
Taphonomy as a Methodological Approach for the Study of Dog Domestication: Application to the Prehistoric Site of Peña Moñuz (Guadalajara, Spain)

 ,
,
Abstract
1. Introduction
- Many prehistoric sites with well-preserved faunal remains feature numerous bones bearing tooth marks. If dogs were present at these sites, it is plausible that they created some of these marks. Identifying the origin of such marks could provide indirect evidence of the actions of dogs in these contexts.
- Testing the hypothesis outlined above would be challenging without a reliable method to classify tooth marks. However, previous research has demonstrated that tooth marks made by different predators can be classified with high accuracy [42,43,44,45,46]. Moreover, studies suggest that tooth pits created by different individuals of the same carnivorous species often exhibit consistent patterns. For example, observations of wolves from distinct populations—such as Flechas and Villardeciervos in Zamora, Cabárceno in Cantabria, and Hosquillo in Cuenca—show that both captive and wild individuals produce similar tooth pits [47]. Similarly, research on other carnivores, including leopards and tigers, indicates that individuals of the same species, even with significant size differences due to sexual dimorphism, leave comparable tooth marks. For instance, male and female leopards, as well as tigers, produce similar patterns of tooth pits [48].
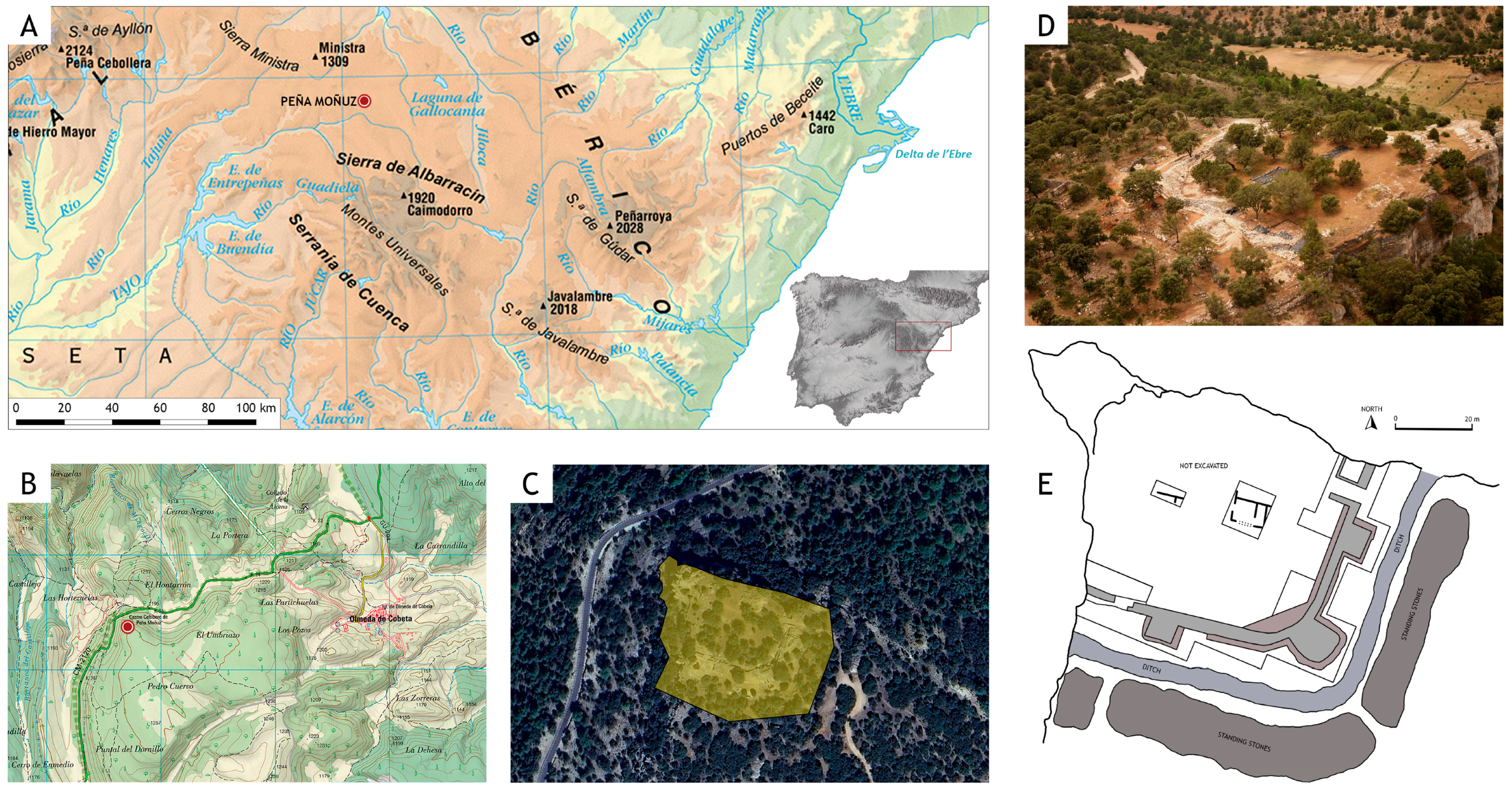
2. The Site of Peña Moñuz
3. Materials and Methods
- Carnivores were provided with whole horse bones (typically radius and tibiae).
- The bones were semi-defleshed before being offered to the animals after feeding.
- The bones were left in enclosures for several days.
- The bones were subsequently collected and boiled in water to preserve tooth marks, avoiding chemical cleaning agents that could degrade them. After boiling and drying, the bones were ready for examination.
- Pits on each bone were identified and located in preparation for further comparative study in the subsequent stages of the research.
4. Results and Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Benecke, N. Studies on early dog remains from Northern Europe. J. Archaeol. Sci. 1987, 14, 31–49. [Google Scholar] [CrossRef]
- Linares-Matás, G.J. La Domesticación del Perro y sus Orígenes. Rev. Soc. Estud. Hist. Etnográficos 2019, 13, 42–49. [Google Scholar]
- Vigne, J.D. L’humérus de chien magdalénien de Erralla (Gipuzkoa, Espagne) et la domestication tardiglaciaire du loup en Europe. Munibe 2005, 57, 279–287. [Google Scholar]
- Germonpré, M.; Sablin, M.V.; Stevens, R.E.; Hedges, R.E.M.; Hofreiter, M.; Stiller, M.; Després, V.R. Fossil dogs and wolves from Palaeolithic sites in Belgium, the Ukraine and Russia: Osteometry, ancient DNA and stable isotopes. J. Archaeol. Sci. 2009, 36, 473–490. [Google Scholar] [CrossRef]
- Shipman, P. The Invaders: How Humans and Their Dogs Drove Neanderthals of Extinction, 1st ed.; The Belknap Press of Harvard University Press: Cambridge, MA, USA; London, UK, 2015. [Google Scholar]
- Wang, X.; Wei, W.; Kang, N.; Zhang, N.; Tang, Z.; Chen, Q.; Wang, L.; Zhang, W.; Tian, H.; Liu, W.; et al. Ancient DNA reveals the origin and history of early dogs in northeastern China. J. Archaeol. Sci. 2024, 168, 106010. [Google Scholar] [CrossRef]
- Janssens, L.; Miller, R.; Van Dongen, S. The morphology of the mandibular coronoid process does not indicate that Canis lupus chanco is the progenitor to dogs. Zoomorphology 2016, 135, 269–277. [Google Scholar] [CrossRef]
- Sablin, M.; Khlopachev, G. The Earliest Ice Age Dogs: Evidence from Eliseevichi 1. Curr. Anthropol. 2002, 43, 795–799. [Google Scholar] [CrossRef]
- Ovodov, N.D.; Crockford, S.J.; Kuzmin, Y.V.; Higham, T.F.G.; Hodgins, G.W.L.; Van der Plicht, J. A 33,000-Year-Old Incipient Dog from the Altai Mountains of Siberia: Evidence of the Earliest Domestication Disrupted by the Last Glacial Maximum. PLoS ONE 2011, 6, e22821. [Google Scholar] [CrossRef]
- Germonpré, M.; Lázničková-Galetová, M.; Sablin, M. Palaeolithic dog skulls at the Gravettian Předmostí site, the Czech Republic. J. Archaeol. Sci. 2012, 39, 184–202. [Google Scholar] [CrossRef]
- Germonpré, M.; Lázničková-Galetová, M.; Losey, R.J.; Räikkönen, J.; Sablin, M.V. Large canids at the Gravettian Předmostí site, the Czech Republic: The mandible. Quat. Int. 2015, 359–360, 261–279. [Google Scholar] [CrossRef]
- Germonpré, M.; Lázničková-Galetová, M.; Jimenez, E.L.; Losey, R.; Sablin, M.; Bocherens, H.; Van den Broeck, M. Consumption of canid meat at the Gravettian Předmostí site, the Czech Republic. Foss. Impr. 2017, 73, 360–382. [Google Scholar] [CrossRef]
- Germonpré, M.; Lázniková-Galetová, M.; Sablin, M.V.; Bocherens, H. Self domestication or human control? The Upper Palaeolithic domestication of the dog. In Hybrid Communities: Biosocial Approaches to Domestication and Other Trans-Species Relationships; Stepanoff, C., Vigne, J.D., Eds.; Routledge: London, UK, 2018; pp. 39–64. [Google Scholar]
- Larson, G.; Karlsson, E.K.; Perri, A.; Webster, M.T.; Ho, S.Y.W.; Peters, J.; Stahl, P.W.; Piper, P.J.; Lingaas, F.; Fredholm, M.; et al. Rethinking dog domestication by integrating genetics, archeology, and biogeography. Proc. Natl. Acad. Sci. USA 2021, 109, 8878–8883. [Google Scholar] [CrossRef] [PubMed]
- Morey, D.F. In search of Paleolithic dogs: A quest with mixed results. J. Archaeol. Sci. 2014, 52, 300–307. [Google Scholar] [CrossRef]
- Boudadi-Maligne, M.; Escarguel, G. A biometric re-evaluation of recent claims for Early Upper Palaeolithic wolf domestication in Eurasia. J. Archaeol. Sci. 2014, 45, 80–89. [Google Scholar] [CrossRef]
- Ledoux, L.; Boudadi-Maligne, M. The contribution of geometric morphometric analysis to prehistoric ichnology: The example of large canid tracks and their implication for the debate concerning wolf domestication. J. Archaeol. Sci. 2015, 61, 25–35. [Google Scholar] [CrossRef]
- Skoglund, P.; Ersmark, E.; Palkopoulou, E.; Dalén, L. Ancient wolf genome reveals an early divergence of domestic dog ancestors and admixture into high-latitude breeds. Curr. Biol. 2015, 25, 1515–1519. [Google Scholar] [CrossRef] [PubMed]
- Bergström, A.; Frantz, L.; Schmidt, R.; Ersmark, E.; Lebrasseur, O.; Girdland-Flink, L.; Lin, A.T.; Storå, J.; Sjögren, K.-G.; Anthony, D.; et al. Origins and genetic legacy of prehistoric dogs. Science 2020, 370, 557–564. [Google Scholar] [CrossRef] [PubMed]
- Olsen, S. Origins of the Domestic Dog: The Fossil Record, 1st ed.; University of Arizona Press: Tucson, AZ, USA, 1985. [Google Scholar]
- Valadez, R. El origen del perro, primera parte (entre el lobo y el perro). AMMVEPE 2000, 11, 75–84. [Google Scholar]
- Valadez, R. El origen del perro (segunda parte): Entre el lobo doméstico y el criadero primitivo. AMMVEPE 2002, 13, 102–111. [Google Scholar]
- Savolainen, P.; Zhang, Y.-P.; Luo, J.; Lundeberg, J.; Leitner, T. Genetic evidence for an East Asian origin of domestic dogs. Science 2002, 298, 1610–1613. [Google Scholar] [CrossRef] [PubMed]
- Pang, J.F.; Kluetsch, C.; Zou, X.J.; Zhang, A.B.; Luo, L.Y.; Angleby, H.; Ardalan, A.; Ekström, C.; Sköllermo, A.; Lundeberg, J.; et al. mtDNA Indicate a single origin for dogs south of Yangtze River, less than 16.300 years ago, from numerous wolves. Mol. Biol. Evol. 2009, 26, 2849–2864. [Google Scholar] [CrossRef]
- Brown, S.K.; Pedersen, N.C.; Jafarishorijeh, S.; Bannasch, D.L.; Ahrens, K.D.; Wu, J.T.; Okon, M.; Sacks, B. La distinción filogenética de los cromosomas y del perro de aldea de Oriente Medio y el sudeste asiático ilumina los orígenes del perro. PLoS ONE 2011, 6, e28496. [Google Scholar]
- Pionnier-Capitan, M.; Bemilli, C.; Bodu, P.; Célérier, G.; Fierrié, J.G.; Fosse, P.; Garcià, M.; Vigne, J.D. New Evidence for Upper Palaeolithic small domestic dogs in South-Western Europe. J. Archaeol. Sci. 2011, 38, 2123–2140. [Google Scholar] [CrossRef]
- Ding, Z.L.; Oskarsson, M.; Ardalan, A.; Angleby, H.; Dahlgren, L.G.; Tepeli, C.; Kirkness, E.; Savolainen, P.; Zhang, Y.P. Origins of domestic dog in Southern East Asia is supported by analysis of Y-chromosome DNA. Heredity 2012, 108, 507–514. [Google Scholar] [CrossRef] [PubMed]
- Street, M.; Napierala, H.; Janssens, L. The late Palaeolithic dog from Bonn-Oberkassel in context. In The Late Glacial Burial from Oberkassel Revisited; Giemsch, L., Schmitz, R.W., Reinische, R.W., Eds.; Rheinische: Ausgrabungen, Germany, 2015; p. 72. [Google Scholar]
- Janssens, L.; Giemsch, L.; Schmitz, R.; Street, M.; Van Dongen, S.; Crombé, P. A new look at an old dog: Bonn-Oberkassel reconsidered. J. Archaeol. Sci. 2018, 92, 126–138. [Google Scholar] [CrossRef]
- Tchernov, E.; Valla, F.F. Two new dogs, and other Natufian Dogs, from the Southern Levant. J. Archaeol. Sci. 1997, 24, 65–95. [Google Scholar] [CrossRef]
- Pitulko, V.V.; Kasparov, A.K. Archaeological dogs from the Early Holocene Zhokhov site in the Eastern Siberian Arctic. J. Archaeol. Sci. Rep. 2017, 13, 491–515. [Google Scholar] [CrossRef]
- Botigué, L.R.; Song, S.; Scheu, A.; Gopalan, S.; Pendleton, A.L.; Oetjens, M.; Taravella, A.M.; Seregély, T.; Zeeb-Lanz, A.; Arbogast, R.M.; et al. Ancient European dog genomes reveal continuity since the Early Neolithic. Nat. Commun. 2017, 8, 16082. [Google Scholar] [CrossRef] [PubMed]
- Germonpré, M.; Fedorov, S.; Danilov, P.; Galeta, P.; Jimenez, E.L.; Sablin, M.; Losey, R.J. Palaeolithic and prehistoric dogs and Pleistocene wolves from Yakutia: Identification of isolated skulls. J. Archaeol. Sci. 2017, 78, 1–19. [Google Scholar] [CrossRef]
- Thalmann, O.; Shapiro, B.; Cui, P.; Schuenemann, V.J.; Sawyer, S.K.; Greenfield, D.L.; Germonpré, M.B.; Sablin, M.V.; López-Giráldez, F.; Domingo-Roura, X.; et al. Complete mitochondrial genomes of ancient canids suggest a European origin of domestic dogs. Science 2013, 342, 871–874. [Google Scholar] [CrossRef] [PubMed]
- Vila, C.; Savolainen, P.; Maldonado, J.E.; Amorim, I.R.; Rice, J.E.; Honeycutt, R.L.; Crandall, K.A.; Lundeberg, J.; Wayne, R.K. Multiple and ancient origins of the domestic dog. Science 1997, 276, 1687–1689. [Google Scholar] [CrossRef] [PubMed]
- Prassack, K.A.; Dubois, J.; Lázničková-Galetová, M.; Germonpré, M.; Ungar, P.S. Dental microwear as a behavioral proxy for distinguishing between canids at the Upper Paleolithic (Gravettian) site of Predmostí, Czech Republic. J. Archaeol. Sci. 2020, 115, 105092. [Google Scholar] [CrossRef]
- Germonpré, M.; Sablin, M.V.; Lázničková-Galetová, M.; Després, V.; Stevens, R.E.; Stiller, M.; Hofreiter, M. Palaeolithic dogs and Plesitocene wolves revisited: A replay to Morey (2014). J. Archaeol. Sci. 2015, 54, 210–216. [Google Scholar] [CrossRef]
- Bocherens, H.; Druckerm, D.G.; Germonpré, M.; Lázničková-Galetová, M.; Naito, Y.I.; Wissing, C.; Bruzek, J.; Oliva, M. Reconstruction of the Gravettian food-web at Predmostí I using multi-isotopic tracking (13C, 15N, 34S) of bone collagen. Quat. Int. 2015, 359, 211–228. [Google Scholar] [CrossRef]
- Crockford, S.J.; Juzmin, Y.V. Comments on Germonpré et al. J. Archaeol. Sci. 36, 2009 “Fossil dogs and wolves from Palaeolithic sites in Belgium, the Ukraine and Russia: Osteometry, ancient DNA and stable isotopes”, and Germonpré, Lázničková-Galetová and Sablin, J. Archaeol. Sci. 39, 2021 “Palaeolithic dog skull at the Gravettian Predmostí site, the Czech Republic”. J. Archaeol. Sci. 2012, 39, 2797–2801. [Google Scholar]
- Welker, M.H.; Byers, D.A.; McClure, S.B. “I wanna be your dog”: Evaluating the efficacy of univariate and multivariate methods for differentiating domestic and wild canids in North America. J. Osteoarchaeol. 2021, 31, 196–206. [Google Scholar] [CrossRef]
- Loog, L.; Thalmann, O.; Sinding, M.H.S.; Schuenemann, V.J.; Perri, A.; Germonpré, M.; Bocherens, H.; Wittm, K.E.; Samaniego-Castruita, J.A.; Velasco, M.S.; et al. Ancient DNA suggests modern wolves trace their origin to a Late Pleistocene Expansion from Beringia. Mol. Ecol. 2019, 29, 1596–1610. [Google Scholar] [CrossRef]
- Courtenay, L.A.; Yravedra, J.; Huguet, R.; Aramendi, J.; Maté-González, M.A.; González-Aguilera, D.; Arriaza, M.C. Combining machine learning algorithms and geometric morphometrics: A study of carnivore tooth marks. Palaeogeogr. Palaeoclimatol. Palaecol. 2019, 522, 28–39. [Google Scholar] [CrossRef]
- Courtenay, L.A.; Herranz-Rodrigo, D.; Huguet, R.; Maté-González, M.A.; González-Aguilera, D.; Yravedra, J. Obtaining new resolutions in carnivore tooth pit morphological analyses: A methodological update for digital taphonomy. PLoS ONE 2020, 15, e0240328. [Google Scholar] [CrossRef] [PubMed]
- Courtenay, L.A.; Herranz-Rodrigo, D.; González-Aguilera, D.; Yravedra, J. Developments in data science solutions for carnivore tooth pit classification. Sci. Rep. 2021, 11, 10209. [Google Scholar] [CrossRef] [PubMed]
- Yravedra, J.; Maté-González, M.A.; Courtenay, L.A.; González-Aguilera, D.; Fernández, M.F. The use of canid tooth marks on bone for the identification of livestock predation. Sci. Rep. 2019, 9, 16301. [Google Scholar] [CrossRef]
- Yravedra, J.; Herranz-Rodrigo, D.; Mendoza, C.; Aragón-Poza, P.; Courtenay, L.A. The use of tooth marks for new research into identifying and understanding the first domestic dogs in Palaeolithic populations. J. Archaeol. Sci. Rep. 2021, 40 Pt B, 103252. [Google Scholar] [CrossRef]
- Courtenay, L.A.; Herranz-Rodrigo, D.; Yravedra, J.; Vázquez-Rodríguez, J.M.; Huguet, R.; Barja, I.; Maté González, M.A.; Fernández, M.F.; Muñoz-Nieto, A.L.; González-Aguilera, D. 3D Insights into the Effects of Captivity on Wolf Mastication and Their Tooth Marks; Implications in Ecological Studies of Both the Past and Present. Animals 2021, 11, 2323. [Google Scholar] [CrossRef] [PubMed]
- Herranz-Rodrigo, D.; Tardáguila-Giacomozzi, S.J.; Courtenay, L.A.; Rodríguez-Alba, J.-J.; Garrucho, A.; Recuero, J.; Yravedra, J. New Geometric Morphometric Insights in Digital Taphonomy: Analyses into the Sexual Dimorphism of Felids through Their Tooth Pits. Appl. Sci. 2021, 11, 7848. [Google Scholar] [CrossRef]
- Arenas Esteban, J.A. Primeros resultados de las investigaciones arqueológicas en la Dehesa de Olmeda de Cobeta. In Actas del Segundo Simposio de Arqueología de Guadalajara; García-Soto, E., García-Valero, M.A., Martínez-Naranjo, J.P., Eds.; Junta de Comunidades de Castilla-La Mancha: Guadalajara, España, 2008; pp. 149–163. [Google Scholar]
- Arenas Esteban, J.A. El poblamiento Prerromano en el Área Alto Tajo-Alto Jalón. Complutum 2011, 22, 129–146. [Google Scholar] [CrossRef]
- Ruiz Zapata, B.; Gil García, M.J.; Martín Arroyo, T.; Arenas Esteban, J.A. El impacto del asentamiento celtibérico de Peña Moñuz (cuenca superior del Tajo) sobre el paisaje vegetal: Análisis polínico. Sociedad Geológica de España Geogaceta 2014, 55, 83–86. [Google Scholar]
- Arenas, J.A.; Marín Aguilera, B. El enclave fortificado de Peña Moñuz (Guadalajara): Diversidad cultural en la plenitud de la Edad del Hierro meseteña. Boletín Asoc. Amigos Mus. Guadalaj. 2020, 11, 35–61. [Google Scholar]
- Estaca-Gómez, V.; Aguilar Molina, L.; Arenas-Esteban, J. El aprovechamiento de los recursos cárnicos en el Castro Celtibérico de Peña Moñuz (Dehesa de Olmeda, Guadalajara). Munibe 2024. submitted. [Google Scholar] [CrossRef]
- Sutclife, A.J. Spotted hyaena: Crusher, gnawer, digester and collector of bones. Nature 1970, 227, 1110–1113. [Google Scholar] [CrossRef] [PubMed]
- Haynes, G. Evidence of carnivore gnawing on Pleistocene and recent mammalian bones. Paleobiology 1980, 6, 341–351. [Google Scholar] [CrossRef]
- Haynes, G. A guide for differentiating mammalian carnivore taxa responsible for gnaw damage to herbivore limb bones. Paleobiology 1983, 9, 164–172. [Google Scholar] [CrossRef]
- Maguire, J.M.; Pemberton, D.; Collet, M.H. The Makapansgat limeworks grey breccia: Hominids, hyenas, hystricids or hillwash? Paleontol. Afr. 1980, 23, 75–98. [Google Scholar]
- Binford, L.R. Bones: Ancient Men and Modern Myths; Academic Press: New York, NY, USA, 1981. [Google Scholar]
- Brain, C.K. The hunters or the Hunted? Introduction to African Cave Taphonomy; University of Chicago Press: Chicago, IL, USA, 1983. [Google Scholar]
- Horwitz, L.K.; Smith, P. The effects of striped hyaena activity on human remains. J. Archaeol. Sci. 1988, 15, 471–481. [Google Scholar] [CrossRef]
- Selvaggio, M.M. Carnivore tooth marks and stone tool butchery marks on scavenged bones: Archaeological implications. J. Hum. Evol. 1994, 27, 215–228. [Google Scholar] [CrossRef]
- Blumenschine, R.J. Percussion marks, tooth marks and the experimental determinations of the timing of hominin and carnivore access to long bones at FLK Zinjanthropus, Olduvai Gorge, Tanzania. J. Hum. Evol. 1995, 29, 21–51. [Google Scholar] [CrossRef]
- Fisher, J. Bone surface modifications in zooarchaeology. J. Archaeol. Method Theory 1995, 2, 7–68. [Google Scholar] [CrossRef]
- D’Errico, F.; Villa, P. Hooles and Grooves: The Contribution of microscopy and taphonomy to the problem of art origins. J. Hum. Evol. 1997, 33, 1–31. [Google Scholar] [CrossRef] [PubMed]
- D’Errico, F.; Villa, P. Modifications des os par les Hyènes et question des origines l’art. In Proceedings of the XVIII Rencontres Internationale d’Archeologie et d’Hisire d’Antibes, Antibes, France, 23–25 October 1997. [Google Scholar]
- Yravedra, J.; Lagos, L.; Bárcena, F. A taphonomic study of wild wolf (Canis lupus) modification of horse bones in northwestern Spain. J. Taphon. 2011, 9, 37–66. [Google Scholar]
- Courtenay, L.A.; Yravedra, J.; Maté-González, M.A.; Vázquez-Rodríguez, J.M.; Fernández-Fernández, M.; González-Aguilera, D. The effects of prey size on carnivore tooth mark morphologies on bone: The case study of Canis lupus signatus. Hist. Biol. 2020, 33, 2760–2772. [Google Scholar] [CrossRef]
- Courtenay, L.; Yravedra, J.; Herranz-Rodrigo, D.; Rodríguez-Alba, J.J.; Serrano-Ramos, A.; Estaca-Gómez, V.; Gonzálo-Aguilera, D.; Solano, J.A.; Jiménez-Arenas, J.M. Deciphering carnivoran competition for animal resources at the 1.46 Ma early Pleistocene site of Barranco León (Orce, Granada, Spain). Quat. Sci. Rev. 2022, 300, 107912. [Google Scholar] [CrossRef]
- Yravedra, J.; Andrés, M.; Fosse, P.; Besson, J.P. Taphonomic analysis of small ungulates modified by fox (Vulpes vulpes) in Southwestern Europe. J. Taphon. 2014, 12, 37–67. [Google Scholar]
- Yravedra, J.; Andrés, M.; Domínguez Rodrigo, M. A taphonomic study of the African wild dog (Lycaon pictus). Archaeol. Anthropol. Sci. 2014, 6, 113–124. [Google Scholar] [CrossRef]
- Rodríguez Alba, J.J.; Linares-Matás, G.; Yravedra, J. First assessments of the taphonomic behaviour of jaguar (Panthera onca). Quat. Int. 2019, 517, 88–96. [Google Scholar] [CrossRef]
- Selvaggio, M.M.; Wilder, J. Identifying the Involvement of Multiple Carnivore Taxa with Archaeological Bone Assemblages. J. Archaeol. Sci. 2001, 28, 465–470. [Google Scholar] [CrossRef]
- Adams, D.C.; Collyer, M.; Otarola-Castillo, E.; Sherratt, E. Geometric Morphometric Analyses of 2D/3D Landmarks Data, version 4.0.9; Package ‘geomorph’; CRAN: Vienna, Austria, 2014. [Google Scholar]
- O’Higgins, P.; Jones, N. Facial growth in Cercocebus torquatus: An application of three-dimensional geometric morphometry techniques to the study of morphological variation. J. Anat. 1998, 193, 251–272. [Google Scholar] [CrossRef] [PubMed]
- Gower, J.C. Generalized Procrustes analysis. Psychometrika 1975, 40, 33–50. [Google Scholar] [CrossRef]
- Ten-Berge, J.M.F. Orthogonal Procrustes rotation for two or more matrices. Psychometrika 1977, 42, 267–276. [Google Scholar] [CrossRef]
- Goodall, C.R. Procrustes methods in the statistical analysis of shape (with discussion). J. R. Stat. Soc. 1991, 53, 285–339. [Google Scholar] [CrossRef]
- Kent, J.T. The complex Bingham distribution and shape analysis. J. R. Stat. Soc. 1994, 56, 285–299. [Google Scholar] [CrossRef]
- Dryden, I.L.; Mardia, K.V. Statistical Shape Analysis, with Applications in R, 2nd ed.; John Miley & Sons: Chichester, UK, 2016; pp. 125–173. [Google Scholar]
- Vega-Toscano, L.G.; Cerdeño-Serrano, L.; Córdoba de Oya, B. El Origen de los Mastines Ibéricos: La Trashumancia entre los Pueblos Prerromanos de la Meseta. Complutum 1998, 9, 117–135. [Google Scholar]
- Koudelka, F. Das Verhältnis der Ossa longa zur Skeletthöhe bei den Säugetieren. Verhandlungen Naturforschenden Vereines Brünn 1885, 24, 127–153. [Google Scholar]
- Iborra, P. La Ganadería y la Caza desde el Bronce Final hasta el Ibérico Final en el Territorio Valenciano; Diputación provincial de Valencia, Memorias del SIP: Valencia, Spain, 2004. [Google Scholar]
- Sanchis, A.; Sarrión, I. Restos de cánidos (Canis familiaris ssp) en yacimientos valencianos de la Edad del Bronce. Arch. Prehist. Levantina 2004, 25, 161–198. [Google Scholar]
- Estaca, V.; De la Torre, A.; Tardaguila, S.; Major, M. Zooarchaeological study of Aldovea (Torrejón de Ardoz, Madrid), a new Bronze Age archaeological site from the Iberian Peninsula inland. J. Archaeol. Sci. Rep. 2024, 57, 104616. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lab Reference | Material | Radiocarbon Date | Average Date | Calibrated Dates |
|---|---|---|---|---|
| Beta 270931 | Bone | 2290 ± 40 BP | 2210 ± 40 BP | 400-350 BC/290-220BC |
| Beta 270932 | Bone | 2180 ± 40 BP | 2100 ± 40 BP | 370-150 BC/140-110BC |
| Beta 270933 | Bone | 2220 ± 40 BP | 2140 ± 40 BP | 390-180BC |
| Beta 270934 | Bone | 2290 ± 40 BP | 2190 ± 40 BP | 400-350 BC/290-220BC |
| Beta 99068 | Bone | 2280 ± 40 BP | 2210 ± 40 BP | 400-355 BC/265-230BC |
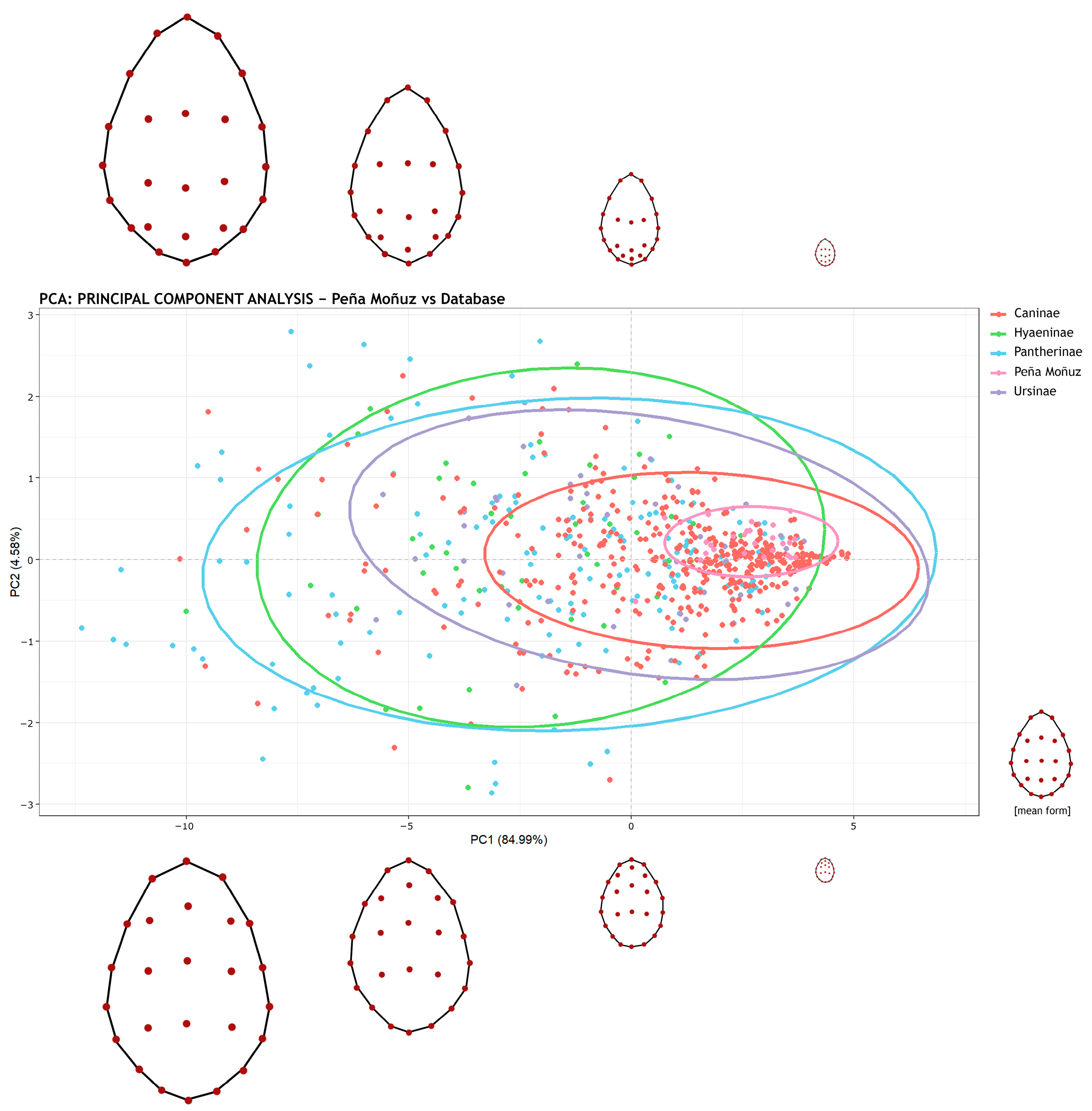
| U. arctos | C. familiaris | V. vulpes | H. sapiens | C. crocuta | P. onca | P. pardus | P. leo | L. pictus | Peña Moñuz | |
|---|---|---|---|---|---|---|---|---|---|---|
| C. familiaris | 0.001 | - | - | - | - | - | - | - | - | - |
| V. vulpes | 0.001 | 0.002 | - | - | - | - | - | - | - | - |
| H. sapiens | 0.068 | 0.174 | 0.052 | - | ||||||
| C. crocuta | 0.002 | 0.001 | 0.001 | 0.005 | - | - | - | - | - | - |
| P. onca | 0.001 | 0.001 | 0.001 | 0.023 | 0.642 | - | - | - | - | - |
| P. pardus | 0.002 | 0.002 | 0.017 | 0.046 | 0.001 | 0.001 | - | - | - | - |
| P. leo | 0.001 | 0.001 | 0.001 | 0.001 | 0.453 | 0.755 | 0.001 | - | - | - |
| L. pictus | 0.052 | 0.001 | 0.001 | 0.071 | 0.434 | 0.260 | 0.004 | 0.022 | - | - |
| Peña Moñuz | 0.001 | 0.023 | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 | - |
| C. lupus | 0.072 | 0.003 | 0.001 | 0.667 | 0.001 | 0.001 | 0.003 | 0.001 | 0.004 | 0.001 |
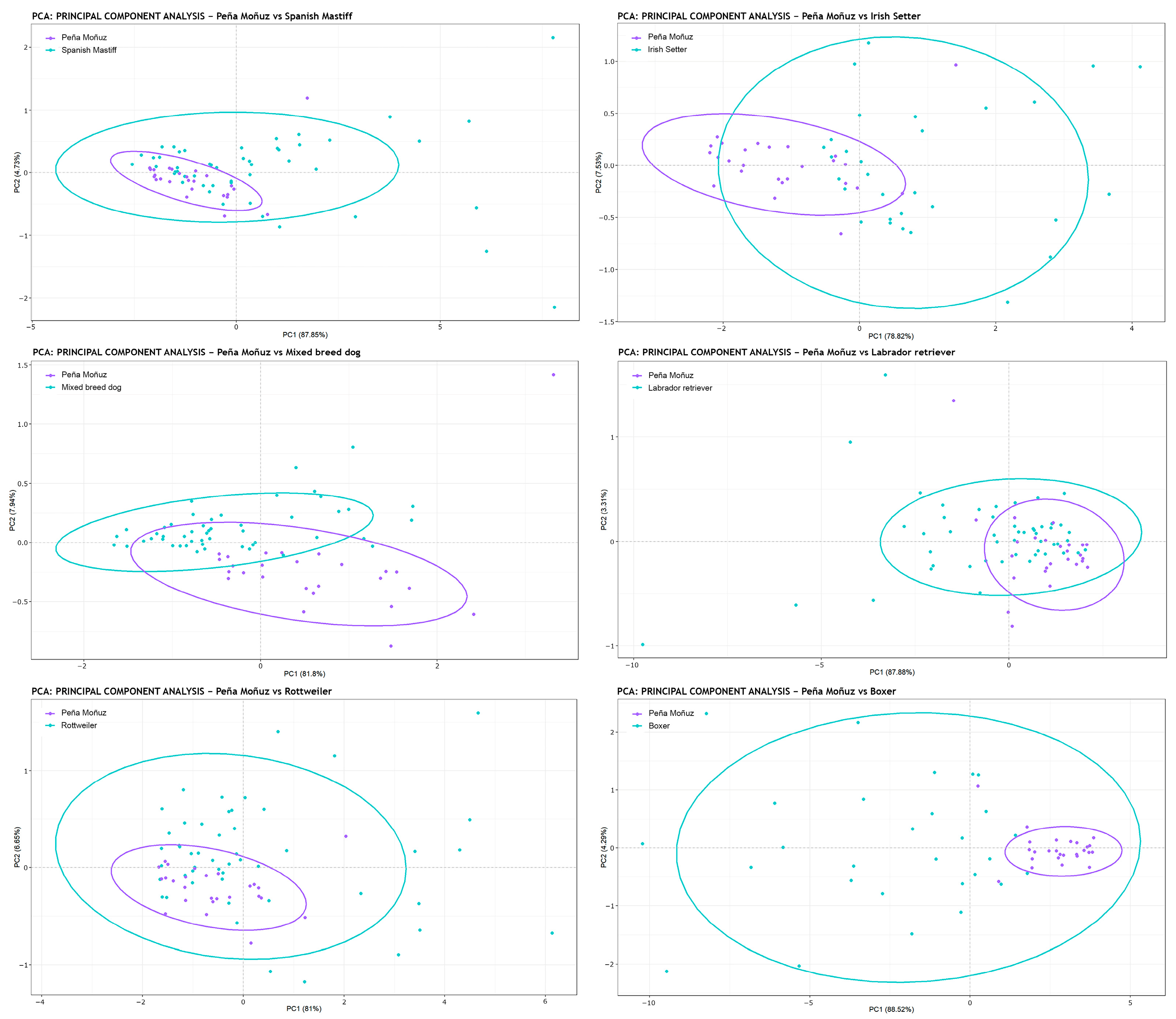
| Boxer | Labrador | Mastiff | Mixed Breed | Peña Moñuz | Rottweiler | |
|---|---|---|---|---|---|---|
| Labrador retriever | 0.001 | - | - | - | - | - |
| Spanish Mastiff | 0.001 | 0.001 | - | - | - | - |
| Mixed breed | 0.001 | 0.001 | 0.223 | - | - | - |
| Peña Moñuz | 0.001 | 0.007 | 0.001 | 0.001 | - | - |
| Rottweiler | 0.050 | 0.001 | 0.001 | 0.001 | 0.001 | - |
| Irish Setter | 0.026 | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 |
| Boxer | Labrador | Mastiff | Mixed Breed | Peña Moñuz | Rottweiler | |
|---|---|---|---|---|---|---|
| Labrador retriever | 0.001 | - | - | - | - | - |
| Spanish Mastiff | 0.001 | 0.001 | - | - | - | - |
| Mixed breed | 0.001 | 0.001 | 0.001 | - | - | - |
| Peña Moñuz | 0.001 | 0.004 | 0.001 | 0.001 | - | - |
| Rottweiler | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 | - |
| Irish Setter | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Claver, I.; Estaca-Gómez, V.; Linares-Matás, G.J.; Arenas-Esteban, J.A.; Yravedra, J. Taphonomy as a Methodological Approach for the Study of Dog Domestication: Application to the Prehistoric Site of Peña Moñuz (Guadalajara, Spain). Heritage 2025, 8, 34. https://doi.org/10.3390/heritage8010034
Claver I, Estaca-Gómez V, Linares-Matás GJ, Arenas-Esteban JA, Yravedra J. Taphonomy as a Methodological Approach for the Study of Dog Domestication: Application to the Prehistoric Site of Peña Moñuz (Guadalajara, Spain). Heritage. 2025; 8(1):34. https://doi.org/10.3390/heritage8010034
Chicago/Turabian StyleClaver, Idoia, Verónica Estaca-Gómez, Gonzalo J. Linares-Matás, Jesús Alberto Arenas-Esteban, and José Yravedra. 2025. "Taphonomy as a Methodological Approach for the Study of Dog Domestication: Application to the Prehistoric Site of Peña Moñuz (Guadalajara, Spain)" Heritage 8, no. 1: 34. https://doi.org/10.3390/heritage8010034
APA StyleClaver, I., Estaca-Gómez, V., Linares-Matás, G. J., Arenas-Esteban, J. A., & Yravedra, J. (2025). Taphonomy as a Methodological Approach for the Study of Dog Domestication: Application to the Prehistoric Site of Peña Moñuz (Guadalajara, Spain). Heritage, 8(1), 34. https://doi.org/10.3390/heritage8010034