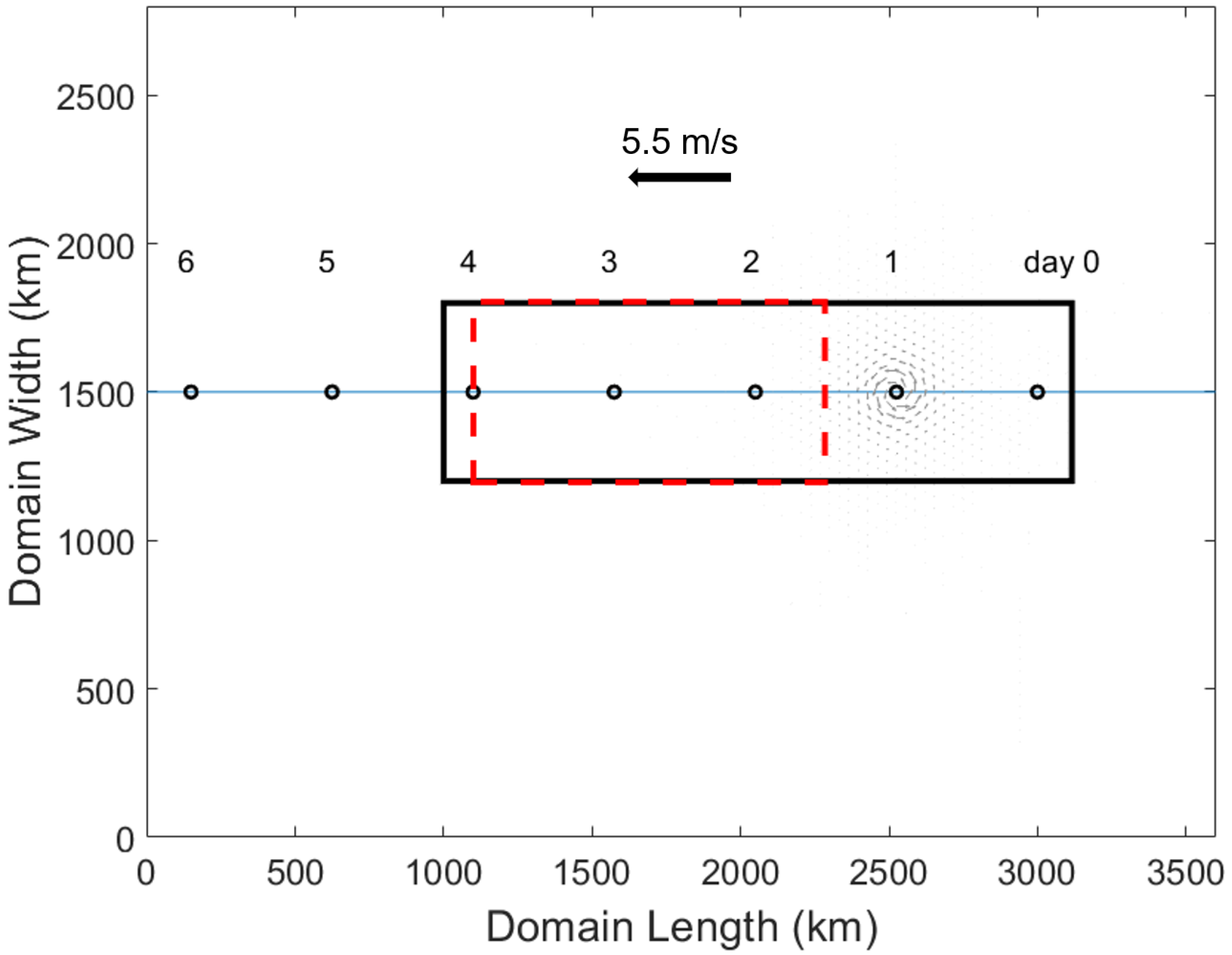
2. Materials and Methods



3. Results
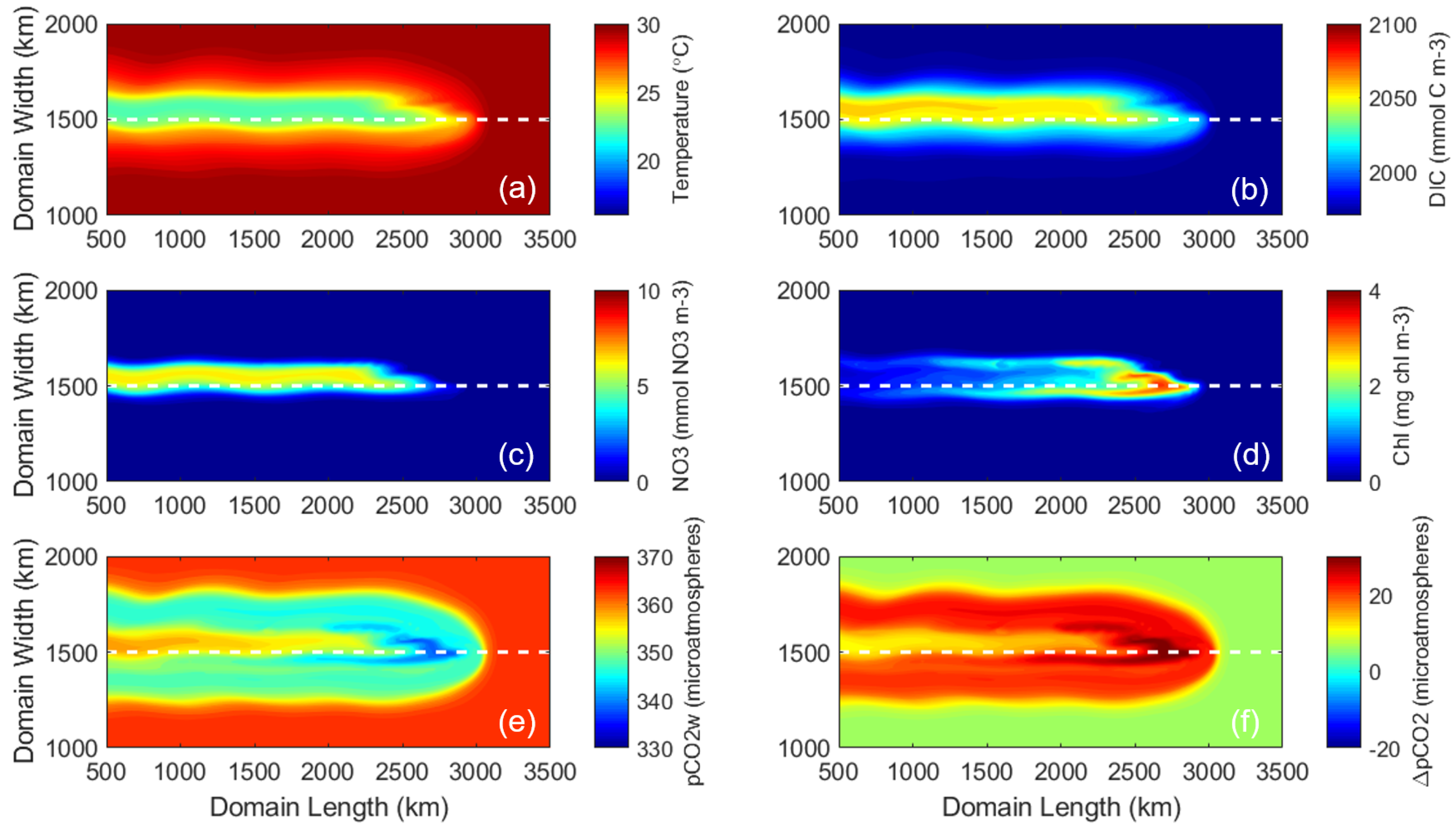
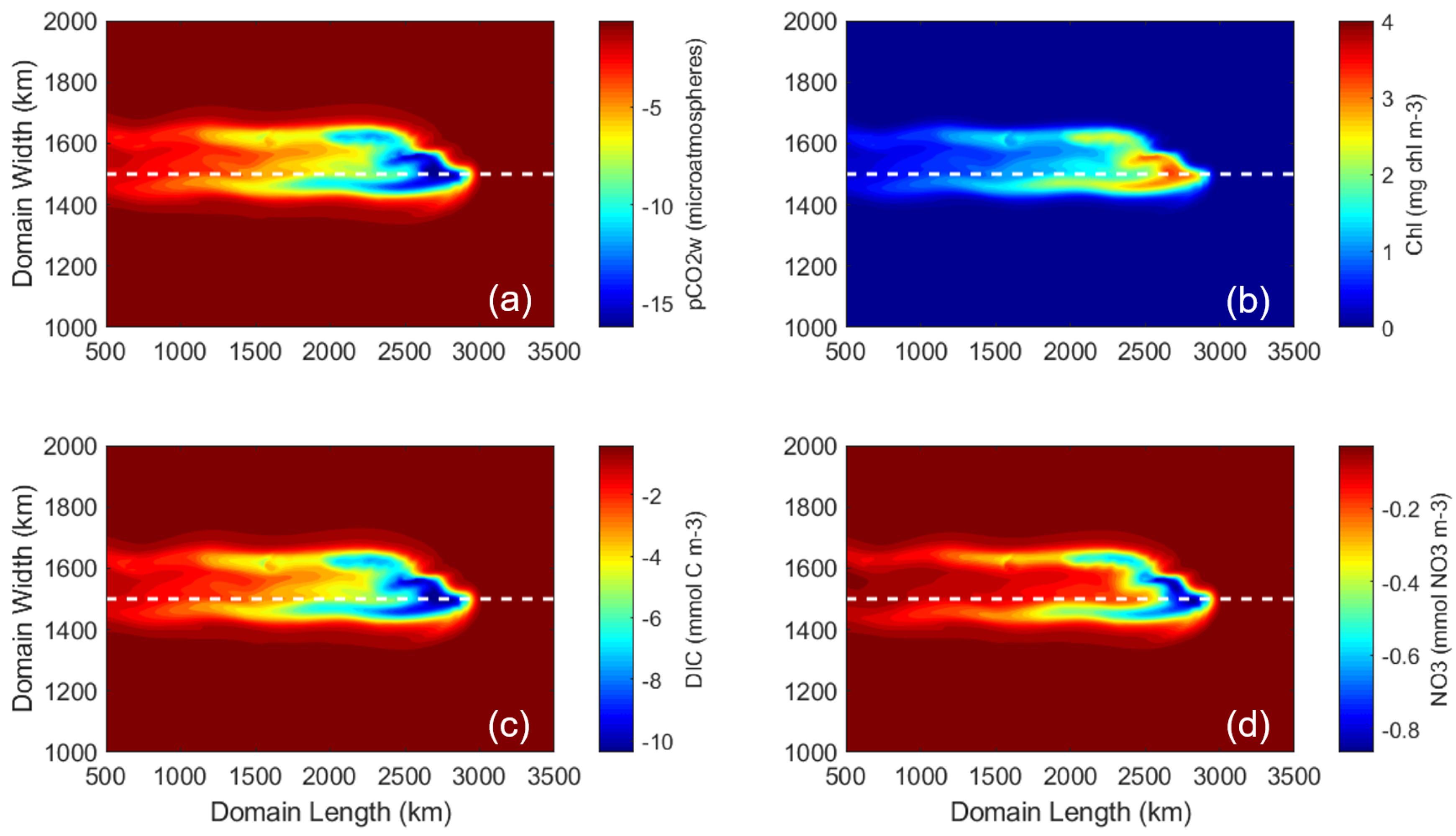
3.1. Control Run
3.2. Sensitivity Experiments
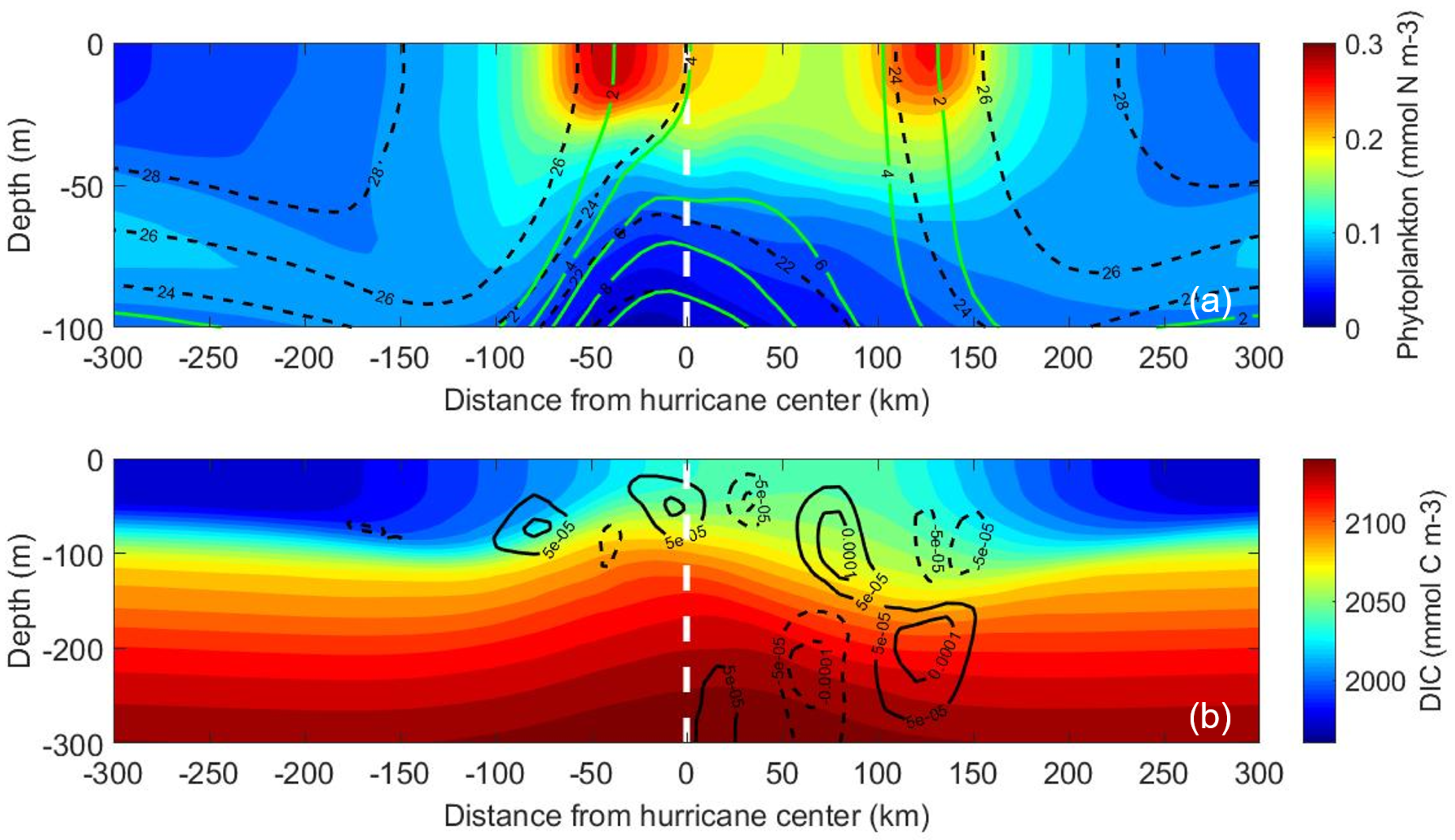
3.2.1. Effects of Biology
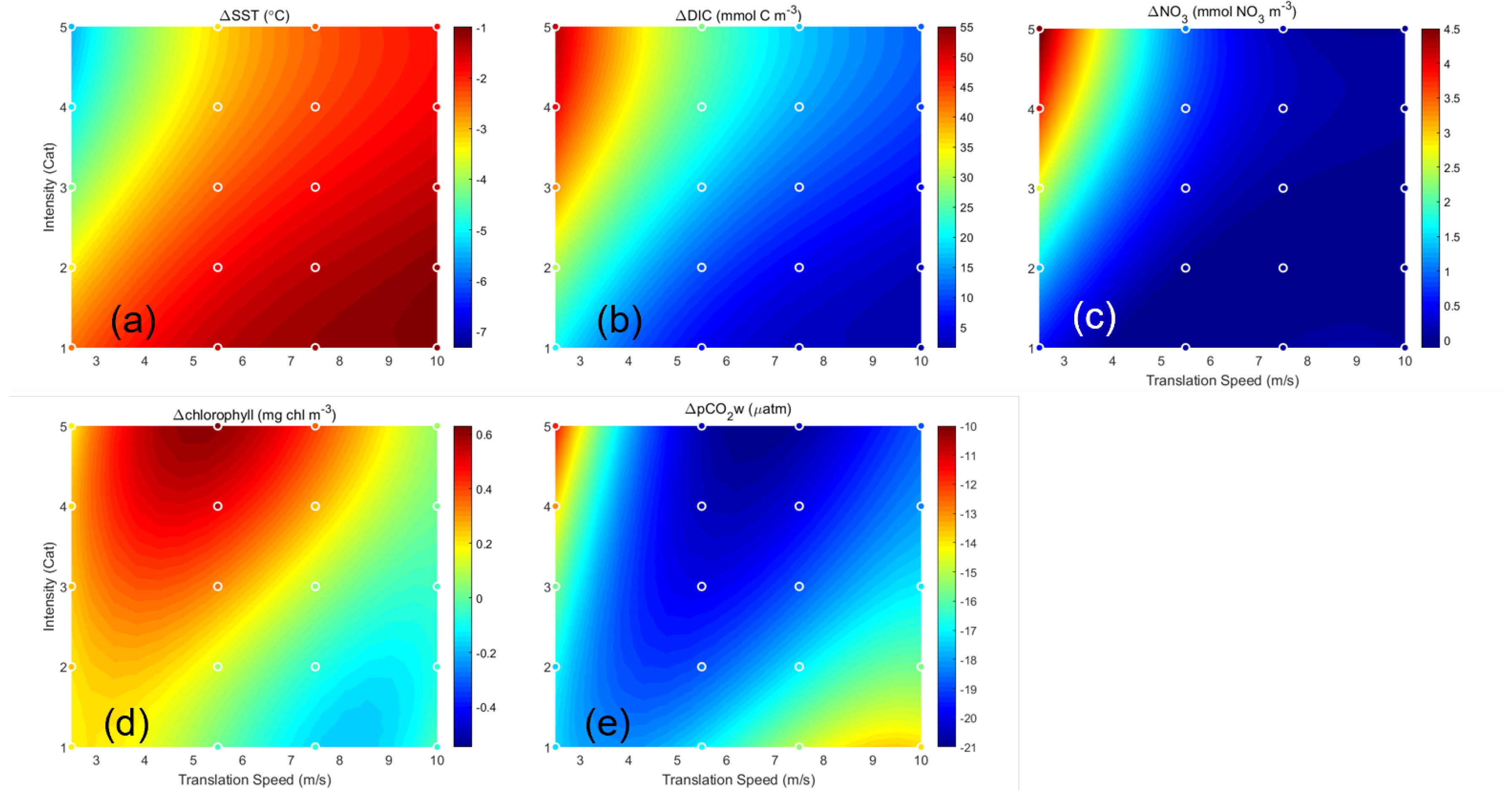
3.2.2. Effects of Translation Speed and Intensity
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| MDPI | Multidisciplinary Digital Publishing Institute |
| DOAJ | Directory of open access journals |
| pCO2w | Partial pressure of carbon dioxide in water |
| pCO2 | Partial pressure of carbon dioxide |
| pCO2a | Partial pressure of carbon dioxide in air |
| DIC | Dissolved inorganic carbon |
| SST | Sea surface temperature |
| ROMS | Regional Ocean Modeling System |
| chl | Chlorophyll |
| NO3 | Nitrate |
| pCO2 | pCO2a-pCO2w |
References
- Webster, P.; Holland, G.J.; Curry, J.A.; Chang, H.R. Changes in Tropical Cyclone Number, Duration, and Intensity in a Warming Environment. Science 2005, 309, 1844–1846. [Google Scholar] [CrossRef] [PubMed]
- Bender, M.A.; Knutson, T.R.; Tuleya, R.E.; Sirutis, J.J.; Vecchi, G.A.; Garner, S.T.; Held, I.M. Modeled Impact of Anthropogenic Warming on the Frequency of Intense Atlantic Hurricanes. Science 2010, 327, 454–458. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Murakami, H.; Knutson, T.R.; Mizuta, R.; Yoshida, K. Tropical cyclone motion in a changing climate. Sci. Adv. 2020, 6, eaaz7610. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.; Imberger, J. Variation of pCO2 in ocean surface water in response to the passage of a hurricane. J. Geophys. Res. 2010, 115, c10024. [Google Scholar] [CrossRef]
- Paerl, H.W.; Bales, J.D.; Ausley, L.W.; Buzzelli, C.P.; Crowder, L.B.; Eby, L.A.; Ramus, J.S. Ecosystem impacts of three sequential hurricanes (Dennis, Floyd, and Irene) on the United States’ largest lagoonal estuary, Pamlico Sound, NC. Proc. Natl. Acad. Sci. USA 2001, 98, 5655–5660. [Google Scholar] [CrossRef] [PubMed]
- Nyadjro, E.; Wang, Z.; Reagan, J.; Cebrian, J.; Shriver, J. Bio-Physical Changes in the Gulf of Mexico During the 2018 Hurricane Michael. IEEE Geosci. Remote Sens. Lett. 2022, 19, 1002205. [Google Scholar] [CrossRef]
- Bates, N.R.; Knap, A.H.; Michaels, A.F. Contribution of hurricanes to local and global estimates of air—Sea exchange of CO2. Nature 1998, 395, 58–61. [Google Scholar] [CrossRef]
- Nemoto, K.; Midorikawa, T.; Wada, A.; Ogawa, K.; Takatani, S.; Kimoto, H.; Ishii, M.; Inoue, H.Y. Continuous observations of atmospheric and oceanic CO2 using a moored buoy in the East China Sea: Variations during the passage of typhoons. Deep. Sea Res. Part Ii Top. Stud. Oceanogr. 2009, 56, 542–553. [Google Scholar] [CrossRef]
- Price, J.F. Upper ocean response to a hurricane. J. Phys. Oceanogr. 1981, 11, 153–175. [Google Scholar] [CrossRef]
- Lemay, J.; Thomas, H.; Craig, S.E.; Burt, W.J.; Fennel, K. Hurricane Arthur and its effect on the short-term variability of pCO2 on the Scotian Shelf, NW Atlantic. Biogeosciences 2018, 15, 2111–2123. [Google Scholar] [CrossRef]
- Yu, P.; Wang, Z.; Churchill, J.; Zheng, M.; Pan, J.; Bai, Y.; Liang, C. Effects of Typhoons on Surface Seawater pCO2 and Air-Sea CO2 Fluxes in the Northern South China Sea. J. Geophys. Res. Ocean. 2020, 125, e2020JC016258. [Google Scholar] [CrossRef]
- Babin, S.M.; Carton, J.A.; Dickey, T.D.; Wiggert, J.D. Satellite evidence of hurricane-induced phytoplankton blooms in an oceanic desert. J. Geophys. Res. Ocean. 2004, 109, c03043. [Google Scholar] [CrossRef]
- Lin, I.I. Typhoon-induced phytoplankton blooms and primary productivity increase in the western North Pacific subtropical ocean. J. Geophys. Res. Ocean. 2012, 117, c03039. [Google Scholar] [CrossRef]
- Shropshire, T.; Li, Y.; He, R.; Dare, R.A.; McBride, J.L. Storm impact of sea surface temperature and chlorophyll a in the Gulf of Mexico and Sargasso Sea based on daily cloud-free satellite data reconstructions. Geophys. Res. Lett. 2016, 43, 12199–12207. [Google Scholar] [CrossRef]
- Haakman, K.; Sayol, J.M.; van der Boog, C.; Katsman, C. Statistical Characterization of the Observed Cold Wake Induced by North Atlantic Hurricanes. Remote Sens. 2019, 11, 2368. [Google Scholar] [CrossRef]
- Eley, E.; Subrahmanyam, B.; Trott, C. Ocean-Atmosphere Interactions during Hurricanes Marco and Laura (2020). Remote Sens. 2021, 13, 1932. [Google Scholar] [CrossRef]
- Ye, H.; Sheng, J.; Tang, D.; Morozov, E.; Kalhoro, M.; Wang, S.; Xu, H. Examining the Impact of Tropical Cyclones on Air-Sea CO2 Exchanges in the Bay of Bengal Based on Satellite Data and In Situ Observations. J. Geophys. Res. Ocean. 2018, 124, 555–576. [Google Scholar] [CrossRef]
- Subrahmanyam, B.; Rao, K.H.; Srinivasa Rao, N.; Murty, V.S.N.; Sharp, R.J. Influence of a tropical cyclone on Chlorophyll-a Concentration in the Arabian Sea. Geophys. Res. Lett. 2002, 29, 22-1–22-4. [Google Scholar] [CrossRef]
- Mahadevan, A.; Tagliabue, A.; Bopp, L.; Lenton, A.; Memery, L.; Levy, M. Impact of episodic vertical fluxes on sea surface pCO2. Philos. Trans. R. Soc. Math. Phys. Eng. Sci. 2011, 369, 2009–2025. [Google Scholar] [CrossRef][Green Version]
- Chen, K.S.; Hung, C.C.; Gong, G.C.; Chou, W.C.; Chung, C.C.; Shih, Y.Y.; Wang, C.C. Enhanced POC export in the oligotrophic northwest Pacific Ocean after extreme weather events. Geophys. Res. Lett. 2013, 40, 5728–5734. [Google Scholar] [CrossRef]
- Hung, C.C.; Chung, C.C.; Gong, G.C.; Jan, S.; Tsai, Y.; Chen, K.S.; Chou, W.C.; Lee, M.A.; Chang, Y.; Chen, M.H.; et al. Nutrient supply in the Southern East China Sea after Typhoon Morakot. J. Mar. Res. 2013, 71, 133–150. [Google Scholar] [CrossRef]
- Bauer, J.E.; Cai, W.J.; Raymond, P.A.; Bianchi, T.S.; Hopkinson, C.S.; Regnier, P.A.G. The changing carbon cycle of the coastal ocean. Nature 2013, 504, 61–70. [Google Scholar] [CrossRef] [PubMed]
- McGee, L.; He, R. Mesoscale and submesoscale mechanisms behind asymmetric cooling and phytoplankton blooms induced by hurricanes: A comparison between an open ocean case and a continental shelf sea case. Ocean. Dyn. 2018, 68, 1443–1456. [Google Scholar] [CrossRef]
- Ho Ko, Y.; Park, G.H.; Kim, D.; Kim, T.W. Variations in Seawater pCO2 Associated With Vertical Mixing During Tropical Cyclone Season in the Northwestern Subtropical Pacific Ocean. Front. Mar. Sci. 2021, 8, 679314. [Google Scholar] [CrossRef]
- Ye, H.; Sheng, J.; Tang, D.; Siswanto, E.; Kalhoro, M.; Sui, Y. Storm-induced changes in pCO2 at the sea surface over the northern South China Sea during Typhoon Wutip. J. Geophys. Res. Ocean. 2017, 122, 4761–4778. [Google Scholar] [CrossRef]
- Ye, H.; Morozov, E.; Tang, D.; Wang, S.; Liu, Y.; Li, Y.; Tang, S. Variation of pCO2 concentrations induced by tropical cyclones “Wind-Pump” in the middle-latitude surface oceans: A comparative study. PLoS ONE 2020, 15, e0226189. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.W.; Anthes, R.A. Numerical simulations of the ocean’s nonlinear, baroclinic response to translating hurricanes. J. Phys. Oceanogr. 1978, 8, 468–480. [Google Scholar] [CrossRef]
- Haidvogel, D.B.; Arango, H.; Budgell, W.P.; Cornuelle, B.D.; Curchitser, E.; Di Lorenzo, E.; Fennel, K.; Geyer, W.R.; Hermann, A.J.; Lanerolle, L.; et al. Ocean forecasting in terrain-following coordinates: Formulation and skill assessment of the Regional Ocean Modeling System. J. Comput. Phys. 2008, 227, 3595–3624. [Google Scholar] [CrossRef]
- Fennel, K.; Wilkin, J.; Levin, J.; Moisan, J.; O’Reilly, J.; Haidvogel, D. Nitrogen cycling in the Middle Atlantic Bight: Results from a three-dimensional model and implications for the North Atlantic nitrogen budget. Glob. Biogeochem. Cycles 2006, 20. [Google Scholar] [CrossRef]
- Fennel, K.; Wilkin, J.; Previdi, M.; Najjar, R. Denitrification effects on air-sea CO2 flux in the coastal ocean: Simulations for the northwest North Atlantic. Geophys. Res. Lett. 2008, 35. [Google Scholar] [CrossRef]
- Fennel, K.; Hetland, R.; Feng, Y.; DiMarco, S. A coupled physical-biological model of the Northern Gulf of Mexico shelf: Model description, validation and analysis of phytoplankton variability. Biogeosciences 2011, 8, 1881–1899. [Google Scholar] [CrossRef]
- Umlauf, L.; Burchard, H. A generic length-scale equation for geophysical turbulence models. J. Mar. Res. 2003, 61, 235–265. [Google Scholar] [CrossRef]
- Warner, J.C.; Sherwood, C.R.; Arango, H.G.; Signell, R.P. Performance of four turbulence closure models implemented using a generic length scale method. Ocean. Model. 2005, 8, 81–113. [Google Scholar] [CrossRef]
- Kantha, L.H.; Clayson, C.A. An improved mixed layer model for geophysical applications. J. Geophys. Res. 1994, 99, 25235–25266. [Google Scholar] [CrossRef]
- Holland, G. An analytic model of the wind and pressure profiles in hurricanes. Mon. Weather Rev. 1980, 108, 1212–1218. [Google Scholar] [CrossRef]
- Huang, S.; Oey, L. Right-side cooling and phytoplankton bloom in the wake of a tropical cyclone. J. Geophys. Res. Ocean. 2015, 120, 5735–5748. [Google Scholar] [CrossRef]
- Sanabia, E.; Jayne, S. Ocean Observations Under Two Major Hurricanes: Evolution of the Response Across the Storm Wakes. AGU Adv. 2020, 1, e2019AV000161. [Google Scholar] [CrossRef]
- Eppley, R.W. Temperature and Phytoplankton Growth in the Sea. Fish. Bull. 1972, 70, 1063–1085. [Google Scholar]
- Kossin, J.P. A global slowdown of tropical-cyclone translation speed. Nature 2018, 558, 104–107. [Google Scholar] [CrossRef]
- Mei, W.; Xie, S.P. Intensification of landfalling typhoons over the northwest Pacific since the late 1970s. Nat. Geosci. 2016, 9, 753–757. [Google Scholar] [CrossRef]
- Gutmann, E.D.; Rasmussen, R.M.; Liu, C.; Ikeda, K.; Bruyere, C.L.; Done, J.M.; Garrè, L.; Friis-Hansen, P.; Veldore, V. Changes in Hurricanes from a 13-Yr Convection-Permitting Pseudo–Global Warming Simulation. J. Clim. 2018, 31, 3643–3657. [Google Scholar] [CrossRef]
- Zhang, W. Extreme Translation Events of Atlantic Tropical Cyclones. Atmosphere 2021, 12, 1032. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hurricane Category | Minimum Central Pressure (mB) | Maximum Wind Speed (m s−1) |
|---|---|---|
| Cat 1 | 975 | 37.5 |
| Cat 2 | 965 | 44.6 |
| Cat 3 | 950 | 54.5 |
| Cat 4 | 930 | 66.8 |
| Cat 5 | 920 | 72.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
McGee, L.; He, R. Hurricane-Induced Oceanic Carbon Changes in the Upper Ocean. Oceans 2022, 3, 114-124. https://doi.org/10.3390/oceans3020010
McGee L, He R. Hurricane-Induced Oceanic Carbon Changes in the Upper Ocean. Oceans. 2022; 3(2):114-124. https://doi.org/10.3390/oceans3020010
Chicago/Turabian StyleMcGee, Laura, and Ruoying He. 2022. "Hurricane-Induced Oceanic Carbon Changes in the Upper Ocean" Oceans 3, no. 2: 114-124. https://doi.org/10.3390/oceans3020010
APA StyleMcGee, L., & He, R. (2022). Hurricane-Induced Oceanic Carbon Changes in the Upper Ocean. Oceans, 3(2), 114-124. https://doi.org/10.3390/oceans3020010