
Noradrenergic Signaling in Astrocytes Influences Mammalian Sleep Homeostasis



{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
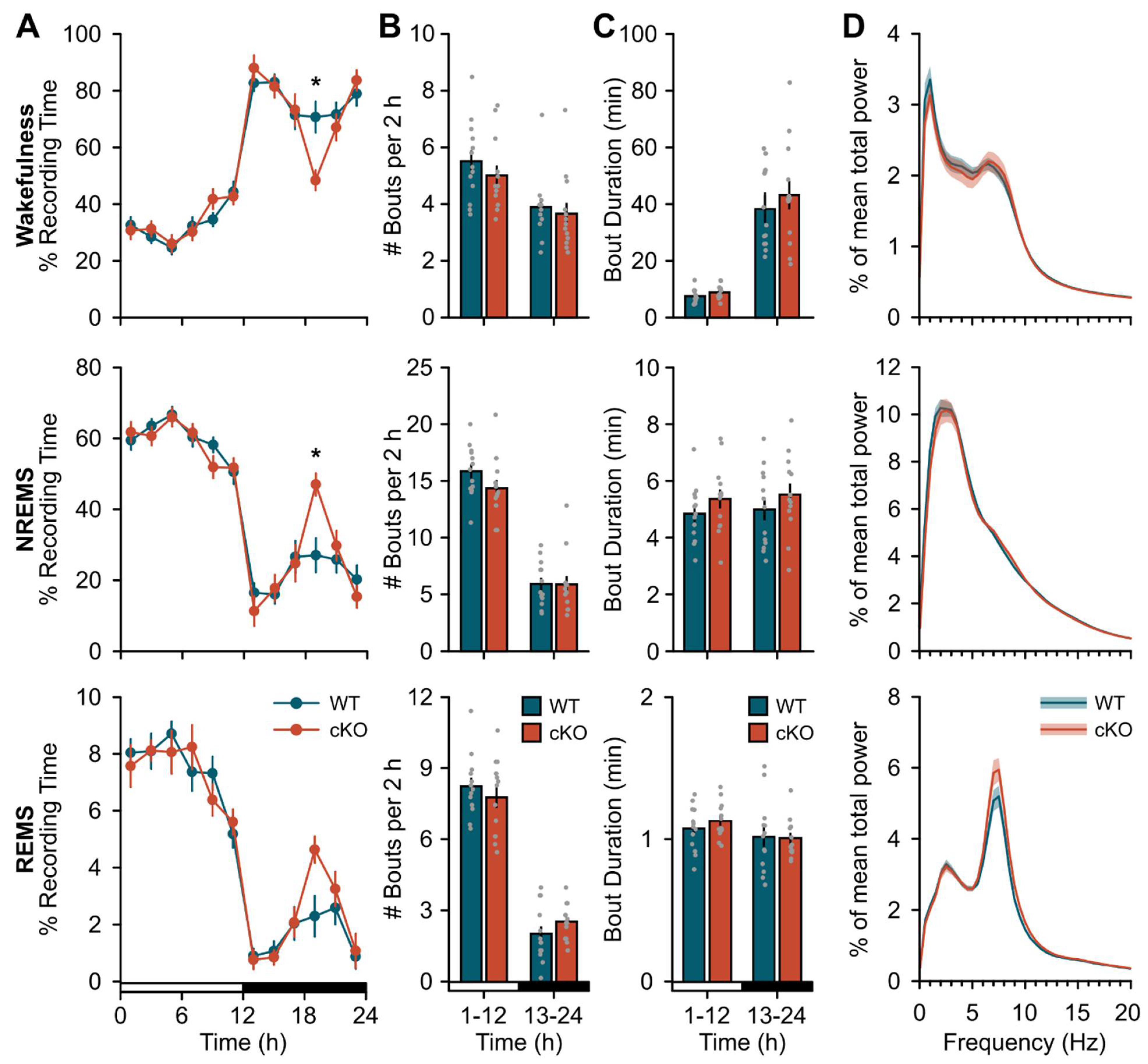
2.1. Baseline Changes in Sleep Expression Following cKO of Astroglial β2-ARs
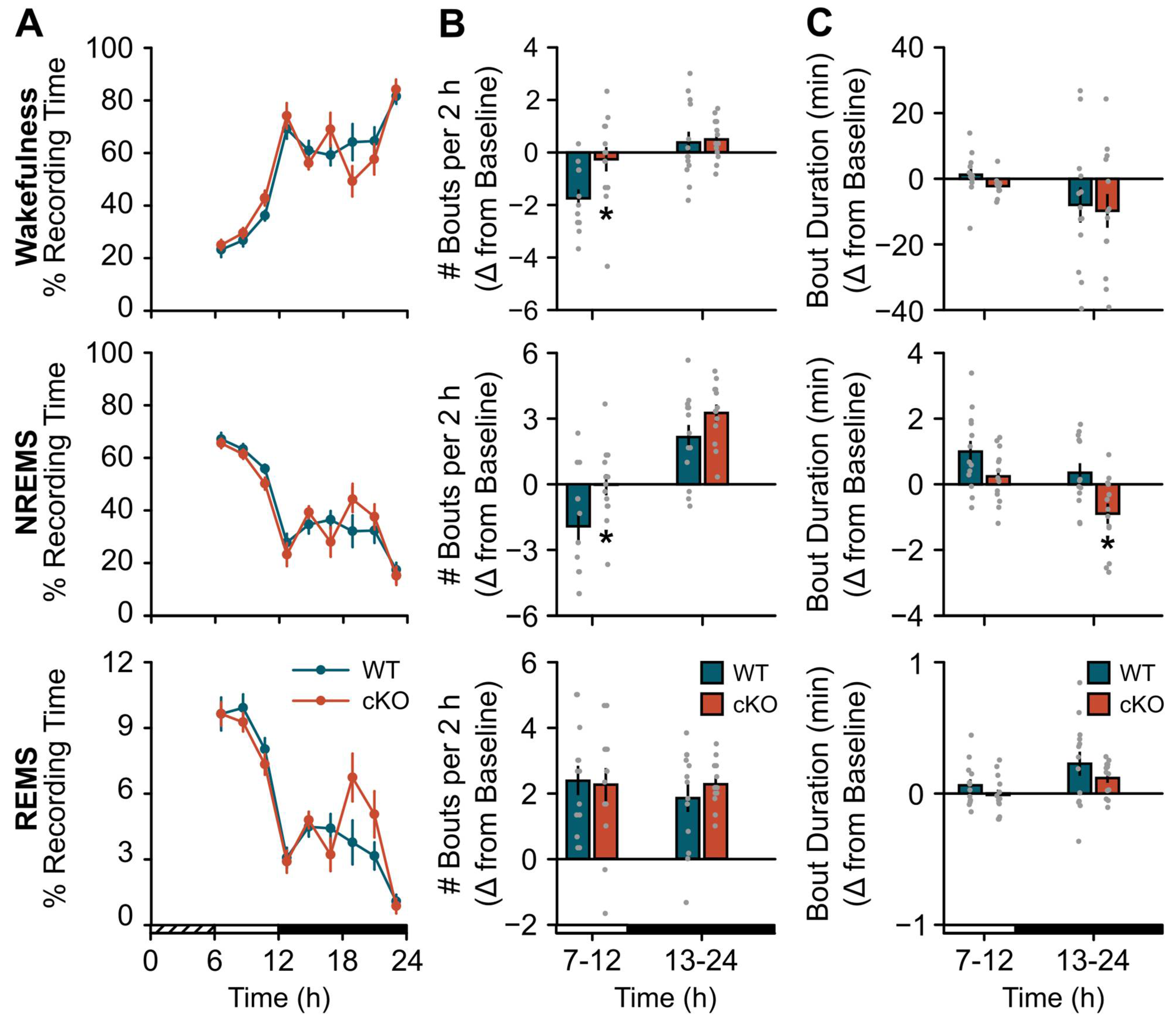
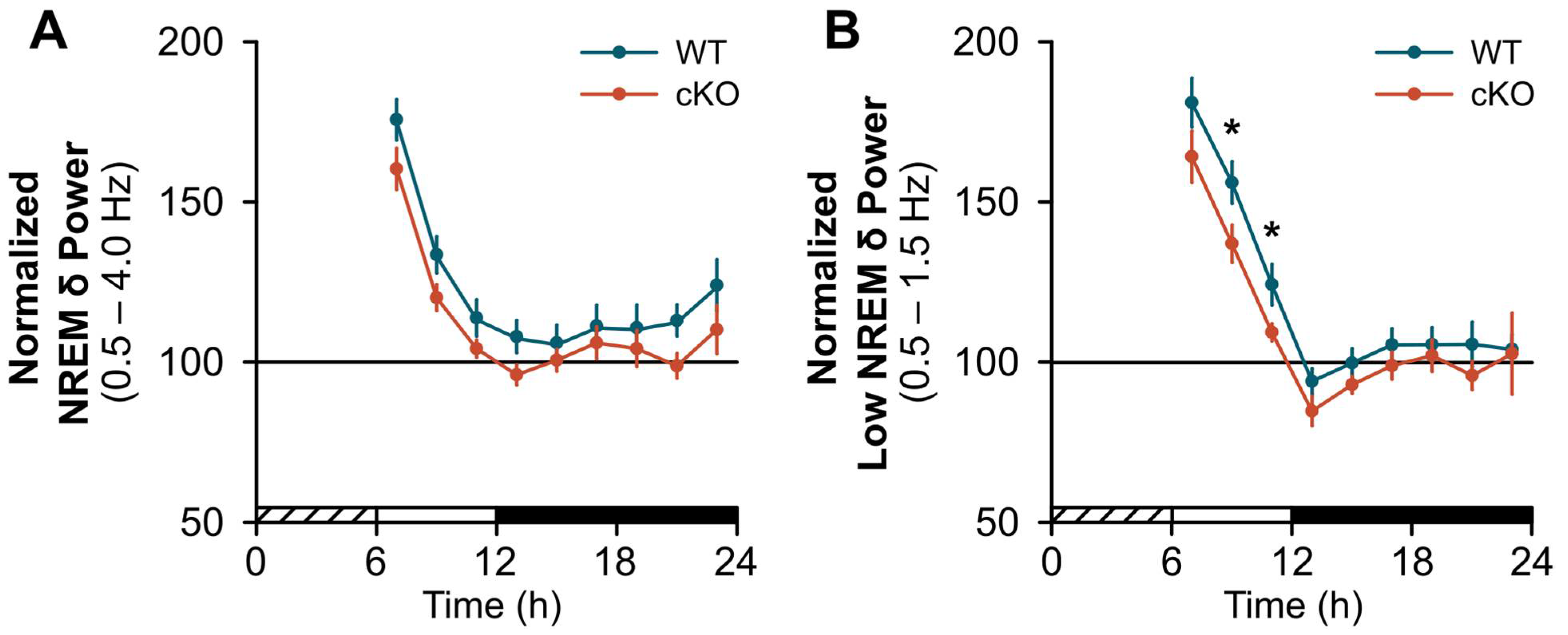
2.2. Changes in Sleep Homeostasis Following cKO of Astroglial β2-ARs
3. Discussion
3.1. Monoamines and Sleep Homeostasis
3.2. Astroglial Signaling and Sleep Homeostasis
3.3. Astroglial β2-AR Signaling and Sleep Homeostasis
3.4. Conclusions and Future Directions
4. Materials and Methods
4.1. Animals
4.2. Surgical Procedures
4.2.1. EEG and EMG Implantation
4.2.2. Cranial Window for Ca2+ Imaging
4.2.3. Telemeter Implantation for Core Body Temperature
4.3. Tamoxifen Injections
4.4. Experimental Procedures
4.4.1. Sleep Phenotyping
4.4.2. Ca2+ Response to Isoproterenol
4.4.3. Daily Core Body Temperature Patterns
4.5. Data Acquisition and Processing
4.5.1. EEG and EMG
4.5.2. Ca2+ Imaging
4.5.3. Core Body Temperature
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dijk, D.J.; Lockley, S.W. Integration of human sleep-wake regulation and circadian rhythmicity. J. Appl. Physiol. 2002, 92, 852–862. [Google Scholar] [CrossRef]
- Halassa, M.M.; Florian, C.; Fellin, T.; Munoz, J.R.; Lee, S.Y.; Abel, T.; Haydon, P.G.; Frank, M.G. Astrocytic modulation of sleep homeostasis and cognitive consequences of sleep loss. Neuron 2009, 61, 213–219. [Google Scholar] [CrossRef]
- Ingiosi, A.M.; Hayworth, C.R.; Harvey, D.O.; Singletary, K.G.; Rempe, M.J.; Wisor, J.P.; Frank, M.G. A role for astroglial calcium in mammalian sleep and sleep regulation. Curr. Biol. 2020, 30, 4373–4383.e4377. [Google Scholar] [CrossRef] [PubMed]
- Bojarskaite, L.; Bjørnstad, D.M.; Pettersen, K.H.; Cunen, C.; Hermansen, G.H.; Åbjørsbråten, K.S.; Chambers, A.R.; Sprengel, R.; Vervaeke, K.; Tang, W.; et al. Astrocytic Ca2+ signaling is reduced during sleep and is involved in the regulation of slow wave sleep. Nat. Commun. 2020, 11, 3240. [Google Scholar] [CrossRef] [PubMed]
- Vaidyanathan, T.V.; Collard, M.; Yokoyama, S.; Reitman, M.E.; Poskanzer, K.E. Cortical astrocytes independently regulate sleep depth and duration via separate gpcr pathways. eLife 2021, 10, e63329. [Google Scholar] [CrossRef] [PubMed]
- Tsunematsu, T.; Sakata, S.; Sanagi, T.; Tanaka, K.F.; Matsui, K. Region-specific and state-dependent astrocyte Ca2+ dynamics during the sleep-wake cycle in mice. J. Neurosci. 2021, 41, 5440–5452. [Google Scholar] [CrossRef] [PubMed]
- Blum, I.D.; Keles, M.F.; Baz, E.S.; Han, E.; Park, K.; Luu, S.; Issa, H.; Brown, M.; Ho, M.C.W.; Tabuchi, M.; et al. Astroglial calcium signaling encodes sleep need in drosophila. Curr. Biol. 2021, 31, 150–162 e157. [Google Scholar] [CrossRef]
- Ingiosi, A.M.; Frank, M.G. Goodnight, astrocyte: Waking up to astroglial mechanisms in sleep. FEBS J. 2022, in press. [CrossRef]
- Frank, M.G. Astroglial regulation of sleep homeostasis. Curr. Opin. Neurobiol. 2013, 23, 812–818. [Google Scholar] [CrossRef]
- Jones, B.E. The role of noradrenergic locus coeruleus neurons and neighboring cholinergic neurons of the pontomesencephalic tegmentum in sleep-wake states. Progress. Brain Res. 1991, 88, 533–543. [Google Scholar]
- Jones, B.E. Basic mechanisms of sleep-waking states. In Principles and Practice of Sleep Medicine, 4th ed.; Kryger, M.H., Roth, T., Dement, W.C., Eds.; Saunders: Philadelphia, PA, USA, 2005; pp. 136–153. [Google Scholar]
- Daniele, T.M.d.C.; de Bruin, P.F.C.; Rios, E.R.V.; de Bruin, V.M.S. Effects of exercise on depressive behavior and striatal levels of norepinephrine, serotonin and their metabolites in sleep-deprived mice. Behav. Brain Res. 2017, 332, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Bellesi, M.; Tononi, G.; Cirelli, C.; Serra, P.A. Region-specific dissociation between cortical noradrenaline levels and the sleep/wake cycle. Sleep 2016, 39, 143–154. [Google Scholar] [CrossRef]
- Bekar, L.K.; He, W.; Nedergaard, M. Locus coeruleus alpha-adrenergic-mediated activation of cortical astrocytes in vivo. Cereb. Cortex 2008, 18, 2789–2795. [Google Scholar] [CrossRef] [PubMed]
- Cirelli, C.; Huber, R.; Gopalakrishnan, A.; Southard, T.L.; Tononi, G. Locus ceruleus control of slow-wave homeostasis. J. Neurosci. 2005, 25, 4503–4511. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ding, F.; O’Donnell, J.; Thrane, A.S.; Zeppenfeld, D.; Kang, H.; Xie, L.; Wang, F.; Nedergaard, M. A1-adrenergic receptors mediate coordinated Ca2+ signaling of cortical astrocytes in awake, behaving mice. Cell Calcium 2013, 54, 387–394. [Google Scholar] [CrossRef]
- Chow, L.M.L.; Zhang, J.; Baker, S. Inducible cre recombinase activity in mouse mature astrocytes and adult neural precursor cells. Trangenic Res. 2008, 17, 919–928. [Google Scholar] [CrossRef]
- Hanada, R.; Leibbrandt, A.; Hanada, T.; Kitaoka, S.; Furuyashiki, T.; Fujihara, H.; Trichereau, J.; Paolino, M.; Qadri, F.; Plehm, R.; et al. Central control of fever and female body temperature by rankl/rank. Nature 2009, 462, 505–509. [Google Scholar] [CrossRef]
- Ganat, Y.M.; Silbereis, J.; Cave, C.; Ngu, H.; Anderson, G.M.; Ohkubo, Y.; Ment, L.R.; Vaccarino, F.M. Early postnatal astroglial cells produce multilineage precursors and neural stem cells in vivo. J. Neurosci. 2006, 26, 8609–8621. [Google Scholar] [CrossRef]
- Kim, J.G.; Suyama, S.; Koch, M.; Jin, S.; Argente-Arizon, P.; Argente, J.; Liu, Z.W.; Zimmer, M.R.; Jeong, J.K.; Szigeti-Buck, K.; et al. Leptin signaling in astrocytes regulates hypothalamic neuronal circuits and feeding. Nat. Neurosci. 2014, 17, 908–910. [Google Scholar] [CrossRef]
- García-Cáceres, C.; Quarta, C.; Varela, L.; Gao, Y.; Gruber, T.; Legutko, B.; Jastroch, M.; Johansson, P.; Ninkovic, J.; Yi, C.X.; et al. Astrocytic insulin signaling couples brain glucose uptake with nutrient availability. Cell 2016, 166, 867–880. [Google Scholar] [CrossRef]
- Franco, C.; Genis, L.; Navarro, J.A.; Perez-Domper, P.; Fernandez, A.M.; Schneuwly, S.; Torres Alemán, I. A role for astrocytes in cerebellar deficits in frataxin deficiency: Protection by insulin-like growth factor i. Mol. Cell. Neurosci. 2017, 80, 100–110. [Google Scholar] [CrossRef] [PubMed]
- Koeppen, J.; Nguyen, A.Q.; Nikolakopoulou, A.M.; Garcia, M.; Hanna, S.; Woodruff, S.; Figueroa, Z.; Obenaus, A.; Ethell, I.M. Functional consequences of synapse remodeling following astrocyte-specific regulation of ephrin-b1 in the adult hippocampus. J. Neurosci. 2018, 38, 5710–5726. [Google Scholar] [CrossRef]
- Jin, S.; Kim, K.K.; Park, B.S.; Kim, D.H.; Jeong, B.; Kang, D.; Lee, T.H.; Park, J.W.; Kim, J.G.; Lee, B.J. Function of astrocyte myd88 in high-fat-diet-induced hypothalamic inflammation. J. Neuroinflam. 2020, 17, 195. [Google Scholar] [CrossRef] [PubMed]
- Gao, V.; Suzuki, A.; Magistretti, P.J.; Lengacher, S.; Pollonini, G.; Steinman, M.Q.; Alberini, C.M. Astrocytic β2-adrenergic receptors mediate hippocampal long-term memory consolidation. Proc. Natl. Acad. Sci. USA 2016, 113, 201605063. [Google Scholar] [CrossRef]
- Hofstetter, J.R.; Svihla-Jones, D.A.; Mayeda, A.R. A QTL on mouse chromosome 12 for the genetic variance in free-running circadian period between inbred strains of mice. J. Circadian Rhythm. 2007, 5, 7. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Franken, P.; Malafosse, A.; Tafti, M. Genetic determinants of sleep regulation in inbred mice. Sleep 1999, 22, 155–169. [Google Scholar]
- Franken, P.; Lopez-Molina, L.; Marcacci, L.; Schibler, U.; Tafti, M. The transcription factor dbp affects circadian sleep consolidation and rhythmic eeg activity. J. Neurosci. 2000, 20, 617–625. [Google Scholar] [CrossRef]
- Jouvet, M. Sleep and serotonin: An unfinished story. Neuropsychopharmacology 1999, 21, 24s–27s. [Google Scholar]
- Greene, R.W.; Frank, M.G. Slow wave activity during sleep: Functional and therapeutic implications. Neuroscientist 2010, 17, 618–633. [Google Scholar] [CrossRef]
- Sallanon, M.; Janin, M.; Buda, C.; Jouvet, M. Serotonergic mechanisms and sleep rebound. Brain Res. 1983, 268, 95–104. [Google Scholar] [CrossRef]
- Tobler, I.; Borbely, A.A. Sleep regulation after reduction of brain serotonin: Effect of p-chlorophenylalanine combined with sleep deprivation in the rat. Sleep 1982, 5, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Gerstner, J.R.; Koberstein, J.N.; Watson, A.J.; Zapero, N.; Risso, D.; Speed, T.P.; Frank, M.G.; Peixoto, L. Removal of unwanted variation reveals novel patterns of gene expression linked to sleep homeostasis in murine cortex. BMC Genom. 2016, 17, 727. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, M.; Hellman, K.; Abel, T.; Thomas, S.A. Adrenergic signaling plays a critical role in the maintenance of waking and in the regulation of rem sleep. J. Neurophysiol. 2004, 92, 2071–2082. [Google Scholar] [CrossRef]
- Mallick, B.N.; Alam, M.N. Different types of norepinephrinergic receptors are involved in preoptic area mediated independent modulation of sleep-wakefulness and body temperature. Brain Res. 1992, 591, 8–19. [Google Scholar] [CrossRef]
- Catus, S.L.; Gibbs, M.E.; Sato, M.; Summers, R.J.; Hutchinson, D.S. Role of β-adrenoceptors in glucose uptake in astrocytes using β-adrenoceptor knockout mice. Br. J. Pharmacol. 2011, 162, 1700–1715. [Google Scholar] [CrossRef]
- Hertz, L.; Lovatt, D.; Goldman, S.A.; Nedergaard, M. Adrenoceptors in brain: Cellular gene expression and effects on astrocytic metabolism and [Ca(2+)]i. Neurochem. Int. 2010, 57, 411–420. [Google Scholar] [CrossRef] [PubMed]
- Oe, Y.; Wang, X.; Patriarchi, T.; Konno, A.; Ozawa, K.; Yahagi, K.; Hirai, H.; Tsuboi, T.; Kitaguchi, T.; Tian, L.; et al. Distinct temporal integration of noradrenaline signaling by astrocytic second messengers during vigilance. Nat. Commun. 2020, 11, 471. [Google Scholar] [CrossRef]
- Wahis, J.; Holt, M.G. Astrocytes, noradrenaline, α1-adrenoreceptors, and neuromodulation: Evidence and unanswered questions. Front. Cell. Neurosci. 2021, 15, 645691. [Google Scholar] [CrossRef]
- Scharf, M.T.; Naidoo, N.; Zimmerman, J.E.; Pack, A.I. The energy hypothesis of sleep revisited. Prog. Neurobiol. 2008, 86, 264–280. [Google Scholar] [CrossRef]
- Petit, J.-M.; Burlet-Godinot, S.; Magistretti, P.J.; Allaman, I. Glycogen metabolism and the homeostatic regulation of sleep. Metab. Brain Dis. 2015, 30, 263–279. [Google Scholar] [CrossRef]
- Benington, J.; Heller, H.C. Restoration of brain energy metabolism as the function of sleep. Prog. Neurobiol. 1995, 45, 347–360. [Google Scholar] [CrossRef]
- Dong, J.-H.; Chen, X.; Cui, M.; Yu, X.; Pang, Q.; Sun, J.-P. Beta2-adrenergic receptor and astrocyte glucose metabolism. J. Mol. Neurosci. 2012, 48, 456–463. [Google Scholar] [CrossRef] [PubMed]
- Pellerin, L.; Magistretti, P.J. Sweet sixteen for anls. J. Cereb. Blood Flow Metab. 2012, 32, 1152–1166. [Google Scholar] [CrossRef]
- Wisor, J.P.; Rempe, M.J.; Schmidt, M.A.; Moore, M.E.; Clegern, W.C. Sleep slow-wave activity regulates cerebral glycolytic metabolism. Cereb. Cortex 2013, 23, 1978–1987. [Google Scholar] [CrossRef] [PubMed]
- Dash, M.B.; Tononi, G.; Cirelli, C. Extracellular levels of lactate, but not oxygen, reflect sleep homeostasis in the rat cerebral cortex. Sleep 2012, 35, 909–919. [Google Scholar] [CrossRef] [PubMed]
- Naylor, E.; Aillon, D.V.; Barrett, B.S.; Wilson, G.S.; Johnson, D.A.; Johnson, D.A.; Harmon, H.P.; Gabbert, S.; Petillo, P.A. Lactate as a biomarker for sleep. Sleep 2012, 35, 1209–1222. [Google Scholar] [CrossRef] [PubMed]
- Petit, J.M.; Gyger, J.; Burlet-Godinot, S.; Fiumelli, H.; Martin, J.L.; Magistretti, P.J. Genes involved in the astrocyte-neuron lactate shuttle (anls) are specifically regulated in cortical astrocytes following sleep deprivation in mice. Sleep 2013, 36, 1445–1458. [Google Scholar] [CrossRef]
- Clasadonte, J.; Scemes, E.; Wang, Z.; Boison, D.; Haydon, P.G. Connexin 43-mediated astroglial metabolic networks contribute to the regulation of the sleep-wake cycle. Neuron 2017, 95, 1365–1380 e1365. [Google Scholar] [CrossRef]
- Bazargani, N.; Attwell, D. Amines, astrocytes, and arousal. Neuron 2017, 94, 228–231. [Google Scholar] [CrossRef]
- Cash, R.; Raisman, R.; Lanfumey, L.; Ploska, A.; Agid, Y. Cellular localization of adrenergic receptors in rat and human brain. Brain Res. 1986, 370, 127–135. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, K.; Sloan, S.A.; Bennett, M.L.; Scholze, A.R.; O’Keeffe, S.; Phatnani, H.P.; Guarnieri, P.; Caneda, C.; Ruderisch, N.; et al. An rna-sequencing transcriptome and splicing database of glia, neurons, and vascular cells of the cerebral cortex. J. Neurosci. 2014, 34, 11929–11947. [Google Scholar] [CrossRef] [PubMed]
- Ramos, B.P.; Arnsten, A.F. Adrenergic pharmacology and cognition: Focus on the prefrontal cortex. Pharmacol. Ther. 2007, 113, 523–536. [Google Scholar] [CrossRef] [PubMed]
- Mulvey, B.; Bhatti, D.L.; Gyawali, S.; Lake, A.M.; Kriaucionis, S.; Ford, C.P.; Bruchas, M.R.; Heintz, N.; Dougherty, J.D. Molecular and functional sex differences of noradrenergic neurons in the mouse locus coeruleus. Cell Rep. 2018, 23, 2225–2235. [Google Scholar] [CrossRef]
- Sun, P.; Wang, J.; Zhang, M.; Duan, X.; Wei, Y.; Xu, F.; Ma, Y.; Zhang, Y.-H. Sex-related differential whole-brain input atlas of locus coeruleus noradrenaline neurons. Front. Neural Circuits 2020, 14, 53. [Google Scholar] [CrossRef] [PubMed]
- Ehlen, J.C.; Jones, K.A.; Pinckney, L.; Gray, C.L.; Burette, S.; Weinberg, R.J.; Evans, J.A.; Brager, A.J.; Zylka, M.J.; Paul, K.N.; et al. Maternal ube3a loss disrupts sleep homeostasis but leaves circadian rhythmicity largely intact. J. Neurosci. 2015, 35, 13587–13598. [Google Scholar] [CrossRef]
- Brancaccio, M.; Patton, A.P.; Chesham, J.E.; Maywood, E.S.; Hastings, M.H. Astrocytes control circadian timekeeping in the suprachiasmatic nucleus via glutamatergic signaling. Neuron 2017, 93, 1420–1435 e1425. [Google Scholar] [CrossRef]
- Brancaccio, M.; Edwards, M.D.; Patton, A.P.; Smyllie, N.J.; Chesham, J.E.; Maywood, E.S.; Hastings, M.H. Cell-autonomous clock of astrocytes drives circadian behavior in mammals. Science 2019, 363, 187–192. [Google Scholar] [CrossRef]
- Astiz, M.; Delgado-García, L.M.; López-Mascaraque, L. Astrocytes as essential time-keepers of the central pacemaker. Glia 2021, 70, 808–819. [Google Scholar] [CrossRef]
- Hinoi, E.; Gao, N.; Jung, D.Y.; Yadav, V.; Yoshizawa, T.; Myers, M.G., Jr.; Chua, S.C., Jr.; Kim, J.K.; Kaestner, K.H.; Karsenty, G. The sympathetic tone mediates leptin’s inhibition of insulin secretion by modulating osteocalcin bioactivity. J. Cell Biol. 2008, 183, 1235–1242. [Google Scholar] [CrossRef]
- Ingiosi, A.M.; Schoch, H.; Wintler, T.; Singletary, K.G.; Righelli, D.; Roser, L.G.; Medina, E.; Risso, D.; Frank, M.G.; Peixoto, L. Shank3 modulates sleep and expression of circadian transcription factors. eLife 2019, 8, e42819. [Google Scholar] [CrossRef]
- Frank, M.G.; Stryker, M.P.; Tecott, L.H. Sleep and sleep homeostasis in mice lacking the 5-ht2c receptor. Neuropsychopharmacology 2002, 27, 869–873. [Google Scholar] [CrossRef]
- Srinivasan, R.; Huang, B.S.; Venugopal, S.; Johnston, A.D.; Chai, H.; Zeng, H.; Golshani, P.; Khakh, B.S. Ca2+ signaling in astrocytes from ip3r2−/− mice in brain slices and during startle responses in vivo. Nat. Neurosci. 2015, 18, 708–717. [Google Scholar] [CrossRef] [PubMed]
- Heuser, K.; Nome, C.G.; Pettersen, K.H.; Åbjørsbråten, K.S.; Jensen, V.; Tang, W.; Sprengel, R.; Taubøll, E.; Nagelhus, E.A.; Enger, R. Ca2+ signals in astrocytes facilitate spread of epileptiform activity. Cereb. Cortex 2018, 28, 4036–4048. [Google Scholar] [CrossRef] [PubMed]
- Bjorness, T.E.; Dale, N.; Mettlach, G.; Sonneborn, A.; Sahin, B.; Fienberg, A.A.; Yanagisawa, M.; Bibb, J.A.; Greene, R.W. An adenosine-mediated glial-neuronal circuit for homeostatic sleep. J. Neurosci. Off. J. Soc. Neurosci. 2016, 36, 3709–3721. [Google Scholar] [CrossRef]
- Ingiosi, A.M.; Raymond, R.M., Jr.; Pavlova, M.N.; Opp, M.R. Selective contributions of neuronal and astroglial interleukin-1 receptor 1 to the regulation of sleep. Brain. Behav. Immun. 2015, 48, 244–257. [Google Scholar] [CrossRef]
- Gonzalez, J.P.; Ramachandran, J.; Xie, L.H.; Contreras, J.E.; Fraidenraich, D. Selective connexin43 inhibition prevents isoproterenol-induced arrhythmias and lethality in muscular dystrophy mice. Sci. Rep. 2015, 5, 13490. [Google Scholar] [CrossRef]
- Ji, S.; Guo, R.; Wang, J.; Qian, L.; Liu, M.; Xu, H.; Zhang, J.; Guan, Y.; Yang, G.; Chen, L. Mpges-1 deletion attenuates isoproterenol-induced myocardial fibrosis in mice. J. Pharmacol. Exp. Ther. 2020, 375, JPET-AR-2020-000023. [Google Scholar] [CrossRef]
- Ingiosi, A.M.; Opp, M.R. Sleep and immunomodulatory responses to systemic lipopolysaccharide in mice selectively expressing interleukin-1 receptor 1 on neurons or astrocytes. Glia 2016, 64, 780–791. [Google Scholar] [CrossRef]
- Franken, P.; Tobler, I.; Borbely, A.A. Sleep homeostasis in the rat: Simulation of the time course of eeg slow-wave activity. Neurosci. Lett. 1991, 130, 141–144. [Google Scholar] [CrossRef]
- Gulati, S.; Cao, V.Y.; Otte, S. Multi-layer cortical Ca2+ imaging in freely moving mice with prism probes and miniaturized fluorescence microscopy. J. Vis. Exp. 2017, e55579. [Google Scholar]
- Weber, F.; Hoang Do, J.P.; Chung, S.; Beier, K.T.; Bikov, M.; Saffari Doost, M.; Dan, Y. Regulation of rem and non-rem sleep by periaqueductal gabaergic neurons. Nat. Commun. 2018, 9, 354. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, J.C.; Su, K.; Goldberg, A.R.; Luna, V.M.; Biane, J.S.; Ordek, G.; Zhou, P.; Ong, S.K.; Wright, M.A.; Zweifel, L.; et al. Anxiety cells in a hippocampal-hypothalamic circuit. Neuron 2018, 97, 670–683.e676. [Google Scholar] [CrossRef] [PubMed]
- Kirschen, G.W.; Shen, J.; Tian, M.; Schroeder, B.; Wang, J.; Man, G.; Wu, S.; Ge, S. Active dentate granule cells encode experience to promote the addition of adult-born hippocampal neurons. J. Neurosci. 2017, 37, 4661–4678. [Google Scholar] [CrossRef] [PubMed]
- Resendez, S.L.; Jennings, J.H.; Ung, R.L.; Namboodiri, V.M.; Zhou, Z.C.; Otis, J.M.; Nomura, H.; McHenry, J.A.; Kosyk, O.; Stuber, G.D. Visualization of cortical, subcortical and deep brain neural circuit dynamics during naturalistic mammalian behavior with head-mounted microscopes and chronically implanted lenses. Nat. Protoc. 2016, 11, 566–597. [Google Scholar] [CrossRef] [PubMed]
- Eban-Rothschild, A.; Rothschild, G.; Giardino, W.J.; Jones, J.R.; de Lecea, L. Vta dopaminergic neurons regulate ethologically relevant sleep-wake behaviors. Nat. Neurosci. 2016, 19, 1356–1366. [Google Scholar] [CrossRef] [PubMed]
- Paukert, M.; Agarwal, A.; Cha, J.; Doze, V.A.; Kang, J.U.; Bergles, D.E. Norepinephrine controls astroglial responsiveness to local circuit activity. Neuron 2014, 82, 1263–1270. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ingiosi, A.M.; Frank, M.G. Noradrenergic Signaling in Astrocytes Influences Mammalian Sleep Homeostasis. Clocks & Sleep 2022, 4, 332-345. https://doi.org/10.3390/clockssleep4030028
Ingiosi AM, Frank MG. Noradrenergic Signaling in Astrocytes Influences Mammalian Sleep Homeostasis. Clocks & Sleep. 2022; 4(3):332-345. https://doi.org/10.3390/clockssleep4030028
Chicago/Turabian StyleIngiosi, Ashley M., and Marcos G. Frank. 2022. "Noradrenergic Signaling in Astrocytes Influences Mammalian Sleep Homeostasis" Clocks & Sleep 4, no. 3: 332-345. https://doi.org/10.3390/clockssleep4030028
APA StyleIngiosi, A. M., & Frank, M. G. (2022). Noradrenergic Signaling in Astrocytes Influences Mammalian Sleep Homeostasis. Clocks & Sleep, 4(3), 332-345. https://doi.org/10.3390/clockssleep4030028