
Cell Fractionation and the Identification of Host Proteins Involved in Plant–Virus Interactions



1
Department of Plant Pathology and Nebraska Center for Virology, University of Nebraska-Lincoln, Lincoln, NE 68583, USA
2
Department of Botany, Faculty of Science, Mansoura University, Mansoura 35516, Egypt
3
Department of Biology, Kutztown University of Pennsylvania, Kutztown, PA 19530, USA
*
Author to whom correspondence should be addressed.
Pathogens 2024, 13(1), 53; https://doi.org/10.3390/pathogens13010053
Submission received: 8 December 2023
/
Revised: 19 December 2023
/
Accepted: 22 December 2023
/
Published: 5 January 2024
(This article belongs to the Special Issue Host-Virus Interactions in Viral Infectious Diseases)
Abstract
:Plant viruses depend on host cellular factors for their replication and movement. There are cellular proteins that change their localization and/or expression and have a proviral role or antiviral activity and interact with or target viral proteins. Identification of those proteins and their roles during infection is crucial for understanding plant–virus interactions and to design antiviral resistance in crops. Important host proteins have been identified using approaches such as tag-dependent immunoprecipitation or yeast two hybridization that require cloning individual proteins or the entire virus. However, the number of possible interactions between host and viral proteins is immense. Therefore, an alternative method is needed for proteome-wide identification of host proteins involved in host–virus interactions. Here, we present cell fractionation coupled with mass spectrometry as an option to identify protein–protein interactions between viruses and their hosts. This approach involves separating subcellular organelles using differential and/or gradient centrifugation from virus-free and virus-infected cells (1) followed by comparative analysis of the proteomic profiles obtained for each subcellular organelle via mass spectrometry (2). After biological validation, prospect host proteins with proviral or antiviral roles can be subject to fundamental studies in the context of basic biology to shed light on both virus replication and cellular processes. They can also be targeted via gene editing to develop virus-resistant crops.
1. Introduction
Plant viruses are obligate intracellular parasites that depend on their host’s biological machinery to replicate and move cell-to-cell and systemically. Plant virus genomes encode a small set of proteins that hijack a variety of cellular processes and interact with host proteins and other biological molecules [1]. In response to viral infection, a subset of host proteins change their subcellular localization and/or accumulation level, and host organelles undergo some functional and structural changes [2,3]. For any virus–host combination, host proteins may not be involved or have a role in the interaction. The latter group can be divided into proteins with antiviral activity or a proviral role. Antiviral proteins actively defend against the virus while proviral proteins support the virus in critical parts of replication or spread within the host [4,5,6]. Therefore, the susceptibility or resistance of plants to viruses is influenced by changes that occur in the host at the protein level. Identification of proviral proteins can establish the foundation to engineer antiviral resistance by eliminating or mutating genes that determine susceptibility [7,8].
Several techniques have been used to identify interaction partners and track and measure protein accumulation, subcellular localization, and biological activity during viral infection, including 2D-gel electrophoresis, protein microarrays, yeast two hybridization, affinity purification, confocal microscopy, and bimolecular fluorescence complementation [9,10,11,12]. The yeast two hybrid (Y2H) approach allows the detection of soluble protein interactions within the cellular environment. However, it can only detect about 25% of interactions as it requires preparation of a library of bait and pray cloned proteins [13]. Other methods such as coimmunoprecipitation (Co-IP), isobaric tags for relative and absolute quantification (iTRAQ), phase separation, proximity labeling [14], and protein interaction reporter technology [15] have also be used for detecting protein–protein interactions in plant–virus combinations. These methods require cloning and tagging individual viral proteins alone or in the context of an infectious virus clone. While these approaches have provided fantastic results for key viral proteins of representative or model species [10,11,16,17], most plant viruses have not been cloned. Additionally, due to strict genome constrains or possible interference with protein function, it is likely that many viruses or viral proteins will not tolerate even simple tags (FLAG, myC, HA, HIS, etc.), as shown for Pea enation mosaic virus 1 (PEMV1) where the addition of 3xHA to the coat protein significantly reduced PEMV1 accumulation [18]. Here, we propose the combination of tag- and clone-independent subcellular fractionation and mass spectrometry to identify, at the proteome level, host proteins interacting with viral proteins.
2. Cellular Organelles Are Involved in Virus Infection
Viruses have evolved to take advantage of host organelles to form their replication compartments and replicate their genomes with help from cellular proteins [19]. This section highlights some examples of organelle involvement in virus replication. Upon infection with Tomato mosaic virus (ToMV), 130 KDa and 180 KDa replication proteins associate with intracellular membranes and bind to host proteins such as TOM1. This binding is required for RNA-dependent RNA polymerase (RdRp) activity and ToMV genome replication [20,21].
The host secretory pathway is used for replication by some viruses. During infection with Tobacco mosaic virus (TMV), the host endoplasmic reticulum (ER) is transformed into large irregular aggregates used to form virus replication compartments that contain viral replication proteins, viral genomic RNA, and other host factors including ER resident chaperones such as BiP, PDI, and SKP1 [22,23,24]. The ER also plays a role in viral transport through cells, and some viruses such as TMV modify the ER tubules or form motile vesicles detached from the ER that are then targeted to endosomes [25].
Chloroplasts play a pivotal role for some plant viruses. The chloroplast unusual positioning protein 1 (CHUP1) facilitates the movement of Cauliflower mosaic virus (CaMV) and its inclusion bodies between cells through interaction with virus protein P6. Accordingly, silencing CHUP1 delays viral movement [26,27]. Moreover, chloroplast membranes serve as platforms for Potexvirus replication, with viral proteins including ribonucleoprotein complex (RNP) associating with host factors such as chloroplast phosphoglycerate kinase (chlPGK) and chaperonin Hsp90 to form viral replication compartments [28,29]
The chloroplast, Golgi apparatus, and ER are involved during infection by Turnip mosaic virus (TuMV). Upon infection, these organelles amalgamate into a perinuclear globular structure within which viral replication takes place [30].
Extracellular vesicles have dual roles in viral infection. In TuMV-infected plants, extracellular vesicles carry host proteins needed for virus replication proteins (poly(A)-binding protein) and antiviral defense proteins (AGO2 and 14-3-3 protein), potentially serving both as viral movement mediators and immune signal transmitters [31].
Other organelles such as mitochondria and the nucleolus are also involved in viral infection. The mitochondria participate in electron transport chain. Infection by Cucumber mosaic virus (CMV) disrupts electron transport chains in chloroplasts and mitochondria, leading to higher electron flow to oxygen (O2) and increased accumulation of hydrogen peroxide (H2O2) [32]. The nucleolus is targeted by some viral proteins such as the 3a movement protein encoded by CMV, protein P3 of Tobacco etch virus, and the coat protein (CP) of Tomato yellow leaf curl virus, which serves as a nuclear shuttle to facilitate the transport of viral DNA into and out of the nucleus, where replication takes place [33]. During infection with Groundnut rosette virus (GRV), the long-distance movement protein (MP) associates with host fibrillarin and travels to the nucleolus. Thus, there is a connection between fibrillarin binding and nucleolar trafficking in plant–virus interactions [34,35].
Great progress has been made in understanding interactions between viral proteins and cellular components using various techniques, such as proteomics, transcriptomics, and advanced imaging technologies [10,15,16,17]. However, there are still significant gaps in our understanding of the repertoire of interactions between plant viral proteins and cellular components. The identity of host factors, their mechanistic role, and their contribution to antiviral defense or viral replication and pathogenesis require further investigation.
3. Host Proteins Participate in Virus Infection
Plant host proteins play several contrasting roles during viral infection, such as being proviral or antiviral. These roles influence the progression of the disease and the outcome of the interaction. Proviral proteins are host factors that viruses exploit for their replication [36,37], cellular movement [38], or for overcoming antiviral defenses [8]. Potyviruses selectively require different eIF4E paralogs to translate their proteins and recruit them through protein–protein interactions using potyviral VPg [39]. Thus, the translation initiation factor eIF(iso)4E is a proviral host protein. Plant susceptibility to Tobacco etch virus (TEV) and TuMV depends on the presence of eIF(iso)4E. Mutant Arabidopsis thaliana plants lacking eIF(iso)4E are resistant to TEV and TuMV [40]. Similarly, reducing the expression of eIF(iso)4E in plum plants confers resistance to Plum pox virus [41].
Host proteins with antiviral activity are involved in various defense pathways such as RNA silencing, the hypersensitive response, autophagy, RNA decay, or pathogenesis-related proteins [42,43,44]. RNA-binding proteins, such as PUMILIO proteins bind to target RNAs using specialized RNA-binding domains and can directly or indirectly contribute to the plant defense system against RNA viruses [6,45]. Components of RNA silencing such as Dicer-like (DCL) proteins, Argonaute (AGO) proteins, and RNA-dependent RNA polymerases (RDRs) have antiviral roles [46,47].
4. Changes in Accumulation and Subcellular Localization of Host Proteins
Viral proteins localize to particular subcellular organelles (Figure 1). A multitude of changes occur in host proteins during viral infection including changes in protein expression, accumulation and/or subcellular distribution (Figure 2). These alterations are part of the plant response to restrict virus infection or are orchestrated by the virus to create a favorable environment for its replication, movement, or antagonization of the host defense response. Interestingly, proteins involved in the interactions partially overlap between resistant and susceptible cultivars [12]. For instance, RDR1 is a critical component of antiviral defense and is activated by virus infection [48,49]. During infection with Turnip crinkle virus, the expression of mitochondrial matrix HSP70 and CPN60 proteins increases in infected plants as a stress response to mitochondrial damage caused by the virus [50]. In response to TMV infection, the abundance of callose synthase, an enzyme involved in the synthesis of callose, increases in resistant tobacco plants. Callose is deposited inside the plasmodesmata and blocks the cell-cell movement of the virus, contributing to antiviral defense. In contrast, in susceptible plants, TMV can circumvent this barrier by increasing the activity of the host pathogenesis-related protein PR2. This leads to an increased deposition of the enzyme β-1,3 glucanase in the plasmodesmata, β-1,3 glucanase hydrolyzes callose, thereby facilitating the movement of the virus through the plasmodesmata [51,52,53].
Another possible change in proviral factors is the expression of specific proteins only during viral infection (Figure 2A) as in the case of Kunitz peptidase inhibitor-like protein (KPILP), which is not expressed in non- infected N. benthamiana. KPILP is involved in chloroplast retrograde signaling regulation and stimulation of intercellular transport of macromolecules and is induced by stress or expressed upon TMV infection. In plants with KPILP knocked-down TMV accumulation and intercellular movement were reduced [54].
Viral infection can also induce changes in the subcellular localization of host proteins. These changes are necessary for viral replication, cell-to-cell movement of the virus, and other steps of the viral infection. A comprehensive list of host proteins that undergo subcellular changes induced by the viral infection is reviewed in [2].
5. Potyviruses as a Model for Host–Viral Protein Interactions
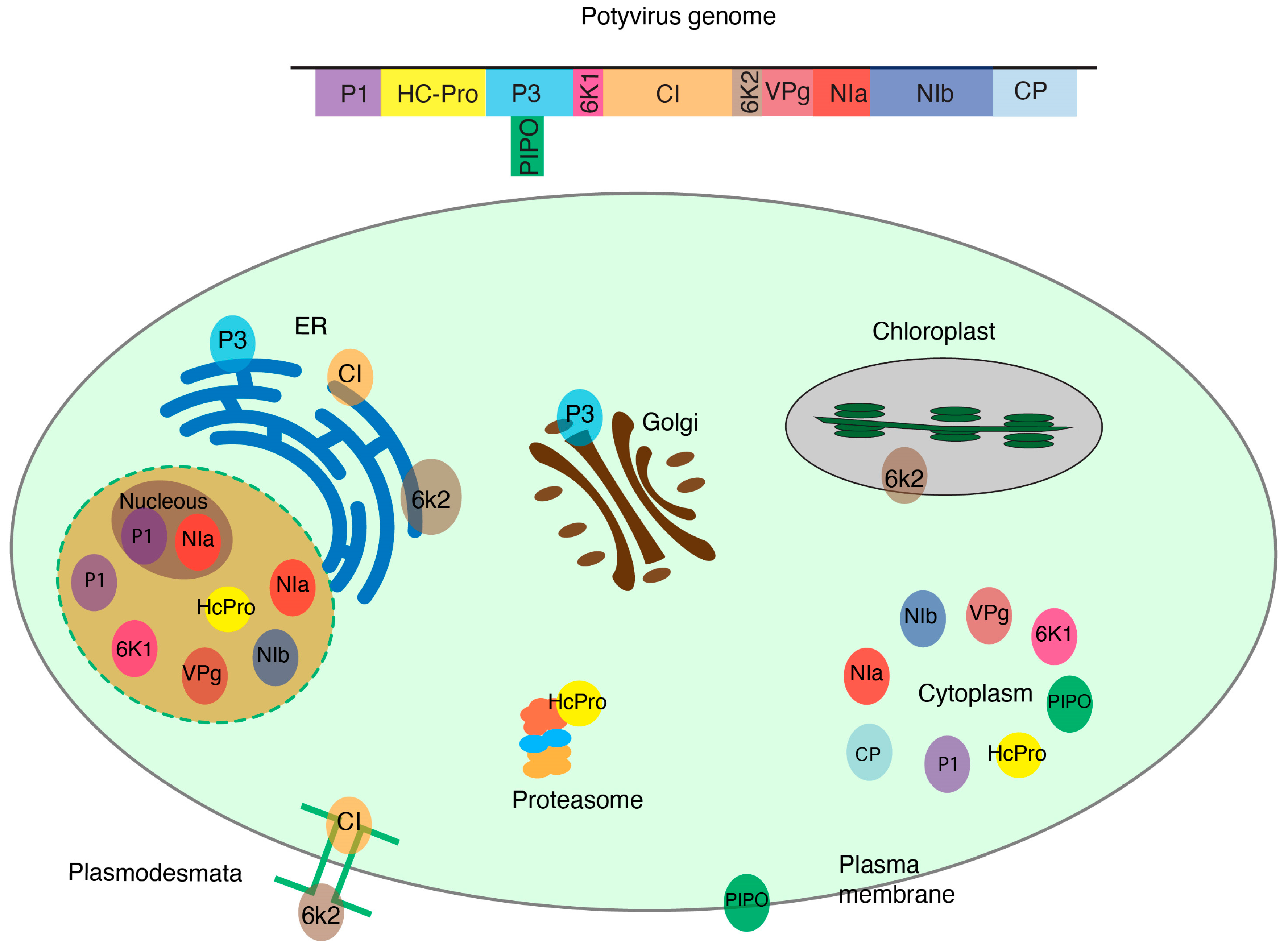
The genus Potyvirus consists of positive single-stranded RNA viruses, is one of the most diverse genera, and includes species that cause significant economic losses [55]. Potyviruses encode 11 conserved proteins (VPg, P1, HcPro, P3, P3N-PIPO, 6K1, CI, 6K2, Pro, NIb, and CP). During infection, each one of the viral proteins is translocated to cellular organelles and interacts with host proteins to facilitate replication, movement, or other aspects of viral infection (Figure 1). Some host–viral protein interactions have been identified and characterized [56,57]. Most remain unexplored.
TEV protein P1 is translocated between the nucleolus and cytoplasm and interacts with the host 80s cytoplasmic ribosomes, binding to the 60s ribosomal subunits. This interaction facilitates translation of viral proteins [58]. Potyviral VPg is covalently linked to the 5′ end of the genome and binds to the host translation initiation factors eIF4E/eIF(iso)4E which is critical for viral protein synthesis [59]. TuMV VPg interacts with and degrades suppressor of gene silencing 3 (SGS3) and RDR6, both important components of the RNA silencing pathway [60]. Potyviral P3 interacts with plant eukaryotic elongation factor 1 (eEF1A) to promote the unfolded protein response and viral pathogenesis [59]. P3N-PIPO interacts with the host plasma-membrane-localized cation binding protein PCaP1, which helps in translocating the viral protein to the plasma membrane and the plasmodesmata. This, in turn, facilitates virus movement between cells [61,62].
In Arabidopsis thaliana plants infected with TuMV, Nlb interacts with the host RNA helicase AtRH9 which is recruited to the viral replication compartments associated with chloroplasts. The interaction with AtRH9 enhances viral replication by stimulating the RNA-dependent RNA polymerase activity of Nlb [63].
The complex network of interactions between potyviral proteins and their host proteins and organelles illustrates the interplay between the virus and its host, which impacts viral replication, movement, and plant defense responses [11,16,57]. Mapping all these interactions is necessary for a thorough understanding of plant–virus interactions and for developing strategies to design antiviral resistance in crops.
Collectively, the 167 species of potyvirus infect approximately 318 host plant species, each with a genome that encodes at least 36,795 proteins [64]. Thus, the number of possible interactions between the 11 potyviral proteins and the 36,795 host proteins is massive. It would be impossible to study these interactions on an individual basis. The task becomes even more challenging when integrating the 26 plant virus families grouping 118 genera and 1516 species of viruses.
6. Cellular Fractionation to Identify Host Proteins
Interactions between plant viruses and their host proteins occur within subcellular compartments (Figure 1). Thus, identification of host proteins important in plant–virus interactions can be achieved by separating cellular compartments into fractions and proteins that co-localize in each subcellular compartment identified using mass spectrometry [15,65,66]. By comparing proteomic profiles between a virus-infected and a virus-free sample, it would be possible to identify host proteins that co-localize with viral proteins in each subcellular compartment and to determine host proteins that are lost during infection in each subcellular compartment (Figure 2).
Several studies have used cell fractionation to elucidate the interaction between viral proteins and their host counterparts (Table 1). Separation of the plasma membrane was used to identify the interaction between SMALL AUXIN UP RNA (SAUR15) and the BRASSINOSTEROID-INSENSITIVE 1 (BRI1) protein in Arabidopsis thaliana plants. The interaction between these two proteins activates the plasma membrane’s H+ ATPase, which stimulates cell growth [67]. Mitochondria-rich fractions of quinoa plants infected with a mitovirus were separated via cell fractionation to analyze the impact of viral infection on the mitochondrial proteome. The analysis revealed that the infection leads to an up-regulation of proteins that modulate stress response to drought [68]. Cell fractionation was also used to elucidate the order of enzymatic activities across the Golgi apparatus. Separation of the Golgi proteins using cell fractionation, followed by mass spectrometry, revealed the presence of differences in the sequences of transmembrane amino acids across the Golgi [69].
A proteomic comparison of a resistant and a susceptible maize cultivar inoculated with Sugarcane mosaic virus (SCMV, genus Potyvirus) showed that most of the differentially expressed proteins are predicted to localize to the chloroplast [12]. Thus, separation of the chloroplast via cell-fractionation and comparison of proteomic profiles in mock-inoculated versus potyvirus-infected plants has potential to identify host and viral proteins that are important for the interaction. Similarly, for any plant–virus combination, separating nuclear and cytoplasmic fractions or soluble and membrane fractions [70] has potential to provide novel insights into the subcellular localization of viral proteins and their possible cellular interaction partners. Furthermore, proteins that re-localize within virus-infected plants have a high probability of executing a proviral role, as described for Poly(A) binding protein 2 (PABP2) and TuMV [70]. The Groundnut rosette virus open reading frame (ORF) 3 protein inters the nucleus, interacts with and re-organizes cajal bodies, and induces their fusion with the nucleolus. Nucleolar localization of the ORF3 protein is essential for the formation of viral ribonucleoprotein particles capable of intercellular movement leading to systemic infection [35]. Tobacco rattle virus (TRV) infection induces nucleolar re-distribution of coilin via a mechanism that is dependent on TRV protein 16K interacting with Poly(ADP-ribose) polymerase 1 (PAR1), a key regulator of salicylic acid mediated defense [71]. This is critical for viruses causing important diseases that have not been extensively studied such as Maize chlorotic mottle virus, a causal agent of maize lethal necrosis disease [72].

{kind=link}
{kind=link}
Table 1.
Representative host proteins important in plant–virus interactions that were identified via cellular fractionation.
Table 1.
Representative host proteins important in plant–virus interactions that were identified via cellular fractionation.
| Host | Virus | Host Protein | Viral Protein | Technique | Ref. |
|---|---|---|---|---|---|
| Transgenic tobacco BY-2 cells | ToMV | Sar1, Sec61, and TOM1 | 130 KDa and 180 KDa replication proteins | Membrane flotation analysis and Sucrose gradient sedimentation analysis | [20] |
| Transgenic tobacco BY-2 cells | ToMV | Tm-1 | 130 K and 180 K | Differential centrifugation | [73] |
| Cucumis sativus | ToRSV | N/A | NTB | Membrane fractionation | [74] |
| Pea or lettuce plants | LMV | 20 s Proteasome | HCPro | 30% sucrose cushion and gel filtration column | [75] |
| N. benthamiana | CiLV-C | N/A | P29, P15, MP, and P24 | Bimolecular Fluorescence Complementation combined with ultracentrifugation | [76] |
| Tomato | TYLCV | HSP70 | CP | Sucrose density gradient | [77] |
| N. benthamiana | PVA | Ck2, CPIP, HSP70, and CHIP | NIb, VPg, and CP | Membrane fractionation | [78] |
| N. benthamiana | CMoV | SUMO1, SUMO2, and SCE1 | ORF4 | Cell wall fractionation | [79] |
Viruses: citrus leprosis virus C (CiLV-C), lettuce mosaic virus (LMV), potato virus A (PVA), tomato yellow leaf curl virus (TYLCV), tomato ringspot virus (ToRSV), tomato mosaic virus (ToMV), and carrot mottle virus (CMoV). Viral proteins: nucleoside triphosphate binding (NTB), nuclear inclusion protein b (NIb), viral protein genome-linked (VPg), coat protein (CP), movement protein (MP), and movement protein (ORF4). Host proteins: These proteins were identified via cellular fractionation: small GTP-binding protein (Sar1), protein transport protein (Sec61), target Of Myb1 Membrane Trafficking Protein (TOM1), small ubiquitin-like modifier (SUMO), SUMO-conjugating enzyme (SCE1), heat shock protein 70 (HSP70), Hsc70-interacting protein (CHIP), protein kinase CK2, and CP-interacting protein (CPIP).
7. The Process of Cellular Fractionation
Cell fractionation involves the disruption of the cell and tissue homogenization using techniques that range from traditional grinding with a mortar and pestle to enzymatic digestion or biochemical processes [80]. Protocols for separating each subcellular compartment (Table 2) and the buffers needed (Table 3) are summarized.
Differential centrifugation is used to enrich the target organelle and eliminate other compartments and contaminants (Figure 2). The speed of centrifugation determines which cellular compartments precipitate based on their size and density. Organelles that are larger and denser, such as non-broken cells, nuclei, or chloroplasts, precipitate at lower centrifugal forces. Mitochondria, Golgi apparatus, ER, and ribosome, on the other hand, precipitate at high speed [81]. While differential centrifugation is efficient in separating organelles, its resolving power is limited. To overcome this limitation, density gradient centrifugation is employed in later stages of the process. This technique involves creating concentration gradients using substances such as sucrose or Ficoll. When the desired fraction to be purified is loaded onto the gradient, its components reach their equilibrium density, facilitating their separation [82].
Table 2.
Organelles markers and the methods for separating plant cellular components via differential and gradient centrifugation. All centrifugation is at 4 °C unless otherwise mentioned.
Table 2.
Organelles markers and the methods for separating plant cellular components via differential and gradient centrifugation. All centrifugation is at 4 °C unless otherwise mentioned.
| Organelle | Marker Proteins | Tissue Type | Homogeniza-tion Buffer | Centrifugation Speed, Condition | Fraction Obtained | Gradient Centrifugation | Purification | Final Obtained Fraction | Ref. |
|---|---|---|---|---|---|---|---|---|---|
| Membrane fraction | See other organelles | Cucumis sativus leaves | Homogenization buffer one | 30,000× g, 30 min | Crude membrane (P30) (pellet) | 20–45% sucrose gradient centrifugate at 143,000× g for 4 h | - | Membranes separated into 13 fractions | [74] |
| Cytoplasm | UDP-glucose pyrophosphorylase (UGPase) [83] | Tomato leaves | Nuclear extraction buffer | Filtrate centrifuged at 1300× g, 10 min | Cytoplasmic fraction concentrated 10 times by ultracentrifugation | 10–50% sucrose gradient centrifugation at 104,000× g for 20 h | - | 10 fractions obtained from the gradient | [77] |
| Rice cell culture | Enzyme buffer | 100,000× g, 1 h | Remove top lipid layer, take supernatant | - | Add trichloroacetic acid to supernatant; centrifuge at 20,000× g for 5 min | Wash the pellet with cold acetone at 20,000× g for 15 min and take the pellet | [84] | ||
| Vacuole | TIPs ( and isoforms), Epsilon subunit of tonoplast H+ ATPase (V-ATPase) [85] and [86] | Arabidopsis Rosette leaves | Protoplast solution | 80× g at 20 °C, 15 min | Pellet protoplast | 10% Ficoll buffer overlayed on 4% Ficoll and vacuole buffer; centrifugation at 50,000× g, 5 min at 10 °C | - | Vacuoles found on the 4% Ficoll buffer/vacuole buffer interface | [87] |
| Chloroplast | Plastocyanin (PC) light harvesting complex b (LHC) [83,85] | N. benthamiana leaves | Enzyme mixture | 300× g, 16 min | Resuspend the pellet | 40% and 80% Percoll gradient and centrifugate at 3000× g, 25 min; collect chloroplasts at the interface of 40%/80% Percoll | - | Resuspend chloroplast in resuspension buffer | [83] |
| ER | HDEL domain [85] | Castor Bean Endosperm | Homogenization buffer two | 1000× g, 15 min | supernatant | 20%, 30%, 40%, and 60% sucrose; centrifuge at 250,000× g, 22 h at 2 °C | Resuspend ER fraction between 20% and 30% and pellet via centrifugation at 250,000× g for 45 min | Resuspend ER pellet | [88] |
| Mitochondria | Voltage-dependent, anion-selective channel protein 1-5 (VDAC1-5) [89] | Citrus pulp | Extraction buffer for mitochondria | 3000× g, 10 min then centrifuge the supernatant at 12,000× g, 15 min | Resuspend the pellet in washing buffer | 18%, 22.5%, and 35% Percoll gradient; ultracentrifugation at 50,000× g, 1 h | Mitochondrial band enriched at 22.5–35% Percoll gradient interface, then diluted with washing buffer, and centrifuged at 1500× g | Resuspend, purified mitochondria pellet in small volume of washing buffer | [90] |
| Golgi | ADP-ribosylation factor 1 (ARF1) [91] | Wheat seedling | Extraction buffer for Golgi membranes | 3000× g, 20 min | supernatant | 25–40% sucrose gradient centrifuge at 100,000× g for 16 h | Ultracentrifuge fractions (1:10) at 100,000× g for 1 h | Resuspend the membrane pellet in 50 μL dilution buffer | [92] |
| Nucleus | Histone H3 [93] | Tomato leaves | Nuclear extraction buffer | 1300× g, 10 min | Pellet | 10–50% sucrose gradient centrifuge at 104,000× g, 20 h | - | 10 Nuclei fractions | [77] |
| Proteasomes | Regulatory Particle Triple-A ATPase subunit 2 (RPT2) and Regulatory Particle Non-ATPase 10 (RPN10) [94] | Arabidopsis seedlings | Extraction buffer for proteasomes | 30,000× g, 15 min | Supernatant | Precipitation with 2% and 10% PEG 8000 then re-clarifying via centrifugation at 30,000× g, 45 min | Anion exchange chromatography column | Precipitation with 10% PEG 8000 then size elution chromatography to obtain peak fraction. | [95] |
| Plasma membrane | P-type H″-ATPase [85] | Arabidopsis seedlings | Homogenizing medium | 2770× g, 10 min, take supernatant; ultracentrifugation at 231,000× g, 35 min | Pellet | - | Multiple ultracentrifugation | Resuspend the pellet in PM-suspension medium | [96] |
| Peroxisome | Catalase [97] | Arabidopsis rosette leaves | Grinding buffer | 5000× g, 1 min | Supernatant free from chloroplast and nuclei | 15–38% (v/v) Percoll gradient; centrifuge at 13,000× g, 12 min | 36% sucrose centrifuge at 39,000× g for 30 min | Leaf peroxisome fraction located at the bottom | [98] |
| Autophagosome | Autophagy-related protein 8 (ATG8) [99] | Tobacco BY-2 cell suspension culture | Lysis buffer | 17,000× g, 5 min | Pellet | 30% Percoll; centrifuge at 50,000× g, 1 h | Place density marker beads on 30% Percoll solution and centrifuge again | Fractionate into 30 fractions | [100] |
| Ribosome | Ribosomal Protein S6 (RPS6) [101] | Arabidopsis seedlings/leaves | Ribosome extraction buffer | 10,000× g, 15 min | Supernatant | Sucrose cushion, 149,000× g, 18 h | Resuspend ribosomal pellet in Staehelin A buffer; spin at 14,000× g for 15 min | Collect the supernatant | [102] |
| Extracellular vesicles | Tetraspanin 8 [103] Syntaxin PENETRATION1 (PEN1) [104] | Arabidopsis rosettes | Vesicle isolation buffer (VIB) | 700× g, 20 min at 2 °C | Supernatant | - | Centrifuge successively at 10,000× g for 60 min, 40,000× g for 60 min, and 100,000× g for 60 min and obtain the pellet each time | Pellet resuspended in VIB | [104] |
Gel electrophoresis can be used to separate protein extracts from each fraction to confirm the purity of organelle fractions during cell fractionation. The separated proteins can then be transferred onto membranes and probed using organelle markers (Table 2) that indicate the enrichment of the target organelle (Figure 2B) or the presence of contamination from non-target organelles within the fraction. This step helps in assessing the purity of the organelle fraction obtained during the fractionation process [81]. For instance, Tonoplast Intrinsic Protein (TIPs) (α and γ isoforms) and Epsilon subunit of tonoplast H+ ATPase (V-ATPase) proteins are used as markers in Western blots to confirm the extraction of the vacuole. Plastocyanin (PC) light-harvesting complex b (LHC) is used as an organelle marker for chloroplasts [83].
Following cell fractionation, mass spectrometry (MS), LC-MS, or LC-MS/MS can be used to identify and quantify the proteins present in each fraction [105]. In MS, samples are first digested using trypsin, and the peptides are then separated for quantification. In LC-MS, the MS analyses of samples are compared, while in LC-MS/MS variant a tandem mass spectrometry (MS/MS) analysis is added to provide more detailed peptide information [81]. The LC-MS approach compares the abundance of peptides in two different runs. The area under the LC-MS profile of a specific precursor ion is measured in each run. The fold change in protein abundance is then determined by comparing the areas corresponding to the same peptide in both runs. The advantage of this method is that it does not require prior peptide identification through MS/MS analysis. Peptide abundance is quantified first, and the MS/MS analysis is performed only for those peptides that show changes in abundance [106].
Table 3.
Buffers for cellular fractionation methods.
| Buffers | Components | Reference |
|---|---|---|
| Homogenization buffer one | 50 mM Tris–HCl (pH 7.4), 15 mM MgCl2, 10 mM KCl, 20% glycerol, 0.1% β-mercaptoethanol, 5 μg/mL leupeptin, and 2 μg/mL aprotinin. | [107] |
| Nuclear extraction buffer | 10 mM MES (pH 5.2), 250 mM sorbitol, 10 mM NaCl, 5 mM NaF, 5 mM EDTA, 10 mM MgCl2, 0.024% Triton X-100, 0.1% bovine serum albumin, 1 mM fresh DTT, and Complete Protease Inhibitor Mixture. | [77] |
| Enzyme buffer | 0.4 M Mannitol, 3.6 mM MES–KOH (pH 5.7), 2.0% (w/v) cellulase Onozuka RS, 0.5% (w/v) pectolyase Y-23, and 1.0% (w/v) Driselase. | [84] |
| Protoplast solution | 1% (w/v) Cellulase Onozuka R10, 1% (w/v) Macerozyme R10, 0.4 M mannitol, 25 mM CaCl2, 5 mM mercaptoethanol, and 10 mM 2-morpholinoethanesulfonic acid (MES)-KOH (pH 5.7). | [87] |
| Vacuole buffer | 0.45 M mannitol and 5 mM sodium phosphate 2 mM EDTA (pH 7.5). Keep on ice. The 200 mM sodium phosphate stock solution (pH 7.5) can be prepared by mixing 84 mL of 200 mM Na2HPO4 and 16 mL of 200 mM NaH2PO4. | [87] |
| Enzyme mixture | 1.5% (w/v) cellulase R-10, 0.5% (w/v) macerozyme R-10, 5 mM 2-morpholinoethanesulfonic acid (MES), 0.1% (w/v) BSA, 10 mM CaCl2, and 0.4 M mannitol (pH 5.8). | [83] |
| Chloroplast resuspension buffer | 0.3 M sorbitol, 20 mM Tricine-KOH (pH 7.6), 5 mM MgCl2, 2.5 mM EDTA. | [83] |
| Homogenization buffer two | 500 mM sucrose, 10 mM KCl, 1 mM EDTA, 1 mM MgCl2 2 mM dithiothreitol (DTT), 0.1 mM phenylmethyl-sulfonyl fluoride (PMSF), and 150 mM Tricine-KOH pH 7.5. | [88] |
| Extraction buffer for mitochondria | 0.4 M sorbitol, 0.2 M MOPS-Tris (pH 7.8), 7.5 mM EDTA, 1.5% (w/v) PVP-40, 0.1% [w/v] bovine serum albumin, and 2 mM DTT. | [90] |
| Washing buffer | 0.33 M sorbitol and 50 mM MOPS-Tris (pH 7.5). | [90] |
| Extraction buffer for Golgi membranes | 50 mM HEPES–KOH (pH 6.8), 0.4 M sucrose, 1 mM dithiothreitol (DTT), 5 mM MnCl2, and 5 mM MgCl2. | [92] |
| Extraction buffer for Proteasomes | 50 mM potassium phosphate (pH 6), 2 mM MgCl2, 5% (v/v) glycerol, and 5 mM 2-mercaptoethanol supplemented with 10 mM ATP, 5% polyvinylpyrrolidone, 0.6% sodium metabisulfite, and 2 mM phenylmethylsulfonyl fluoride. 0.8% plant protease inhibitor mixture is added just before use. | [95] |
| Homogenizing medium | 0.5 M sorbitol, 50 mM MOPS–KOH (pH 7.6), 5 mM EGTA, 5 mM EDTA, 1.5% (w/v) polyvinylpyrrolidone 40 (PVP-40, molecular weight 40,000), 0.5% (w/v) defatted-BSA, 2 mM phenylmethanesulfonyl fluoride (PMSF), 4 mM salicylhydroxamic acid (SHAM), and 2.5 mM 1,4-dithiothreitol (DTT). | [96] |
| Plasma membrane (PM)-suspension medium | 10 mM MOPSKOH (pH 7.3), 1 mM EGTA, 0.3 M sucrose, 1 mM DTT. Store the stock solution without DTT at 4 °C. | [96] |
| Grinding Buffer | 170 mM Tricine-KOH (pH 7.5), 1.0 M sucrose, 2 mM EDTA, 1% (w/v) BSA, 10 mM KCl, 1 mM MgCl2, 0.5% (w/v) PVP-40, and 5 mM DTT. | [98] |
| Lysis buffer | 50 mM HEPES–KOH (pH 7.5) buffer containing 1 mM EDTA, 10 μM leupeptin, 10 μM pepstatin A, 1 mM AEBSF, and 0.4 M sorbitol. Mix 50 mL of 0.1 M HEPES–KOH (pH 7.5); 20 mL of 2 M sorbitol; 29 mL of water; and 1 mL of 0.1 M EDTA–NaOH (pH 8.0). Store at 4 °C. Take 10–20 mL of lysis buffer and add 1/100 volume of 1 mM leupeptin, 1/100 volume of 1 mM pepstatin A, and 1/100 volume of 0.1 M AEBSF immediately before use. | [100] |
| Staehelin A buffer | 20 mM Tris–HCl (pH 7.5), 5 mM MgCl2, 1 mM sodium molybdate, and 1 mM dithiothreitol. | [102] |
| Ribosome extraction buffer | 200 mM Tris–HCl (pH 7.5), 200 mM KCl, 25 mM EGTA, 36 mM MgCl2, 1 mM sodium molybdate, 1 mM dithiothreitol, 50 μg/mL cycloheximide, 50 μg/mL chloramphenicol, 80 mM β-glycerophosphate, 1% (v/v) Triton X-100, 1% (v/v) Brij 35, 1% (v/v) Tween 40, and 1% (v/v) NP40. | [102] |
| Vesicle isolation buffer VIB | 20 mM MES, 2 mM CaCl2, and 0.1 M NaCl (pH 6). | [104] |
8. Experimental Challenges of Cell Fractionation
Subcellular fractionation combined with mass spectrometry comes with challenges. One of them is that obtaining a pure organelle fraction might be difficult because overlapping can occur even when applying density gradient centrifugation [82]. Another is that some subcellular compartments that are challenging to fractionate or may be lost during the fractionation process. This could lead to the underrepresentation or omission of important protein–protein interactions [81]. Moreover, protein–protein interactions between plants and viruses can be highly dynamic, transient, or dependent on specific stages of infection or cellular conditions. Since cell fractionation captures protein composition at a specific time point, it may miss short-lived or context-dependent interactions that occur during specific stages of the infection [108]. Another challenge relates to the solubility of the viral and host proteins. Some membrane-associated proteins and other proteins can be difficult to solubilize or extract during fractionation, which can result in the loss or incomplete representation of these interactions in the fractionation and subsequent mass spectrometry analysis [109,110].
Proteins accumulates at variable quantities and cellular organelles vary in abundance. In published information (Table 1), the starting material was protoplasts, or 1 g to 500 g of leaf tissue. Thus, it can be predicted that the amount of tissue to start cell fractionation will also need to be adjusted for every plant–virus combination.
Mass spectrometry-based proteomics techniques also have sensitivity and dynamic range limitations that may impact the detection of low-abundance or weakly interacting proteins. They can also generate false positives and false negatives. False positives can arise from non-specific protein–protein interactions or contaminants introduced during sample processing. False negatives, on the other hand, may occur if the proteins of interest are present below the detection limit or if certain protein–protein interactions are not compatible with the fractionation or mass spectrometry techniques employed [111].
The functional relevance and significance of proteins that co-localize interactions require further validation and functional assays. Follow-up studies are necessary to understand the biological implications of the identified protein–protein interactions in the context of plant–virus interactions [112].
To overcome the limitations listed above, it is important to complement cell fractionation and mass spectrometry with other techniques, such as co-immunoprecipitation, fluorescence-based assays, or functional studies, to validate and further investigate the identified protein–protein interactions. Integrating multiple approaches can provide a more comprehensive understanding of the complex interactions between plants and viruses [113].
9. Conclusions
Identification and characterization of proteins that mediated host–viral interactions is critical for fundamental understanding of virus–host interactions, virus replication, cellular processes, and for developing strategies to engineer resistance to viruses in crops. Proteome-wide interaction analyses offer a promising alternative to traditional techniques, are not dependent on cloning or tagging individual proteins alone or in the context of an infectious clone, and allow for the comprehensive identification of host proteins important in host–viral interactions. To determine changes in subcellular localization and accumulation of host proteins during viral infection, we propose the following: (i) To extract proteins from a specific location in the cell in the form of purified organelle from virus-free and virus-infected plant using the methods listed in Table 1 and Table 2; (ii) Visualize proteins in the obtained fractions using sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and probe with the organelle marker antibody for quality control (Table 2); (iii) Identify and quantify proteins in each fraction using mass spectrometry; (iv) compare the changes in identity, abundance, and localization of proteins between the same fractions of virus-free and virus-infected samples.
Insights gained from studying viral–plant protein interactions have far-reaching implications for plant virology, agriculture, and our understanding of fundamental cellular processes. By unraveling the roles of host proteins in viral infection, we can potentially identify new targets for antiviral strategies, develop novel approaches to engineer resistant crops, and improve our understanding of the mechanisms governing cellular responses to viral infection.
Author Contributions
A.E.G. and H.G.-R. conceived the paper. A.E.G., K.E.M., E.P. and H.G.-R. wrote the paper. All authors have read and agreed to the published version of the manuscript.
Funding
Amany E. Gomaa was funded by the Ministry of Higher Education of the Arab Republic of Egypt for her Ph.D. scholarship (GM 1147) at the University of Lincoln Nebraska, USA. This research was supported by the NIH grant R01GM120 to Hernan Garcia-Ruiz and by the Nebraska Agricultural Experiment Station with funding from the Hatch Act (Accession Number 1007272) through the USDA National Institute of Food and Agriculture. Open access costs were provided by the same grant.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest. The founding sponsors had no role in the design of the study, the collection and analyses or interpretation of data, in the writing of the manuscript, or in the decision to publish the results.
References
- Hong, J.-S.; Ju, H.-J. The plant cellular systems for plant virus movement. Plant Pathol. J. 2017, 33, 213. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Peña, R.; Mounadi, K.E.; Garcia-Ruiz, H. Changes in subcellular localization of host proteins induced by plant viruses. Viruses 2021, 13, 677. [Google Scholar] [CrossRef] [PubMed]
- Wei, T.; Huang, T.-S.; McNeil, J.; Laliberté, J.-F.; Hong, J.; Nelson, R.S.; Wang, A. Sequential recruitment of the endoplasmic reticulum and chloroplasts for plant potyvirus replication. J. Virol. 2010, 84, 799–809. [Google Scholar] [CrossRef] [PubMed]
- Hyodo, K.; Okuno, T. Hijacking of host cellular components as proviral factors by plant-infecting viruses. In Advances in Virus Research; Elsevier: Amsterdam, The Netherlands, 2020; Volume 107, pp. 37–86. [Google Scholar]
- Das, P.P.; Lin, Q.; Wong, S.-M. Comparative proteomics of Tobacco mosaic virus-infected Nicotiana tabacum plants identified major host proteins involved in photosystems and plant defence. J. Proteom. 2019, 194, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Musidlak, O.; Nawrot, R.; Goździcka-Józefiak, A. Which plant proteins are involved in antiviral defense? Review on in vivo and in vitro activities of selected plant proteins against viruses. Int. J. Mol. Sci. 2017, 18, 2300. [Google Scholar] [CrossRef] [PubMed]
- Akhter, M.S.; Nakahara, K.S.; Masuta, C. Resistance induction based on the understanding of molecular interactions between plant viruses and host plants. Virol. J. 2021, 18, 176. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Ruiz, H. Susceptibility genes to plant viruses. Viruses 2018, 10, 484. [Google Scholar] [CrossRef]
- Wang, L.; Tan, H.; Medina-Puche, L.; Wu, M.; Garnelo Gomez, B.; Gao, M.; Shi, C.; Jimenez-Gongora, T.; Fan, P.; Ding, X. Combinatorial interactions between viral proteins expand the potential functional landscape of the tomato yellow leaf curl virus proteome. PLoS Pathog. 2022, 18, e1010909. [Google Scholar] [CrossRef]
- Ivanov, K.I.; Eskelin, K.; Basic, M.; De, S.; Lohmus, A.; Varjosalo, M.; Makinen, K. Molecular insights into the function of the viral RNA silencing suppressor HCPro. Plant J. 2016, 85, 30–45. [Google Scholar] [CrossRef]
- Dufresne, P.J.; Thivierge, K.; Cotton, S.; Beauchemin, C.; Ide, C.; Ubalijoro, E.; Laliberte, J.F.; Fortin, M.G. Heat shock 70 protein interaction with Turnip mosaic virus RNA-dependent RNA polymerase within virus-induced membrane vesicles. Virology 2008, 374, 217–227. [Google Scholar] [CrossRef]
- Wu, L.; Han, Z.; Wang, S.; Wang, X.; Sun, A.; Zu, X.; Chen, Y. Comparative proteomic analysis of the plant-virus interaction in resistant and susceptible ecotypes of maize infected with sugarcane mosaic virus. J. Proteom. 2013, 89, 124–140. [Google Scholar] [CrossRef] [PubMed]
- Mendez-Rios, J.; Uetz, P. Global approaches to study protein–protein interactions among viruses and hosts. Future Microbiol. 2010, 5, 289–301. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Song, G.; Lal, N.K.; Nagalakshmi, U.; Li, Y.; Zheng, W.; Huang, P.J.; Branon, T.C.; Ting, A.Y.; Walley, J.W.; et al. TurboID-based proximity labeling reveals that UBR7 is a regulator of N NLR immune receptor-mediated immunity. Nat. Commun. 2019, 10, 3252. [Google Scholar] [CrossRef] [PubMed]
- DeBlasio, S.L.; Chavez, J.D.; Alexander, M.M.; Ramsey, J.; Eng, J.K.; Mahoney, J.; Gray, S.M.; Bruce, J.E.; Cilia, M. Visualization of Host-Polerovirus Interaction Topologies Using Protein Interaction Reporter Technology. J. Virol. 2016, 90, 1973–1987. [Google Scholar] [CrossRef] [PubMed]
- Martinez, F.; Rodrigo, G.; Aragones, V.; Ruiz, M.; Lodewijk, I.; Fernandez, U.; Elena, S.F.; Daros, J.A. Interaction network of tobacco etch potyvirus NIa protein with the host proteome during infection. BMC Genom. 2016, 17, 87. [Google Scholar] [CrossRef] [PubMed]
- DeBlasio, S.L.; Johnson, R.; Mahoney, J.; Karasev, A.; Gray, S.M.; MacCoss, M.J.; Cilia, M. Insights into the polerovirus-plant interactome revealed by coimmunoprecipitation and mass spectrometry. Mol. Plant Microbe Interact. 2015, 28, 467–481. [Google Scholar] [CrossRef]
- Chkuaseli, T.; White, K.A. Complex and simple translational readthrough signals in pea enation mosaic virus 1 and potato leafroll virus, respectively. PLoS Pathog. 2022, 18, e1010888. [Google Scholar] [CrossRef] [PubMed]
- den Boon, J.A.; Ahlquist, P. Organelle-like membrane compartmentalization of positive-strand RNA virus replication factories. Annu. Rev. Microbiol. 2010, 64, 241–256. [Google Scholar] [CrossRef]
- Nishikiori, M.; Dohi, K.; Mori, M.; Meshi, T.; Naito, S.; Ishikawa, M. Membrane-bound tomato mosaic virus replication proteins participate in RNA synthesis and are associated with host proteins in a pattern distinct from those that are not membrane bound. J. Virol. 2006, 80, 8459–8468. [Google Scholar] [CrossRef]
- Nishikiori, M.; Mori, M.; Dohi, K.; Okamura, H.; Katoh, E.; Naito, S.; Meshi, T.; Ishikawa, M. A host small GTP-binding protein ARL8 plays crucial roles in tobamovirus RNA replication. PLoS Pathog. 2011, 7, e1002409. [Google Scholar] [CrossRef]
- Heinlein, M.; Padgett, H.S.; Gens, J.S.; Pickard, B.G.; Casper, S.J.; Epel, B.L.; Beachy, R.N. Changing patterns of localization of the tobacco mosaic virus movement protein and replicase to the endoplasmic reticulum and microtubules during infection. Plant Cell 1998, 10, 1107–1120. [Google Scholar] [CrossRef] [PubMed]
- Reichel, C.; Beachy, R.N. Tobacco mosaic virus infection induces severe morphological changes of the endoplasmic reticulum. Proc. Natl. Acad. Sci. USA 1998, 95, 11169–11174. [Google Scholar] [CrossRef] [PubMed]
- Verchot, J. Wrapping membranes around plant virus infection. Curr. Opin. Virol. 2011, 1, 388–395. [Google Scholar] [CrossRef] [PubMed]
- Solovyev, A.G.; Atabekova, A.K.; Lezzhov, A.A.; Solovieva, A.D.; Chergintsev, D.A.; Morozov, S.Y. Distinct mechanisms of endomembrane reorganization determine dissimilar transport pathways in plant RNA viruses. Plants 2022, 11, 2403. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, D.; Chakraborty, S. Chloroplast: The Trojan horse in plant–virus interaction. Mol. Plant Pathol. 2018, 19, 504–518. [Google Scholar] [CrossRef]
- Angel, C.A.; Lutz, L.; Yang, X.; Rodriguez, A.; Adair, A.; Zhang, Y.; Leisner, S.M.; Nelson, R.S.; Schoelz, J.E. The P6 protein of Cauliflower mosaic virus interacts with CHUP1, a plant protein which moves chloroplasts on actin microfilaments. Virology 2013, 443, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Budziszewska, M.; Obrępalska-Stęplowska, A. The role of the chloroplast in the replication of positive-sense single-stranded plant RNA viruses. Front. Plant Sci. 2018, 9, 1776. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.-F.; Huang, Y.-P.; Chen, L.-H.; Hsu, Y.-H.; Tsai, C.-H. Chloroplast phosphoglycerate kinase is involved in the targeting of Bamboo mosaic virus to chloroplasts in Nicotiana benthamiana plants. Plant Physiol. 2013, 163, 1598–1608. [Google Scholar] [CrossRef]
- Grangeon, R.; Agbeci, M.; Chen, J.; Grondin, G.; Zheng, H.; Laliberté, J.-F. Impact on the endoplasmic reticulum and Golgi apparatus of turnip mosaic virus infection. J. Virol. 2012, 86, 9255–9265. [Google Scholar] [CrossRef]
- Zhou, Q.; Ma, K.; Hu, H.; Xing, X.; Huang, X.; Gao, H. Extracellular vesicles: Their functions in plant–pathogen interactions. Mol. Plant Pathol. 2022, 23, 760–771. [Google Scholar] [CrossRef]
- Song, X.S.; Wang, Y.J.; Mao, W.H.; Shi, K.; Zhou, Y.H.; Nogués, S.; Yu, J.Q. Effects of cucumber mosaic virus infection on electron transport and antioxidant system in chloroplasts and mitochondria of cucumber and tomato leaves. Physiol. Plant. 2009, 135, 246–257. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Ryabov, E.; Brown, J.; Taliansky, M. Involvement of the nucleolus in plant virus systemic infection. Biochem. Soc. Trans. 2004, 32, 557–560. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; MacFarlane, S.; Kalinina, N.O.; Rakitina, D.V.; Ryabov, E.V.; Gillespie, T.; Haupt, S.; Brown, J.W.; Taliansky, M. Interaction of a plant virus-encoded protein with the major nucleolar protein fibrillarin is required for systemic virus infection. Proc. Natl. Acad. Sci. USA 2007, 104, 11115–11120. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Ryabov, E.V.; Kalinina, N.O.; Rakitina, D.V.; Gillespie, T.; MacFarlane, S.; Haupt, S.; Brown, J.W.; Taliansky, M. Cajal bodies and the nucleolus are required for a plant virus systemic infection. EMBO J. 2007, 26, 2169–2179. [Google Scholar] [CrossRef]
- Hyodo, K.; Okuno, T. Host factors used by positive-strand RNA plant viruses for genome replication. J. Gen. Plant Pathol. 2014, 80, 123–135. [Google Scholar] [CrossRef]
- Nagy, P.D.; Pogany, J. The dependence of viral RNA replication on co-opted host factors. Nat. Rev. Microbiol. 2012, 10, 137–149. [Google Scholar] [CrossRef]
- Hipper, C.; Brault, V.; Ziegler-Graff, V.; Revers, F. Viral and cellular factors involved in phloem transport of plant viruses. Front. Plant Sci. 2013, 4, 154. [Google Scholar] [CrossRef]
- Gallois, J.L.; Charron, C.; Sanchez, F.; Pagny, G.; Houvenaghel, M.C.; Moretti, A.; Ponz, F.; Revers, F.; Caranta, C.; German-Retana, S. Single amino acid changes in the turnip mosaic virus viral genome-linked protein (VPg) confer virulence towards Arabidopsis thaliana mutants knocked out for eukaryotic initiation factors eIF(iso)4E and eIF(iso)4G. J. Gen. Virol. 2010, 91, 288–293. [Google Scholar] [CrossRef]
- Lellis, A.D.; Kasschau, K.D.; Whitham, S.A.; Carrington, J.C. Loss-of-susceptibility mutants of Arabidopsis thaliana reveal an essential role for eIF (iso) 4E during potyvirus infection. Curr. Biol. 2002, 12, 1046–1051. [Google Scholar] [CrossRef]
- Wang, X.; Kohalmi, S.E.; Svircev, A.; Wang, A.; Sanfaçon, H.; Tian, L. Silencing of the host factor eIF (iso) 4E gene confers plum pox virus resistance in plum. PLoS ONE 2013, 8, e50627. [Google Scholar] [CrossRef]
- Li, F.; Wang, A. RNA-targeted antiviral immunity: More than just RNA silencing. Trends Microbiol. 2019, 27, 792–805. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Ruiz, H. Host factors against plant viruses. Mol. Plant Pathol. 2019, 20, 1588–1601. [Google Scholar] [CrossRef] [PubMed]
- Kudchodkar, S.B.; Levine, B. Viruses and autophagy. Rev. Med. Virol. 2009, 19, 359–378. [Google Scholar] [CrossRef] [PubMed]
- Huh, S.U.; Kim, M.J.; Paek, K.H. Arabidopsis Pumilio protein APUM5 suppresses Cucumber mosaic virus infection via direct binding of viral RNAs. Proc. Natl. Acad. Sci. USA 2013, 110, 779–784. [Google Scholar] [CrossRef] [PubMed]
- Kwon, J.; Kasai, A.; Maoka, T.; Masuta, C.; Sano, T.; Nakahara, K.S. RNA silencing-related genes contribute to tolerance of infection with potato virus X and Y in a susceptible tomato plant. Virol. J. 2020, 17, 149. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Ruiz, H.; Carbonell, A.; Hoyer, J.S.; Fahlgren, N.; Gilbert, K.B.; Takeda, A.; Giampetruzzi, A.; Garcia Ruiz, M.T.; McGinn, M.G.; Lowery, N.; et al. Roles and programming of Arabidopsis ARGONAUTE proteins during Turnip mosaic virus infection. PLoS Pathog. 2015, 11, e1004755. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Ruiz, H.; Takeda, A.; Chapman, E.J.; Sullivan, C.M.; Fahlgren, N.; Brempelis, K.J.; Carrington, J.C. Arabidopsis RNA-dependent RNA polymerases and dicer-like proteins in antiviral defense and small interfering RNA biogenesis during Turnip Mosaic Virus infection. Plant Cell 2010, 22, 481–496. [Google Scholar] [CrossRef]
- Xie, Z.; Fan, B.; Chen, C.; Chen, Z. An important role of an inducible RNA-dependent RNA polymerase in plant antiviral defense. Proc. Natl. Acad. Sci. USA 2001, 98, 6516–6521. [Google Scholar] [CrossRef]
- Blake, J.A.; Lee, K.W.; Morris, T.J.; Elthon, T.E. Effects of turnip crinkle virus infection on the structure and function of mitochondria and expression of stress proteins in turnips. Physiol. Plant. 2007, 129, 698–706. [Google Scholar] [CrossRef]
- Kozieł, E.; Otulak-Kozieł, K.; Bujarski, J.J. Plant cell wall as a key player during resistant and susceptible plant-virus interactions. Front. Microbiol. 2021, 12, 656809. [Google Scholar] [CrossRef]
- Bucher, G.L.; Tarina, C.; Heinlein, M.; Di Serio, F.; Meins Jr, F.; Iglesias, V.A. Local expression of enzymatically active class I β-1, 3-glucanase enhances symptoms of TMV infection in tobacco. Plant J. 2001, 28, 361–369. [Google Scholar] [CrossRef] [PubMed]
- Iglesias, V.A.; Meins Jr, F. Movement of plant viruses is delayed in a β-1, 3-glucanase-deficient mutant showing a reduced plasmodesmatal size exclusion limit and enhanced callose deposition. Plant J. 2000, 21, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Ershova, N.; Kamarova, K.; Sheshukova, E.; Antimonova, A.; Komarova, T. A novel cellular factor of Nicotiana benthamiana susceptibility to tobamovirus infection. Front. Plant Sci. 2023, 14, 1224958. [Google Scholar] [CrossRef] [PubMed]
- Adams, M.; Zerbini, F.; French, R.; Rabenstein, F.; Stenger, D.; Valkonen, J. Family potyviridae. In Virus Taxonomy, Ninth Report of the International Committee on Taxonomy of Viruses; King, A.M.Q., Adams, M.J., Carstens, E.B., Lefkowitz, E.J., Eds.; Elsevier: Amsterdam, The Netherlands, 2011; pp. 1069–1089. [Google Scholar]
- Ivanov, K.I.; Eskelin, K.; Lohmus, A.; Makinen, K. Molecular and cellular mechanisms underlying potyvirus infection. J. Gen. Virol. 2014, 95, 1415–1429. [Google Scholar] [CrossRef]
- Pollari, M.; De, S.; Wang, A.; Makinen, K. The potyviral silencing suppressor HCPro recruits and employs host ARGONAUTE1 in pro-viral functions. PLoS Pathog. 2020, 16, e1008965. [Google Scholar] [CrossRef]
- Martínez, F.; Daròs, J.-A. Tobacco etch virus protein P1 traffics to the nucleolus and associates with the host 60S ribosomal subunits during infection. J. Virol. 2014, 88, 10725–10737. [Google Scholar] [CrossRef] [PubMed]
- Saha, S.; Mäkinen, K. Insights into the functions of eIF4E-binding motif of VPg in potato virus A infection. Viruses 2020, 12, 197. [Google Scholar] [CrossRef]
- Cheng, X.; Wang, A. The potyvirus silencing suppressor protein VPg mediates degradation of SGS3 via ubiquitination and autophagy pathways. J. Virol. 2017, 91, e01478-01416. [Google Scholar] [CrossRef]
- Cheng, G.; Dong, M.; Xu, Q.; Peng, L.; Yang, Z.; Wei, T.; Xu, J. Dissecting the molecular mechanism of the subcellular localization and cell-to-cell movement of the sugarcane mosaic virus P3N-PIPO. Sci. Rep. 2017, 7, 9868. [Google Scholar] [CrossRef]
- Chai, M.; Wu, X.; Liu, J.; Fang, Y.; Luan, Y.; Cui, X.; Zhou, X.; Wang, A.; Cheng, X. P3N-PIPO interacts with P3 via the shared N-terminal domain to recruit viral replication vesicles for cell-to-cell movement. J. Virol. 2020, 94, e01898-01819. [Google Scholar] [CrossRef]
- Shen, W.; Shi, Y.; Dai, Z.; Wang, A. The RNA-dependent RNA polymerase NIb of potyviruses plays multifunctional, contrasting roles during viral infection. Viruses 2020, 12, 77. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Sánchez, O.; Pérez-Rodríguez, P.; Delaye, L.; Tiessen, A. Plant proteins are smaller because they are encoded by fewer exons than animal proteins. Genom. Proteom. Bioinform. 2016, 14, 357–370. [Google Scholar] [CrossRef] [PubMed]
- Chan, Z.; Qin, G.; Xu, X.; Li, B.; Tian, S. Proteome approach to characterize proteins induced by antagonist yeast and salicylic acid in peach fruit. J. Proteome Res. 2007, 6, 1677–1688. [Google Scholar] [CrossRef]
- Takac, T.; Vadovic, P.; Pechan, T.; Luptovciak, I.; Samajova, O.; Samaj, J. Comparative proteomic study of Arabidopsis mutants mpk4 and mpk6. Sci. Rep. 2016, 6, 28306. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Liu, C.; Hepworth, S.R.; Ma, C.; Li, H.; Li, J.; Wang, S.-M.; Yin, H. SAUR15 interaction with BRI1 activates plasma membrane H+-ATPase to promote organ development of Arabidopsis. Plant Physiol. 2022, 189, 2454–2466. [Google Scholar] [CrossRef] [PubMed]
- Di Silvestre, D.; Passignani, G.; Rossi, R.; Ciuffo, M.; Turina, M.; Vigani, G.; Mauri, P.L. Presence of a Mitovirus Is Associated with Alteration of the Mitochondrial Proteome, as Revealed by Protein–Protein Interaction (PPI) and Co-Expression Network Models in Chenopodium quinoa Plants. Biology 2022, 11, 95. [Google Scholar] [CrossRef] [PubMed]
- Parsons, H.T.; Stevens, T.J.; McFarlane, H.E.; Vidal-Melgosa, S.; Griss, J.; Lawrence, N.; Butler, R.; Sousa, M.M.; Salemi, M.; Willats, W.G. Separating Golgi proteins from cis to trans reveals underlying properties of cisternal localization. Plant Cell 2019, 31, 2010–2034. [Google Scholar] [CrossRef] [PubMed]
- Beauchemin, C.; Laliberte, J.F. The poly(A) binding protein is internalized in virus-induced vesicles or redistributed to the nucleolus during turnip mosaic virus infection. J. Virol. 2007, 81, 10905–10913. [Google Scholar] [CrossRef]
- Spechenkova, N.; Samarskaya, V.O.; Kalinina, N.O.; Zavriev, S.K.; MacFarlane, S.; Love, A.J.; Taliansky, M. Plant Poly(ADP-Ribose) Polymerase 1 Is a Potential Mediator of Cross-Talk between the Cajal Body Protein Coilin and Salicylic Acid-Mediated Antiviral Defence. Viruses 2023, 15, 1282. [Google Scholar] [CrossRef]
- Wamaitha, M.J.; Nigam, D.; Maina, S.; Stomeo, F.; Wangai, A.; Njuguna, J.N.; Holton, T.A.; Wanjala, B.W.; Wamalwa, M.; Lucas, T.; et al. Metagenomic analysis of viruses associated with maize lethal necrosis in Kenya. Virol. J. 2018, 15, 90. [Google Scholar] [CrossRef]
- Ishibashi, K.; Ishikawa, M. The resistance protein Tm-1 inhibits formation of a Tomato mosaic virus replication protein-host membrane protein complex. J. Virol. 2013, 87, 7933–7939. [Google Scholar] [CrossRef] [PubMed]
- Han, S.; Sanfacon, H. Tomato ringspot virus proteins containing the nucleoside triphosphate binding domain are transmembrane proteins that associate with the endoplasmic reticulum and cofractionate with replication complexes. J. Virol. 2003, 77, 523–534. [Google Scholar] [CrossRef] [PubMed]
- Ballut, L.; Drucker, M.; Pugniere, M.; Cambon, F.; Blanc, S.; Roquet, F.; Candresse, T.; Schmid, H.P.; Nicolas, P.; Gall, O.L.; et al. HcPro, a multifunctional protein encoded by a plant RNA virus, targets the 20S proteasome and affects its enzymic activities. J. Gen. Virol. 2005, 86, 2595–2603. [Google Scholar] [CrossRef] [PubMed]
- Leastro, M.O.; Kitajima, E.W.; Silva, M.S.; Resende, R.O.; Freitas-Astúa, J. Dissecting the subcellular localization, intracellular trafficking, interactions, membrane association, and topology of citrus leprosis virus C proteins. Front. Plant Sci. 2018, 9, 1299. [Google Scholar] [CrossRef]
- Gorovits, R.; Moshe, A.; Kolot, M.; Sobol, I.; Czosnek, H. Progressive aggregation of Tomato yellow leaf curl virus coat protein in systemically infected tomato plants, susceptible and resistant to the virus. Virus Res. 2013, 171, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Lohmus, A.; Hafren, A.; Mäkinen, K. Coat protein regulation by CK2, CPIP, HSP70, and CHIP is required for potato virus A replication and coat protein accumulation. J. Virol. 2017, 91, e01316. [Google Scholar] [CrossRef]
- Jiang, J.; Kuo, Y.W.; Salem, N.; Erickson, A.; Falk, B.W. Carrot mottle virus ORF4 movement protein targets plasmodesmata by interacting with the host cell SUMOylation system. New Phytol. 2021, 231, 382–398. [Google Scholar] [CrossRef]
- Millar, A.H.; Taylor, N.L. The isolation of plant organelles and structures in the post-genomic era. In Isolation of Plant Organelles and Structures: Methods and Protocols; Springer: Berlin/Heidelberg, Germany, 2017; pp. 1–11. [Google Scholar]
- Agrawal, G.K.; Bourguignon, J.; Rolland, N.; Ephritikhine, G.; Ferro, M.; Jaquinod, M.; Alexiou, K.G.; Chardot, T.; Chakraborty, N.; Jolivet, P. Plant organelle proteomics: Collaborating for optimal cell function. Mass Spectrom. Rev. 2011, 30, 772–853. [Google Scholar] [CrossRef]
- Zhu, D.; Zhang, M.; Gao, C.; Shen, J. Protein trafficking in plant cells: Tools and markers. Sci. China Life Sci. 2020, 63, 343–363. [Google Scholar] [CrossRef]
- Zhao, J.; Xu, J.; Chen, B.; Cui, W.; Zhou, Z.; Song, X.; Chen, Z.; Zheng, H.; Lin, L.; Peng, J.; et al. Characterization of Proteins Involved in Chloroplast Targeting Disturbed by Rice Stripe Virus by Novel Protoplast-Chloroplast Proteomics. Int. J. Mol. Sci. 2019, 20, 253. [Google Scholar] [CrossRef]
- Lao, J.; Smith-Moritz, A.M.; Mortimer, J.C.; Heazlewood, J.L. Enrichment of the Plant Cytosolic Fraction. In Isolation of Plant Organelles and Structures; Springer: Berlin/Heidelberg, Germany, 2017; pp. 213–232. [Google Scholar]
- Jaquinod, M.; Villiers, F.; Kieffer-Jaquinod, S.; Hugouvieux, V.; Bruley, C.; Garin, J.; Bourguignon, J. A proteomics dissection of Arabidopsis thaliana vacuoles isolated from cell culture. Mol. Cell. Proteom. 2007, 6, 394–412. [Google Scholar] [CrossRef] [PubMed]
- Collins, C.A.; LaMontagne, E.D.; Anderson, J.C.; Ekanayake, G.; Clarke, A.S.; Bond, L.N.; Salamango, D.J.; Cornish, P.V.; Peck, S.C.; Heese, A. EPSIN1 Modulates the Plasma Membrane Abundance of FLAGELLIN SENSING2 for Effective Immune Responses. Plant Physiol. 2020, 182, 1762–1775. [Google Scholar] [CrossRef] [PubMed]
- Zouhar, J. Isolation of Vacuoles and the Tonoplast. In Isolation of Plant Organelles and Structures; Springer: Berlin/Heidelberg, Germany, 2017; pp. 113–118. [Google Scholar]
- Simon, W.J.; Maltman, D.J.; Slabas, A.R. Isolation and fractionation of the endoplasmic reticulum from castor bean (Ricinus communis) endosperm for proteomic analyses. In 2D PAGE: Sample Preparation and Fractionation; Springer: Berlin/Heidelberg, Germany, 2008; pp. 203–215. [Google Scholar]
- Belykh, E.S.; Velegzhaninov, I.O.; Garmash, E.V. Responses of genes of DNA repair, alternative oxidase, and pro-/antioxidant state in Arabidopsis thaliana with altered expression of AOX1a to gamma irradiation. Int. J. Radiat. Biol. 2022, 98, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Chai, Y.; Yang, H.; Tian, Z.; Li, C.; Xu, R.; Shi, C.; Zhu, F.; Zeng, Y.; Deng, X. Isolation and comparative proteomic analysis of mitochondria from the pulp of ripening citrus fruit. Hortic. Res. 2021, 8, 31. [Google Scholar] [CrossRef]
- Hurný, A.; Cuesta, C.; Cavallari, N.; Ötvös, K.; Duclercq, J.; Dokládal, L.; Montesinos, J.C.; Gallemí, M.; Semerádová, H.; Rauter, T.; et al. SYNERGISTIC ON AUXIN AND CYTOKININ 1 positively regulates growth and attenuates soil pathogen resistance. Nat. Commun. 2020, 11, 2170. [Google Scholar] [CrossRef] [PubMed]
- Zeng, W.; Ebert, B.; Parsons, H.T.; Rautengarten, C.; Bacic, A.; Heazlewood, J.L. Enrichment of Golgi membranes from Triticum aestivum (wheat) seedlings. In Isolation of Plant Organelles and Structures; Springer: Berlin/Heidelberg, Germany, 2017; pp. 131–150. [Google Scholar]
- Jensen, G.S.; Fal, K.; Hamant, O.; Haswell, E.S. The RNA Polymerase-Associated Factor 1 Complex Is Required for Plant Touch Responses. J. Exp. Bot. 2016, 68, 499–511. [Google Scholar] [CrossRef] [PubMed]
- Shibahara, T.; Kawasaki, H.; Hirano, H. Identification of the 19S regulatory particle subunits from the rice 26S proteasome. Eur. J. Biochem. 2002, 269, 1474–1483. [Google Scholar] [CrossRef] [PubMed]
- Marshall, R.S.; Gemperline, D.C.; Vierstra, R.D. Purification of 26S proteasomes and their subcomplexes from plants. In Isolation of Plant Organelles and Structures: Methods and Protocols; Springer: Berlin/Heidelberg, Germany, 2017; pp. 301–334. [Google Scholar]
- Minami, A.; Takahashi, D.; Kawamura, Y.; Uemura, M. Isolation of plasma membrane and plasma membrane microdomains. In Isolation of Plant Organelles and Structures; Springer: Berlin/Heidelberg, Germany, 2017; pp. 199–212. [Google Scholar]
- Xu, Y.; Li, X.; Huang, J.; Peng, L.; Luo, D.; Zhang, Q.; Dan, Z.; Xiao, H.; Yang, F.; Hu, J. A simplified method to isolate rice mitochondria. Plant Methods 2020, 16, 1–10. [Google Scholar] [CrossRef]
- Reumann, S.; Singhal, R. Isolation of Leaf Peroxisomes from Arabidopsis for Organelle Proteome Analyses. In Plant Proteomics: Methods and Protocols; Jorrin-Novo, J.V., Komatsu, S., Weckwerth, W., Wienkoop, S., Eds.; Humana Press: Totowa, NJ, USA, 2014; pp. 541–552. [Google Scholar]
- Bao, Y.; Mugume, Y.; Bassham, D. Biochemical methods to monitor autophagic responses in plants. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 2017; Volume 588, pp. 497–513. [Google Scholar]
- Takatsuka, C.; Inoue-Aono, Y.; Moriyasu, Y. Isolation of autolysosomes from tobacco BY-2 Cells. In Isolation of Plant Organelles and Structures; Springer: Berlin/Heidelberg, Germany, 2017; pp. 151–161. [Google Scholar]
- Williams, A.J.; Werner-Fraczek, J.; Chang, I.-F.; Bailey-Serres, J. Regulated phosphorylation of 40S ribosomal protein S6 in root tips of maize. Plant Physiol. 2003, 132, 2086–2097. [Google Scholar] [CrossRef]
- Klang Årstrand, H.; Turkina, M.V. Isolation of cytosolic ribosomes. In Isolation of Plant Organelles and Structures; Springer: Berlin/Heidelberg, Germany, 2017; pp. 241–247. [Google Scholar]
- Chen, A.; He, B.; Jin, H. Isolation of extracellular vesicles from arabidopsis. Curr. Protoc. 2022, 2, e352. [Google Scholar] [CrossRef]
- Rutter, B.D.; Innes, R.W. Extracellular vesicles isolated from the leaf apoplast carry stress-response proteins. Plant Physiol. 2017, 173, 728–741. [Google Scholar] [CrossRef] [PubMed]
- Tuli, L.; Ressom, H.W. LC–MS based detection of differential protein expression. J. Proteom. Bioinform. 2009, 2, 416. [Google Scholar] [CrossRef] [PubMed]
- Vandenbogaert, M.; Li-Thiao-Té, S.; Kaltenbach, H.M.; Zhang, R.; Aittokallio, T.; Schwikowski, B. Alignment of LC-MS images, with applications to biomarker discovery and protein identification. Proteomics 2008, 8, 650–672. [Google Scholar] [CrossRef] [PubMed]
- Schaad, M.C.; Jensen, P.E.; Carrington, J.C. Formation of plant RNA virus replication complexes on membranes: Role of an endoplasmic reticulum-targeted viral protein. Embo J. 1997, 16, 4049–4059. [Google Scholar] [CrossRef] [PubMed]
- Jean Beltran, P.M.; Federspiel, J.D.; Sheng, X.; Cristea, I.M. Proteomics and integrative omic approaches for understanding host–pathogen interactions and infectious diseases. Mol. Syst. Biol. 2017, 13, 922. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.L.; Garrison, D.J.; May, J.P. Phase separation of a plant virus movement protein and cellular factors support virus-host interactions. PLoS Pathog. 2021, 17, e1009622. [Google Scholar] [CrossRef] [PubMed]
- Doucette, A.A.; Tran, J.C.; Wall, M.J.; Fitzsimmons, S. Intact proteome fractionation strategies compatible with mass spectrometry. Expert Rev. Proteom. 2011, 8, 787–800. [Google Scholar] [CrossRef]
- Low, T.Y.; Syafruddin, S.E.; Mohtar, M.A.; Vellaichamy, A.; A Rahman, N.S.; Pung, Y.-F.; Tan, C.S.H. Recent progress in mass spectrometry-based strategies for elucidating protein–protein interactions. Cell. Mol. Life Sci. 2021, 78, 5325–5339. [Google Scholar] [CrossRef]
- Li, X.; Wang, W.; Chen, J. From pathways to networks: Connecting dots by establishing protein–protein interaction networks in signaling pathways using affinity purification and mass spectrometry. Proteomics 2015, 15, 188–202. [Google Scholar] [CrossRef]
- Yakubu, R.R.; Nieves, E.; Weiss, L.M. The methods employed in mass spectrometric analysis of posttranslational modifications (PTMs) and protein–protein interactions (PPIs). In Advancements of Mass Spectrometry in Biomedical Research; Springer: Berlin/Heidelberg, Germany, 2019; pp. 169–198. [Google Scholar]
Figure 1.
Subcellular distribution of potyviral proteins withing plant cell organelles. Potyviruses encode 11 proteins that localize to specific subcellular compartments where they interact with host proteins. Cellular proteins may change their accumulation or localization during virus infection.
Figure 1.
Subcellular distribution of potyviral proteins withing plant cell organelles. Potyviruses encode 11 proteins that localize to specific subcellular compartments where they interact with host proteins. Cellular proteins may change their accumulation or localization during virus infection.
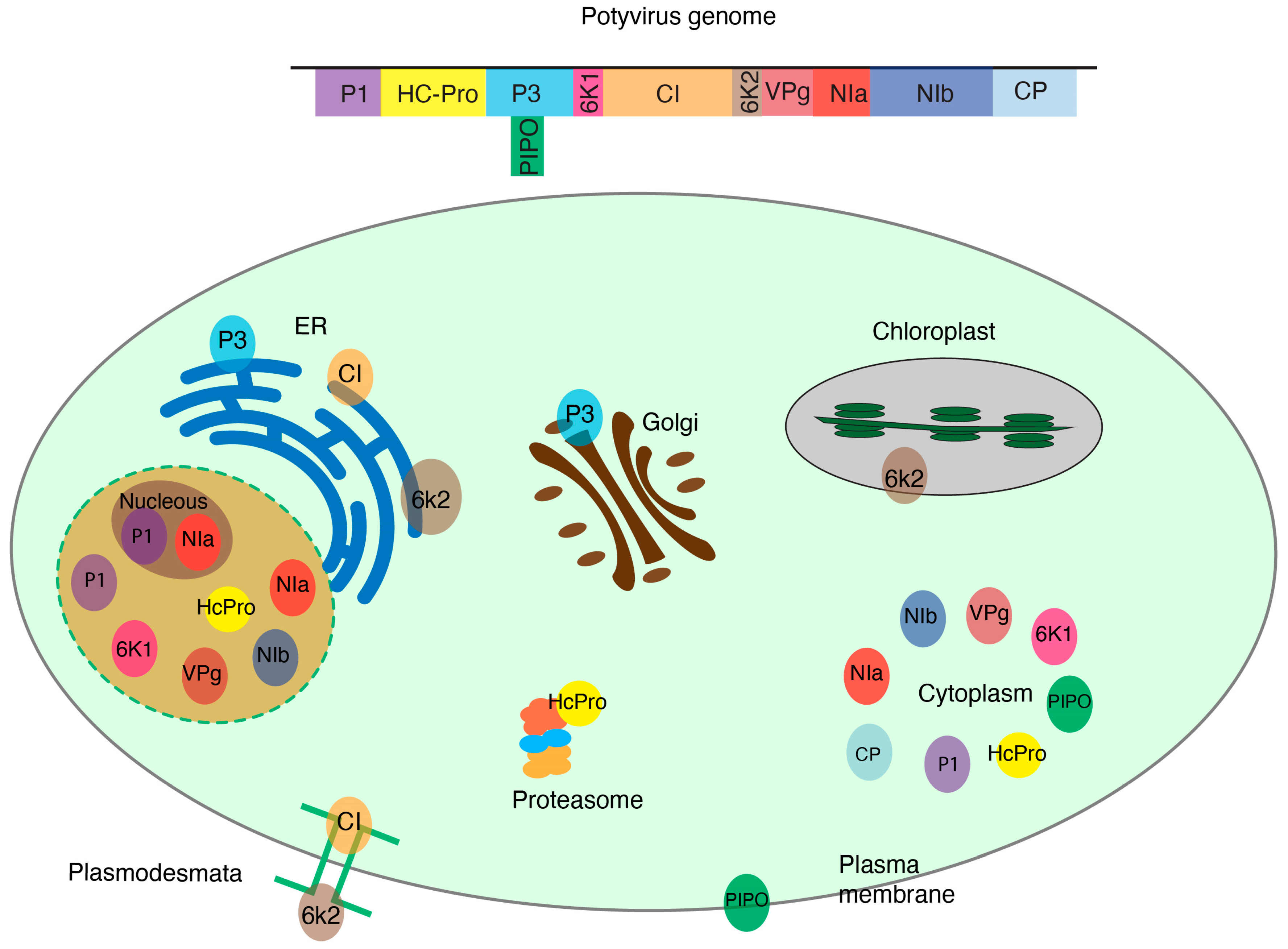
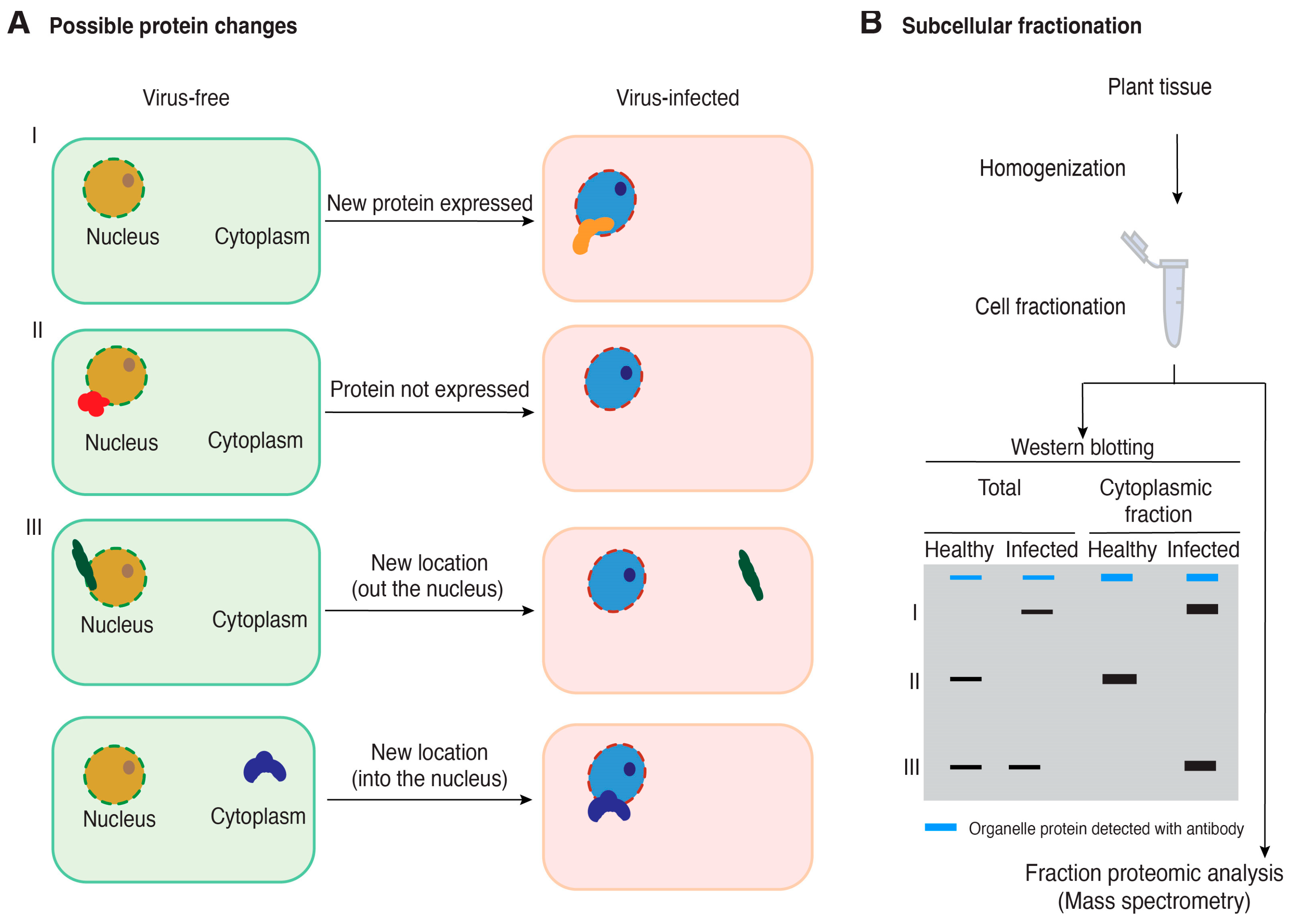
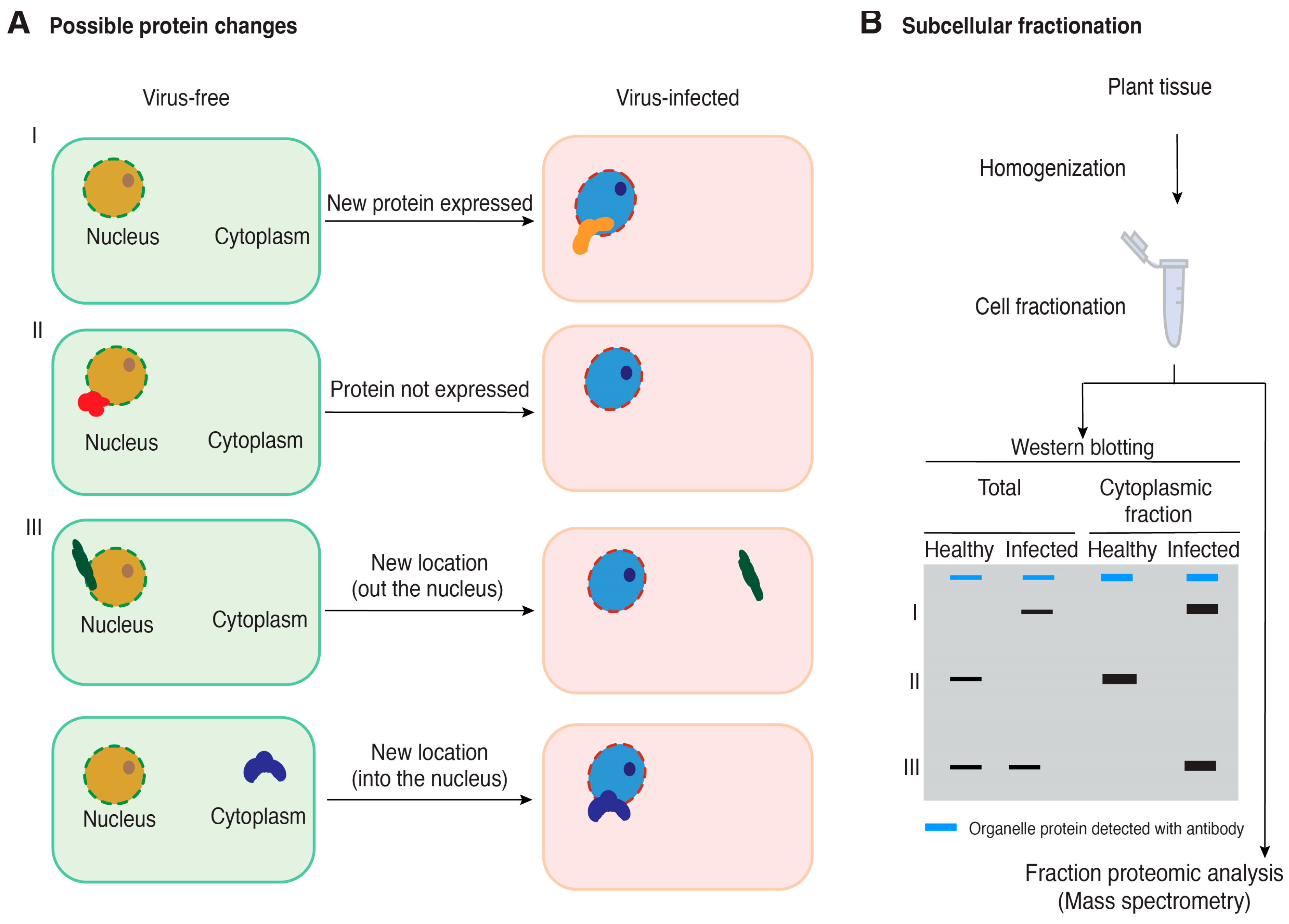
Figure 2.
A model for the possible changes in host protein accumulation and subcellular localization during virus infection and the use of cell fractionation coupled with mass spectrometry to identify proteins relevant to plant–virus interactions. (A) Possible changes in host protein accumulation and subcellular localization in virus-infected plants relative to healthy plants. (B) Cell fractionation and Western blotting for quality control before performing mass spectrometry to identify cellular proteins that change accumulation or subcellular localization in virus-infected plants. A blue band represents a cellular protein used as a marker to be detected using an antibody. Black bands represent hypothetical changes in accumulation and localization of host proteins for which antibodies are not available.
Figure 2.
A model for the possible changes in host protein accumulation and subcellular localization during virus infection and the use of cell fractionation coupled with mass spectrometry to identify proteins relevant to plant–virus interactions. (A) Possible changes in host protein accumulation and subcellular localization in virus-infected plants relative to healthy plants. (B) Cell fractionation and Western blotting for quality control before performing mass spectrometry to identify cellular proteins that change accumulation or subcellular localization in virus-infected plants. A blue band represents a cellular protein used as a marker to be detected using an antibody. Black bands represent hypothetical changes in accumulation and localization of host proteins for which antibodies are not available.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Gomaa, A.E.; El Mounadi, K.; Parperides, E.; Garcia-Ruiz, H. Cell Fractionation and the Identification of Host Proteins Involved in Plant–Virus Interactions. Pathogens 2024, 13, 53. https://doi.org/10.3390/pathogens13010053
AMA Style
Gomaa AE, El Mounadi K, Parperides E, Garcia-Ruiz H. Cell Fractionation and the Identification of Host Proteins Involved in Plant–Virus Interactions. Pathogens. 2024; 13(1):53. https://doi.org/10.3390/pathogens13010053
Chicago/Turabian StyleGomaa, Amany E., Kaoutar El Mounadi, Eric Parperides, and Hernan Garcia-Ruiz. 2024. "Cell Fractionation and the Identification of Host Proteins Involved in Plant–Virus Interactions" Pathogens 13, no. 1: 53. https://doi.org/10.3390/pathogens13010053
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.