Abstract
Biochar (Bc) and liquid vermicompost extracts (LVEs) are increasingly being used as biofertilizers in agriculture to promote soil-microbe-crop interactions. However, although both these products can potentially act synergistically due to their complementary characteristics, their co-application in different soils has not yet been investigated. Therefore, firstly, an LVE-activated biochar (BLVE) was experimentally formulated and the persistence of LVE bacteria over a 60-day storage period was determined. The total number of LVE bacteria increased by 10-fold after 7 days and was stable throughout the entire biochar storage period. In addition, changes in the composition of the bacterial community were observed after 30 days of storage, indicating that taxa less represented in pure LVE may be advantaged upon biochar colonization. Secondly, a microcosm experiment was performed to evaluate whether the biological fertility and enzyme activities of two soils, differing in organic matter content, could be enhanced by the addition of LVE-activated biochar. In this experiment, three different doses of Bc, LVE, and BLVE against the carbon-related biological fertility index (i.e., biological fertility index, BFI) and three enzyme activities over a 21-day incubation period were tested. The BLVE treatment yielded the best results (i.e., BFI +32%, enzyme activities +38%). This indicates that Bc and LVEs can act synergistically to promote soil fertility, quality, and microbial activity. By integrating LVE-activated biochar into their soil management practices, farmers could achieve higher crop yields and healthier products.
1. Introduction
Animal and plant biomass, and particularly products derived from its biological and thermochemical conversion (e.g., vermicompost and biochar, respectively), are attracting increasing attention in agriculture for their ability to improve soil quality and crop yield [1,2].
Vermicompost is the product of the decomposition of organic waste by microorganisms through the digestive tract of earthworms [3]. Both water-soluble nutrients and microorganisms can be further extracted during steeping of vermicompost in water and formulated into nutrient-rich and microbiologically active liquid vermicompost extracts (LVEs) [4]. Bacteria are essential components of LVEs, as they fulfill multiple functions that promote both soil and plant health, including nutrient solubilization, biocontrol, or the active release of plant growth stimulators [5,6]. On the other hand, the appeal of biochar, the solid byproduct of biomass pyrolysis, stems primarily from its capacity to improve the soil structure and quality in a wide array of ways; for instance, biochar can improve soil carbon sequestration, soil water- and nutrient-holding capacity, and soil porosity and thus aeration [7,8,9]. Interestingly, biochar has recently emerged as a promising carrier material for the delivery of beneficial microorganisms in the soil, as it provides a stable environment for bacterial survival and activity, potentially enhancing their longevity and effectiveness in the soil [10,11].
Although LVEs and biochar are both considered useful and sustainable soil organic amendments, their broad-scale application has not yet been widely adopted as contrasting findings have been reported on their effects on soil health. As an example, depending on the soil type, the sole application of LVEs can be prone to runoff and leaching under excessive rainfall/irrigation conditions [12], while variable biochar dosages can increase soil salinization in the long term, decrease soil fertility, and/or adversely affect soil microbial activity [13,14]. In this scenario, there is a growing consensus that these potentially adverse effects of LVEs and biochar could be mitigated by their co-application, as both amendments have been shown to act in synergy given their different, yet potentially complementary attributes [15,16,17]. The co-application of LVEs and biochar has usually involved a prior soil amendment with biochar followed by the application of LVEs by fertigation [18,19]. While this strategy has been shown to provide benefits in terms of both plant growth and protection [20,21,22,23], recent studies have investigated the formulation of “pre-conditioned” or “activated” biochars [24]. The process of biochar activation offers a more targeted approach, allowing more controlled integration of both organic amendments and facilitating the contact of LVE-associated microorganisms and biochar particles without the interference of the soil matrix [25]. In addition, effective biochar formulations could also increase its ease of application, transport, and commercialization, while minimizing the potential water wastage that may occur in the dual application process. However, analyses of the bacteria present in LVEs are scarce [4,26], and no study exists on their persistence in biochar particles after activation. Moreover, previous studies have focused on the use of vermicompost-activated biochars in the bioremediation context, showing the synergistic effects of microorganisms and biochar in metal-metribuzin- and cadmium-contaminated soils [27,28,29]. However, the functional consequences of amendment with LVE-activated biochar for soil quality have not yet been investigated.
In the first part of this study, both in-plate isolation methods and 16SrRNA-based sequencing were used to monitor bacterial composition and persistence in LVE-activated biochar. In the second part, the efficacy of LVE-activated biochar was evaluated in two soils with very low and moderate organic matter content. We hypothesized that (1) the biochar can be a suitable environment for the growth and persistence of the inoculated bacteria, and (2) the soil quality and biological fertility could be improved by the addition of LVE-activated biochar.
2. Materials and Methods
2.1. Biochar and LVE Characteristics
Biochar was produced from pruning residues’ woodchips (G30–G50) of a consortium of selected tree species (Abies sp., Alnus sp., Castanea sativa, Fraxinus sp., Quercus sp., and Robinia pseudoacacia) by a pyrolysis process (Bio-Esperia S.r.l., Arezzo, Italy). Biochar production was characterized by a 10 h process duration with an average heating rate of 75–80 °C min−1 before reaching a peak of 1100 °C. The parameters for the characterization of biochar were analyzed through certified methods approved by Italian regulations (Law Decree 75/2015): 2–5 mm particle diameter, 400% water-holding capacity (WHC), 9.8 pH, 69% total organic carbon (TOC), <0.1% total carbonate (total CaCO3), <0.4% total nitrogen (TN), 0.034% available phosphorus (P), and 115 cmol(+) kg−1 cation-exchange capacity (CEC). The TOC content was Class 1 following the Guidelines for Certification of the International Biochar Initiative [30].
The liquid vermicompost extract (LVE) was derived from cattle and horse manure vermicompost produced by Centro di Lombricoltura Toscano (CLT S.r.l, Pisa, Italy). Manure was converted into vermicompost in an open-air litter with two earthworm species (i.e., Eisenia fetida and Eisenia andrei) in a 6-month composting process; after the vermicompost was fully matured, a portion was cold water-extracted using air insufflation over a 48 h period to produce the LVE. The chemical properties of the LVE were as follows: 4.8 pH, 4.05 mS cm−1 electrical conductivity (EC), 0.1% TOC, 101 ppm TN, 100 ppm total organic nitrogen (TON), and 10 C/N ratio.
2.2. Formulation of LVE-Activated Biochar and Microbiological Analysis
2.2.1. Biochar Sterilization and Contamination Check
Biochar was sterilized to remove the presence of inherent microbial populations. Briefly, 1 g of biochar was transferred in triplicate (n = 3) to 100 mL flasks and autoclaved at 121 °C for 20 min. Flasks were closed tightly and stored for five days at 25 °C in the dark. At each storage day, flasks were filled with 20 mL of Luria Bertani (LB) medium, composed as follows (g L−1): 10 tryptone, 5 yeast extract, 10 sodium chloride (NaCl) at 7.0 pH, and incubated overnight in a rotary shaker (120 rpm). Subsequently, a 50 µL aliquot from these flasks was serially diluted up to the 105-fold, and each dilution spread on LB agar (1.5%, w/w) plates, which were incubated for five days at 25 °C. Finally, the number of colony-forming units (CFUs) was counted and recorded. The same experiment was repeated using non-autoclaved biochar as the control. Both the sterilization process and the activation of biochar with the LVE were performed at the Biology Department of the University of Florence (Italy).
2.2.2. Preparation of Water- and LVE-Activated Biochars
An amount of 1 g of autoclaved biochar was mixed either with sterile distilled water (BC = biochar as control) or with LVE (BLVE = LVE-activated biochar) at 1:20 (w:v) ratio in 100 mL flasks, and incubated statically at 25 °C for 24 h. Flasks were then drained, closed tightly, and stored for 1, 7, 15, 30, and 60 days at 25 °C in the dark for subsequent analyses (Figure 1).
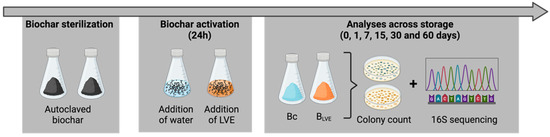
Figure 1.
Flowchart illustrating the different tasks carried out throughout the formulation of water- and LVE-activated biochars (BC and BLVE, respectively).
2.2.3. Bacterial Quantification
At each storage day, CFUs were quantified through the drop-plate method as follows: both BC and BLVE were mixed with a sterile 0.8% NaCl solution at 1:20 (w:v) ratio and incubated in a rotary shaker at 25 °C and 150 rpm for 2 h. A 20 µL aliquot of each mixture was serially diluted in a 0.8% NaCl solution up to the 108-fold, and 10 µL of each dilution dropped on LB agar and incubated overnight at 25 °C. Dilutions containing between 30 and 300 CFUs were counted, and their concentration expressed as CFU g−1 of biochar. The number of CFUs was also determined in the pure LVE as described for BC and BLVE, even if, in this case, their concentration was expressed as CFU mL−1. All experiments in each storage day were performed in triplicate.
2.2.4. Bacterial Isolation and Molecular Identification
Isolation of bacteria from pure LVE, Bc, and BLVE was carried out from the above-described serial dilutions performed. Hence, 50 µL from the dilutions, containing between 30 and 300 CFUs, was plated on LB agar and incubated for one week at 25 °C. Plates were thoroughly observed under a stereomicroscope and unique morphotypes were isolated and re-plated as many times as necessary until pure colonies were obtained. The assay was performed in triplicate.
Molecular identification of bacteria was carried out through the amplification and sequencing of the partial 16S recombinant DNA region using universal forward primers 104F 5′-GCACGGGTGAGTAACACGTG-3′ [31] and reverse 1392R 5′-ACGGGCGGTGTGTRC-3′ [32]. A single colony was resuspended in sterile water and the DNA extracted with thermal shock (96 °C for 10 min and cooled at −20 °C for 30 min). Samples were then centrifuged, and the supernatant used as the template for amplification. Each PCR reaction was performed in 25 µL; the mixture contained the following: 5 µL of 5X Colorless GoTaq® Reaction Buffer (Promega, Madison, WI, USA) (1X final concentration), 0.5 µL of forward primer (final concentration 0.5 µM), 0.5 µL of reverse primer (final concentration 0.5 µM), 0.5 µL of dNTPs mix (10 mM), 0.5 µL magnesium chloride (25 mM), 0.2 µL of Taq polymerase (1 U/sample), 1 µL of DNA, and sterile deionized water to 25 µL. Amplification protocol: initial denaturation at 94 °C for 3 min, followed by 30 cycles of 94 °C for 30 s; 56 °C for 40 s and 72 °C for 1 min; and a final elongation step at 72 °C for 5 min. The PCR products were purified using NucleoFast® 96 PCR Plate, 96-well ultrafiltration plate for PCR clean-up (MACHEREY-NAGEL, Düren, Germany), and sequenced with the Sanger method. DNA sequences were deposited into the GenBank repository. The taxonomic affiliations and respective accession numbers of the isolated bacteria are listed in Table S1.
2.3. pH and EC Determination in LVE-Activated Biochar
Both BC and BLVE were mixed with distilled water at 1:20 (w:v) ratio, and mixtures were left to stand for 30 min. Subsequently, both the pH and EC were measured using a pH meter (Edge® HI2002, HANNA Instruments Inc., Woonsocket, RI, USA) and an EC meter (BASIC 30, Crison Strumenti SpA, Carpi, Italy), respectively [33]. These measurements were carried out in triplicate.
2.4. Validation of the Biochar-Activation Protocol on Two Different Soils
2.4.1. Soil Experimental Setup
The soil experiment was conducted twice to increase the amount of data and improve the robustness of the statistical analysis. To differentiate between the effects of the BC, LVE, and BLVE doses, each soil experiment was conducted in triplicate with nine treatments for both soil types, using 250 mL microcosms (a total of 60 microcosms). This study applied four treatments to evaluate the soil parameter changes: a control (Cnt), and low, medium, and high doses of Bc, LVE, and BLVE at 0.5%, 1%, and 2% (w/w), respectively. The treatments (BC, LVE, and BLVE) were combined with the acronyms L- (low), M- (medium), and H- (high) based on the indicated dose (0.5%, 1%, and 2%). The Cnt, BC, and BLVE samples were moistened with deionized water at 60% of WHC, which is considered an ideal moisture value for soil biological activity. The LVE samples were moistened with LVE solutions diluted in deionized water to achieve 0.5%, 1%, and 2% (w/w) doses, reaching 60% of WHC. The microcosms used in the experiment were made of glass containers sealed with rubber stoppers. The soil inside the containers was moistened daily to maintain the water content at the initial WHC. The jars were opened daily to aerate the samples and measure soil respiration. The samples were kept in the dark for an incubation period of 21 days at 25 ± 1 °C. At the end of this period, the samples were stored at 4 °C for chemical and enzyme analyses.
2.4.2. Soil Recovering
The surface (0–15 cm) of two agricultural sandy soils was collected from two locations. The first one was derived from a field owned by the agricultural research center “Enrico Avanzi” of the University of Pisa (Pisa, Italy; Lat. 43°39′38.96″ N; Long. 10°18′22.17″ E; 1 m above sea level) and it was given the name “Weak Soil” (W-S), due to its very low (1.0%) soil organic matter (SOM) content. The second one was collected in a countryside field at Colle di Compito (Lucca, Italy; Lat. 43°46′21.34″ N; Long. 10°36′24.86″ E; 42 m above sea level) and, for its moderate (2.2%) SOM content, was named “Middle Soil” (M-S). The soil samples were collected in September 2023 by drilling 20 cores, each measuring 5 cm in diameter and 15 cm in depth. The soils were air-dried and passed through a 2 mm sieve to remove larger residues.
2.4.3. Soil Analyses
The main physical and chemical parameters of both soils (i.e., W-S and M-S) are listed in Table 1. The parameters, such as texture, WHC, pH in water, and CEC, were measured according to standard methods (n = 3) [34].

Table 1.
Main soil physical and chemical parameters (mean ± SD; n = 3) of “Weak Soil” (W-S) and “Middle Soil” (M-S).
Total organic carbon was determined by subtracting the inorganic carbon (total CaCO3) amount from the total carbon (TC) content. A vario Micro Element Analyzer (Elementar Italia Srl, Lomazzo, Como, Italy) was used to conduct the TC analysis. Soil organic matter content was derived using the van Bemmelen coefficient (1.724). The total CaCO3 was measured using a Scheibler apparatus (Gabrielli Technology, Calenzano, Firenze, Italy) (n = 6).
To determine the soil’s microbial biomass carbon (MB-C), a method was used where organic C was extracted from both fumigated and non-fumigated soils using 1 N K2SO4 [35]. The extracted organic C was then measured by QBD1200 Laboratory TOC Analyzer (Hach Co., Loveland, CO, USA). The difference in soluble C between the fumigated and non-fumigated soils was then converted into microbial biomass carbon (MB-C) using an extraction efficiency coefficient (Kc) of 0.45 (n = 6).
During the experimental period, a 21-day aerobic incubation was carried out to determine the sample’s potential to mineralize organic C. The evolution of carbon dioxide (CO2) was monitored daily from day 1 to day 21. An amount of 100 g of soil was placed in 250 mL glass containers, which were sealed with rubber stoppers. The soil was moistened to 60% of WHC and incubated at 25 ± 1 °C. The CO2 that evolved was trapped in NaOH solution, and the excess alkali was titrated with HCl [36]. The results were normalized with respect to time and expressed as mg of mineralized C 100 g−1 of dry soil (n = 6).
The soil biological fertility index (BFI) [37] is based on six variables as follows:
where Res_Bas (ppm) is the basal respiration, Res_Cum (ppm) is the cumulative respiration, MB-C (ppm C-CO2) is the microbial biomass carbon, SO (%) is the organic matter, qCO2 (mg C-CO2·10−2·h−1·mg C mic−1) is the specific respiration of biomass, with an increase in CO2 being related to microbial stress [38] or to changes in the bacterial-to-fungal ratio [39,40], and, finally, qM (%) is the mineralization quotient expressing the ratio of cumulative respiration to organic C content (Res_Cum·TOC−1·100). The values for each variable that contribute to the BFI were grouped into five levels [41], based on previous studies [42,43]. These levels were then added together to create a BFI score that ranges from 1 to 30. The BFI levels indicate the different levels of soil fertility as follows: (I) BFI < 9 represents stressed soils with very low fertility, (II) 9 < BFI < 12 represents pre-stress soils, (III) 13 < BFI < 18 represents soils with intermediate fertility, (IV) 19 < BFI < 24 represents good fertility soils, and (V) BFI > 24 represents soils with very high fertility.
All the enzyme activities were analyzed by spectrophotometric methods (n = 3). Alkaline phosphatase activity (APA) was measured using p-nitrophenyl phosphate incubated with soil samples at pH 11 and 37 °C for 60 min. The reaction product (p-nitrophenol) was extracted by dilute alkali (0.5 M CaCl2 and 0.5 M NaOH) and determined at 410 nm [44]. β-glucosidase activity (βGA) was determined using a substrate of 4-nitrophenyl-β-D-glucopyranoside. After incubation at 37 °C for 60 min, the production of p-nitrophenol was measured at 410 nm [45] as for APA analysis. Both APA and βGA activities were expressed as μmol p-nitrophenol·g−1 dry soil·h−1. Urease activity (UA) was spectrophotometrically (690 nm) measured according to Kandeler and Gerber [46] by analyzing the ammonia production after a 2 h incubation of soil samples with urea substrate at 37 °C; UA activity was expressed as µg of NH4+-N g−1 soil 2 h−1 [46].
The effect of the treatments on the soil quality was assessed by determining the soil alteration index 3 (SAI3), as defined by Puglisi et al. [47]. This index was calculated by processing the enzyme data according to the following formula:
where the enzyme activities were expressed in μmol of p-nitrophenol g−1 of dry soil h−1 (for phosphatase and β-glucosidase), and in μg of urea g−1 of dry soil 2 h−1 (urease).
2.5. Statistical Analyses
All results are presented as mean ± SD and the Levene’s test was performed to assess the equality of variances among the samples. The data regarding both biochars’ pH and EC approached a normal distribution (Shapiro-Wilk test, p < 0.05), and hence a Student’s t-test for independent samples was used to check for statistically significant differences in these parameters between BC and BLVE at each storage day.
The statistical analysis of the soil chemical and biochemical results was conducted using JMP software v. 17.2 (SAS Institute, Inc., Cary, NC, USA). The soil data were calculated on a dry weight basis. One-way ANOVA was used to check for differences among treatments. The Tukey-Kramer post-hoc test was performed to identify significantly different means at p < 0.05.
3. Results
3.1. Biochar Sterilization Test
Biochar sterilization through autoclaving was effective in removing the biochar’s inherent bacterial populations, as shown in Table 2. On the other hand, 2.5 and 9.3 × 103 CFU g−1 were recovered from non-autoclaved biochar after 24 and 48 h, respectively, where the CFU number stabilized after 72 h (20 × 103 CFU g−1) (Table 2).
Table 2.
Number of CFU (mean CFU g−1 × 103 ± SE), grown in LB plates from non-autoclaved and autoclaved biochar after 24, 48, 72, and 96 h.
3.2. Quantitative and Qualitative Analyses of Bacteria across BC and BLVE Storage
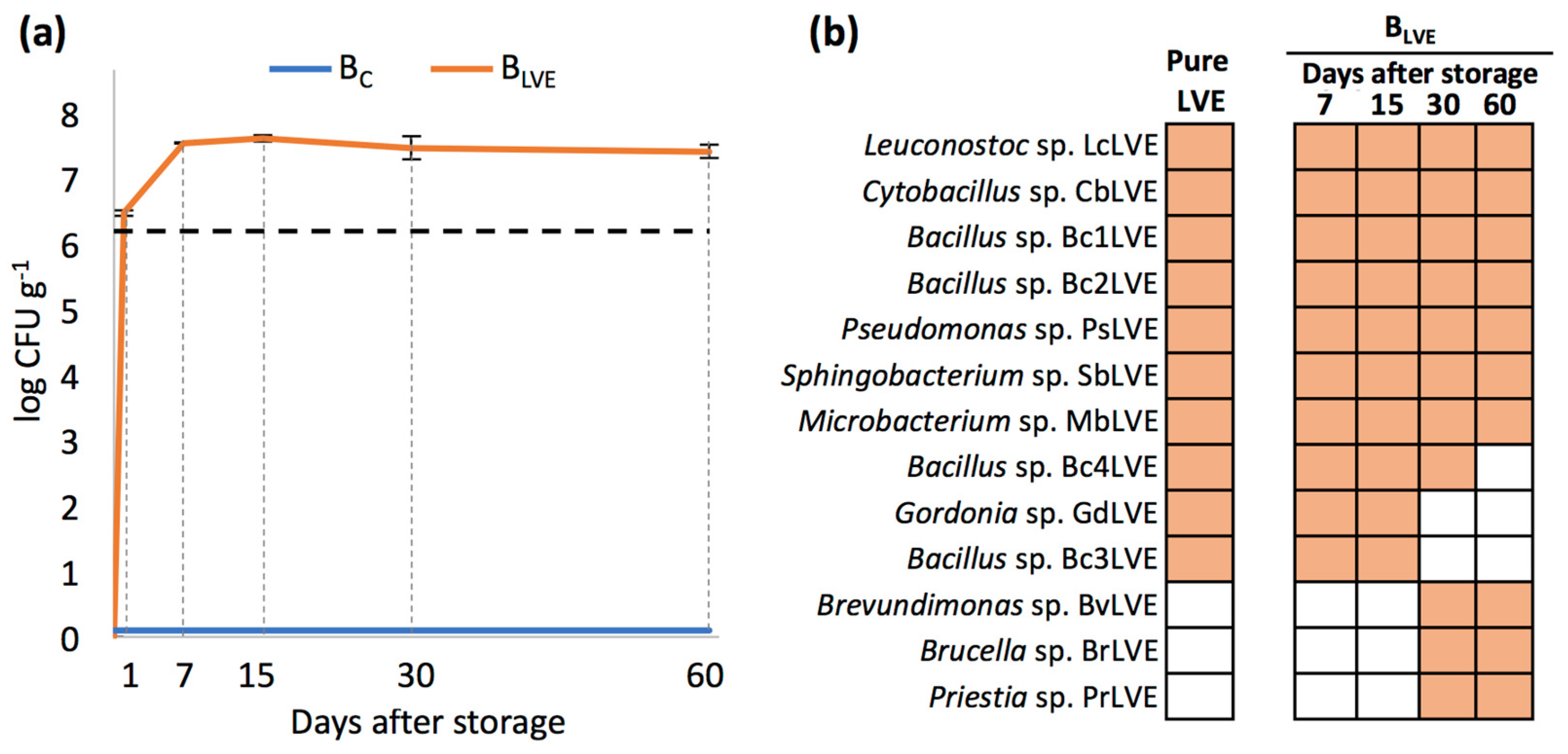
Regardless of the storage time examined, no CFUs were recovered in the Bc. On the other hand, nearly 3 × 106 CFUs g−1 were recovered from the BLVE one day after storage, more than double compared to those present in the pure LVE (~1.3 × 106 CFUs mL−1). The bacterial population in the BLVE reached its maximum at 7 days after storage (~4 × 107 CFUs g−1) and remained around this order of magnitude until 60 days after storage (Figure 2a).
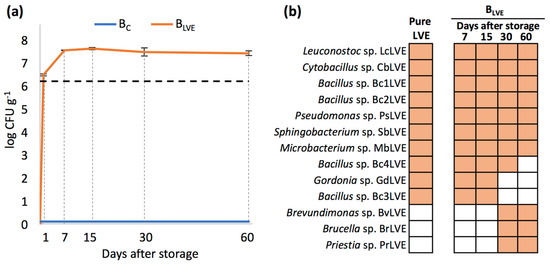
Figure 2.
(a) Total number of CFUs (mean CFU g−1 ± SD) isolated from BC and BLVE across the different days after storage. The dashed line represents the number of CFUs counted in the initial pure LVE; (b) presence of each bacterium in the pure LVE and in BLVE across the different days after storage.
As for qualitative analyses, ten bacterial morphotypes were isolated from the pure LVE, which were all recovered from the BLVE both 7 and 15 days after storage. Most of these bacteria belonged to the phyla Bacillota (i.e., Leuconostoc sp. LcLVE, Cytobacillus sp. CbLVE, Bacillus sp. Bc1LVE, Bacillus sp. Bc2LVE, Bacillus sp. Bc3LVE, and Bacillus sp. Bc4LVE) followed by Actinomycetota (i.e., Microbacterium sp. MbLVE and Gordonia sp. GdLVE), Pseudomonadota (i.e., Pseudomonas sp. PsLVE), and Bacteroidota (i.e., Sphingobacterium sp. SbLVE). On the other hand, two bacterial genera, including Gordonia sp. GdLVE and Bacillus sp. Bc3LVE, were not detected in the BLVE 30 days after storage, while three previously undetected genera, including Bervundimonas sp. BvLVE, Brucella sp. BrLVE, and Priestia sp. PrLVE, were recovered for the first time at this stage. Bacterial members recovered 30 days after storage, except for Bacillus sp. Bc4LVE, were also recovered from the BLVE 60 days after storage (Figure 2b).
3.3. pH and EC Analyses of BC and BLVE
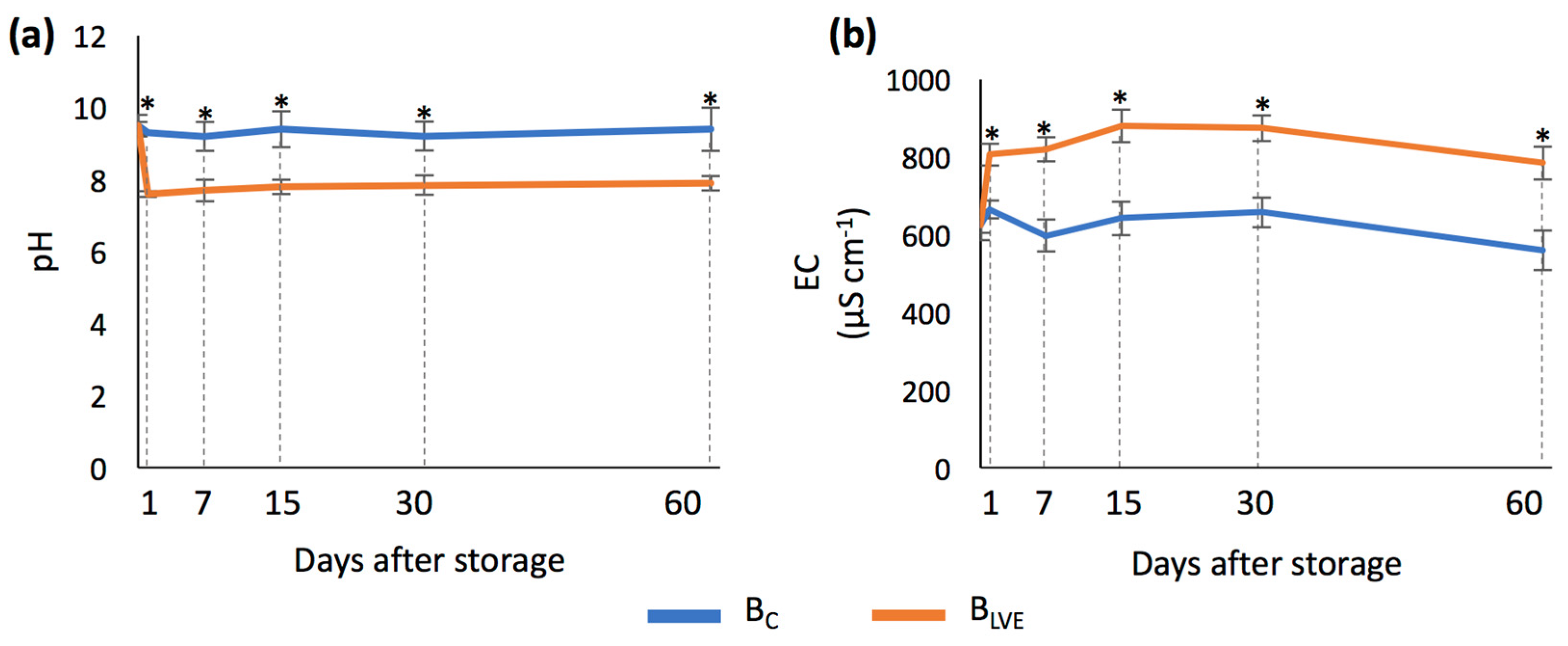
The activation of the biochar with the LVE led to significant changes in both the pH and EC (Figure 3a,b). Compared to the BC, activation with the LVE decreased the biochar’s pH from 9.3 to 7.6, while it increased the EC from 665 to 806 µS cm−1 on the first day of storage. The extent of these changes was maintained throughout the entire 60-day storage period.
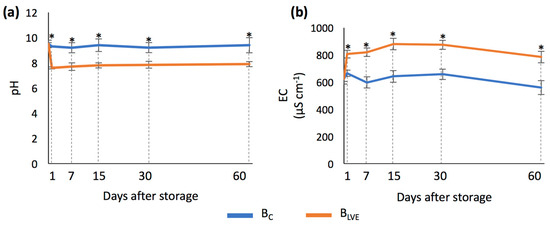
Figure 3.
(a) pH (mean ± SD) of BC and BLVE across the different days after storage; (b) electrical conductivity (EC) (mean ± SD) of BC and BLVE across the different days after storage. Asterisks indicate significant differences between BC and BLVE each day after storage.
3.4. Testing the Effect of Increasing Doses on Soil Quality
3.4.1. Carbon Fractions and BFI
In both soils, the M-S and W-S, the application of the BC and BLVE treatments resulted in an increase in the SOM. Regardless of the concentration tested, the average increase recorded in the M-S was 69% for both the BC and BLVE treatments (Table 3). The W-S showed an average increase of 128% for the BC treatments and 134% for the BLVE treatments (Table 4).
Table 3.
Soil fertility parameters comprising the soil biological fertility index (BFI) and its level (BFI lv.) of M-S (mean ± SD; n = 6). Means with the same letters within the column are not significantly different from each other at p < 0.05 according to Tukey-Kramer test.
Table 4.
Soil fertility parameters comprising the soil biological fertility index (BFI) and its level (BFI lv.) of W-S (mean ± SD; n = 6). Means with the same letters within the column are not significantly different from each other at p < 0.05 according to Tukey-Kramer test.
The different soil types had varying effects on the microbial biomass carbon (MB-C) based on the treatment used. In the M-S, the BC alone treatment increased the MB-C by an average of 18%, while the LVE alone treatment showed positive effects only at the M and H dose with an average increase of 12%. On the other hand, the BLVE treatment showed a general increase of 32% (Table 3). In the W-S, only the highest dose showed statistically significant effects, with the BC increasing by 18%, LVE by 19%, and BLVE by 30% (Table 4).
No statistical differences were found between individual treatments in the soil Res_Bas. However, a significant increase in the parameter was observed in the BLVE treatment for both the M-S and W-S. The average increase was 21% in the M-S (Table 3) and 26% in the W-S, but only for the M and H doses (Table 4). As for the Res_Bas, the soil Res_Cum followed a similar trend for both analyzed soils. It was an increase in the M-S at the highest dose of BC (+8%), and for the M and H doses of BLVE (+23% on average, Table 3). In the W-S, this increase is only observed for the H-BLVE, with an increase of 47% (Table 4).
The trend of the qCO2 varied depending on the soil type and treatments. In the case of the M-S, there was a general decrease in the BC-based treatments with a reduction of 15% for the BC and 8% for the BLVE, and a decrease of 7% for the M and H doses of the LVE (Table 3). As for the W-S, only the BC-based treatments showed a significant difference and had opposite trends compared to the Cnt, with a decrease of 13% for the H-BC, and an increase of 6% for the L and M doses of BLVE (Table 4). The trend of the qM is also dose-dependent, unlike qCO2. As reported in Table 3 for the M-S and in Table 4 for the W-S, in both soils, there was a decrease observed for the M-BC and H-BC doses (−20% in M-S and −19% in W-S) and LVE-based treatments (−35% in M-S and −52% in W-S). On the other hand, for the BLVE treatments, negative results were obtained for the L-BLVE dose (−47% in M-S and −60% in W-S), while an increase of 37% and 41% for each soil was observed for the M and H doses.
The final calculation of the BFI and the corresponding levels indicated that treatments based on the BC and BLVE positively influenced soil fertility. For the M-S, the BC and BLVE treatments increased the BFI by 17% and 33%, respectively (Table 3), while for the W-S, the H-BC dose increased the BFI by 25% and an average of 31% for the M and H doses of the BLVE (Table 4). A potential synergistic effect may occur between the BLVE treatments at the M and H doses in both soils.
3.4.2. Enzyme Activities and SAI3
The enzyme activities showed an overall positive response to both the individual and combined treatments (Table 5). Compared to the Cnt, the phosphatase activity increased in both soils with the application of increasing doses of BC (+32%), LVE (+24%), and BLVE (+33%). In the M-S, the activity of urease was increased by 18% for the BC and 17% for the LVE. However, when amended (BLVE), the increase became more significant (+22%). In the W-S, only the BC- and BLVE-based treatments showed a substantial increase, with 44% for the BC and 129% for the BLVE. Out of all the enzymes analyzed, β-glucosidase showed a different trend. This enzyme activity decreased by 16% and 17% in both the M-S and W-S when treated with the BC-based doses, and by 18% and 12% when treated with the BLVE. Only in the M-S was there a decrease of 22% in enzyme activity when treated with the LVE. Despite the decreasing trend of β-glucosidase, the SAI3 calculation showed an overall improvement in soil quality as the doses increased. Regarding any synergistic effect between the materials in the BLVE, it was only observed in the M-S at the highest dose (H). However, in the W-S, differences were visible at the M dose compared to the other treatments.
Table 5.
Soil quality parameters comprising the soil alteration index 3 (SAI3) of M-S and W-S (mean ± SD; n = 6). Means with the same letters within the column are not significantly different from each other at p < 0.05 according to Tukey-Kramer test.
4. Discussion
4.1. Potential Properties of LVE-Activated Biochar
The biochar activation method used allowed the efficient colonization and active multiplication of the LVE bacteria in biochar particles. This effect was clearly depicted by the 10-fold increase in bacterial abundance in the BLVE during the first week of storage and by the maintenance of these population levels for at least 60 days of storage. On the other hand, the observed changes in community composition across biochar storage indicated that taxa lying outside detectability thresholds in pure LVE can later prove to be advantaged upon biochar colonization, as observed for Bervundimonas sp. BvLVE, Brucella sp. BrLVE, and Priestia sp. PrLVE. The colonization of biochar by these taxa could contribute to increasing the microbial diversity in the soil upon amendment with LVE-activated biochar, which could further enhance nutrient availability through nutrient mineralization and organic matter decomposition [48]. Nevertheless, it is important to consider that the specific consequences of amending soils with activated biochar can vary depending on factors such as the type of biochar, the composition of the soil, and the specific microorganisms involved. In addition, although the 16S rRNA gene can provide poor discriminatory power at the species level, the top 100 blast hits for the first two genera showed a high correspondence to the species Brevundimonas bullata and Priestia aryabbhatai, respectively. Studies correlating the presence of B. bullata with improved soil quality are missing. However, Li et al. [49] characterized the antagonistic activity of this species against plant pathogenic nematodes. Furthermore, P. aryabhattai has been shown to significantly promote plant growth and drought tolerance in different plant species, including Arabidopsis, tobacco, and maize [50,51] and to have phosphate-solubilizing and N-fixing roles, which may potentially contribute to nutrient availability and uptake by plants [52]. Conversely, other bacteria present in LVE may gradually decrease their abundance in biochar particles, as observed for Gordonia sp. GdLVE, Bacillus sp. Bc3LVE, and Bacillus sp. Bc4LVE. A possible reason for the decrease in these taxa may be linked to functional redundancy with other phylogenetically close members and/or faster-growing microorganisms present in LVE, or even to antagonistic interactions among specific taxa.
Another explanation for the observed changes in community composition over time could lie in the varying ability of the isolated bacteria to persist in biochar particles. In this regard, the top 100 blast hits for Leuconostoc sp. LcLVE, Bacillus sp. Bc1LVE, and Bacillus sp. Bc2LVE showed a high correspondence to the species Leuconostoc mesenteroides, Bacillus subtilis, and Bacillus cereus, which are well known for their capability to secrete extracellular polymeric substances that attach to surfaces and promote biofilm formation [53,54,55]. However, although biochar can potentially serve as a substrate for biofilms, further research is needed on the role of biofilm formation in enhancing bacterial survival in biochar. In addition, it is reasonable to assume that microbial communities may experience some degree of fluctuation in structure due to their high metabolic activity; this can occur particularly in OM-rich environments, which can induce a more active production of microbial metabolites that can ultimately affect the overall community structure [56,57].
Finally, activation with the LVE led to a significant decrease in the pH of the biochar and an increase in its EC. As for the pH, this effect may have positive consequences on plant growth, as the alkalinity of biochar can lead to the unavailability of assimilable forms of nutrients to plants [58]. On the other hand, the increase in EC could indicate LVE as an additional source of soluble salts, which could compete in uptake with the availability of important nutrients, leading the plant to a nutritional and metabolic imbalance and decreased or increased cell osmotic potential. However, the potential impacts of increased EC on the measured soil quality parameters remain to be elucidated.
4.2. Validation of LVE-Activated Biochar on Soil
4.2.1. Effects on Linked C Parameters and Biological Fertility of Soil
As expected, the addition of Bc and BLVE to the soil increased the SOM content in both soil types (i.e., M-S and W-S), with a greater dose leading to a higher increase. This agrees with other studies [59,60], which have shown that the use of organic fertilizers such as biochar and composted materials (i.e., vermicompost) can enhance the SOM. This is primarily due to biochar contributing stable organic carbon to the soil [61].
Most studies suggest that biochar applications increase the MB-C, due to its ability to create a favorable habitat for microorganisms [62]. In this study, the addition of BC and BLVE modified the biological activity in the soils tested and it was observed that the positive impact of Bc was more evident at higher doses and in the M-S. This could be attributed to the fact that the M-S had a better initial condition of biological fertility in terms of the SOM and MB-C, with respect to the W-S. Other research also supported the ability of high doses of Bc to enhance the MB-C in soils that share similar characteristics to the M-S [63,64]. The higher MB-C values in the BLVE-treated soils, than Bc-treated ones, suggest that the LVE and/or soil microorganisms could be increased by the biochar. On the other hand, several studies demonstrated the ability of vermicompost to increase the MB-C [65,66,67] and this would also explain the positive effect of the high dose of LVE treatment in both soils.
The addition of the BLVE increased the soil respiration parameters (i.e., Res_Cum and Res_Bas) in both soils and at higher application doses, indicating a potential role of LVE bacteria in the promotion of a higher and more efficient soil respiration process. Some studies show that biochar does not increase soil respiration [68,69], while LVE can improve it [65,70]. The increased effect of the BLVE on soil respiration could be attributed to (1) the activity of inoculated microorganisms on the native SOM and (2) the ability of the biochar to promote the activity of both inoculated and native microorganisms.
The qCO2 measures the respiration rate of CO2-C per unit of MB-C [71]. It is widely used as an indicator of ecosystem succession [72], during which it is expected to decline, and maturity [73]. It has been seen that a low qCO2 value indicates that soil conditions were improved by the addition of organic matter to soil [74]. Overall, here, the treatments contributed to a reduction in the qCO2 in both soils. However, the reduction was more significant in the M-S than in the W-S. This study confirms that the use of biochar, vermicompost, or a combination of both can reduce this parameter and this observation is in line with other similar studies [64,75,76]. The different trend observed between the two soils could be attributed to the lower amount of native SOM in the W-S compared to the abundance of microorganisms.
The qM represents the TOC fraction that has been mineralized throughout an incubation period [77]. Biochar-based treatments substantially reduced this parameter in both soils, confirming the results of previous studies [78,79] and qualifying it as an effective tool for increasing C storage in the soil.
The BFI is a robust multi-domain indicator of fertility developed for Italian soils [37]. In the BLVE treatments, the biochar and LVE showed a mutual enhancement of their properties, resulting in an increase in the BFI level in both soils, hence reflecting an improved biological activity and soil quality. Although the effect was observable in the M-S, it was more pronounced in the W-S. Indeed, even with the lowest dose of BLVE, there was a significant increase in the BFI in the W-S.
4.2.2. Effects on Enzyme Activities and Quality of Soils
In general, the different treatments positively influenced the APA and UA at various doses, but a negative trend for the βGA was observed. Although in other studies [80,81,82,83] the biochar had positive effects on the UA and APA, in our case, the Bc treatments led to a decrease in the βGA. This is in line with Gunal et al. [84], who found that the addition of biochar reduced the βGA and the decline was higher in sandy loam soils compared to loamy soils. Foster et al. [85] suggest that the surface area and pore size distribution of biochar are key indicators of potential biochar-enzyme interactions. They assume that the decline in the βGA is mainly a result of direct absorption. At the same time, adsorption of a small amount of substrate can also occur, since the polar glucose substrate can easily adsorb onto negatively charged solid phases. Other factors may also have influenced the βGA, including the presence in the biochar of inhibiting compounds such as phenols and polyphenols.
In general, vermicompost-based treatments enhance soil enzyme activities and our results align with those of other researchers [19,86,87]. However, it is possible that the decrease in the βGA, as a result of the LVE treatment in the M-S, was due to a reduction in the enzyme present in the vermicompost used to produce the extract. Previous studies have shown that the βGA in the vermicompost decreases as it matures [88,89]. This decrease can be attributed to the presence of more humified organic substances and a reduction in cellobiose and other disaccharides. It is possible that LVE-based treatments have altered the composition of the SOM, thereby reducing the need to produce this enzyme by microorganisms.
Although the βGA showed a negative trend, the SAI3 revealed that the soil quality increased with increasing doses of the BLVE-based treatments in both soils. It is interesting to note that even the lowest dose showed a clear improvement, indicating the magnification of the interaction between the biochar and inoculated microorganisms.
Moreover, the observed changes in the soil enzyme activities may partly lie on the identity and metabolic attributes of the bacteria present in the activated biochar. Microbial APA has often been recognized as a plant- and soil-promoting attribute due to its role in making P more accessible from the SOM into more available forms [90]. On the other hand, microbial UA is a key component of soil biological activity, since it promotes nutrient cycling and the enhancement of N in the soil, and thus contributes to the maintenance of soil health and fertility [91]. In this regard, several members of the phylum Bacillota, including B. subtilis, B. cereus, and P. aryabhattai, are known for their high APA and UA [92,93,94,95]. Moreover, other identified bacterial species from the phylum Pseudomonadota, including B. bullata as well as several Brucella and Pseudomonas species, can produce diverse types of phosphatases [93,96,97,98,99,100] and ureases [101,102].
5. Conclusions
The present study showed that biochar can be successfully activated with LVE and that LVE-associated bacteria can undergo compositional shifts across biochar storage. However, while the presence of LVE bacteria could be correlated to the observed beneficial effects on soil quality, further characterization of these isolates is necessary to allow their safe application in agriculture, including the evaluation of potential ecological impacts, the assessment of potential risks of horizontal gene transfer, and the examination of potential side-effects on non-target organisms resulting from the introduction of non-native bacteria into the environment. Moreover, to better understand the role of vermicompost bacteria in soil improvement, further genetic analysis, metabolic profiling, and functional assessments are needed. These additional investigations could provide insights into the genetic composition and metabolic capabilities of the bacteria, as well as their specific functions in enhancing soil health and nutrient cycling. It is important to conduct these studies to gain a comprehensive understanding of the mechanisms underlying the beneficial properties of vermicompost bacteria and to address potential ecological impacts, risks of horizontal gene transfer, and effects on non-target organisms.
In addition, the use of LVE-activated biochar significantly enhanced the soil parameters of C-related soil biological fertility and enzyme-linked soil quality. This study found that activated biochar can significantly improve soil quality and fertility, particularly in less fertile soil (i.e., W-S). However, the increase was also substantial in medium-fertile soils (i.e., M-S), making activated biochar a highly valuable tool for enhancing soil quality and fertility. By incorporating LVE-activated biochar into soil management practices, farmers could achieve higher yields and healthier crops.
However, it is necessary to conduct more comprehensive research to confirm the effectiveness of this approach, considering different types of soils with different physical, chemical, and biological properties. Furthermore, it would be essential to conduct long-term respiration tests and a BLVE storage period in various environmental conditions on other biochars and LVEs produced using different matrices and methods.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/soilsystems8020054/s1, Table S1: list of bacteria isolated from the liquid vermicompost extract (LVE) (genus and assigned strain name), primers used for amplification, and respective accession numbers.
Author Contributions
Conceptualization, P.C. and M.B.; methodology, P.C. and M.B.; software, P.C. and M.B.; validation, P.C. and M.B.; formal analysis, P.C. and M.B.; investigation, P.C. and M.B.; data curation, P.C. and M.B.; writing—original draft preparation, P.C., M.B. and S.C.; writing—review and editing, P.C., M.B., S.C., S.L. and R.C.; visualization, R.F.; supervision, S.C., S.L. and R.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data and materials can be made available upon reasonable request by the corresponding author.
Acknowledgments
We appreciate the support from Bio-Esperia S.r.l. (Arezzo, Italy) and the Centro di Lombricoltura Toscano S.r.l. (Pisa, Italy), for having produced and provided the biochar and the liquid vermicompost extracts, respectively. We are also grateful to C.I.B.I.A.C.I. (Centro Interdipartimentale di Servizi per le Biotecnologie di interesse Agrario, Chimico, Industriale) (University of Florence) for the sequencing service and to Priscilla Bettini (Biology Department, University of Florence, Sesto Fiorentino) for the provision of the infrastructures for the biochar activation process. We thank also the project funded under the National Recovery and Resilience Plan (NRRP), Mission 4 Component 2 Investment 1.4—Call for tender No. 3138 of 16 December 2021, rectified by Decree n.3175 of 18 December 2021 of Italian Ministry of University and Research funded by the European Union—NextGenerationEU. Project code CN_00000033, Concession Decree No. 1034 of 17 June 2022 adopted by the Italian Ministry of University and Research, CUP B63C22000650007, Project title “National Biodiversity Future Center—NBFC”.
Conflicts of Interest
We declare no known competing financial and non-financial interests concerning the current research. The producers of the biochar and liquid vermicompost extracts had no role in the study design, nor in the collection, analyses, and/or interpretation of the data, nor in the writing of the manuscript, nor in the decision to publish the results.
References
- Sanchez-Hernandez, J.C.; Ro, K.S.; Díaz, F.J. Biochar and earthworms working in tandem: Research opportunities for soil bioremediation. Sci. Total Environ. 2019, 688, 574–583. [Google Scholar] [CrossRef] [PubMed]
- Nyambo, P.; Zhou, L.; Chuma, T.; Sokombela, A.; Malobane, M.E.; Musokwa, M. Prospects of Vermicompost and Biochar in Climate Smart Agriculture. In Vermicomposting for Sustainable Food Systems in Africa; Springer Nature: Singapore, 2023; pp. 145–159. [Google Scholar]
- Álvarez Bernal, D.; Lastiri Hernández, M.A.; Buelna Osben, H.R.; Contreras Ramos, S.M.; Mora, M. Vermicompost as an alternative of management for water hyacinth. Rev. Int. Contam. 2016, 32, 425–433. [Google Scholar] [CrossRef]
- Koskey, G.; Avio, L.; Turrini, A.; Sbrana, C.; Bàrberi, P. Biostimulatory effect of vermicompost extract enhances soil mycorrhizal activity and selectively improves crop productivity. Plant Soil 2023, 484, 183–199. [Google Scholar] [CrossRef]
- Choudhary, R.C.; Bairwa, H.L.; Kumar, U.; Javed, T.; Asad, M.; Lal, K.; Abdelsalam, N.R. Influence of organic manures on soil nutrient content, microbial population, yield and quality parameters of pomegranate (Punica granatum L.) cv. Bhagwa. PLoS ONE 2022, 17, e0266675. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Jha, G. Utilization of microbiome to develop disease resistance in crop plants against phytopathogens. Front. Plant Sci. 2023, 14, 1204896. [Google Scholar] [CrossRef] [PubMed]
- Karhu, K.; Mattila, T.; Bergström, I.; Regina, K. Biochar addition to agricultural soil increased CH4 uptake and water holding capacity—Results from a short-term pilot field study. Agric. Ecosyst. Environ. 2011, 140, 309–313. [Google Scholar] [CrossRef]
- Qayyum, M.F.; Haider, G.; Iqbal, M.; Hameed, S.; Ahmad, N.; ur Rehman, M.Z.; Ali, S. Effect of alkaline and chemically engineered biochar on soil properties and phosphorus bioavailability in maize. Chemosphere 2021, 266, 128980. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Fonte, S.J.; Parikh, S.J.; Six, J.; Scow, K.M. Biochar additions can enhance soil structure and the physical stabilization of C in aggregates. Geoderma 2017, 303, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Bolan, S.; Hou, D.; Wang, L.; Hale, L.; Egamberdieva, D.; Tammeorg, P.; Bolan, N. The potential of biochar as a microbial carrier for agricultural and environmental applications. Sci. Total Environ. 2023, 886, 163968. [Google Scholar] [CrossRef]
- Shi, X.; Zhou, G.; Liao, S.; Shan, S.; Wang, G.; Guo, Z. Immobilization of cadmium by immobilized Alishewanella sp. WH16-1 with alginate-lotus seed pods in pot experiments of Cd-contaminated paddy soil. J. Hazard. Mater. 2018, 357, 431–439. [Google Scholar] [CrossRef]
- Quaik, S.; Embrandiri, A.; Rupani, P.F.; Ibrahim, M.H. Potential of vermicomposting leachate as organic foliar fertilizer and nutrient solution in hydroponic culture: A review. In Proceedings of the 2nd International Conference on Environment and BioScience IPCBEE, Phnom Penh, Cambodia, 28–29 September 2012; IACSIT Press: Singapore, 2012; Volume 44, pp. 43–47. [Google Scholar]
- Brtnicky, M.; Datta, R.; Holatko, J.; Bielska, L.; Gusiatin, Z.M.; Kucerik, J.; Pecina, V. A critical review of the possible adverse effects of biochar in the soil environment. Sci. Total Environ. 2021, 796, 148756. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, J.D.; Chaves, L.H.; Mendes, J.S.; Chaves, I.B.; Tito, G.A. Alterations in soil salinity with the use of different biochar doses. Rev. Ciênc. 2019, 42, 89–98. [Google Scholar]
- Jahan, S.; Ahmad, F.; Rasul, F.; Amir, R.; Shahzad, S. Physicochemical Analysis of Vermicompost-Perlite Based Activated Biochar and its Influence on Wheat (Triticum aestivum L.) Growth Under Water Stress. J. Soil Sci. Plant Nutr. 2023, 23, 3034–3050. [Google Scholar] [CrossRef]
- Karki, R.; Solberg, S.Ø. Activated BIOCHAR with VERMICOMPOST as an alternative PEAT substitute. Int. J. Biosci. Healthc. Agric. Technol. 2018, 9, 64. [Google Scholar]
- Yuvaraj, A.; Thangaraj, R.; Karmegam, N.; Ravindran, B.; Chang, S.W.; Awasthi, M.K.; Kannan, S. Activation of biochar through exoenzymes prompted by earthworms for vermibiochar production: A viable resource recovery option for heavy metal contaminated soils and water. Chemosphere 2021, 278, 130458. [Google Scholar] [CrossRef] [PubMed]
- Becagli, M.; Guglielminetti, L.; Cardelli, R. Effects of Combined Biochar and Vermicompost Solution on Leachate Characterization and Nitrogen Balance from a Greenhouse Tomato (Solanum lycopersicum) Cultivation Soil. Commun. Soil Sci. Plant Anal. 2021, 52, 1879–1893. [Google Scholar] [CrossRef]
- Becagli, M.; Arduini, I.; Cardelli, R. Using Biochar and Vermiwash to Improve Biological Activities of Soil. Agriculture 2022, 12, 178. [Google Scholar] [CrossRef]
- Abbey, L.; Cai, J.; Gunupuru, L.R.; Ijenyo, M.; Esan, E.O.; Lin, S. Nutrient Release Pattern and Greenhouse-Grown Swiss Chard Response to Biochar Inoculated with Vermicast. Int. J. Agron. 2020, 2020, 7852187. [Google Scholar] [CrossRef]
- Dura, O.; Dura, S.; Kepenekci, H. Nematicidal potential of vermicompost tea against Meloidogyne incognita on tomato. In Proceedings of the IV. Balkan Agricultural Congress, Edirne, Turkey, 31 August–2 September 2022; pp. 742–748. [Google Scholar]
- Edenborn, S.L.; Johnson, L.M.; Edenborn, H.M.; Albarran-Jack, M.R.; Demetrion, L.D. Amendment of a hardwood biochar with compost tea: Effects on plant growth, insect damage and the functional diversity of soil microbial communities. Biol. Agric. Hortic. 2018, 34, 88–106. [Google Scholar] [CrossRef]
- Mondal, S.; Ghosh, S.; Mukherjee, A. Application of biochar and vermicompost against the rice root-knot nematode (Meloidogyne graminicola): An eco-friendly approach in nematode management. J. Plant Dis. Prot. 2011, 128, 819–829. [Google Scholar] [CrossRef]
- Brtnicky, M.; Mustafa, A.; Hammerschmiedt, T.; Kintl, A.; Trakal, L.; Beesley, L.; Holatko, J. Pre-activated biochar by fertilizers mitigates nutrient leaching and stimulates soil microbial activity. Chem. Biol. Technol. Agric. 2023, 10, 57. [Google Scholar] [CrossRef]
- Schommer, V.A.; Nazari, M.T.; Melara, F.; Braun, J.C.A.; Rempel, A.; Dos Santos, L.F.; Piccin, J.S. Techniques and mechanisms of bacteria immobilization on biochar for further environmental and agricultural applications. Microbiol. Res. 2023, 278, 127534. [Google Scholar] [CrossRef] [PubMed]
- Fritz, J.I.; Franke-Whittle, I.H.; Haindl, S.; Insam, H.; Braun, R. Microbiological community analysis of vermicompost tea and its influence on the growth of vegetables and cereals. Can. J. Microbiol. 2012, 58, 836–847. [Google Scholar] [CrossRef] [PubMed]
- Ke, Y.; Si, S.; Zhang, Z.; Geng, P.; Shen, Y.; Wang, J.; Zhu, X. Synergistic passivation performance of cadmium pollution by biochar combined with sulfate reducing bacteria. Environ. Technol. Innov. 2023, 32, 103356. [Google Scholar] [CrossRef]
- Ouyang, P.; Narayanan, M.; Shi, X.; Chen, X.; Li, Z.; Luo, Y.; Ma, Y. Integrating biochar and bacteria for sustainable remediation of metal-contaminated soils. Biochar 2023, 5, 63. [Google Scholar] [CrossRef]
- Xiao, Y.; Zheng, L.; Wang, S.; Dong, M.; Gao, A.; Han, Z.; Zhang, H. Bioremediation of metribuzin-contaminated soil by corn straw biochar-immobilized Bacillus cereus N1. Process Biochem. 2023, 130, 520–533. [Google Scholar] [CrossRef]
- International Biochar Initiative. 2024. Available online: http://www.european-biochar.org/en/ebc-ibi (accessed on 11 January 2024).
- Hristov, A.N.; Callaway, T.R.; Lee, C.; Dowd, S.E. Rumen bacterial, archaeal, and fungal diversity of dairy cows in response to ingestion of lauric or myristic acid. J. Anim. Sci. 2012, 90, 4449–4457. [Google Scholar] [CrossRef] [PubMed]
- Lane, D.J. 16S/23S rRNA sequencing. In Nucleic Acid Techniques in Bacterial Systematics; Wiley: Hoboken, NJ, USA, 1991; pp. 115–175. [Google Scholar]
- Celletti, S.; Bergamo, A.; Benedetti, V.; Pecchi, M.; Patuzzi, F.; Basso, D.; Baratieri, M.; Cesco, S.; Mimmo, T. Phytotoxicity of hydrochars obtained by hydrothermal carbonization of manure-based digestate. J. Environ. Manag. 2021, 280, 111635. [Google Scholar] [CrossRef]
- Colombo, C.; Miano, T. Metodi di Analisi Chimica del Suolo; Pubblicità & Stampa: Modugno, BA, USA, 2015. [Google Scholar]
- Levi-Minzi, R.; Riffaldi, R.; Saviozzi, A. Carbon mineralization in soil amended with different organic materials. Agric. Ecosyst. Environ. 1990, 31, 325–335. [Google Scholar] [CrossRef]
- Vance, E.D.; Brookes, C.; Jenkinson, D.S. An extraction method for measuring soil microbial biomass carbon. Soil Biol. Biochem. 1987, 19, 403–707. [Google Scholar] [CrossRef]
- Renzi, G.; Canfora, L.; Salvati, L.; Benedetti, A. Validation of the soil Biological Fertility Index (BFI) using a multidimensional statistical approach: A country-scale exercise. Catena 2017, 149, 294–299. [Google Scholar] [CrossRef]
- Andersen, C.P. Source-sink balance and carbon allocation below ground in plants exposed to ozone. New Phytol. 2003, 157, 213–228. [Google Scholar] [CrossRef] [PubMed]
- Landi, L.; Renella, G.; Moreno, J.L.; Falchini, L.; Nannipieri, P. Influence of cadmium on the metabolic quotient, L-: D-glutamic acid respiration ratio and enzyme activity: Microbial biomass ratio under laboratory conditions. Biol. Fertil. Soils 2000, 32, 8–16. [Google Scholar] [CrossRef]
- Nannipieri, P.; Ascher, J.; Ceccherini, M.; Landi, L.; Pietramellara, G.; Renella, G. Microbial diversity and soil functions. Eur. J. Soil Sci. 2003, 54, 655–670. [Google Scholar] [CrossRef]
- Sequi, P.; Benedetti, A.; Dell’Abate, M.T.; ATLAS. Atlante di Indicatori della Qualità del Suolo; Franco Angeli: Milano, Italy, 2006. [Google Scholar]
- Brookes, P.C. The use of microbial parameters inmonitoring soil pollution by heavy metals. Biol. Fertil. Soils 1995, 19, 269–279. [Google Scholar] [CrossRef]
- Bloem, J.; Benedetti, A.; Hopkins, D. Microbial Methods Assessing Soil Quality; CABI: London, UK, 2006. [Google Scholar]
- Eivazi, F.; Tabatabai, M.A. Phosphatases in soil. Soil Biol. Biochem. 1997, 9, 167–172. [Google Scholar] [CrossRef]
- Eivazi, F.; Tabatabai, M.A. Glucosidases and galactosidases in soils. Soil Biol. Biochem. 1988, 20, 601–606. [Google Scholar] [CrossRef]
- Kandeler, E.; Gerber, H. Short-term assay of soil urease activity using colorimetric determination of ammonium. Biol. Fertil. Soils 1988, 6, 68–72. [Google Scholar] [CrossRef]
- Puglisi, E.; Del Re, A.A.M.; Rao, M.A.; Gianfreda, L. Development and validation of numerical indexes integrating enzyme activities of soils. Soil Biol. Biochem. 2006, 38, 1673–1681. [Google Scholar] [CrossRef]
- Maron, P.A.; Sarr, A.; Kaisermann, A.; Lévêque, J.; Mathieu, O.; Guigue, J.; Ranjard, L. High microbial diversity promotes soil ecosystem functioning. Appl. Environ. Microbiol. 2018, 84, e02738-17. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Ding, M.; Sun, X.; Li, Z.; Xu, L.; Li, L. Characterization of Nematicidal Activity and Nematode-Toxic Metabolites of a Soilborne Brevundimonas bullata Isolate. Pathogens 2022, 11, 708. [Google Scholar] [CrossRef] [PubMed]
- Castelo Sousa, H.; Gomes de Sousa, G.; de Araújo Viana, T.V.; Prudêncio de Araújo Pereira, A.; Nojosa Lessa, C.I.; Pires de Souza, M.V.; Barbosa da Silva, F.D. Bacillus aryabhattai Mitigates the Effects of Salt and Water Stress on the Agronomic Performance of Maize under an Agroecological System. Agriculture 2023, 13, 1150. [Google Scholar] [CrossRef]
- Xu, H.; Gao, J.; Portieles, R.; Du, L.; Gao, X.; Borras-Hidalgo, O. Endophytic bacterium Bacillus aryabhattai induces novel transcriptomic changes to stimulate plant growth. PLoS ONE 2022, 17, e0272500. [Google Scholar] [CrossRef] [PubMed]
- Deng, C.; Zhang, N.; Liang, X.; Huang, T.; Li, B. Bacillus aryabhattai LAD impacts rhizosphere bacterial community structure and promotes maize plant growth. J. Sci. Food Agric. 2022, 102, 6650–6657. [Google Scholar] [CrossRef] [PubMed]
- Dergham, Y.; Sanchez-Vizuete, P.; Le Coq, D.; Deschamps, J.; Bridier, A.; Hamze, K.; Briandet, R. Comparison of the genetic features involved in Bacillus subtilis biofilm formation using multi-culturing approaches. Microorganisms 2021, 9, 633. [Google Scholar] [CrossRef]
- Leathers, T.D.; Bischoff, K.M. Biofilm formation by strains of Leuconostoc citreum and L. mesenteroides. Biotechnol. Lett. 2011, 33, 517–523. [Google Scholar] [CrossRef]
- Majed, R.; Faille, C.; Kallassy, M.; Gohar, M. Bacillus cereus biofilms—Same, only different. Front. Microbiol. 2016, 7, 1054. [Google Scholar] [CrossRef]
- Blanchet, M.; Pringault, O.; Bouvy, M.; Catala, P.; Oriol, L.; Caparros, J.; Joux, F. Changes in bacterial community metabolism and composition during the degradation of dissolved organic matter from the jellyfish Aurelia aurita in a Mediterranean coastal lagoon. Environ. Sci. Pollut. Res. 2015, 22, 13638–13653. [Google Scholar] [CrossRef]
- Gowda, K.; Ping, D.; Mani, M.; Kuehn, S. Genomic structure predicts metabolite dynamics in microbial communities. Cell 2022, 185, 530–546. [Google Scholar] [CrossRef]
- Fornes, F.; Belda, R.M.; Fernández de Córdova, P.; Cebolla-Cornejo, J. Assessment of biochar and hydrochar as minor to major constituents of growing media for containerized tomato production. J. Sci. Food Agric. 2017, 97, 3675–3684. [Google Scholar] [CrossRef]
- Agegnehu, G.; Bass, A.M.; Nelson, P.N.; Bird, M.I. Benefits of biochar, compost and biochar-compost for soil quality, maize yield and greenhouse gas emissions in a tropical agricultural soil. Sci. Total Environ. 2016, 543, 295–306. [Google Scholar] [CrossRef]
- Plaza, C.; Giannetta, B.; Fernández, J.M.; López-de-Sá, E.G.; Polo, A.; Gascó, G.; Méndez, A.; Zaccone, C. Response of different soil organic matter pools to biochar and organic fertilizers. Agric. Ecosyst. Environ. 2016, 225, 150–159. [Google Scholar] [CrossRef]
- Gross, A.; Bromm, T.; Glaser, B. Soil organic carbon sequestration after biochar application: A global meta-analysis. Agronomy 2021, 11, 2474. [Google Scholar] [CrossRef]
- Lehmann, J.; Rillig, M.C.; Thies, J.; Masiello, C.A.; Hockaday, W.C.; Crowley, D. Biochar effects on soil biota—A review. Soil Biol. Biochem. 2011, 43, 1812–1836. [Google Scholar] [CrossRef]
- Lo Piccolo, E.; Becagli, M.; Lauria, G.; Cantini, V.; Ceccanti, C.; Cardelli, R.; Massai, R.; Remorini, D.; Guidi, L.; Landi, M. Biochar as a soil amendment in the tree establishment phase: What are the consequences for tree physiology, soil quality and carbon sequestration? Sci. Total Environ. 2022, 844, 157175. [Google Scholar] [CrossRef]
- Zhou, H.; Zhang, D.; Wang, P.; Liu, X.; Cheng, K.; Li, L.; Zheng, J.; Zhang, X.; Zheng, J.; Crowley, D.; et al. Changes in microbial biomass and the metabolic quotient with biochar addition to agricultural soils: A Meta-analysis. Agric. Ecosyst. Environ. 2017, 239, 80–89. [Google Scholar] [CrossRef]
- Moridi, A.; Zarei, M.; Moosavi, A.A.; Ronaghi, A. Effect of liquid organic fertilizers and soil moisture status on some biological and physical properties of soil. Polish J. Soil Sci. 2021, 54, 41–58. [Google Scholar] [CrossRef]
- Sabir, M.S.; Shahzadi, F.; Ali, F.; Shakeela, Q.; Niaz, Z.; Ahmed, S. Comparative Effect of Fertilization Practices on Soil Microbial Diversity and Activity: An Overview. Curr. Microbiol. 2021, 78, 3644–3655. [Google Scholar] [CrossRef]
- Wu, Q.; Zhang, J.; Liu, X.; Chang, T.; Wang, Q.; Shaghaleh, H.; Hamoud, Y.A. Effects of biochar and vermicompost on microorganisms and enzymatic activities in greenhouse soil. Front. Environ. Sci. 2023, 10, 1060277. [Google Scholar] [CrossRef]
- Ge, X.; Cao, Y.; Zhou, B.; Wang, X.; Yang, Z.; Li, M.H. Biochar addition increases subsurface soil microbial biomass but has limited effects on soil CO2 emissions in subtropical moso bamboo plantations. Appl. Soil Ecol. 2019, 142, 155–165. [Google Scholar] [CrossRef]
- Weng, Z.H.; Van Zwieten, L.; Singh, B.P.; Kimber, S.; Morris, S.; Cowie, A.; Macdonald, L.M. Plant-biochar interactions drive the negative priming of soil organic carbon in an annual ryegrass field system. Soil Biol. Biochem. 2015, 90, 111–121. [Google Scholar] [CrossRef]
- Pant, A.P.; Radovich, T.J.K.; Hue, N.V.; Miyasaka, S.C. Pak Choi (Brassica rapa, Chinensis group) yield, phytonutrient content, and soil biological properties as affected by vermicompost-to-water ratio used for extraction. HortScience 2012, 47, 395–402. [Google Scholar] [CrossRef]
- Anderson, T.H.; Domsch, K.H. The metabolic quotient for CO2 (qCO2) as a specific activity parameter to assess the effects of environmental conditions, such as pH, on the microbial biomass of forest soils. Soil Biol. Biochem. 1993, 25, 393–395. [Google Scholar] [CrossRef]
- Insam, H.; Haselwandter, K. Metabolic quotient of the soil microflora in relation to plant succession. Oecologia 1989, 79, 174–178. [Google Scholar] [CrossRef] [PubMed]
- Anderson, T.H.; Domsch, K.H. Determination of ecophysiological maintenance carbon requirements of soil microorganisms in a dormant state. Biol. Fertil. Soils 1985, 1, 81–89. [Google Scholar] [CrossRef]
- Powlson, D.S.; Jenkinson, D.S. A comparison of the organic matter, biomass, adenosine triphosphate, and mineralizable nitrogen contents of ploughed and direct drilled soils. J. Agric. Sci. 1981, 97, 713–721. [Google Scholar] [CrossRef]
- Trivedi, P.; Singh, K.; Pankaj, U.; Verma, S.K.; Verma, R.K.; Patra, D.D. Effect of organic amendments and microbial application on sodic soil properties and growth of an aromatic crop. Ecol. Eng. 2017, 102, 127–136. [Google Scholar] [CrossRef]
- Yilmaz, F.I.; Kurt, S. The effects of biochar and vermicompost applications on some enzyme activities in rhizosphere root zone of corn (Zea mays L.) plant. Comptes Rendus L’académie Bulg. Sci. 2020, 73, 58–65. [Google Scholar]
- Moscatelli, M.C.; Lagomarsino, A.; Marinari, S.; De Angelis, P.; Grego, S. Soil microbial indices as bioindicators of environmental changes in a poplar plantation. Ecol. Indic. 2005, 5, 171–179. [Google Scholar] [CrossRef]
- Luis Moreno, J.; Bastida, F.; Díaz-López, M.; Li, Y.; Zhou, Y.; López-Mondéjar, R.; Benavente-Ferraces, I.; Rojas, R.; Rey, A.; Carlos García-Gil, J.; et al. Response of soil chemical properties, enzyme activities and microbial communities to biochar application and climate change in a Mediterranean agroecosystem. Geoderma 2022, 407, 115536. [Google Scholar] [CrossRef]
- Das, S.K. Soil carbon footprint, budgeting, and dynamics in a biomass conversion-based long-term organic production system. Biomass Convers. Biorefin. 2023. [Google Scholar] [CrossRef]
- Pokharel, P.; Ma, Z.; Chang, S.X. Biochar increases soil microbial biomass with changes in extra- and intracellular enzyme activities: A global meta-analysis. Biochar 2020, 2, 65–79. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, J.; Feng, Y. The effects of biochar addition on soil physicochemical properties: A review. Catena 2021, 202, 105284. [Google Scholar] [CrossRef]
- Joseph, S.; Cowie, A.L.; Van Zwieten, L.; Bolan, N.; Budai, A.; Buss, W.; Lehmann, J. How biochar works, and when it doesn’t: A review of mechanisms controlling soil and plant responses to biochar. Gcb Bioenergy 2021, 13, 1731–1764. [Google Scholar] [CrossRef]
- Ghodszad, L.; Reyhanitabar, A.; Maghsoodi, M.R.; Lajayer, B.A.; Chang, S.X. Biochar affects the fate of phosphorus in soil and water: A critical review. Chemosphere 2021, 283, 131176. [Google Scholar] [CrossRef]
- Günal, E.; Erdem, H.; Demirbaş, A. Effects of three biochar types on activity of β-glucosidase enzyme in two agricultural soils of different textures. Arch. Agron. Soil Sci. 2018, 64, 1963–1974. [Google Scholar] [CrossRef]
- Foster, E.J.; Fogle, E.J.; Cotrufo, M.F. Sorption to biochar impacts β-glucosidase and phosphatase enzyme activities. Agriculture 2018, 8, 158. [Google Scholar] [CrossRef]
- Jayaswal, K.; Christian, J.; Singh, N.K.; Padhiyar, H.; Yadav, M.; Sanghvi, G. Effect of vermicompost on soil quality parameters for different land use patterns. IOP Conf. Ser. Earth Environ. Sci. 2023, 1280, 012054. [Google Scholar] [CrossRef]
- Zuo, Y.; Zhang, J.; Zhao, R.; Dai, H.; Zhang, Z. Application of vermicompost improves strawberry growth and quality through increased photosynthesis rate, free radical scavenging and soil enzymatic activity. Sci. Hortic. 2018, 233, 132–140. [Google Scholar] [CrossRef]
- Fernández-Gómez, M.J.; Nogales, R.; Insam, H.; Romero, E.; Goberna, M. Continuous-feeding vermicomposting as a recycling management method to revalue tomato-fruit wastes from greenhouse crops. Waste Manag. 2010, 30, 2461–2468. [Google Scholar] [CrossRef]
- García-Sánchez, M.; Taušnerová, H.; Hanč, A.; Tlustoš, P. Stabilization of different starting materials through vermicomposting in a continuous-feeding system: Changes in chemical and biological parameters. Waste Manag. 2017, 62, 33–42. [Google Scholar] [CrossRef]
- Pan, L.; Cai, B. Phosphate-Solubilizing Bacteria: Advances in Their Physiology, Molecular Mechanisms and Microbial Community Effects. Microorganisms 2023, 11, 2904. [Google Scholar] [CrossRef]
- Koçak, B. Importance of urease activity in soil. In Proceedings of the International Scientific and Vocational Studies Congress—Science and Health (BILMES SH 2020), Ankara, Turkey, 12–15 December 2020. [Google Scholar]
- Aruna, S.; Manikandavelu, D.; Uma, A.; Antony, C.; Jayakumar, N. Assessment of phosphorus mobilizing capacity of Bacillus spp. isolated from mangrove rhizospheric sediment and its potential application in aquaculture. Indian J. Anim. Res. 2023, 57, 1506–1511. [Google Scholar] [CrossRef]
- Fitriatin, B.N.; Arief, D.H.; Simarmata, T.; Santosa, D.A.; Joy, B. Phosphatase-producing bacteria isolated from Sanggabuana forest and their capability to hydrolyze organic phosphate. J. Soil Sci. Environ. 2011, 2, 299–303. [Google Scholar]
- Hwang, T.K.; Park, Y.J.; Kim, M.J.; Park, M.K.; Kim, M.C.; Jung, M.; Shin, J.H. Isolation of Bacillus aryabhattai GW320 from the Cucumber Rhizospheric Soil and Evaluation of Plant Growth Promoting Activity. Korean J. Soil. Sci. Fert. 2021, 54, 126–139. [Google Scholar] [CrossRef]
- Parhamfar, M.; Badoei-Dalfard, A.; Parhamfar, M.; Fahimi Rad, S. Purification and Characterization of an Extracellular Phosphatase Enzyme From Bacillus spp. J. Cell Mol. Res. 2016, 8, 90–97. [Google Scholar]
- Ramesh, A.; Sharma, S.K.; Joshi, O.P.; Khan, I.R. Phytase, phosphatase activity and P-nutrition of soybean as influenced by inoculation of Bacillus. Indian J. Microbiol. 2011, 51, 94–99. [Google Scholar] [CrossRef] [PubMed]
- Abraham, W.R.; Estrela, A.B.; Nikitin, D.I.; Smit, J.; Vancanneyt, M. Brevundimonas halotolerans sp. nov., Brevundimonas poindexterae sp. nov. and Brevundimonas staleyi sp. nov., prosthecate bacteria from aquatic habitats. Int. J. Syst. Evol. Microbiol. 2010, 60, 1837–1843. [Google Scholar] [CrossRef]
- Bagewadi, Z.K.; Yaraguppi, D.A.; Mulla, S.I.; Deshpande, S.H. Response surface methodology based optimization, partial purification and characterization of alkaline phosphatase isolated from Pseudomonas asiatica strain ZKB1 and its application in plant growth promotion. Mol. Biotechnol. 2022, 64, 984–1002. [Google Scholar] [CrossRef]
- Paul, D.; Sinha, S.N. Phosphate solubilization potential and phosphatase activity of some bacterial strains isolated from thermal power plant effluent exposed water of river Ganga. CIBTech J. Microbiol. 2013, 2, 1–7. [Google Scholar]
- Riedel, R.; Commichau, F.M.; Benndorf, D.; Hertel, R.; Holzer, K.; Hoelzle, L.E.; Martienssen, M. Biodegradation of selected aminophosphonates by the bacterial isolate Ochrobactrum sp. BTU1. Microbiol. Res. 2024, 280, 127600. [Google Scholar] [CrossRef] [PubMed]
- Baldi, P.C.; Giambartolomei, G.H. Brucella. In Molecular Medical Microbiology; Academic Press: Cambridge, MA, USA, 2024; pp. 1657–1679. [Google Scholar]
- Cheng, W.M.; Liu, J.D.; Feng, Y.; Hu, X.M.; Zhao, Y.Y.; Liu, Y. Study on the cooperation mechanism of urea-hydrolysis bacteria and biosurfactant bacteria for dust suppression. Chem. Eng. J. 2024, 480, 148008. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).