Abstract
In tropical areas, the simultaneous transmission of multiple vector-borne diseases is common due to ecological factors shared by arthropod vectors. Malaria and dengue virus, transmitted by Anopheles and Aedes mosquitoes, respectively, are among the top vector-borne diseases that cause significant morbidity and mortality in endemic areas. Notably, tropical areas often have suitable conditions for the co-existence of these mosquito species, highlighting the importance of identifying markers that accurately indicate the risk of acquiring each specific disease entity. Aedes are daytime-biting mosquitoes, while Anopheles preferentially bite during the night. These biting patterns raise the possibility of concurrent exposure to bites from both species. This is important because mosquito saliva, deposited in the skin during blood feeding, induces immune responses that modulate pathogen establishment and infection. Previous studies have focused on characterizing such effects on the vector–pathogen interface for an individual pathogen and its mosquito vector. In this study, we evaluated associations between immune responses to salivary proteins from non-dengue and non-malaria vector mosquito species with clinical characteristics of malaria and dengue, respectively. Surprisingly, antibody responses against Anopheles antigens in dengue patients correlated with red blood cell count and hematocrit, while antibody responses against Aedes proteins were associated with platelet count in malaria patients. Our data indicate that concurrent exposure to multiple disease-carrying mosquito vectors and their salivary proteins with differing immunomodulatory properties could influence the transmission, pathogenesis, and clinical presentation of malaria, dengue fever, and other vector-borne illnesses.
1. Introduction
Malaria and dengue fever are vector-borne diseases of significant public health concern in tropical and subtropical areas around the globe [1]. Dengue virus (DENV) is endemic in Central and South America, with occasional outbreaks in the US, particularly in Texas and Florida [2,3]. Importantly, several recent reports have indicated the occurrence of local malaria transmission in the continental US, while this disease remains endemic in Latin America [4,5].
During transmission, mosquito-borne pathogens are usually deposited in the skin of the vertebrate host along with arthropod saliva during blood-feeding [6]. Compelling evidence suggests that mosquito saliva induces an important immunomodulatory effect on vertebrate responses at the bite site, eventually systemically enhancing pathogen transmission [7,8]. Furthermore, immunogenic salivary proteins induce antibody responses that vary depending on age, seasonality, and vector abundance [9,10]. Thus, immunoglobulin G (IgG) antibody responses to mosquito salivary proteins have been used as a proxy for exposure to mosquito bites and indirect markers for disease risk [11,12], with some studies suggesting potential sex-dependent responses to mosquito saliva [13], but such findings have been partially explained by physiological factors, host genetics, and gender-related social determinants, resulting in differences in exposure [14,15]. With the increase in global mosquito-borne infections, a better understanding of sex-dependent host responses may be critical to mitigating the negative consequences of vector–pathogen transfer and hormone-related antigen responses.
Currently, control of vector-borne diseases relies heavily on decreasing human–vector contact through physical devices (i.e., bed nets) or insecticide treatment [16,17], since effective vaccines or drugs are scarce [18,19]. Importantly, the increase in insecticide and drug resistance calls for the design and implementation of new tools for disease control and new tools to estimate the potential risk of acquisition of vector-borne diseases, which will guide public health policy. Recently, an Ae. aegypti peptide, Nterm-34kDa, has been recommended as a tool to measure exposure to Aedes bites since a positive correlation was observed between the intensity of exposure, mosquito abundance, and anti-Nterm-34kDa IgG antibody levels [20,21]. Also, the gSG6-P1 peptide, identified from the An. gambiae SG6 salivary protein [22] has been extensively validated as a biomarker of exposure against Anopheles mosquitoes from the subgenus Cellia and Anopheles [23]. Both peptides have been used successfully to determine the level of exposure to mosquito bites associated with mosquito control interventions or risk of disease [12,24,25,26]. However, the SG6 protein is absent in the subgenus Nyssorhynchus, which includes An. albimanus and An. darlingi, we have designed several peptides from proteins present in these mosquitoes. Specifically, we have tested the An. albimanus peptides Peroxi-P1, Trans-1, and Trans-2 and the An. darlingi peptide AnDarApy-1, to be used in addition to the gSG6-P1 peptide, to have a better understanding of human–vector contact in Latin America. The peptides have previously been tested in several regions of Colombia, where we found that high IgG antibody levels are associated with malaria infection [27,28,29].
Prior work suggests that saliva induces skin responses associated with the potential for pathogen establishment [30]. In certain areas endemic for malaria or dengue, people can be exposed to hundreds of mosquito bites per day, and an increase in mosquito abundance is often associated with an increase in transmission [31]. It is well known that, in a specific geographical area, several mosquito species are in circulation simultaneously, although their feeding behavior may be different [32,33]. For instance, Anopheles mosquitoes are preferential nocturnal feeders, while Aedes mosquitoes are diurnal biters [34]. Therefore, it is possible that an individual may sustain Aedes mosquito bites during the day, followed by Anopheles mosquito bites at night with the potential for saliva from different mosquito species to have different immunomodulatory effects on the course of the infections with these pathogens [34]. To our knowledge, there are currently no studies that describe the effect of exposure to bites from different mosquito species on skin immune responses and pathogen replication. Thus, there is a critical need to better understand how contact with salivary proteins from different mosquito species due to sequential exposure to diurnal and nocturnal biters may impact human arbovirus acquisition and anti-viral immune responses.
In this study, our primary goal was to evaluate the levels of antibodies against salivary antigens of different arthropod vectors of human disease in people with either malaria or dengue fever to assess whether responses against vector saliva are associated with blood parameters leading to severe clinical presentation. Since it is rare that only a single species of mosquito is found in a specific area, and several arthropod-borne diseases are common in the tropics, we hypothesized that exposure to salivary proteins with different immunomodulatory properties would impact infection and progression to disease. Transmission of Plasmodium and DENV often co-occur in tropical areas, mainly because of the overlap in the ecological niches preferred by the main vectors of these infections [35,36]. We, therefore, leveraged our ongoing malaria and dengue surveillance study in Norte de Santander, Colombia, to evaluate exposure to Anopheles and Aedes mosquito saliva and compare these data with blood parameters at diagnosis [37]. In this area, all DENV serotypes have circulated over the years. In addition, malaria caused by Plasmodium falciparum and Plasmodium vivax has been reported, with P. vivax as the most prevalent. To our knowledge, this is the first study exploring a potential correlation between exposure to the saliva of non-vectors in the clinical presentation of disease reflected by blood parameters.
2. Materials and Methods
2.1. Human Sample Collection and Diagnosis
All protocols involving human subjects were reviewed and approved by Universidad de Pamplona and by the IRB of Kansas State University (IRB#1206). Written informed consent was obtained from all subjects, and blood samples were collected from each subject living in two areas with different endemicity levels for malaria and DENV in the department of Norte de Santander, Colombia. For the dengue study, we included samples from Los Patios (n = 76), Ocana (n = 19) and Cucuta (n = 29). For the malaria study, we included samples collected in Tibu (n = 34), Villa del Rosario (n = 3), and Tarra (n = 8). The sample size and ages of volunteers included in these studies are described in Table 1. Malaria diagnosis was completed using the Rapid Diagnostic Test Malaria (Xerion) and a thick blood smear evaluated by at least two experienced microscopists. Gametocyte carriage was performed by qRT-PCR on the Pvs25 and Pvs230 genes of P. vivax parasites, following the methods described elsewhere [38,39]. DENV diagnosis was performed using the Rapid Diagnostic Test, Cassette Dengue AG (Xerion) and confirmed by qRT-PCR [40]. We also tested 48 human samples collected from healthy volunteers living in Kansas (USA) and 55 living in Los Patios (Colombia) to measure antibody levels against the peptides in healthy/non-infected individuals living in both non-endemic areas (USA) and endemic areas (Colombia).

Table 1.
Description of the study population ages in the malaria endemic areas (Tibu and Tarra, Norte de Santander, Colombia) and dengue endemic area (Los Patios and Cucuta, Norte de Santander, Colombia) and healthy individuals from Los Patios (Colombia) and Manhattan, Kansas (USA).
2.2. Salivary Antigens
Salivary gland extracts (SGE) from An. albimanus (STECLA strain) were prepared as previously published [41]. In this study, SGE and the previously reported salivary peptides Nterm-34kDa (Ae. aegypti) and gSG6-P1 (An. gambiae), AnDarApy-1 (An. darlingi) [28], Peroxi-P1, Trans1, and Trans-2 (An. albimanus) [27], were used to evaluate exposure to mosquito bites [27]. Peptides were synthesized by Genscript (Piscataway, NJ, USA), dissolved in ultrapure water, and frozen at −80 °C until used as antigens in ELISA assays.
2.3. Human IgG Antibody Detection by ELISA
The level of human IgG antibodies against mosquito salivary proteins was determined by an indirect ELISA, following the methods published by Londono-Renteria et al [29,41,42]. Briefly, 96-well ELISA plates (Nunc-MaxiSorp, Nalgene Nunc International, Rochester, NY, USA) were coated with 50 μL/well of Ae. aegypti or An. albimanus salivary gland extract (SGE) in a final concentration of 1 µg/mL prepared in a coating solution (1X PBS). Each salivary peptide was used in a final concentration of 2 µg/mL. Serum samples were tested in duplicate in a 1/100 dilution. After washes, plates were incubated with horseradish peroxidase-conjugated goat anti-human IgG (1: 1000) (Abcam, Ab81202), and colorimetric development was obtained using tetra-methyl-benzidine (one-solution micro-well, Gene-Script, Piscataway, NJ, USA). The reaction was terminated with 1 M phosphoric acid, and the absorbance was measured at 450 nm. Two controls were included on each plate: (1) control blank: two wells with antigen and without sample as a control for nonspecific induction of color for any of the reagents used in the test; and (2) positive control: 1 control per plate to test plate variation and normalize OD (optical density) values. IgG antibody levels are reported as ΔOD = Average patient OD value (duplicate) less the Blank OD.
2.4. Data Analysis
The median OD value was selected for all IgG levels against salivary antigens to determine high (above the median) or low (below the median) antibody levels. Differences between two independent groups were tested using the nonparametric Mann–Whitney U test. Correlation analysis between age and antigens was performed using the Spearman correlation test. All differences were considered significant at p < 0.05. All statistical tests were performed using Prism version 10 (Graph Pad Software Inc., La Jolla, CA, USA).
3. Results
3.1. Plasmodium vivax Malaria and Exposure to the Non-Malaria Vector Aedes aegypti
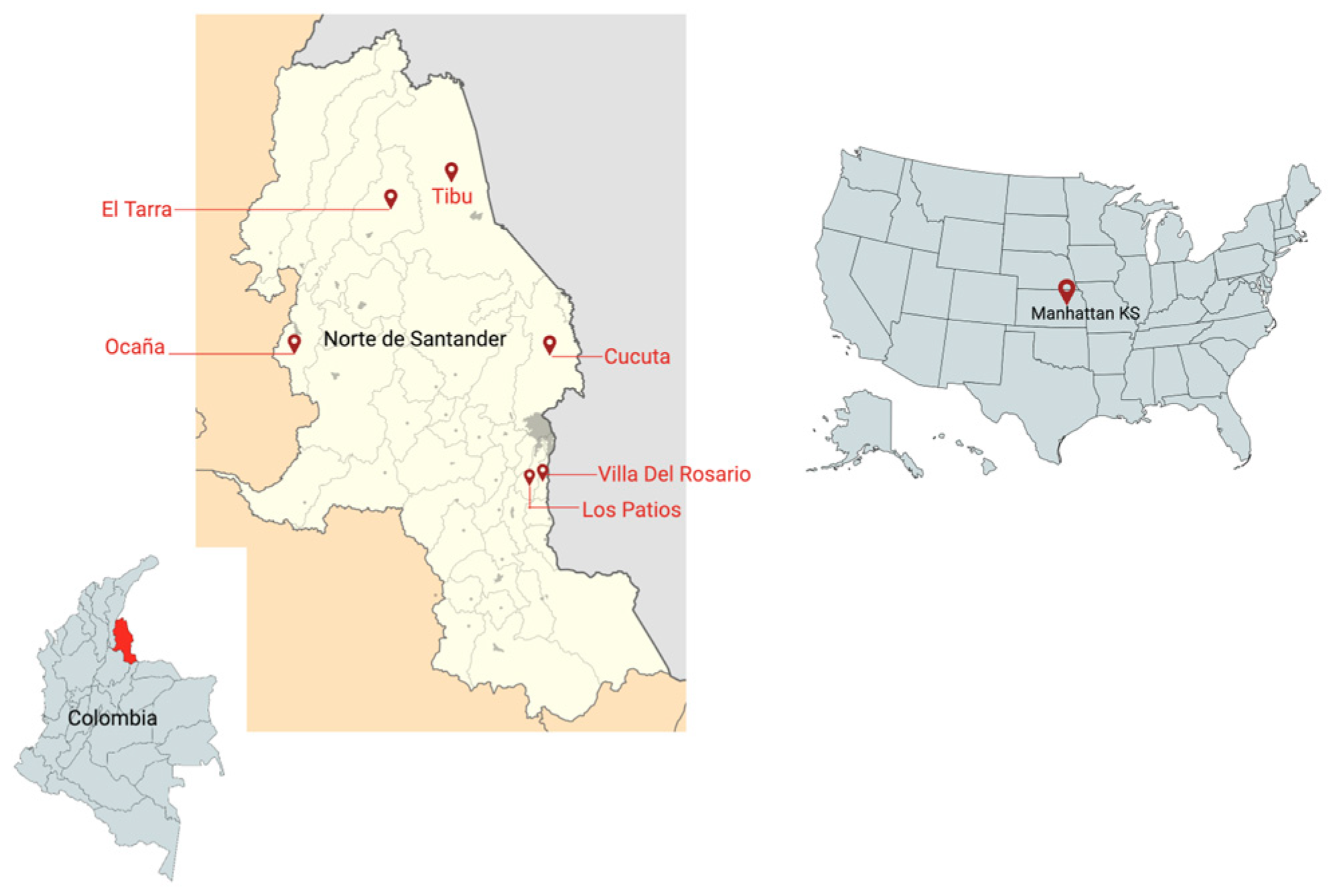
A total of 49 participants with current P. vivax malaria infections, diagnosed by microscopy and a rapid diagnostic test (RDT), from the areas of Tarra and Tibu in Norte de Santander, Colombia, were included in the study from 2018 to 2019 (Table 1) (Figure 1).
Figure 1.
Map with the study sites in Colombia and the United States of America.
The mean parasite count in our study population was 6665 parasites/µL (from 420 to 26,480 parasites/µL). Several malaria vectors circulate in our study area, including An. albimanus and An. nunestovary (Nyssorhynchus) and An. pseudopunctipennis and An. punctimacula (Anopheles), so we tested IgG antibodies using whole An. albimanus SGE, An. albimanus, An. darlingi, and gSG6-P1 peptides. We also measured exposure to Aedes bites in malaria-infected people, testing the IgG antibodies against the Ae. aegypti peptide Nterm34kDa. Since gametocytes are the Plasmodium parasite infective stages for the mosquito vector, we determined gametocyte carriage by measuring transcript levels of the Pvs25 and Pvs230 genes and comparing them with the IgG antibody levels against the mosquito salivary antigens. Our study did not reveal any correlation between the anti-saliva antibodies and parasite count or expression of the Pvs25 gene (gametes, ookinetes); however, IgG antibodies against An. albimanus SGE (Spearman correlation rs = −0.6099, p = 0.0269), Trans-1 (rs = −0.7510, p = 0.0031), gSG6-P1 (rs = −0.6648, p = 0.0132), and Nterm34kDa (Spearman correlation rs = −0.7253, p = 0.0050) showed a significant negative correlation with the level of expression of Pvs230 (gametocytes) in female participants (Table 2) (Supplementary Table S1).
Table 2.
Correlation analysis between IgG antibody responses against the Ae. Aegypti peptide Nterm34kDa and parasite count/gametocytemia by gender in Plasmodium-positive volunteers. Data are presented using Spearman correlation rs and significance p < 0.005.
We next tested the correlation between IgG anti-salivary protein antibody levels and blood parameters such as red blood cell (RBC) count, white blood cell (WBC) count, platelet count, hemoglobin, hematocrit, and parasite count (Table 3) (Supplementary Table S2). We found that the levels of IgG antibodies against the Ae. aegypti peptide Nterm34kDa (non-malaria vector) were negatively correlated with RBC count in males, while females presented a significant positive correlation between anti-Nterm34kDa antibodies and WBC count. We also compared antibody levels with age and observed a positive correlation with the level of antibodies against whole An. albimanus SGE, gSG6-P1, and Trans-1 in females but not males (Table 4) (Supplementary Table S3).
Table 3.
Correlation analysis between IgG antibody responses against Ae. aegypti Nterm34kDa peptide and blood parameters by gender in Plasmodium-positive volunteers. Data are presented using Spearman correlation rs and significance p < 0.005.
Table 4.
Correlation analysis between IgG antibody responses against each peptide and age by gender in P. vivax-infected patients. Data are presented using Spearman correlation rs and significance p < 0.005.
3.2. DENV and Exposure to the Non-DENV Vector Anopheles albimanus
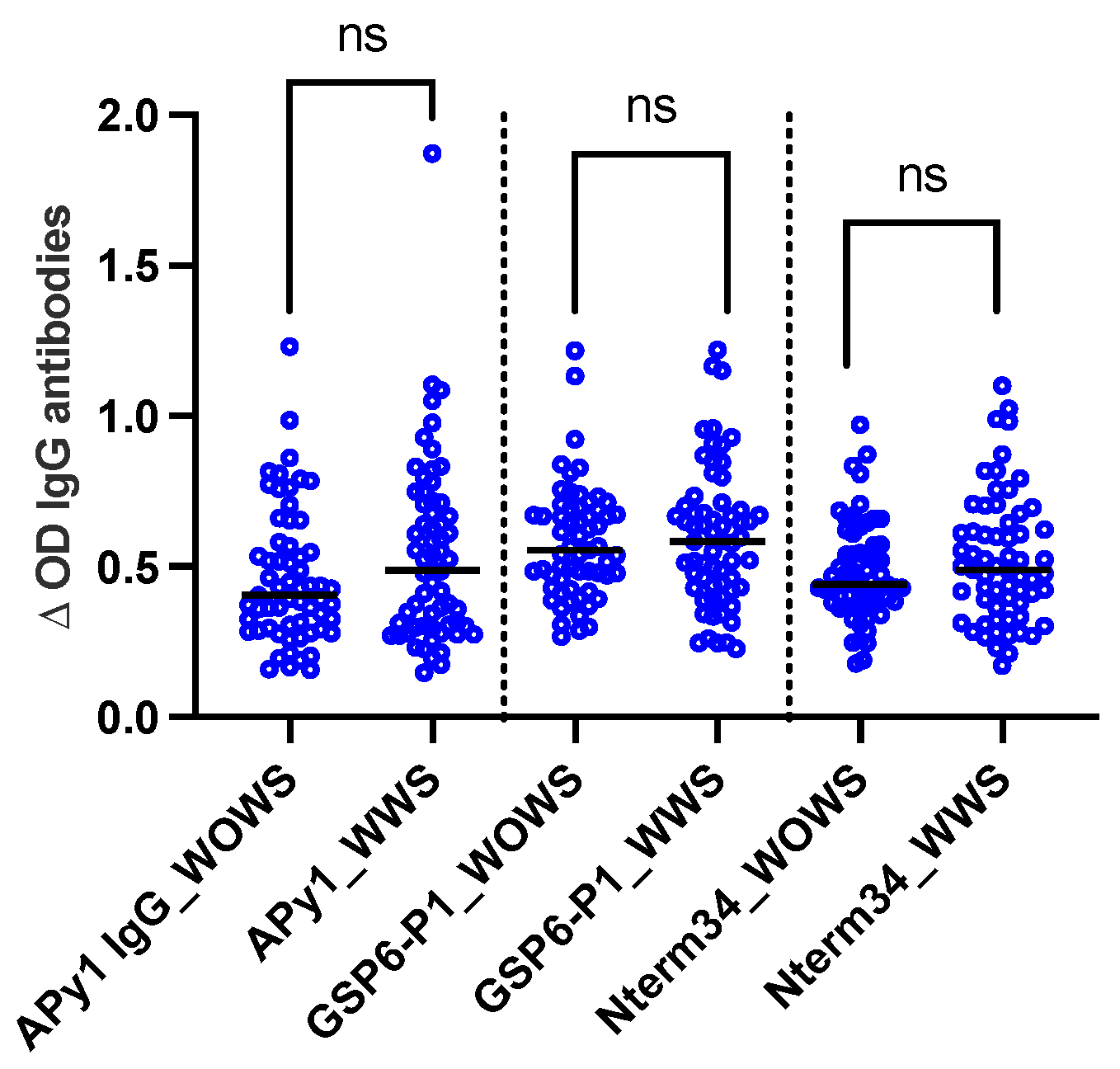
A total of 124 DENV-positive volunteers living in Los Patios (n = 75), Cucuta (n = 24), and Ocana (n = 25) between October 2018 and September 2020 were included in this study. Following DENV classification according to WHO guidelines, we included 63 DENV patients with warning signs and 61 DENV patients without warning signs. We did not observe significant differences in the level of IgG antibody levels against Nterm34kDa, AnDarApy1, and gSG6-P1 peptides when comparing DENV groups (Mann–Whitney test, p > 0.05) (Figure 2).
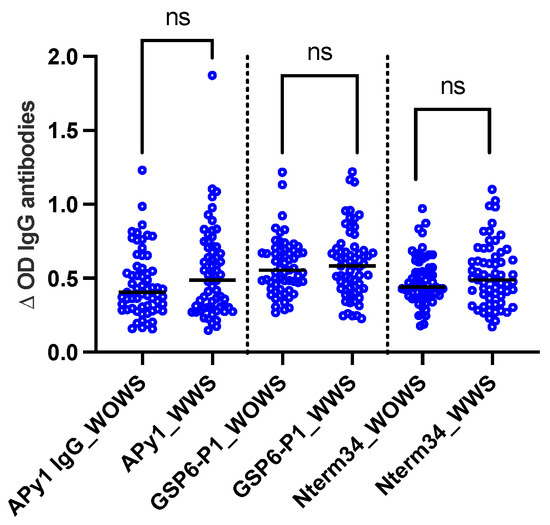
Figure 2.
Scatterplot of IgG antibodies against the mosquito salivary peptides Nterm34kDa, AnDarApy1, and gSG6-P1 in people with dengue fever with warning signs (WWS) and dengue without warning signs. Mann–Whitney test, p > 0.05, ns = not significant.
However, we observed a significant positive correlation between RBC count and IgG antibody levels against the Nterm34kDa peptide (Spearman correlation rs = −0.2107, p = 0.0193) and gSG6-P1 (rs = 0.1807, p = 0.0455) in males but not females. All comparisons are found in Table 5.
Table 5.
Correlation analysis between IgG antibody responses against Anopheles antigens and blood parameters by gender. In dengue fever patients. Data are presented using Spearman correlation rs and significance p < 0.005.
3.3. Healthy Individuals from Endemic and Non-Endemic Areas
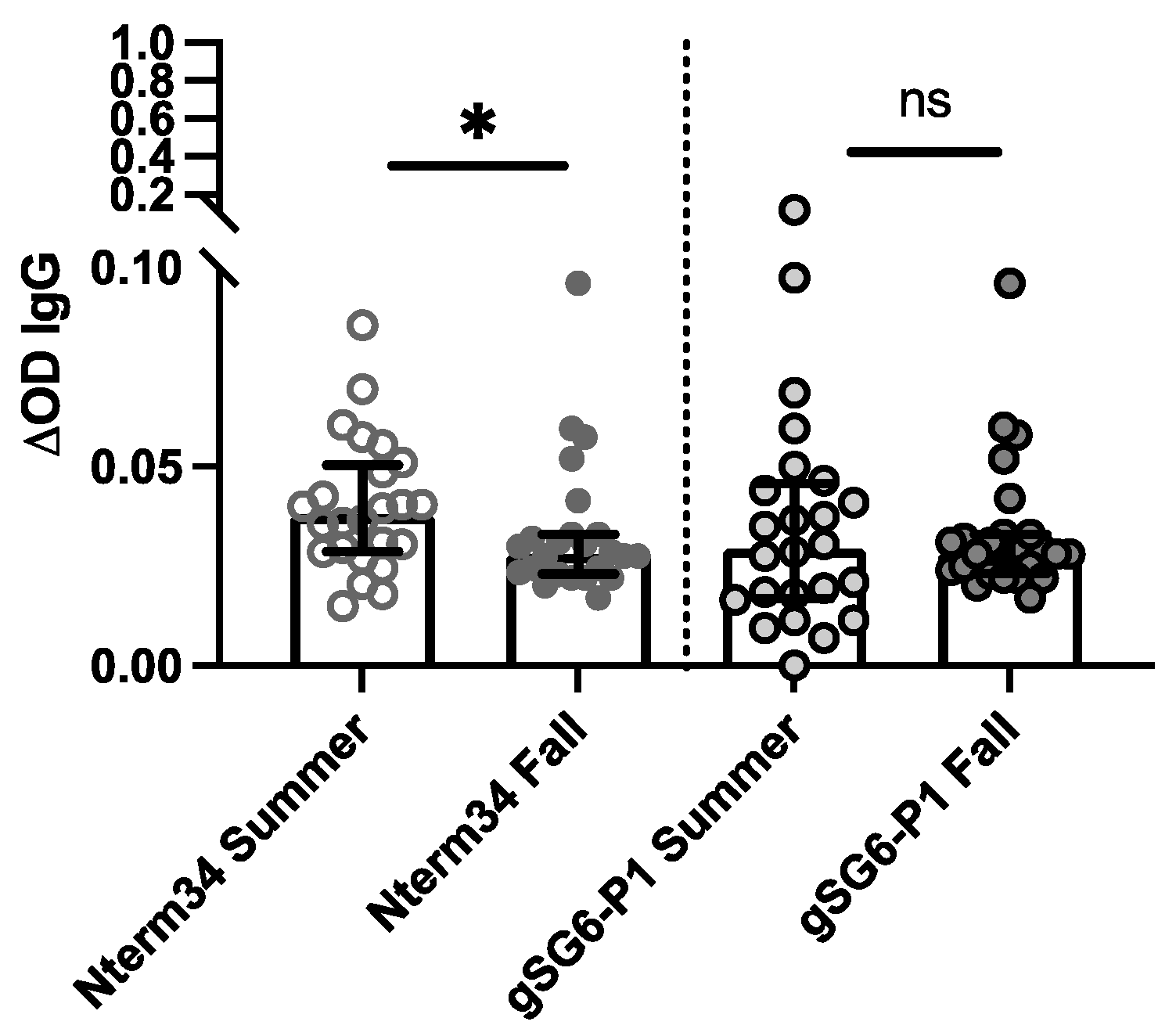
In 2018, we recruited participants in Manhattan, Kansas for a study focused on evaluating exposure to blood-sucking arthropods. A total of 27 participants donated blood samples in the summer (June–August) and fall (September–October). Our analyses revealed a significant reduction in antibody levels against Nterm-34kDa but not for gSG6-P1 in the Fall (Figure 3), suggesting an association between the intensity of exposure to mosquito bites and IgG antibody levels against mosquito salivary peptides.
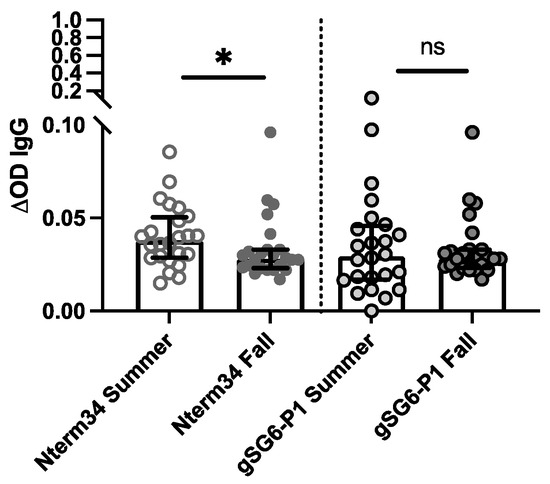
Figure 3.
IgG antibody levels against the mosquito salivary peptides in healthy participants residents of Kansas (USA) in the summer of 2018 compared to levels in the IgG antibody levels found in volunteers followed during the Fall of 2018 (n = 25). Significance was measured by the Mann–Whitney test p < 0.05 (* = 0.0, and ns = not significant).
Next, we compared antibody levels in healthy US participants with levels in healthy individuals living in areas with endemic DENV and malaria. In 2018, samples were collected from 54 healthy volunteers living in houses where a DENV case was reported in Los Patios, Norte de Santander (Colombia). The samples were tested for the presence of asymptomatic infections through rapid tests and RT-PCR [43]. We observed a significant negative correlation between age and IgG antibodies against Nterm34kDa (rs = 0.4182, p = 0.000) and gSG6-P1 (rs = −0.3553, p = 0.0003). This significant negative correlation between age and anti-Nterm34kDa remained even when stratifying the data by gender (Table 5). When stratifying data by location, we observed that people from the US demonstrated significant negative correlations between antibody levels against both peptides and age, while healthy individuals from Colombia showed a positive correlation between age and gSG6-P1, but not Nterm-34kDa (Table 6).
Table 6.
Correlation analysis between IgG antibody responses against each peptide and age. Data are presented using Spearman correlation rs.
3.4. Blood Parameters and Concordant Vector–Pathogen Interactions
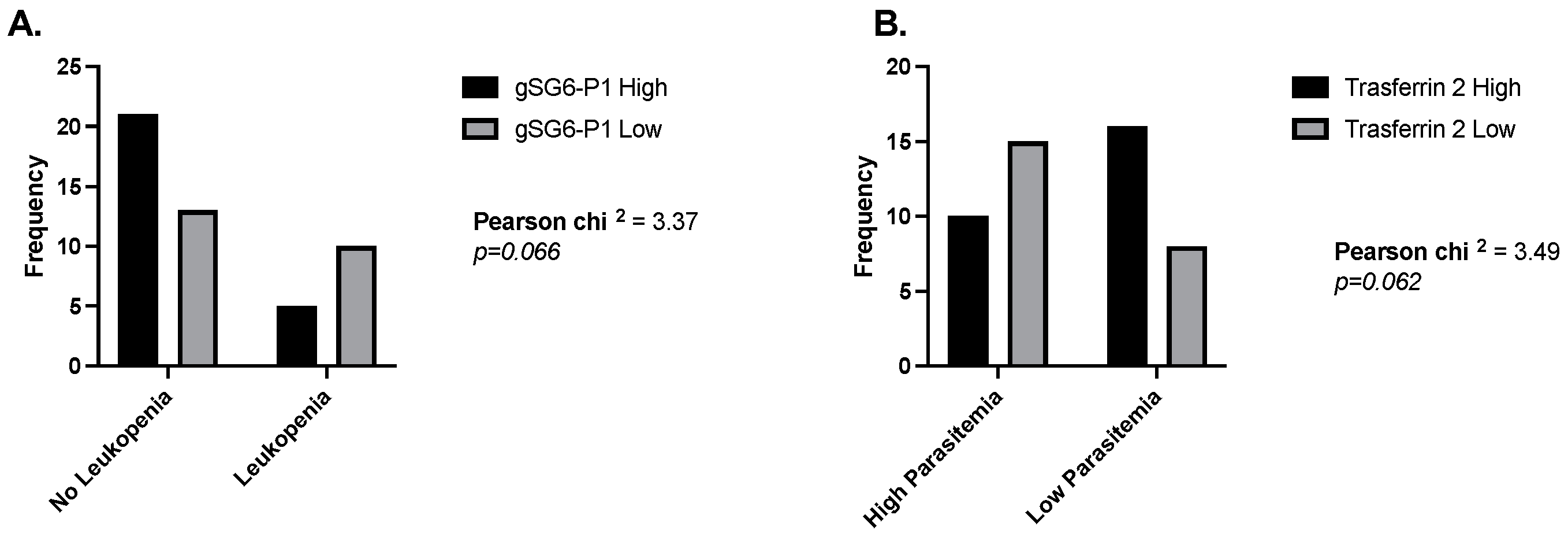
We have described the blood parameters that highlight a potential correlation or association between exposure to a pathogen and its non-vector salivary antigens. However, we also observed interesting results when analyzing the IgG responses to the salivary antigens of Anopheles mosquitoes in malaria patients. Specifically, when measuring odds ratios, we found that malaria patients with normal leucocyte counts were 3.3 times more likely to present high IgG antibodies against the gSG6-P1 peptide (Figure 4A), and people with high parasitemia were 3.3 times more likely to have low antibodies against Trans-2 (Figure 4B). We also observed a significant negative correlation between platelet count and each Anopheles antigen in people with malaria. In the case of dengue patients and the Aedes peptide Nterm34kDa, we observed a significant positive correlation between this peptide and RBC count.
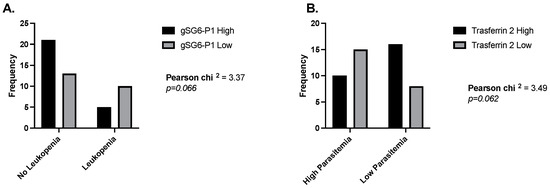
Figure 4.
Odds ratios comparing levels of IgG antibodies against gSG6-P1 and Trans-2 peptide in people with and without leukemia (A) and low and high parasitemia levels (B).
4. Discussion
Mosquito saliva is composed of a plethora of molecules that are injected into the skin to counteract host responses and facilitate blood uptake. This saliva also induces the production of host antibodies that correlate with the intensity of mosquito exposure. Notably, levels of IgG antibodies against whole or specific salivary proteins have been categorized as a reliable tool to measure exposure to mosquito bites and disease transmission intensity [44,45]. In malaria, Plasmodium gametocytes in vertebrate blood are the infectious stages for the mosquito vector. Interestingly, previous studies showed that the carriage of specific parasite stages influenced host attractiveness to mosquitoes, with people who harbored gametocytes in their blood being more attractive to mosquitoes than those presenting parasitemia with only asexual stages [38,39,46,47]. Here, we evaluated gametocyte carriage by measuring the expression of Plasmodium vivax Pvs25, while Pvs230 was used to determine gametocyte carriage [48]. Upon analysis of the level of antibodies against mosquito saliva, we observed that the expression of Pvs230 was significantly negatively correlated with IgG antibody levels of all mosquito antigens tested in this study, including the non-malaria vector Aedes peptide Nterm34kDa. However, this association was not observed with Pvs25, suggesting that expression of Pvs230 may be associated with exposure to mosquito bites from both Anopheles and Aedes mosquitoes. Detection of parasite carriage in the malaria elimination era is crucial to determine the risk of reemergence or roadblocks in control interventions. Previous studies have determined gametocyte carriage by detecting transcripts of both Pvs25 and Pvs230. Although Pvs25 is expressed in the mosquito stages, there is evidence that the transcription of this gene starts at the mature gametocyte stage, but the protein is expressed only in the zygote when it is transforming into ookinete [48,49,50]. Studies suggest that Pvs25 is expressed in female gametocytes and Pvs230 is considered specific to male gametocytes [51]. Thus, we aimed to evaluate if there was a relationship between gametocyte carriage with these two genes. It was interesting to see a correlation between anti-saliva antibodies and Pvs230 but not Pvs25, and it may be associated with the fact that each male gametocyte (microgametocyte) maturates into eight microgametes while one female gametocyte (macrogametocyte) develops into one macrogamete [52]. However, more studies are needed to establish this correlation.
Previous entomological data from our studies sites in Norte de Santander indicate that Anopheles and Aedes mosquitoes are prevalent in the area [53,54]. However, a limitation of this study is that the gSG6-P1 and the Nterm34kDa peptides we evaluated as markers of mosquito exposure have not been extensively validated for their correlation with mosquito abundance [55,56,57], and we did not collect specific mosquito data from the sites in our study, which could be used to confirm exposure to these mosquito species.
One of the objectives of this study was to determine if antibody levels against mosquito saliva were correlated with blood parameters. In the case of the malaria cohort, we observed that IgG antibodies against the non-malaria vector Aedes peptide Nterm34kDa were negatively associated with RBC count. This finding is notable as one of the signs of severe malaria is anemia, yet we did not observe correlations between any of the Anopheles salivary antigens and RBC count. The observed negative correlation between IgG anti-Nter34kDa and RBC count suggests that individuals with the lowest RBC counts may have the highest exposure to non-malaria vector Ae. aegypti or may produce more antibodies against their saliva. Interestingly, a previous study reported that anemia accelerates blood intake by Ae. aegypti, although it may negatively impact egg production [58]. Also, prior work has demonstrated that DENV acquisition by Ae. aegypti was inversely correlated with iron concentration in human serum [59]. Thus, it is possible that DENV benefits from the Aedes’ preference to feed on anemic individuals. Indeed, among our DENV-infected participants, we observed a significant positive correlation between RBC and IgG antibody responses to both Nterm34kDa and gSG6-P1 peptides, suggesting increased exposure to mosquito bites while infected. Moreover, a recent study suggested that infection with DENV increased attractiveness for Aedes mosquitoes [60], similar to observations made with Anopheles mosquitoes and Plasmodium gametocyte carriers [39,46].
Although the Ae. aegypti peptide Nterm34kDa has been reported useful for evaluating exposure to Aedes mosquitoes around the world, the gSG6-P1 is restricted to the evaluation of exposure to the bite of Anopheles mosquitoes from the subgenera Cellia and Anopheles since the gSG6 protein is absent in the Nyssorhynchus subgenus. Thus, we designed several peptides to evaluate exposure to An. darlingi and An. albimanus, major members of this subgenus and important malaria vectors in Latin America [27,28]. However, we acknowledge that there is a potential for cross-reactivity when using these peptides. Specifically, we reported that the An. darlingi apyrase, where the peptides were designed, has a 63% identity with an apyrase of An. gambiae (AGAP011971), 49% with an apyrase from Ae. aegypti (AAEL006347), and an apyrase of Cx. quinquefasciatus (CPIJ011010), but no significant similarity was found with An. albimanus apyrases. However, the An. darlingi salivary oxidase/peroxidase has an 87.8% identity with the An. albimanus peroxidase (AAD22196.1), a 53% similarity with An. gambiae (AGAP010735), while the similarity with Ae. aegypti (AAEL000507) and Cx. quinquefasciatus (CPIJ017579) was 48% [27,28]. Thus, it is possible that some of the IgG antibodies measured against these Anopheles-based peptides may reflect some exposure to Ae. aegypti. Our group is currently evaluating the potential cross-reactivity of both An. darlingi and An. albimanus peptides with Culex and Aedes species based on entomological collections in Norte de Santander in hopes of validating the usefulness of such peptides in vector-borne endemic areas with multiple mosquito species.
Mosquito abundance is associated with temperature, humidity, and other environmental factors [61,62,63]. In temperate and subtropical regions, mosquito abundance drastically changes in the summer vs. fall seasons, with higher mosquito populations in the former [64,65]. Previously, we observed a significant decrease in anti-tick antibodies during the fall months [66]. Interestingly, we found a significant decrease in IgG antibody levels against the Nter34kDa peptide in the fall compared to summer in people living in Kansas. Our results are in agreement with previous studies, which suggest a significant decrease in antibodies against Ae. aegypti mosquitoes after cessation of exposure [42]. However, we did not observe a decrease in antibodies against gSG6P1, although prior reports have indicated that antibodies against the gSG6 protein are short-lived16, suggesting a potential continued exposure. In Kansas, An. quadrimaculatus and An. pseudopunctipennis are abundantly found in the summer [67]. Ae. aegypti and Ae. albopictus have been collected in Kansas. However, only Ae. albopictus is found in Manhattan. Previous reports suggest that Nterm34kDa may cross-react with several Aedes species [21,40]. We were unable to find recent seasonal data on Anopheles captures, although a prior report describes the mosquito species present in Kansas from 1936 and 1937 [68]. Importantly, this previous report notes that extreme weather conditions were present in Kansas during the mosquito collections. Thus, it is possible that the species reported were behaving differently due to these unusual weather conditions. Despite the extreme weather conditions, the study reported several Anopheles species captured from June to November, which overlaps nicely with the two time points of our study (Summer 2018 (May–June) and Fall 2018 (September–November)) and could explain why the levels of antibodies against the gSG6-P1 peptide did not decrease during our follow-up. Because we could not find recent studies with the specific seasonality of Anopheles and Aedes mosquitoes endemic in Kansas, we also searched for any studies conducted in other countries with temperate climates in this hemisphere. A study in Northern Argentina indicated that An. pseudopunctipennis presents two peaks of abundance, one in the spring and another in fall/autumn [69]. Together, these studies provide a potential explanation for the persistence of antibody levels in samples collected in the early fall. Although this prior report did not include Ae. aegypti or Ae. albopictus, there is a description of Ae. vexans and Ae. triseriatus from the early summer to late fall, which could explain the minimal difference in antibody levels against the Nterm34kDa observed in our study.
We observed that healthy volunteers living in a non-endemic area (Kansas) for either malaria or dengue (Kansas) presented negative correlations between age and anti-saliva antibodies, while healthy people living in Los Patios, Norte de Santander showed a positive correlation. We found a negative correlation between age and antibodies against Ae. aegypti whole salivary gland extract in healthy individuals living in Norte de Santander in 2015 [9] that could be associated with the development of tolerance as observed in other studies [45,70]. In this study, we included samples from different regions in the State of Norte de Santander. However, in another study we did not observe any associations with age [43], as observed in similar studies [13], suggesting that several factors may contribute to these discrepancies. Similarly, our study demonstrated that two different localities showed differences in antibody responses with age. Taking all these studies into consideration, it is possible that the age correlation is associated with local environmental and entomological data. Further larger studies need to be performed to establish the relationship between age and response to arthropod salivary antigens.
In addition to previous studies showing sex-associated differences in DENV incidence, geographical area also appears to strongly influence the rates of association and occurrence. For instance, a study including DENV cases from at least six Asian countries showed a higher incidence of dengue fever in males [71], while other studies in Central and South America demonstrated a higher incidence in females [72,73]. Herein, we observed that males and females respond differently to mosquito salivary antigens. These results may have implications for pathogen transmission and clinical disease presentation. A major implication of our study is that characteristics, including age, seasonality, and vector control must be considered when assessing serum levels of IgG against Ae. aegypti salivary proteins as a surrogate of the risk of human exposure to mosquito bites and pathogen transmission. Such information is needed in epidemiological studies aimed at control and prevention of mosquito-borne diseases. Furthermore, data on biological sex at the population level could potentially be used to inform calculations of the total disease burden in regions where vector-borne diseases have the highest impact.
We also observed an interesting association between blood parameters and antibody responses against the arthropod vector or both Plasmodium and DENV. First, we found a significant negative correlation between Anopheles antigens and platelets in malaria patients and a positive association between Nterm34kDa and RBC cells in dengue patients. This is interesting because, although platelets may be associated with pathology during malaria [74], usually RBC count and anemia are hallmarks of people living in malaria endemic areas. The opposite has been observed for dengue fever, where a low platelet count can be associated with a severe presentation of dengue fever [75,76]. Yet, previous studies have described a significantly lower platelet count in males with malaria during the high transmission season [77], suggesting a sex-dependent association. In general, thrombocytopenia in malaria has been associated with an increase in endothelial activation [78] and with a widespread activation of platelets by an interaction between the parasite PfEMP1 and the platelet’s CD36 and other receptors [79], but the sex-dependent relationships are still under investigation. Several anticoagulants in mosquito saliva are known to interact directly with platelets and other modulate endothelial responses [80,81], but the significance of the levels of antibodies against the An. albimanus antigens and not against gSG6-P1 or Nterm34kDa needs more investigation. In the case of IgG antibodies against Nterm34kDa and RBC, the association was positive in dengue patients but negative in malaria patients. One of the characteristics of malaria is the rupture of red blood cells during the parasite replication, leading to anemia, meaning a malaria patient already has characteristics in the blood that may be more attractive to the Aedes mosquitoes. Also, IgG antibodies against Nterm34kDa in a DENV-infected patient may have resulted from the infected bite—precluding the clinical presentation. The correlation observed in the malaria group does not mean that all Aedes mosquitoes will only bite anemic patients. Mosquitoes feed on the hosts that are available, and, in a malaria endemic area, there may be far more anemic individuals available, as compared to a dengue endemic area, where Aedes mosquitos will have to feed on the individuals that are available, whether they are anemic or not. Our primary objective in this study was, therefore, to start the conversation on how the presence of several species of mosquitoes in a determined area could affect the course of diseases carried by these species. Our observations lead us to believe that there are parameters that require more attention from the scientific community that may help us to make significant advances in the control of diseases like dengue and malaria.
In summary, our data indicate that individuals living in malaria and dengue endemic areas possess antibodies against the salivary proteins from several different mosquito species that reside within these areas. Moreover, exposure to malaria and dengue mosquito vectors may be associated with clinical and immune responses to infection with the reciprocal pathogens transmitted by these specific species. Thus, our findings suggest that concurrent exposure to multiple disease-carrying mosquito vectors and their salivary proteins with differing immunomodulatory properties could influence the transmission and pathogenesis of malaria, dengue fever, and other vector-borne illnesses. A better understanding of the molecular mechanisms underlying how exposure to multiple and sequential bites from numerous mosquito species influences immune responses and pathogen transmission will advance the development of immune-targeting interventions to reduce disease spread. Additionally, including data on antibody responses against the main mosquito species found in specific areas will provide new knowledge on the interplay of such species, pathogen transmission, and disease severity in humans, which will help to inform the development of more effective vector control and disease prevention efforts.
5. Conclusions
Antibodies against salivary proteins are reliable markers for exposure intensity to bites from insect vectors. The data we have presented here support the use of IgG antibodies against salivary proteins as biomarkers of the intensity of exposure to mosquito bites and further indicate that these measurements can provide critical knowledge on how exposure to mosquito salivary proteins influences the transmission of vector-borne diseases. Moreover, including the potential effects of sequential exposure to saliva from different mosquito species will promote a better understanding of the pathogenesis of diseases borne by the vectors themselves as well as in the context of simultaneous exposure to non-vector mosquitos.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/pathogens13010052/s1, Table S1: Correlation analysis between IgG antibody responses against each Anopheles salivary antigen and parasite count/gametocytemia by gender in Plasmodium positive volunteers. Data is presented in Spearman rs and significance p < 0.005; Table S2: Correlation analysis between IgG antibody responses against Anopheles peptides and blood parameters by gender in Plasmodium positive volunteers. Data is presented in Spearman correlation rs and significance p < 0.005; Table S3: Correlation analysis between IgG antibody responses against the Ae. aegypti peptide Nterm34kDa and blood parameters by gender. In dengue fever patients. Data is presented in Spearman correlation rs and significance p < 0.005; Supplementary data S4.
Author Contributions
Conceptualization, B.L.-R. and J.A.M.; methodology, B.L.-R.; sample collection, J.C.C., C.A.P.-H., M.U.G.-P., L.Y.G.-S. and L.J.-V.; sample testing, M.M.H. and O.M.O.; formal analysis, M.M.H., O.M.O. and B.L.-R.; investigation, B.L.-R. and J.A.M.; resources, B.L.-R. and J.A.M.; writing—original draft preparation, B.L.-R., J.A.M., J.G.S. and B.M.W.; draft revisions B.L.-R., J.A.M., J.G.S. and B.M.W.; project administration, B.L.-R. and J.A.M.; funding acquisition, B.L.-R. and J.A.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Tulane Center of Excellence in Emergent Infectious Diseases (CEEIRD) and A Studio in The Woods Program.
Institutional Review Board Statement
All protocols involving human subjects were reviewed and approved by Universidad de Pamplona and by the IRB of Kansas State University (IRB#1206).
Informed Consent Statement
Written informed consent was obtained from all subjects.
Data Availability Statement
The data presented in this study is available as a Supplementary data.
Acknowledgments
The authors thank the residents of Los Patios, Cucuta, Tarra and Tibu (Colombia) and Manhattan, Kansas (USA) for their participation in this study.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Gebremariam, T.T.; Schallig, H.D.F.H.; Kurmane, Z.M.; Danquah, J.B. Increasing prevalence of malaria and acute dengue virus coinfection in Africa: A meta-analysis and meta-regression of cross-sectional studies. Malar. J. 2023, 22, 300. [Google Scholar] [CrossRef] [PubMed]
- Brathwaite Dick, O.; San Martín, J.L.; Montoya, R.H.; del Diego, J.; Zambrano, B.; Dayan, G.H. The history of dengue outbreaks in the Americas. Am. J. Trop. Med. Hyg. 2012, 87, 584–593. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Romieu, A.C.; Adams, L.E.; Paz-Bailey, G. Opportunities for Improved Dengue Control in the US Territories. JAMA 2023, 330, 19–20. [Google Scholar] [CrossRef] [PubMed]
- Bagcchi, S. Locally acquired malaria cases in the USA. Lancet Infect. Dis. 2023, 23, e401. [Google Scholar] [CrossRef] [PubMed]
- Agudelo Higuita, N.I.; Franco-Paredes, C.; Henao-Martínez, A.F.; Mendez Rojas, B.; Suarez, J.A.; Naranjo, L.; Alger, J. Migrants in transit across Central America and the potential spread of chloroquine resistant malaria-a call for action. Lancet Reg. Health Am. 2023, 22, 100505. [Google Scholar] [CrossRef] [PubMed]
- Coutinho-Abreu, I.V.; Guimaraes-Costa, A.B.; Valenzuela, J.G. Impact of insect salivary proteins in blood feeding, host immunity, disease, and in the development of biomarkers for vector exposure. Curr. Opin. Insect Sci. 2015, 10, 98–103. [Google Scholar] [CrossRef] [PubMed]
- Arca, B.; Ribeiro, J.M. Saliva of hematophagous insects: A multifaceted toolkit. Curr. Opin. Insect Sci. 2018, 29, 102–109. [Google Scholar] [CrossRef]
- Vogt, M.B.; Lahon, A.; Arya, R.P.; Kneubehl, A.R.; Spencer Clinton, J.L.; Paust, S.; Rico-Hesse, R. Mosquito saliva alone has profound effects on the human immune system. PLoS Negl. Trop. Dis. 2018, 12, e0006439. [Google Scholar] [CrossRef]
- Londoño-Rentería, B.; Cárdenas, J.C.; Giovanni, J.E.; Cárdenas, L.; Villamizar, P.; Rolón, J.; Chisenhall, D.M.; Christofferson, R.C.; Carvajal, D.J.; Pérez, O.G.; et al. Aedes aegypti anti-salivary gland antibody concentration and dengue virus exposure history in healthy individuals living in an endemic area in Colombia. Biomedica 2015, 35, 572–581. [Google Scholar] [CrossRef]
- Londono-Renteria, B.L.; Shakeri, H.; Rozo-Lopez, P.; Conway, M.J.; Duggan, N.; Jaberi-Douraki, M.; Colpitts, T.M. Serosurvey of Human Antibodies Recognizing Aedes aegypti D7 Salivary Proteins in Colombia. Front. Public Health 2018, 6, 111. [Google Scholar] [CrossRef]
- Londono-Renteria, B.; Drame, P.M.; Weitzel, T.; Rosas, R.; Gripping, C.; Cardenas, J.C.; Alvares, M.; Wesson, D.M.; Poinsignon, A.; Remoue, F.; et al. An. gambiae gSG6-P1 evaluation as a proxy for human-vector contact in the Americas: A pilot study. Parasit. Vectors 2015, 8, 533. [Google Scholar] [CrossRef]
- Ndille, E.E.; Dubot-Pérès, A.; Doucoure, S.; Mouchet, F.; Cornelie, S.; Sidavong, B.; Fournet, F.; Remoue, F. Human IgG antibody response to Aedes aegypti Nterm-34 kDa salivary peptide as an indicator to identify areas at high risk for dengue transmission: A retrospective study in urban settings of Vientiane city, Lao PDR. Trop. Med. Int. Health 2014, 19, 576–580. [Google Scholar] [CrossRef]
- Peng, Z.; Rasic, N.; Liu, Y.; Simons, F.E. Mosquito saliva-specific IgE and IgG antibodies in 1059 blood donors. J. Allergy Clin. Immunol. 2002, 110, 816–817. [Google Scholar] [CrossRef]
- Guerra-Silveira, F.; Abad-Franch, F. Sex bias in infectious disease epidemiology: Patterns and processes. PLoS ONE 2013, 8, e62390. [Google Scholar] [CrossRef]
- Pathak, S.; Rege, M.; Gogtay, N.J.; Aigal, U.; Sharma, S.K.; Valecha, N.; Bhanot, G.; Kshirsagar, N.A.; Sharma, S. Age-dependent sex bias in clinical malarial disease in hypoendemic regions. PLoS ONE 2012, 7, e35592. [Google Scholar] [CrossRef]
- Bardach, A.E.; Garcia-Perdomo, H.A.; Alcaraz, A.; Lopez, E.T.; Gandara, R.A.R.; Ruvinsky, S.; Ciapponi, A. Interventions for the control of Aedes aegypti in Latin America and the Caribbean: Systematic Review and Meta-Analysis. Trop. Med. Int. Health 2019, 24, 530–552. [Google Scholar] [CrossRef]
- Dhiman, S. Are malaria elimination efforts on right track? An analysis of gains achieved and challenges ahead. Infect. Dis. Poverty 2019, 8, 14. [Google Scholar] [CrossRef]
- Huang, Z.; Zhang, Y.; Li, H.; Zhu, J.; Song, W.; Chen, K.; Zhang, Y.; Lou, Y. Vaccine development for mosquito-borne viral diseases. Front. Immunol. 2023, 14, 1161149. [Google Scholar] [CrossRef]
- Qian, X.; Qi, Z. Mosquito-Borne Flaviviruses and Current Therapeutic Advances. Viruses 2022, 14, 1226. [Google Scholar] [CrossRef]
- Elanga Ndille, E.; Doucoure, S.; Poinsignon, A.; Mouchet, F.; Cornelie, S.; D’Ortenzio, E.; DeHecq, J.S.; Remoue, F. Human IgG Antibody Response to Aedes Nterm-34kDa Salivary Peptide, an Epidemiological Tool to Assess Vector Control in Chikungunya and Dengue Transmission Area. PLoS Negl. Trop. Dis. 2016, 10, e0005109. [Google Scholar] [CrossRef]
- Elanga Ndille, E.; Doucoure, S.; Damien, G.; Mouchet, F.; Drame, P.M.; Cornelie, S.; Noukpo, H.; Yamadjako, S.; Djenontin, A.; Moiroux, N.; et al. First attempt to validate human IgG antibody response to Nterm-34kDa salivary peptide as biomarker for evaluating exposure to Aedes aegypti bites. PLoS Negl. Trop. Dis. 2012, 6, e1905. [Google Scholar] [CrossRef]
- Poinsignon, A.; Cornelie, S.; Mestres-Simon, M.; Lanfrancotti, A.; Rossignol, M.; Boulanger, D.; Cisse, B.; Sokhna, C.; Arcà, B.; Simondon, F.; et al. Novel peptide marker corresponding to salivary protein gSG6 potentially identifies exposure to Anopheles bites. PLoS ONE 2008, 3, e2472. [Google Scholar] [CrossRef]
- Lombardo, F.; Ronca, R.; Rizzo, C.; Mestres-Simòn, M.; Lanfrancotti, A.; Currà, C.; Fiorentino, G.; Bourgouin, C.; Ribeiro, J.M.; Petrarca, V.; et al. The Anopheles gambiae salivary protein gSG6: An anopheline-specific protein with a blood-feeding role. Insect Biochem. Mol. Biol. 2009, 39, 457–466. [Google Scholar] [CrossRef]
- Drame, P.M.; Poinsignon, A.; Dechavanne, C.; Cottrell, G.; Farce, M.; Ladekpo, R.; Massougbodji, A.; Cornelie, S.; Courtin, D.; Migot-Nabias, F.; et al. Specific antibodies to Anopheles gSG6-P1 salivary peptide to assess early childhood exposure to malaria vector bites. Malar. J. 2015, 14, 285. [Google Scholar] [CrossRef]
- Drame, P.M.; Poinsignon, A.; Besnard, P.; Cornelie, S.; Le Mire, J.; Toto, J.C.; Foumane, V.; Dos-Santos, M.A.; Sembène, M.; Fortes, F.; et al. Human antibody responses to the Anopheles salivary gSG6-P1 peptide: A novel tool for evaluating the efficacy of ITNs in malaria vector control. PLoS ONE 2010, 5, e15596. [Google Scholar] [CrossRef]
- Sagna, A.B.; Yobo, M.C.; Elanga Ndille, E.; Remoue, F. New Immuno-Epidemiological Biomarker of Human Exposure to Aedes Vector Bites: From Concept to Applications. Trop. Med. Infect. Dis. 2018, 3, 80. [Google Scholar] [CrossRef]
- Londono-Renteria, B.; Drame, P.M.; Montiel, J.; Vasquez, A.M.; Tobon-Castano, A.; Taylor, M.; Vizcaino, L.; Lenhart, A.A.E. Identification and Pilot Evaluation of Salivary Peptides from Anopheles albimanus as Biomarkers for Bite Exposure and Malaria Infection in Colombia. Int. J. Mol. Sci. 2020, 21, 691. [Google Scholar] [CrossRef]
- Londono-Renteria, B.; Montiel, J.; Calvo, E.; Tobon-Castano, A.; Valdivia, H.O.; Escobedo-Vargas, K.; Romero, L.; Bosantes, M.; Fisher, M.L.; Conway, M.J.; et al. Antibody Responses against Anopheles darlingi Immunogenic Peptides in Plasmodium Infected Humans. Front. Cell Infect. Microbiol. 2020, 10, 455. [Google Scholar] [CrossRef]
- Montiel, J.; Carbal, L.F.; Tobón-Castaño, A.; Vásquez, G.M.; Fisher, M.L.; Londono-Rentería, B. IgG antibody response against Anopheles salivary gland proteins in asymptomatic Plasmodium infections in Narino, Colombia. Malar. J. 2020, 19, 42. [Google Scholar] [CrossRef]
- Visser, I.; Koenraadt, C.J.M.; Koopmans, M.P.G.; Rockx, B. The significance of mosquito saliva in arbovirus transmission and pathogenesis in the vertebrate host. One Health 2023, 16, 100506. [Google Scholar] [CrossRef]
- Bellone, R.; Failloux, A.B. The Role of Temperature in Shaping Mosquito-Borne Viruses Transmission. Front. Microbiol. 2020, 11, 584846. [Google Scholar] [CrossRef]
- Rozo-Lopez, P.; Mengual, X. Updated list of the mosquitoes of Colombia (Diptera: Culicidae). Biodivers. Data J. 2015, e4567. [Google Scholar] [CrossRef]
- Cano-Pérez, E.; González-Beltrán, M.; Ampuero, J.S.; Gómez-Camargo, D.; Morrison, A.C.; Astete, H. Prevalence of Mosquito Populations in the Caribbean Region of Colombia with Important Public Health Implications. Trop. Med. Infect. Dis. 2022, 8, 11. [Google Scholar] [CrossRef]
- Baik, L.S.; Nave, C.; Au, D.D.; Guda, T.; Chevez, J.A.; Ray, A.; Holmes, T.C. Circadian Regulation of Light-Evoked Attraction and Avoidance Behaviors in Daytime- versus Nighttime-Biting Mosquitoes. Curr. Biol. 2020, 30, 3252–3259.e3253. [Google Scholar] [CrossRef]
- Tazeen, A.; Abdullah, M.; Hisamuddin, M.; Ali, S.; Naqvi, I.H.; Verma, H.N.; Ahmed, A.; Parveen, S. Concurrent Infection with Plasmodium vivax and the Dengue and Chikungunya Viruses in a Paediatric Patient from New Delhi, India in 2016. Intervirology 2017, 60, 48–52. [Google Scholar] [CrossRef]
- Eder, M.; Cortes, F.; Teixeira de Siqueira Filha, N.; Araujo de Franca, G.V.; Degroote, S.; Braga, C.; Ridde, V.; Turchi Martelli, C.M. Scoping review on vector-borne diseases in urban areas: Transmission dynamics, vectorial capacity and co-infection. Infect. Dis. Poverty 2018, 7, 90. [Google Scholar] [CrossRef]
- Cardenas, J.C.; Giraldo-Parra, S.Y.; Gonzalez, M.U.; Gutierrez-Silva, L.Y.; Jaimes-Villamizar, L.; Roa-Parra, A.L.; Carvajal, D.J.; Valdivia, H.O.; Sanchez, J.F.; Colpitts, T.M.; et al. Laboratory Findings in Patients with Probable Dengue Diagnosis from an Endemic Area in Colombia in 2018. Viruses 2021, 13, 1401. [Google Scholar] [CrossRef]
- Robinson, A.; Busula, A.O.; Voets, M.A.; Beshir, K.B.; Caulfield, J.C.; Powers, S.J.; Verhulst, N.O.; Winskill, P.; Muwanguzi, J.; Birkett, M.A.; et al. Plasmodium-associated changes in human odor attract mosquitoes. Proc. Natl. Acad. Sci. USA 2018, 115, E4209–E4218. [Google Scholar] [CrossRef]
- Busula, A.O.; Bousema, T.; Mweresa, C.K.; Masiga, D.; Logan, J.G.; Sauerwein, R.W.; Verhulst, N.O.; Takken, W.; de Boer, J.G. Gametocytemia and Attractiveness of Plasmodium falciparum-Infected Kenyan Children to Anopheles gambiae Mosquitoes. J. Infect. Dis. 2017, 216, 291–295. [Google Scholar] [CrossRef]
- Yobo, C.M.; Sadia-Kacou, C.A.M.; Adja, M.A.; Elanga-Ndille, E.; Sagna, A.B.; Guindo-Coulibaly, N.; Poinsignon, A.; Remoue, F.; Koudou, B.G. Evaluation of Human Exposure to Aedes Bites in Rubber and Palm Cultivations Using an Immunoepidemiological Biomarker. Biomed. Res. Int. 2018, 2018, 3572696. [Google Scholar] [CrossRef]
- Londono-Renteria, B.L.; Eisele, T.P.; Keating, J.; James, M.A.; Wesson, D.M. Antibody response against Anopheles albimanus (Diptera: Culicidae) salivary protein as a measure of mosquito bite exposure in Haiti. J. Med. Entomol. 2010, 47, 1156–1163. [Google Scholar] [CrossRef] [PubMed]
- Londono-Renteria, B.; Cardenas, J.C.; Cardenas, L.D.; Christofferson, R.C.; Chisenhall, D.M.; Wesson, D.M.; McCracken, M.K.; Carvajal, D.; Mores, C.N. Use of anti-Aedes aegypti salivary extract antibody concentration to correlate risk of vector exposure and dengue transmission risk in Colombia. PLoS ONE 2013, 8, e81211. [Google Scholar] [CrossRef] [PubMed]
- Olajiga, O.M.; Marin-Lopez, A.; Cardenas, J.C.; Gutierrez-Silva, L.Y.; Gonzales-Pabon, M.U.; Maldonado-Ruiz, L.P.; Worges, M.; Fikrig, E.; Park, Y.; Londono-Renteria, B. Aedes aegypti anti-salivary proteins IgG levels in a cohort of DENV-like symptoms subjects from a dengue-endemic region in Colombia. Front. Epidemiol. 2022, 2, 1002857. [Google Scholar] [CrossRef]
- Rizzo, C.; Lombardo, F.; Ronca, R.; Mangano, V.; Sirima, S.B.; Nèbiè, I.; Fiorentino, G.; Modiano, D.; Arcà, B. Differential antibody response to the Anopheles gambiae gSG6 and cE5 salivary proteins in individuals naturally exposed to bites of malaria vectors. Parasit. Vectors 2014, 7, 549. [Google Scholar] [CrossRef] [PubMed]
- Doucoure, S.; Mouchet, F.; Cournil, A.; Le Goff, G.; Cornelie, S.; Roca, Y.; Giraldez, M.G.; Simon, Z.B.; Loayza, R.; Misse, D.; et al. Human antibody response to Aedes aegypti saliva in an urban population in Bolivia: A new biomarker of exposure to Dengue vector bites. Am. J. Trop. Med. Hyg. 2012, 87, 504–510. [Google Scholar] [CrossRef] [PubMed]
- Debebe, Y.; Hill, S.R.; Birgersson, G.; Tekie, H.; Ignell, R. Plasmodium falciparum gametocyte-induced volatiles enhance attraction of Anopheles mosquitoes in the field. Malar. J. 2020, 19, 327. [Google Scholar] [CrossRef] [PubMed]
- Lacroix, R.; Mukabana, W.R.; Gouagna, L.C.; Koella, J.C. Malaria infection increases attractiveness of humans to mosquitoes. PLoS Biol. 2005, 3, e298. [Google Scholar] [CrossRef] [PubMed]
- Wampfler, R.; Mwingira, F.; Javati, S.; Robinson, L.; Betuela, I.; Siba, P.; Beck, H.P.; Mueller, I.; Felger, I. Strategies for detection of Plasmodium species gametocytes. PLoS ONE 2013, 8, e76316. [Google Scholar] [CrossRef]
- Baton, L.A.; Ranford-Cartwright, L.C. Do malaria ookinete surface proteins P25 and P28 mediate parasite entry into mosquito midgut epithelial cells? Malar. J. 2005, 4, 15. [Google Scholar] [CrossRef]
- Essuman, E.; Grabias, B.; Verma, N.; Chorazeczewski, J.K.; Tripathi, A.K.; Mlambo, G.; Addison, E.A.; Amoah, A.G.B.; Quakyi, I.; Oakley, M.S.; et al. A Novel Gametocyte Biomarker for Superior Molecular Detection of the Plasmodium falciparum Infectious Reservoirs. J. Infect. Dis. 2017, 216, 1264–1272. [Google Scholar] [CrossRef]
- Schneider, P.; Reece, S.E.; van Schaijk, B.C.; Bousema, T.; Lanke, K.H.; Meaden, C.S.; Gadalla, A.; Ranford-Cartwright, L.C.; Babiker, H.A. Quantification of female and male Plasmodium falciparum gametocytes by reverse transcriptase quantitative PCR. Mol. Biochem. Parasitol. 2015, 199, 29–33. [Google Scholar] [CrossRef] [PubMed]
- Dash, M.; Sachdeva, S.; Bansal, A.; Sinha, A. Gametogenesis in Plasmodium: Delving Deeper to Connect the Dots. Front. Cell Infect. Microbiol. 2022, 12, 877907. [Google Scholar] [CrossRef] [PubMed]
- Carrillo, M.A.; Cardenas, R.; Yañez, J.; Petzold, M.; Kroeger, A. Risk of dengue, Zika, and chikungunya transmission in the metropolitan area of Cucuta, Colombia: Cross-sectional analysis, baseline for a cluster-randomised controlled trial of a novel vector tool for water containers. BMC Public Health 2023, 23, 1000. [Google Scholar] [CrossRef] [PubMed]
- Montoya-Lerma, J.; Solarte, Y.A.; Giraldo-Calderón, G.I.; Quiñones, M.L.; Ruiz-López, F.; Wilkerson, R.C.; González, R. Malaria vector species in Colombia: A review. Mem. Inst. Oswaldo Cruz 2011, 106 (Suppl. S1), 223–238. [Google Scholar] [CrossRef] [PubMed]
- Poinsignon, A.; Cornelie, S.; Ba, F.; Boulanger, D.; Sow, C.; Rossignol, M.; Sokhna, C.; Cisse, B.; Simondon, F.; Remoue, F. Human IgG response to a salivary peptide, gSG6-P1, as a new immuno-epidemiological tool for evaluating low-level exposure to Anopheles bites. Malar. J. 2009, 8, 198. [Google Scholar] [CrossRef] [PubMed]
- Sagna, A.B.; Sarr, J.B.; Gaayeb, L.; Drame, P.M.; Ndiath, M.O.; Senghor, S.; Sow, C.S.; Poinsignon, A.; Seck, M.; Hermann, E.; et al. gSG6-P1 salivary biomarker discriminates micro-geographical heterogeneity of human exposure to Anopheles bites in low and seasonal malaria areas. Parasit. Vectors 2013, 6, 68. [Google Scholar] [CrossRef]
- Ndo, C.; Elanga-Ndille, E.; Cheteug, G.; Metitsi, R.D.; Wanji, S.; Moukoko, C.E.E. IgG antibody responses to Anopheles gambiae gSG6-P1 salivary peptide are induced in human populations exposed to secondary malaria vectors in forest areas in Cameroon. PLoS ONE 2022, 17, e0276991. [Google Scholar] [CrossRef]
- Shieh, J.N.; Rossingnol, P.A. Opposite influences of host anaemia on blood feeding rate and fecundity of mosquitoes. Parasitology 1992, 105 Pt 2, 159–163. [Google Scholar] [CrossRef]
- Zhu, Y.; Tong, L.; Nie, K.; Wiwatanaratanabutr, I.; Sun, P.; Li, Q.; Yu, X.; Wu, P.; Wu, T.; Yu, C.; et al. Host serum iron modulates dengue virus acquisition by mosquitoes. Nat. Microbiol. 2019, 4, 2405–2415. [Google Scholar] [CrossRef]
- Zhang, H.; Zhu, Y.; Liu, Z.; Peng, Y.; Peng, W.; Tong, L.; Wang, J.; Liu, Q.; Wang, P.; Cheng, G. A volatile from the skin microbiota of flavivirus-infected hosts promotes mosquito attractiveness. Cell 2022, 185, 2510–2522.e2516. [Google Scholar] [CrossRef]
- Minakawa, N.; Sonye, G.; Mogi, M.; Githeko, A.; Yan, G. The effects of climatic factors on the distribution and abundance of malaria vectors in Kenya. J. Med. Entomol. 2002, 39, 833–841. [Google Scholar] [CrossRef] [PubMed]
- Asgarian, T.S.; Moosa-Kazemi, S.H.; Sedaghat, M.M. Impact of meteorological parameters on mosquito population abundance and distribution in a former malaria endemic area, central Iran. Heliyon 2021, 7, e08477. [Google Scholar] [CrossRef] [PubMed]
- Hwang, M.J.; Kim, H.C.; Klein, T.A.; Chong, S.T.; Sim, K.; Chung, Y.; Cheong, H.K. Comparison of climatic factors on mosquito abundance at US Army Garrison Humphreys, Republic of Korea. PLoS ONE 2020, 15, e0240363. [Google Scholar] [CrossRef] [PubMed]
- Reisen, W.K.; Cayan, D.; Tyree, M.; Barker, C.M.; Eldridge, B.; Dettinger, M. Impact of climate variation on mosquito abundance in California. J. Vector Ecol. 2008, 33, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Chuang, T.W.; Ionides, E.L.; Knepper, R.G.; Stanuszek, W.W.; Walker, E.D.; Wilson, M.L. Cross-correlation map analyses show weather variation influences on mosquito abundance patterns in Saginaw County, Michigan, 1989–2005. J. Med. Entomol. 2012, 49, 851–858. [Google Scholar] [CrossRef] [PubMed]
- Maldonado-Ruiz, L.P.; Montenegro-Cadena, L.; Blattner, B.; Menghwar, S.; Zurek, L.; Londono-Renteria, B. Differential Tick Salivary Protein Profiles and Human Immune Responses to Lone Star Ticks (Amblyomma americanum) from the Wild vs. a Laboratory Colony. Front. Immunol. 2019, 10, 1996. [Google Scholar] [CrossRef]
- Robertson, L.C.; Prior, S.; Apperson, C.S.; Irby, W.S. Bionomics of Anopheles quadrimaculatus and Culex erraticus (Diptera: Culicidae) in the Falls Lake basin, North Carolina: Seasonal changes in abundance and gonotrophic status, and host-feeding patterns. J. Med. Entomol. 1993, 30, 689–698. [Google Scholar] [CrossRef]
- Noblesse DeMoss, H. Biological and Taxonomic Observations on the Mosquitoes of Kansas. Trans. Kans. Acad. Sci. 1939, 42, 255–265. [Google Scholar]
- Dantur Juri, M.J.; Claps, G.L.; Santana, M.; Zaidenberg, M.; Almirón, W.R. Abundance patterns of Anopheles pseudopunctipennis and Anopheles argyritarsis in northwestern Argentina. Acta Trop. 2010, 115, 234–241. [Google Scholar] [CrossRef]
- Peng, Z.; Ho, M.K.; Li, C.; Simons, F.E. Evidence for natural desensitization to mosquito salivary allergens: Mosquito saliva specific IgE and IgG levels in children. Ann. Allergy Asthma Immunol. 2004, 93, 553–556. [Google Scholar] [CrossRef]
- Anker, M.; Arima, Y. Male-female differences in the number of reported incident dengue fever cases in six Asian countries. West. Pac. Surveill. Response J. 2011, 2, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, J.E.; Eliason, D.A.; Moore, M.; Sather, G.E.; Schonberger, L.B.; Cabrera-Coello, L.; Fernandez de Castro, J. Epidemiologic investigations of dengue infection in Mexico, 1980. Am. J. Epidemiol. 1983, 117, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Trravassos da Rosa, A.P.; Vasconcelos, P.F.; Travassos Da Rosa, E.S.; Rodrigues, S.G.; Mondet, B.; Cruz, A.C.; Sousa, M.R.; Travassos Da Rosa, J.F. Dengue epidemic in Belém, Pará, Brazil, 1996–1997. Emerg. Infect. Dis. 2000, 6, 298–301. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, J.M.; O’Donnell, J.S. Platelets in malaria pathogenesis. Blood 2018, 132, 1222–1224. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Abreu, C.; Harris, M.; Shrader, J.; Sarvepalli, S. Severe Thrombocytopenia Associated with Dengue Fever: An Evidence-Based Approach to Management of Thrombocytopenia. Case Rep. Hematol. 2022, 2022, 3358325. [Google Scholar] [CrossRef]
- de Azeredo, E.L.; Monteiro, R.Q.; de-Oliveira Pinto, L.M. Thrombocytopenia in Dengue: Interrelationship between Virus and the Imbalance between Coagulation and Fibrinolysis and Inflammatory Mediators. Mediat. Inflamm. 2015, 2015, 313842. [Google Scholar] [CrossRef] [PubMed]
- Ghanchi, N.K.; Khan, M.H.; Arain, M.A.; Zubairi, M.B.A.; Raheem, A.; Khan, M.A.; Beg, M.A. Hematological Profile and Gametocyte Carriage in Malaria Patients from Southern Pakistan. Cureus 2019, 11, e4256. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; van der Heijden, W.; Reuling, I.J.; Wan, J.; Yan, Q.; de Laat-Kremers, R.M.W.; Van der Ven, A.J.; de Groot, P.G.; McCall, M.; Sauerwein, R.W.; et al. Functional changes in hemostasis during asexual and sexual parasitemia in a controlled human malaria infection. PLoS ONE 2022, 17, e0271527. [Google Scholar] [CrossRef]
- Cox, D.; McConkey, S. The role of platelets in the pathogenesis of cerebral malaria. Cell. Mol. Life Sci. 2010, 67, 557–568. [Google Scholar] [CrossRef]
- Islam, A.; Emran, T.B.; Yamamoto, D.S.; Iyori, M.; Amelia, F.; Yusuf, Y.; Yamaguchi, R.; Alam, M.S.; Silveira, H.; Yoshida, S. Anopheline antiplatelet protein from mosquito saliva regulates blood feeding behavior. Sci. Rep. 2019, 9, 3129. [Google Scholar] [CrossRef]
- Champagne, D.E.; Smartt, C.T.; Ribeiro, J.M.; James, A.A. The salivary gland-specific apyrase of the mosquito Aedes aegypti is a member of the 5′-nucleotidase family. Proc. Natl. Acad. Sci. USA 1995, 92, 694–698. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).