Dairy Cow Health and Greenhouse Gas Emission Intensity



Abstract
1. Background
- Global consumption of animal protein will continue to increase because of population growth and taking into account per capita consumption of animal protein in many countries.
- Restricted resources (e.g., water, land, energy, capital) and environmental changes, including climate change, will drive complex agricultural decisions with impacts on research needs.
- Current and foreseeable rapid advances in basic biological sciences provide an unparalleled opportunity to maximize the yield of investments in animal science research and development.
2. Importance of Feed Intake in Conjunction with Animal Health
3. Animal Yield and Emissions
4. Influence of Productive Life and Fertility on the CF of Lactating Cows
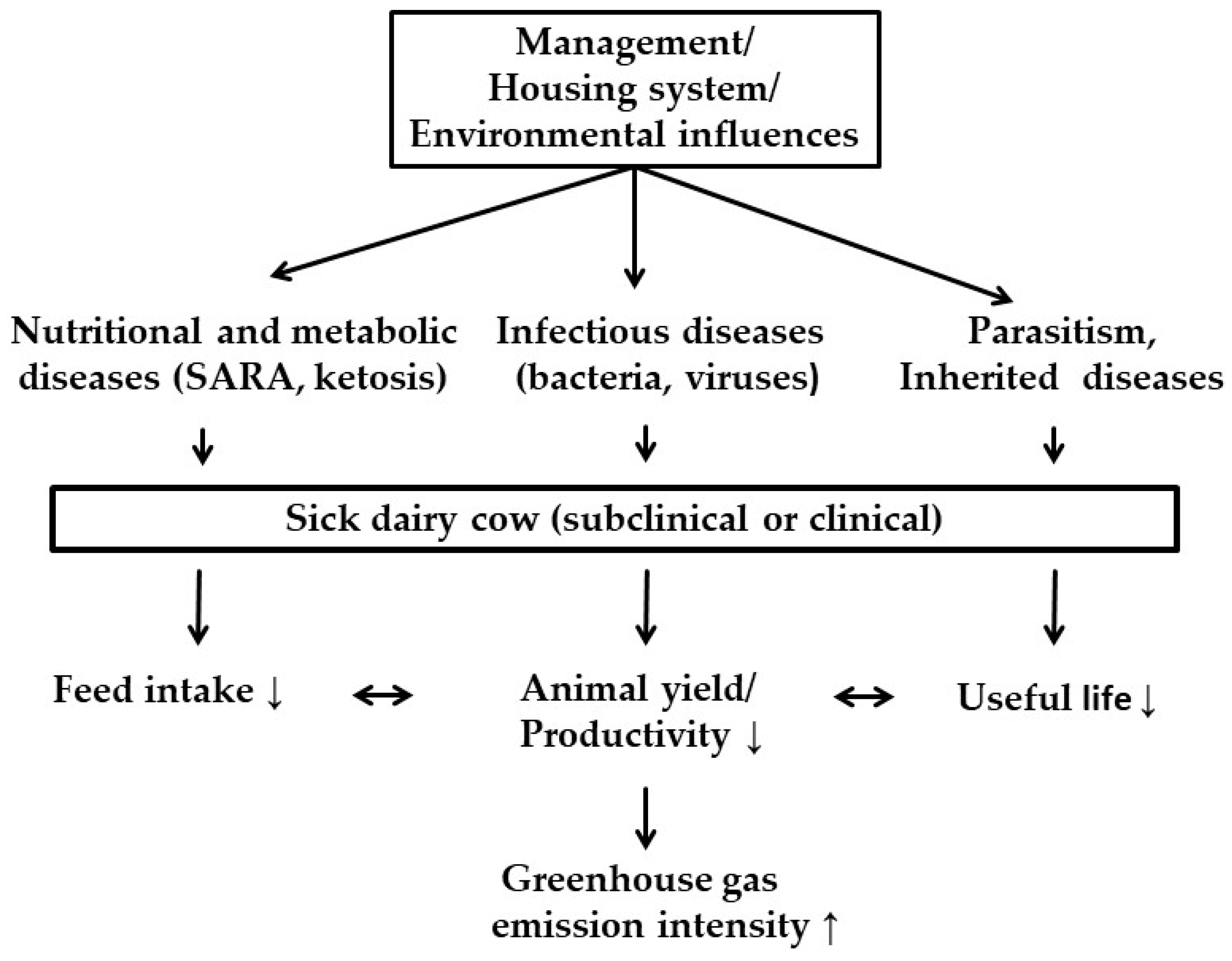
5. Dairy Cow Diseases and GHG Emissions
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Mottet, A.; de Haan, C.; Falcucci, A.; Tempio, G.; Opio, C.; Gerber, P. Livestock: On our plates or eating at our table? A new analysis of the feed/food debate. Glob. Food Secur. 2017, 14, 1–8. [Google Scholar] [CrossRef]
- Keyzer, M.A.; Merbis, M.D.; Pavel, I.F.P.W.; van Wesenbeeck, C.F.A. Diet shifts towards meat and the effects on cereal use: Can we feed the animals in 2030? Ecol. Econ. 2005, 55, 187–202. [Google Scholar] [CrossRef]
- Flachowsky, G.; Brade, W. Reduction potentials for methane emissions from ruminants. Züchtungskunde 2007, 79, 417–465. [Google Scholar]
- National Research Council of the National Academies (NRC). Critical Role of Animal Science Research in Food Security and Sustainability; The National Academies Press: Washington, DC, USA, 2015; p. 436. [Google Scholar]
- Souci, S.W.; Fachmann, W.; Kraut, H. Food Composition and Nutrition Tables, 8th ed.; Wissenschaftliche Verlagsgesellschaft: Stuttgart, Germany, 2016. [Google Scholar]
- Wennemer, H.; Flachowsky, G.; Hoffmann, V. Protein, Population, Politics—How Protein Can Be Supplied Sustainable in the 21st Century; Plexus Verlag: Milttenberg, Switzerland; Frankfurt/Main, Germany, 2006. [Google Scholar]
- Food and Agriculture Organization of the United Nations (FAO); World Health Organization (WHO) United Nations University (UNU). Protein and Amino Acid Requirements in Human Nutrition: Report of A Joint FAO/WHO/UNU Expert Consultation; World Health Organization: Geneva, Switzerland, 2007. [Google Scholar]
- Pillai, R.R.; Kurpad, A.V. Amino acid requirements: Quantitative estimates. In Amino Acids in Human Nutrition and Health; D’Mello, J.P.F., Ed.; CAB International: Wallingford, UK, 2011; pp. 267–290. [Google Scholar]
- Food and Agriculture Organisation of the United Nations (FAO). Improving Diets and Nutrition: Food-Based Approaches; Thompson, B., Amoroso, L., Eds.; CAB International and FAO: Rome, Italy, 2014. [Google Scholar]
- Flachowsky, G.; Kamphues, J. Carbon footprints for food of animal origin: What are the most preferable criteria to measure animal yields? Animals 2012, 2, 108–126. [Google Scholar] [CrossRef]
- Earth System Research Laboratory. Global Greenhouse Gas Reference Network: Trends in Atmospheric Carbon Dioxide. Available online: http://www.esrl.noaa.gov/gmd/ccgg/trends/?utm_source=www.uoota.com (accessed on 22 February 2019).
- Du Pont, Y.R.; Meinshausen, M. Warming assessment of the bottom-up paris agreement emissions pledges. Nat. Commun. 2018, 9, 4810. [Google Scholar] [CrossRef]
- Deckers, J. Should the consumption of farmed animal products be restricted, and if so, by how much? Food Policy 2010, 35, 497–503. [Google Scholar] [CrossRef]
- Godfray, H.C.; Beddington, J.R.; Crute, I.R.; Haddad, L.; Lawrence, D.; Muir, J.F.; Pretty, J.; Robinson, S.; Thomas, S.M.; Toulmin, C. Food security: The challenge of feeding 9 billion people. Science 2010, 327, 812–818. [Google Scholar] [CrossRef]
- Westhoek, H.; Lesschen, J.P.; Rood, T.; Wagner, S.; De Marco, A.; Murphy-Bokern, D.; Leip, A.; van Grinsven, H.; Sutton, M.A.; Oenema, O. Food choices, health and environment: Effects of cutting europe’s meat and dairy intake. Glob. Environ. Chang. Hum. Policy Dimens. 2014, 26, 196–205. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations (FAO). The Future of Food and Agriculture: Trends and Challenges; FAO: Rome, Italy, 2017. [Google Scholar]
- Eisler, M.C.; Lee, M.R.; Tarlton, J.F.; Martin, G.B.; Beddington, J.; Dungait, J.A.; Greathead, H.; Liu, J.; Mathew, S.; Miller, H.; et al. Agriculture: Steps to sustainable livestock. Nature 2014, 507, 32–34. [Google Scholar] [CrossRef]
- Schader, C.; Muller, A.; Scialabba Nel, H.; Hecht, J.; Isensee, A.; Erb, K.H.; Smith, P.; Makkar, H.P.; Klocke, P.; Leiber, F.; et al. Impacts of feeding less food-competing feedstuffs to livestock on global food system sustainability. J. R. Soc. Interface 2015, 12, 20150891. [Google Scholar] [CrossRef]
- Ertl, P.; Steinwidder, A.; Schönauer, M.; Krimberger, K.; Knaus, W.; Zollitsch, W. Net food production of different livestock: A national analysis for austria including relative occupation of different land categories. J. Land Manag. Food Environ. 2016, 67, 91. [Google Scholar]
- Flachowsky, G.; Meyer, U.; Südekum, K.H. Land use for edible protein of animal origin—A review. Animals 2017, 7, 25. [Google Scholar] [CrossRef] [PubMed]
- IPCC (Intergovernmental Panel on Climate Change). Guidelines for national greenhouse gas inventories; agriculture, forestry and other land use. In The National Greenhouse Gas Inventories Programme; Institute for Global Environmental Strategies (IGES): Hayama, Japan, 2006. [Google Scholar]
- Flachowsky, G. Carbon footprints of food of animal origin. In Livestock Production and Climate Change; Malik, P.K., Bhatta, R., Takahashi, J., Kohn, R.A., Prasad, C.S., Eds.; CAB International: Wallingford, UK, 2015; pp. 125–145. [Google Scholar]
- Gerber, P.J.; Steinfeld, H.; Henderson, B.; Mottet, A.; Opio, C.; Dijkman, J.; Falcucci, A.; Tempio, G. Tackling Climate Change through Livestock—A Global Assessment of Emissions and Mitigation Opportunities; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2013. [Google Scholar]
- Vigne, M.; Blanfort, V.; Vayssières, J.; Lecomte, P.; Steinmetz, P. Livestock farming constraints in developing countries—From adaptation to mitigation in ruminant production systems. In Climate Change and Agriculture Worldwide; Torquebiau, E., Ed.; Springer: Dordrecht, The Netherlands, 2016; pp. 127–141. [Google Scholar]
- Foley, J.A.; Ramankutty, N.; Brauman, K.A.; Cassidy, E.S.; Gerber, J.S.; Johnston, M.; Mueller, N.D.; O’Connell, C.; Ray, D.K.; West, P.C.; et al. Solutions for a cultivated planet. Nature 2011, 478, 337–342. [Google Scholar] [CrossRef]
- Powell, J.M.; MacLeod, M.; Vellinga, T.V.; Opio, C.; Falcucci, A.; Tempio, G.; Steinfeld, H.; Gerber, P. Feed-milk-manure nitrogen relationships in global dairy production systems. Livest. Sci. 2013, 152, 261–272. [Google Scholar] [CrossRef]
- Caro, D.; Kebreab, E.; Mitloehner, F.M. Mitigation of enteric methane emissions from global livestock systems through nutrition strategies. Clim. Chang. 2016, 137, 467–480. [Google Scholar] [CrossRef]
- Pandey, A.K.; Das, N.; Muthu, K.A.; Rao, S. Methanogens in the environment: An insight of methane yield and impact on global climate change. Int. Lett. Nat. Sci. 2015, 37, 51–60. [Google Scholar] [CrossRef]
- Hristov, A.N.; Ott, T.; Tricarico, J.; Rotz, A.; Waghorn, G.; Adesogan, A.; Dijkstra, J.; Montes, F.; Oh, J.; Kebreab, E.; et al. Special topics—Mitigation of methane and nitrous oxide emissions from animal operations: Iii. A review of animal management mitigation options. J. Anim. Sci. 2013, 91, 5095–5113. [Google Scholar] [CrossRef]
- Grace, D.; Bett, B.; Lindahl, J.; Robinson, T. Climate and Livestock Disease: Assessing the Vulnerability of Agricultural Systems to Livestock Pests under Climate Change Scenarios; CCAFS Working Paper; CGSpace: Copenhagen, Denmark, 2015; p. 29. [Google Scholar]
- Niemann, H.; Kuhla, B.; Flachowsky, G. Perspectives for feed-efficient animal production. J. Anim. Sci. 2011, 89, 4344–4363. [Google Scholar] [CrossRef]
- Nijdam, D.; Rood, T.; Westhoek, H. The price of protein: Review of land use and carbon footprints from life cycle assessments of animal food products and their substitutes. Food Policy 2012, 37, 760–770. [Google Scholar] [CrossRef]
- Carabano, M.J.; Logar, B.; Bormann, J.; Minet, J.; Vanrobays, M.L.; Diaz, C.; Tychon, B.; Gengler, N.; Hammami, H. Modeling heat stress under different environmental conditions. J. Dairy Sci. 2016, 99, 3798–3814. [Google Scholar] [CrossRef]
- Dahl, G.E.; Tao, S.; Monteiro, A.P.A. Effects of late-gestation heat stress on immunity and performance of calves. J. Dairy Sci. 2016, 99, 3193–3198. [Google Scholar] [CrossRef] [PubMed]
- Özkan, Ş.; Vitali, A.; Lacetera, N.; Amon, B.; Bannink, A.; Bartley, D.J.; Blanco-Penedo, I.; de Haas, Y.; Dufrasne, I.; Elliott, J.; et al. Challenges and priorities for modelling livestock health and pathogens in the context of climate change. Environ. Res. 2016, 151, 130–144. [Google Scholar] [CrossRef] [PubMed]
- Perano, K.M.; Usack, J.G.; Angenent, L.T.; Gebremedhin, K.G. Production and physiological responses of heat-stressed lactating dairy cattle to conductive cooling. J. Dairy Sci. 2015, 98, 5252–5261. [Google Scholar] [CrossRef]
- Polsky, L.; von Keyserlingk, M.A.G. Invited review: Effects of heat stress on dairy cattle welfare. J. Dairy Sci. 2017, 100, 8645–8657. [Google Scholar] [CrossRef]
- McIntyre, K.M.; Setzkorn, C.; Hepworth, P.J.; Morand, S.; Morse, A.P.; Baylis, M. Systematic assessment of the climate sensitivity of important human and domestic animals pathogens in europe. Sci. Rep. 2017, 7, 7134. [Google Scholar] [CrossRef] [PubMed]
- Global Research Alliance on Argriculture Greenhouse Gases (GRA). Animal Health and Greenhouse Gas Emission Intensity Network. Summary of Workshops 2013–2016. Available online: http://globalresearchalliance.org/wp-content/uploads/2017/05/Animal-Health-Summary-of-Workshops-2013-2016.pdf (accessed on 24 January 2019).
- GfE (Gesellschaft für Ernährungsphysiologie). Recommendations for Energy and Nutrient Requirements of Dairy Cattle and Heifers; DLG Verlag: Frankfurt am Main, Germany, 2001. [Google Scholar]
- Dämmgen, U.; Haenel, H.D. Emissions of greenhouse gases and gaseous air pollutants—A challenge for animal nutrition. In Proceedings of the Society of Nutrition Physiology, Göttingen, Germany, 1–4 April 2008; pp. 163–167. [Google Scholar]
- Herdt, T.H. Ruminant adaptation to negative energy balance: Influences on the etiology of ketosis and fatty liver. Vet. Clin. N. Am. Food Anim. Pract. 2000, 16, 215–230. [Google Scholar] [CrossRef]
- Kleen, J.L.; Cannizzo, C. Incidence, prevalence and impact of sara in dairy herds. Anim. Feed Sci. Technol. 2012, 172, 4–8. [Google Scholar] [CrossRef]
- Vanholder, T.; Papen, J.; Bemers, R.; Vertenten, G.; Berge, A.C. Risk factors for subclinical and clinical ketosis and association with production parameters in dairy cows in the netherlands. J. Dairy Sci. 2015, 98, 880–888. [Google Scholar] [CrossRef]
- Humer, E.; Petri, R.M.; Aschenbach, J.R.; Bradford, B.J.; Penner, G.B.; Tafaj, M.; Südekum, K.H.; Zebeli, Q. Invited review: Practical feeding management recommendations to mitigate the risk of subacute ruminal acidosis in dairy cattle. J. Dairy Sci. 2018, 101, 872–888. [Google Scholar] [CrossRef]
- Kleen, J.L.; Hooijer, G.A.; Rehage, J.; Noordhuizen, J.P. Subacute ruminal acidosis (sara): A review. J. Vet. Med. A Physiol. Pathol. Clin. Med. 2003, 50, 406–414. [Google Scholar] [CrossRef] [PubMed]
- Ertl, P.; Zebeli, Q.; Zollitsch, W.; Knaus, W. Feeding of by-products completely replaced cereals and pulses in dairy cows and enhanced edible feed conversion ratio. J. Dairy Sci. 2015, 98, 1225–1233. [Google Scholar] [CrossRef] [PubMed]
- Ertl, P.; Klocker, H.; Hortenhuber, S.; Knaus, W.; Zollitsch, W. The net contribution of dairy production to human food supply: The case of austrian dairy farms. Agric. Syst. 2015, 137, 119–125. [Google Scholar] [CrossRef]
- Zehetmeier, M.; Baudracco, J.; Hoffmann, H.; Heissenhuber, A. Does increasing milk yield per cow reduce greenhouse gas emissions? A system approach. Animal 2012, 6, 154–166. [Google Scholar] [CrossRef] [PubMed]
- Brade, W. Feed intake and efficiency of dairy cows: Opportunities and risks from the perspective of breeding and animal health. Tieraerztliche Umschau 2015, 70, 60. [Google Scholar]
- O’Brien, D.; Shalloo, L.; Grainger, C.; Buckley, F.; Horan, B.; Wallace, M. The influence of strain of holstein-friesian cow and feeding system on greenhouse gas emissions from pastoral dairy farms. J. Dairy Sci. 2010, 93, 3390–3402. [Google Scholar] [CrossRef] [PubMed]
- Bell, M.J.; Wall, E.; Russell, G.; Simm, G.; Stott, A.W. The effect of improving cow productivity, fertility, and longevity on the global warming potential of dairy systems. J. Dairy Sci. 2011, 94, 3662–3678. [Google Scholar] [CrossRef]
- Crosson, P.; Shalloo, L.; O’Brien, D.; Lanigan, G.J.; Foley, P.A.; Boland, T.M.; Kenny, D.A. A review of whole farm systems models of greenhouse gas emissions from beef and dairy cattle production systems. Anim. Feed Sci. Technol. 2011, 166–167, 29–45. [Google Scholar] [CrossRef]
- Wall, E.; Simm, G.; Moran, D. Developing breeding schemes to assist mitigation of greenhouse gas emissions. Animal 2010, 4, 366–376. [Google Scholar] [CrossRef]
- Garnsworthy, P.C. The environmental impact of fertility in dairy cows: A modelling approach to predict methane and ammonia emissions. Anim. Feed Sci. Technol. 2004, 112, 211–223. [Google Scholar] [CrossRef]
- LeBlanc, S.J.; Lissemore, K.D.; Kelton, D.F.; Duffield, T.F.; Leslie, K.E. Major advances in disease prevention in dairy cattle. J. Dairy Sci. 2006, 89, 1267–1279. [Google Scholar] [CrossRef]
- Dechow, C.D.; Goodling, R.C. Mortality, culling by sixty days in milk, and production profiles in high- and low-survival pennsylvania herds. J. Dairy Sci. 2008, 91, 4630–4639. [Google Scholar] [CrossRef] [PubMed]
- Dänicke, S.; Meyer, U.; Kersten, S.; Frahm, J. Animal models to study the impact of nutrition on the immune system of the transition cow. Res. Vet. Sci. 2018, 116, 15–27. [Google Scholar] [CrossRef]
- Mostert, P.F.; van Middelaar, C.E.; Bokkers, E.A.M.; de Boer, I.J.M. The impact of subclinical ketosis in dairy cows on greenhouse gas emissions of milk production. J. Clean. Prod. 2018, 171, 773–782. [Google Scholar] [CrossRef]
- Mostert, P.F.; van Middelaar, C.E.; de Boer, I.J.M.; Bokkers, E.A.M. The impact of foot lesions in dairy cows on greenhouse gas emissions of milk production. Agric. Syst. 2018, 167, 206–212. [Google Scholar] [CrossRef]
- Özkan, Ş.; Ahmadi, B.V.; Bonesmo, H.; Østerås, O.; Stott, A.; Harstad, O.M. Impact of animal health on greenhouse gas emissions. Adv. Anim. Biosci. 2015, 6, 24–25. [Google Scholar] [CrossRef]
- Özkan, Ş.; Ahmadi, B.V.; Stott, A.W. Impact of subclinical mastitis on greenhouse gas emissions intensity and profitability of dairy cows in norway. Prev. Vet. Med. 2018, 150, 19–29. [Google Scholar] [CrossRef]
- Salmon, G.R.; Marshall, K.; Tebug, S.F.; Missohou, A.; Robinson, T.P.; MacLeod, M. The greenhouse gas abatement potential of productivity improving measures applied to cattle systems in a developing region. Anim. Int. J. Anim. Biosci. 2017, 12, 844–852. [Google Scholar] [CrossRef]
- Houdijk, J.G.M.; Tolkamp, B.J.; Rooke, J.A.; Hutchings, M.R. Animal health and greenhouse gas intensity: The paradox of periparturient parasitism. Int. J. Parasitol. 2017, 47, 633–641. [Google Scholar] [CrossRef]
- McAloon, C.G.; Whyte, P.; More, S.J.; Green, M.J.; O’Grady, L.; Garcia, A.; Doherty, M.L. The effect of paratuberculosis on milk yield—A systematic review and meta-analysis. J. Dairy Sci. 2016, 99, 1449–1460. [Google Scholar] [CrossRef]


{kind=link}
| Protein Source | Body Weight (kg) | Performance | Edible Fraction (% of Product or of Body Mass) | Protein in Edible Fraction (g/kg) | Edible Protein (g/d) | Edible Protein (g/kg of BW) |
|---|---|---|---|---|---|---|
| Milk (kg/d) | ||||||
| Dairy cow | 650 | 10 | 95 | 34 | 323 | 0.7 |
| 20 | 646 | 1.0 | ||||
| 40 | 1292 | 2.0 | ||||
| Dairy goat | 60 | 2 | 95 | 36 | 68 | 1.2 |
| 5 | 170 | 2.8 | ||||
| Body weight gain (g/d) | ||||||
| Beef cattle | 350 | 500 | 50 | 190 | 48 | 0.14 |
| 1000 | 95 | 0.27 | ||||
| 1500 | 143 | 0.41 | ||||
| Growing/Fattening pig | 80 | 500 | 60 | 150 | 45 | 0.56 |
| 700 | 63 | 0.8 | ||||
| 1000 | 90 | 1.1 | ||||
| Broiler chicken | 1.5 | 40 | 60 | 200 | 4.8 | 3.2 |
| 60 | 7.2 | 4.8 | ||||
| 80 | 9.6 | 6.4 | ||||
| Laying performance (%) | ||||||
| Laying hens | 1.8 | 50 | 95 | 120 | 3.4 | 1.9 |
| 70 | 4.8 | 2.7 | ||||
| 90 | 6.2 | 3.4 |
| Protein Source | Studies | kg CO2-eq/kg Product | kg CO2-eq/kg Protein |
|---|---|---|---|
| Cow milk | 14 | 1–2 | 28–43 |
| Beef, intensive systems | 11 | 9–42 | 45–210 |
| Meadow, suckler herds | 8 | 23–52 | 114–250 |
| Extensive pastoral systems | 4 | 12–129 | 58–643 |
| Mutton and lamb | 5 | 10–150 | 51–750 |
| Pork | 11 | 4–11 | 20–55 |
| Poultry meat | 5 | 2–6 | 10–30 |
| Eggs | 5 | 2–6 | 15–42 |
| Seafood from fisheries | 18 | 1–86 | 4–540 |
| Seafood from agriculture | 11 | 3–15 | 4–75 |
| Parameter | Dry Matter Intake (kg/d) | ||||
|---|---|---|---|---|---|
| 10 | 15 | 20 | 25 | 30 | |
| Energy intake (MJ NEL/d) | 70 | 105 | 140 | 175 | 210 |
| Energy maintenance (% of total NEL†-intake) | 53.9 | 35.9 | 26.9 | 21.5 | 18.0 |
| Theoretical milk yield (3.3 MJ NEL/kg milk) | 9.8 | 20.4 | 31.0 | 41.6 | 52.2 |
| MJ NEL/kg of milk including energy for maintenance | 7.1 | 5.1 | 4.5 | 4.2 | 4.0 |
| Protein yield (g/cow and day) | 333 | 694 | 1054 | 1414 | 1775 |
| Methane emission♦ | |||||
| (g/d) | 240 | 360 | 480 | 600 | 720 |
| (g/kg milk) | 24.5 | 17.6 | 15.5 | 14.4 | 13.8 |
| Carbon footprint (CF)§ (g of CO2eq/kg of milk) | 825 | 605 | 530 | 495 | 475 |
| Protein Source | kg BW | Dry Matter Intake (kg/day) | Performance/Yield per Day | N-Excretion (% of intake) | Methane Emission; (g/d) | Emissions (kg/kg Edible Protein) | ||
|---|---|---|---|---|---|---|---|---|
| Milk (kg/d) | N | CH4 | CO2eq | |||||
| Dairy cow | 650 | 9 | 5 | 80 | 250 | 1.0 | 1.6 | 45 |
| 12 | 10 | 75 | 310 | 0.65 | 1.0 | 30 | ||
| 16 | 20 | 70 | 380 | 0.44 | 0.6 | 16 | ||
| 25 | 40 | 65 | 520 | 0.24 | 0.4 | 12 | ||
| Dairy goat | 60 | 2 | 2 | 75 | 50 | 0.5 | 0.8 | 20 |
| 2.5 | 5 | 65 | 60 | 0.2 | 0.4 | 10 | ||
| Body weight gain (g/d) | ||||||||
| Beef cattle | 350 | 6.5 | 500 | 90 | 170 | 2.3 | 3.5 | 110 |
| 7.0 | 1000 | 84 | 175 | 1.3 | 1.7 | 55 | ||
| 7.5 | 1500 | 80 | 180 | 1.0 | 1.2 | 33 | ||
| Growing/fattening pig | 80 | 1.8 | 500 | 85 | 5 | 1.0 | 0.12 | 16 |
| 2.0 | 700 | 80 | 5 | 0.7 | 0.08 | 12 | ||
| 2.2 | 900 | 75 | 5 | 0.55 | 0.05 | 10 | ||
| Broilers | 1.5 | 0.07 | 40 | 70 | Traces | 0.35 | 0.01 | 4 |
| 0.08 | 60 | 60 | 0.25 | 0.01 | 3 | |||
| Laying performance (%) | ||||||||
| Laying hens | 1.8 | 0.10 | 50 | 80 | Traces | 0.6 | 0.03 | 7 |
| 0.11 | 70 | 65 | 0.4 | 0.02 | 5 | |||
| 0.12 | 90 | 55 | 0.3 | 0.02 | 3 | |||
| Number of Lactations | 1 | 2 | 3 | 5 | 8 |
|---|---|---|---|---|---|
| kg CO2eq (CF1) per cow | 10,200 | 15,400 | 20,600 | 31,000 | 46,600 |
| g CO2eq (CF1) per kg milk | 1280 | 960 | 860 | 770 | 730 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
von Soosten, D.; Meyer, U.; Flachowsky, G.; Dänicke, S. Dairy Cow Health and Greenhouse Gas Emission Intensity. Dairy 2020, 1, 20-29. https://doi.org/10.3390/dairy1010003
von Soosten D, Meyer U, Flachowsky G, Dänicke S. Dairy Cow Health and Greenhouse Gas Emission Intensity. Dairy. 2020; 1(1):20-29. https://doi.org/10.3390/dairy1010003
Chicago/Turabian Stylevon Soosten, Dirk, Ulrich Meyer, Gerhard Flachowsky, and Sven Dänicke. 2020. "Dairy Cow Health and Greenhouse Gas Emission Intensity" Dairy 1, no. 1: 20-29. https://doi.org/10.3390/dairy1010003
APA Stylevon Soosten, D., Meyer, U., Flachowsky, G., & Dänicke, S. (2020). Dairy Cow Health and Greenhouse Gas Emission Intensity. Dairy, 1(1), 20-29. https://doi.org/10.3390/dairy1010003