Abstract
Mining and industrial activity are contributing to the increase in heavy metal (HM) pollution in soils. Phytoremediation coupled to selected rhizosphere microbiota is an environmentally friendly technology designed to promote HM bioremediation in soils. In this study, sunflower (Helianthus annuus L.) was used together with Rhizophagus irregularis, an arbuscular mycorrhizal fungi (AMF), and Cupriavidus sp. strain 1C2, a plant growth promoting rhizobacteria (PGPR), as a phytoremediation strategy to remove Zn and Cd from an industrial soil (599 mg Zn kg−1 and 1.2 mg Cd kg−1). The work aimed to understand if it is possible to gradually remediate the tested soil while simultaneously obtaining significant yields of biomass with further energetic values by comparison to the conventional growth of the plant in agricultural (non-contaminated) soil. The H. annuus biomass harvested in the contaminated industrial soil was 17% lower than that grown in the agricultural soil—corresponding to yields of 19, 620, 199 and 52 g m−2 of roots, stems, flowers and seeds. It was possible to remove ca. 0.04 and 0.91% of the Zn and Cd of the industrial soil, respectively, via the HM accumulation on the biomass produced. The survival of applied microbiota was indicated by a high root colonization rate of AMF (about 50% more than in non-inoculated agricultural soil) and identification of strain 1C2 in the rhizosphere at the end of the phytoremediation assay. In this study, a phytoremediation strategy encompassing the application of an energetic crop inoculated with known beneficial microbiota applied to a real contaminated soil was successfully tested, with the production of plant biomass with the potential for upstream energetic valorisation purposes.
1. Introduction
Soil is a vital resource for living organisms and the sustainability of our species highly depends on it. Nevertheless, soil contamination has been severely rising since the industrial revolution and there are currently more than 235 million hectares of distressed land all over the world [1], with the EU accounting for more than 3.6 million contaminated locations (FAO, 2018), the USA with 0.6 million brownfields [2], while developing countries have enormous unaccounted soil polluted areas [2].
Heavy metal (HM) pollution is of great distress, as these compounds are not degradable but only exchangeable from one matrix to another. The area of land contaminated with HM has been growing dramatically, primarily due to mining and metallurgical production, but also due to the burning of fossil fuels, to the application of fertilizers and pesticides, and to the discharge of urban and industrial wastes and sewages [3]. These compounds are severely toxic to life, many of them at very low concentrations, having deleterious effects on microbial diversity of the soil itself [4], on plant establishment and growth [5] and further affecting the food chain, and consequently animals and humans [6].
Numerous technologies for soil remediation have been developed based on physical, chemical or thermal procedures, which can be very expensive, ecosystem disturbing and not in line with the sustainability expected for an environmentally focused intervention [7]. Phytoremediation, a biologically based technology that uses plants and associated rhizosphere microbiota to remove or immobilize HM from contaminated soils [7], is attracting attention—ca. 50% of papers concerning the subject have been published recently [2], as it preserves soil and the installed ecosystems, with some successful reports of field application [8,9]. Amongst these studies, the term phytomanagement—producing revenues from biomass growing in contaminated land [10]—is emerging as a way to increase the sustainability of the technique. Besides promoting other advantages, such as soil erosion control and clean-up, and maintaining biodiversity [11], the establishment of a vegetation cover allows for additional profit through the use of the originated biomass for energetic or amendment purposes [12]. The use of energy crops simultaneously tolerant to metal species is appropriate for applying such a kind of strategy. Sunflower (Helianthus annuus L.)—an annual plant belonging to the family of Asteraceae—is a good candidate, as it is not only a well-known and widely used energy crop, which has also shown to possess phytoremediation capability in Zn- and Cd-contaminated soils [13].
The adaptability mechanisms of such plants under abiotic stress can, thus, be used to endure metal toxicity and improve plant resistance against the installed conditions [14]. Research has indicated that the inoculation with efficient microbes to the soil ecosystem not only improves soil quality, and consequently crop yield and quality [15], but also promotes the efficiency in the phytoremediation capacities of the selected plant species [16]. Rhizospheric microorganisms such as arbuscular mycorrhizal fungi (AMF) have been successfully used in the phytoremediation of HM polluted soils [7,17,18]. AMF form symbiotic associations with most terrestrial plant species, and are able to survive the exposure to very high levels of HM [19]. Mycorrhized plants generally present improved nutrition and, consequently, higher biomass production rates [20], and AMF have an important role on adapting the plant’s response to HM toxicity by increasing root absorption surface areas and regulating the interface between metals in the soils and plant roots [21]. Earlier work has demonstrated the importance of such organisms on the development of plants exposed to HM [17], in particular of the inoculation with the AMF Rhizophagus irregularis of H. annuus under abiotic stress [22]. Plant-growth-promoting rhizobacteria (PGPR) have also been used for improving phytoremediation strategies with success [23], and have been described, not only for their valuable growth promotion characteristics, but also as being able to decrease plant stress under harsh abiotic conditions [24]. Similarly, the importance of selected PGPR inocula has been our subject of study formerly, demonstrating that when inoculating H. annuus with PGPR, and in particular with the bacteria Cupriavidus sp. strain 1C2, plants were able to thrive and have their growth increased developing in soils containing up to 500 mg Zn kg−1 and 30 mg Cd kg−1 [13]. These rhizospheric microorganisms are able to improve the plant’s nutrient procurement through nutrient solubilization; stimulate plant growth through the synthesis of indole-3-acetic acid (IAA); decrease susceptibility to pathogens, through the release of antibiotics, competition for nutrients with pathogens or induction of systemic resistance in the host plant; and diminish plants’ stress-related hormones by producing specific enzymes [24]. In fact, previous work has shown that, when screening Cupriavidus sp. strain 1C2 for its in vitro PGP traits, it demonstrated the production of indole acetic acid, hydrogen cyanide, ammonia and siderophores [25], and it showed tolerance to Cd and Zn up to levels of 500 mg L−1 in liquid culture [26]. Additionally, the possibility of promoting plant growth and survival to different abiotic stresses using a mixed microbiota, composed of PGPR and AMF, was also demonstrated in our previous work to have a great potential, due to the symbiotic relationships established between AMF, PGPR, the soil microbiome and the plant [17,22].
The aim of this work was to evaluate the possibility of producing high-energetic-value biomass in baren contaminated soils, that otherwise are not prone to valorization, and compare the obtained yields and biomass contamination with the one of plants grown in agricultural-driven soils. In fact, although this phytoremediation-derived biomass may represent an opportunity instead of a problem, as biomass is a high-energy source material, only few studies [27,28,29] focused on its valorisation for energetic purposes. Nevertheless, none of the performed studies has reported an integrated strategy comprising the monitoring of the soil remediation and the further utilization of all the plant parts and resulting products. The present work involved the assessment of differences in soil status and on the establishment, biomass production yield and HM accumulation abilities of H. annuus in an industrial soil contaminated with zinc (Zn) and cadmium (Cd), with the assistance of selected AMF and PGPR mixed inocula, using the growth of H. annuus in an agricultural soil as a comparison. The ultimate objective is to understand if this phytomanagement approach serves the needs of biomass production for the generation of several energy products hypothesis, which is described more thoroughly further on, in the report of Paulo et al. [30].
2. Materials and Methods
2.1. Experimental Design and Plant Growth
The soils used in this study were an agricultural soil and an industrial soil from Estarreja, both from the North of Portugal (the properties of both soils are shown in Table 1). The industrial site has a long history of metal contamination, as for many years the discharge of solid residues and of the industrial wastewaters directly into the ground induced an HM contamination scenario [31]. Soil was collected randomly in the selected area, to a 40 cm depth. According to Kabata Pendias [32], the usual total metal levels in non-contaminated soils is below 1 mg Cd kg−1 and 300 mg Zn kg−1. The agricultural soil used in the present work is within this range, while the industrial soil presents levels higher than the thresholds established. Therefore, the agricultural soil was used as control soil, representing the optimal conditions required for H. annuus growth.

Table 1.
Soil properties of the agricultural and industrial soils (analytical method).
The experiment consisted of a design with 2 soil conditions (A- agricultural soil; I—industrialized soil inoculated with plant growth promoting microbiota). Each condition consisted of a 1 m3 pot (1 m × 1 m × 1 m) filled with 1 ton of the corresponding soil.
A mixture of Rhizophagus irregularis, an arbuscular mycorrhizal fungi (AMF) from a commercial product used in previous studies [17,22], and of the bacterial isolate Cupriavidus sp. strain 1C2, a plant growth promoting rhizobacteria (PGPR) [33], was used as plant-growth-promoting microbiota. Ten liters of the commercially AMF product (INOQ, GmbH, Hamburg, Germany) was mixed in the Estarreja’s soil before seeding. The same volume of sterilized vermiculite was added to the agricultural soil. The bacterial strain 1C2 was grown at 30 °C and 150 rpm overnight, using Luria–Bertani’s (LB) medium. Resulting pellets were subsequently washed twice and re-suspended to obtain an inoculum density of ca. 108 CFU mL−1 using 10 mM phosphate buffer, pH 8.0 [22]. After seeding, the industrial soil was inoculated with 2 L of bacterial solution, while 2 L of sterilized phosphate buffer was added to the agricultural soil.
H. annuus seeds (variety IBERICO from LusoSem, Algés, Portugal) were surface-disinfected for 10 min with 0.5% (v/v) NaOCl and were then washed several times with sterilized deionized water [22]. All seeds were propagated directly in the tested soils, and, after seeding, were reduced to 100 per pot (at a distance of 10 × 10 cm).
2.2. Plant Biomass and Metal Accumulation Monitoring
H. annuus plants were harvested after 120 days, subdivided into root, stems (stems and leaves were not separated and are considered in this portion), flowers and seeds, and subsequently washed with deionized water and HCl 0.1 M. Samples were air dried and seeds were extracted; then, remaining tissues were oven-dried at 70 °C for 48 h, after which the biomass dry weight was measured. Plant samples were then grinded and digested at a high temperature in a PerkinElmer MicroWave 3000 following the 3052 USEPA method. Cadmium and Zn content was determined using Flame Atomization-Atomic Absorbance Spectrometry (FA-AAS) of the digests [34] in a Unicam 960 spectrophotometer (Waltham, MA, USA), with a detection limit for Cd of 0.0028 mg L−1 and 0.0033 mg L−1 for Zn. The BCR (Community Bureau of Reference) reference sample CRM 279 (sea lettuce) was analyzed using the above-described Cd and Zn determination analytical method. The values obtained by FA-AAS (0.28 ± 0.01 and 52.8 ± 0.9 mg kg−1 for Cd and Zn respectively) confirmed the accuracy and precision of the method by comparison with the certified value (0.274 ± 0.022 and 51.3 ± 1.2 mg kg−1 for Cd and Zn, respectively).
2.3. Bioaccumulation Factor and Remediation Ratio
Bioaccumulation (or bioconcentration) factors (BFs) and remediation ratios (RRs) were calculated according to Mani et al. [35], and translocation factors (TFs), according to Moreira et al. [36], as shown in Equations (1)–(3):
BFs, TFs and RRs were also calculated for the agricultural soil in order to also have information on the pattern of Zn and Cd uptake by the plant, when these metals are also present in the soil at lower concentrations.
2.4. Cd and Zn Mobilization in Soil
The soil of each pot was collected at the beginning and at the end of the experiment in order to assess the water and the ammonium acetate (NH4-Ac) extractable Cd and Zn fractions. Solutions of 1:5 soil water and 1:5 soil NH4-Ac [37] were incubated in rotating flasks at 20 °C for 2 h. The extracts were centrifuged for 10 min at 38,000 rpm and the supernatants were then filtered over a 0.45 µm cellulose acetate filter and collected in test tubes. Cd and Zn contents were determined using the FA-AAS method, as described above.
2.5. Monitoring of Bacterial Community in Soils
A DNeasy PowerSoil Kit (Qiagen, Hilden, Germany) was used for DNA extraction from duplicate soil samples, collected at the initial and final stages of plant growth, for both soil conditions, according to manufacturer’s instructions. Qubit (Thermo Fisher Scientific, Waltham, MA, USA) was used for measuring the DNA concentration and the extracted DNA was stored at −20 °C for future use.
NGS of the extracted DNA was performed at GATC-Eurofins (Konstanz, Germany), which included DNA amplification, libraries’ preparation, sequencing and bioinformatics data analysis. The 16S rRNA phylogenetic gene paired-end sequencing was performed to cover the V3-V4 hypervariable region (Illumina MiSeq platform) by using two primers (357F—TACGGGAGGCAGCAG [38]; 800R—CCAGGGTATCTAATCC [39]). The microbiome evaluation and profiling was executed as described in Paulo at al. [40]. The raw sequence data were deposited in the Sequence Read Archive (SRA) from the NCBI database, within the BioProject with accession number PRJNA830831. BLAST from NCBI was used (https://blast.ncbi.nlm.nih.gov/Blast.cgi, accessed on 2 September 2022) for a more complete identification of operational taxonomic unit (OTU) sequences from bacterial families with high relative abundance, but not identified up to the genus level.
2.6. Root Colonization by AMF
A sub-sample of fresh fine roots was collected from the H. annuus plants harvested from each soil condition (agricultural and industrial), and were divided into ca. 1–2 cm fragments, immersed in 10% (w/v) KOH and heated in a 80 °C water bath for 30 min, following a procedure adapted from Vierheilig et al. [41]. Following this, the KOH solution was discarded and the root’s fragments were cleansed for 10 min by adding HCl 3% (v/v). Afterwards, the roots were tainted using a staining solution consisting of 5% ink (Pelican 4001, Brilliant black, Fountain Pen Ink) diluted in 5% (v/v) acetic acid. After this, the roots were boiled for 4 min at 80 °C. Following staining, the roots were rinsed several times with tap water. Stained root samples were examined microscopically to assess the percentage of mycorrhizal colonization using the grid-line intersect method [42].
2.7. Statistical Analysis
Statistical analysis of the differences in the parameters in all tested treatments was performed applying one-way ANOVA (to compare metal accumulation in each analyzed plant tissue for each one of the tested soils) and t-tests (for the comparison of the same parameter between both soil conditions) using the IBM SPSS Statistics program (IBM, Armonk, NY, USA, version 28.0). To establish the significance of the differences among the means, Duncan tests (p < 0.05) were performed.
3. Results
3.1. H. annuus Biomass Production Yields
The biomass of the different plant sections at harvest was determined. H. annuus plants growing in agricultural soil always presented higher biomass yields for any plant section than plants grown on industrial soil (Table 2). Differences in plant growth were visible, particularly for the stems (being less thick and with a lower number of leaves for plants grown in industrial soil) and flowers (number and size were lower for plants grown in industrial soil), and can be seen in Figure 1. Values per pot (m2) ranged from ca. 63 g for the seeds and ca. 750 g for the stems of H. annuus plants growing in agricultural soil, and from ca. 52 g for the seeds to ca. 620 g for the stems of H. annuus plants growing in industrial soil. Biomass production was higher for the stems, followed by the flowers, the seeds and the roots, independently from soil type. The root was more affected by the soil matrix, reducing ca. 42% in the industrial soil compared to the agricultural soil, with stems and seeds being similarly affected by soil condition, decreasing ca. 17% in the contaminated soil in comparison to the agricultural soil. The formation of flowers from plants grown in industrial soil was 10% lower compared to the agricultural soil. Considering the entire plant as a sum of all plant sections, the biomass reduction of overall H. annuus growing in the industrial soil was ca. 17%, when comparing to plants grown in the agricultural soil.
Table 2.
Biomass production yields per pot (m2 soil) for H. annuus plants grown in agricultural and industrial soils.

Figure 1.
H. annuus development in agricultural soil (A) and industrial soil (B) throughout the experiment.
3.2. Metal Accumulation in H. annuus Plants
The effect of growing H. annuus plants in metal-contaminated soil was assessed and levels of Zn and Cd for the different plant sections are given in Table 3. Values for Zn and Cd were higher in plants grown in industrial soil, being also higher in roots compared to seeds, for both soil conditions. The highest accumulation of Cd and Zn was registered for the roots independently of the soil matrix and the accumulation profile was as follows: roots > stems > flowers > seeds, for both soil conditions and metals; no Cd was detected in the flowers and seeds of H. annuus grown in agricultural soil. Zn values ranged from an average of 2 mg kg−1 in seeds to 67 mg kg−1 in roots of plants grown in agricultural soil and from an average of 4 mg kg−1 in seeds to 434 mg kg−1 in roots of plants grown in an industrial soil. For Cd, values ranged from the non-detection in seeds to 1.6 mg kg−1 in the roots of plants grown in agricultural soil and from an average of 0.5 mg kg−1 in seeds to 24 mg kg−1 in the roots of plants grown in industrial soil.
Table 3.
Zn and Cd accumulation in the different tissues of H. annuus plants grown in agricultural and industrial soil.
In all plant tissues, except for the seeds, significant (p < 0.05) differences were shown between the levels of Zn and Cd, with plants grown in the industrial soil presenting higher accumulation than those grown in the agricultural soil. The accumulation levels in the seeds showed no significant (p < 0.05) difference between soils.
3.3. Bioaccumulation and Translocation Factors and Remediation Ratios
Bioaccumulation factors (BFs), translocation factors (TFs) and remediation ratios (RRs) were calculated and are represented in Figure 2. All BFs, TFs and RRs were generally higher for Cd than for Zn. For Zn, BFs were higher for plants growing in the agricultural soil (maximum of ca. 1.4), while for Cd the factors were higher for plants exposed to the industrial soil (maximum ca. 11.2). The same trend was also verified for the RRs, with the highest (ca. 9.8) and lowest (ca. 0.39) values being shown for Cd and Zn for H. annuus grown in industrial soil, respectively (Figure 2). For TFs, values were generally lower than 1 and were maximized both for Zn and Cd in plants grown in the agricultural soil (maximums of ca. 0.76 and 1.33 respectively).
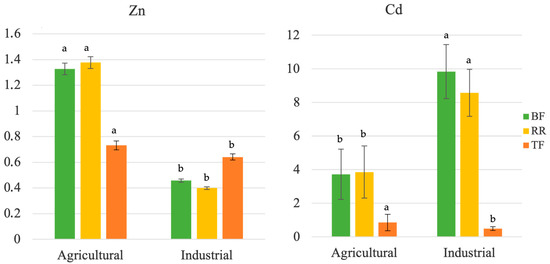
Figure 2.
Bioaccumulation factors (BFs), translocation factors (TFs) and remediation ratios (RRs) of Zn and Cd for H. annuus plants growing in agricultural and industrial soil. Means for the same parameter showing different letters are significantly different from each other (p < 0.05). Independent sample t-tests were performed for each parameter to determine the effect of soil type and results are: t(BFZn) = 6.709, t(BFCd) = 0.002, t(RRZn) = 7.685, t(RRCd) = 0.131, t(TFZn) = 0.771, and t(TFCd) = 2.503.
3.4. Cd and Zn Mobilization in Soils
No metals were detected in the water extracts at the beginning and at the end of the experiment. Metal levels in soil ammonium acetate extracts at the beginning and at the end of the experiment are described in Table 4. No bioavailable Cd was detected in any of the samples. Either at the beginning or at the end of the experiment, the levels of bioavailable Zn were always significantly (p < 0.05) higher for the industrial soil. Also, and for both tested soils, levels of bioavailable Zn increased significantly (p < 0.05) at the end of the study (Table 4).
Table 4.
Metal levels in soil ammonium acetate extracts at the beginning and at the end of the experiment.
3.5. Monitoring of Bacterial Community Present in Soils
The microbial community diversity at the phylum level was found to be similar in both soil conditions, mostly regarding the abundance of Actinobacteria, Proteobacteria, Acidobacteria and Firmicutes (Figure 3). Actinobacteria dominated the bacterial community in both soils, representing more than 45% and 53% of the total relative abundance in the agricultural and the industrial soil, respectively. Proteobacteria presented between 18 and 29% of relative abundance in both soil conditions. Acidobacteria were present with a similar relative abundance in agricultural soil throughout plant growth (10–16%), increasing from 4 to 10% in the industrial soil. Firmicutes presented similar relative abundances between both soil conditions (2–5.5%). Gemmatimonadetes were not identified in any of the industrial soil samples but were detected in all agricultural soil samples, increasing their relative abundance at the end of plant growth. Nitrospirae were present in higher relative abundance in the agricultural soil compared to the industrial soil, where their numbers increased at the end of plant growth. Cyanobacteria were only identified in the agricultural soil (relative abundance between 0.6 and 1.3%).
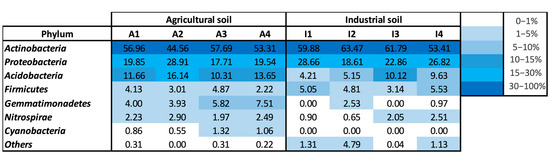
Figure 3.
Heatmap of the relative abundance of bacterial phyla present in the soil samples. A—agricultural soil; I—industrial soil; 1,2—duplicate samples of soil from the beginning of plant growth; 3,4—duplicate samples of soil from the end of plant growth.
Both soil conditions presented a high relative abundance of Actinobacteriia, Rubrobacteria and Alphaproteobacteria classes (Figure 4). Acidobacteriia and Nitrospira presented higher and stable relative abundance in the agricultural soil, compared to the industrial soil and Acidimicrobiia and Gemmatimonadetes, which were mostly detected in the agricultural soil, while Betaproteobacteria were more abundant in the industrial soil. Thermoleophilia, Bacilli and Alphaproteobacteria were identified in similar relative abundance in both types of soil. Comparing the beginning and end of plant growth, Gemmatimonadetes increased while Betaproteobacteria decreased in the agricultural soil. In the industrial soil, both Acidobacteria and Nitrospira increased while Actinobacteria decreased in relative abundance, at the harvest.
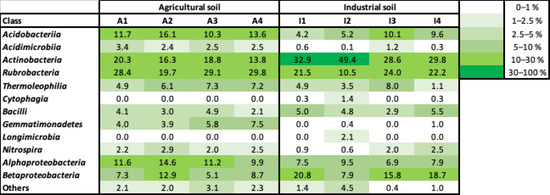
Figure 4.
Heatmap of the relative abundance of the bacterial classes present in the soil samples. A—agricultural soil; I—industrial soil; 1,2—duplicate samples of soil from the beginning of plant growth; 3,4—duplicate samples of soil from the end of plant growth.
The ten dominant bacterial genera accounted for more than 70% of the total relative abundance in most of the soil samples (except for I2). Several bacterial genera were present with similar relative abundance in both soil conditions, at the beginning and end of plant growth (Figure 5). This was the case for Gaiella, which presented the highest relative abundance in all soil samples, and Sphingomonas, Nakamurella and Bacillus, present with similar relative abundance in both soils. However, the agricultural soil presented several bacteria with higher relative abundance compared to the industrial soil. Acidobacterium and Nitrosospira were present with higher relative abundance in the agricultural soil compared to the industrial soil, as well as Solirubrobacter, Gemmatirosa, Aciditerrimonas, Gemmatimonas and Candidatus Koribacter. On the other hand, the industrial soil presented a higher relative abundance of Conexibacter, Geodermatophilus, Blastococcus and Arthrobacter bacterial genera.
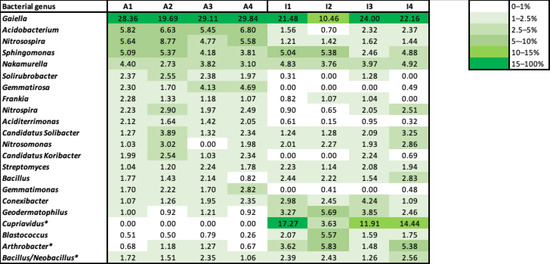
Figure 5.
Heatmap for the most abundant bacterial genera in the soil samples, considering the top 10 bacterial genera identified in each soil sample. Bacterial genera indicated with an asterisk (*) were further identified using BLAST from NCBI. A—agricultural soil; I—industrial soil; 1,2—duplicate samples of soil from the beginning of plant growth; 3,4—duplicate samples of soil from the end of plant growth.
Although most of the bacterial genera kept their relative abundance between the beginning and the end of plant growth, Gemmatirosa increased in relative abundance in the agricultural soil, while Acidobacterium, Nitrospira and Candidatus Koribacter increased in the industrial soil.
Ten OTU sequences belonging to the Burkholderiaceae family were further identified in the industrial soil, using BLAST from NCBI. All sequences were found to be highly similar (>99% identity) to the 16S rRNA sequence of the Cupriavidus sp. 1C2 strain, inoculated at the beginning of plant growth in the industrial soil. According to this information, Cupriavidus sp. 1C2 was identified in the industrial soil, presenting a high relative abundance at the beginning and end of plant growth (between 12 and 17%), except for sample I2, possibly due to experimental variations, associated with the size of the soil sample collected for DNA extraction, and/or NGS sequencing errors. This bacterial genus was not identified in the agricultural soil.
3.6. Mycorrhizal Colonization of Roots
Mycorrhizal colonization was determined for root samples from H. annuus plants grown in both tested soils at harvest, and percent colonisations are described in Table 5. Colonization was observed for plants growing in both soil conditions, with plants growing in industrial soil (inoculated with the selected AMF Rhizophagus irregularis showing a higher degree of colonization (ca. 43%) when comparing to non-inoculated plants grown in agricultural soil (ca. 27%).
Table 5.
Mycorrhizal colonization of roots of H. annuus growing in agricultural and industrial soils.
4. Discussion
H. annuus inoculated with beneficial microbiota was grown in an industrialized Zn- and Cd-contaminated soil in order to test the biomass production abilities for further energy conversion of this phytoremediation strategy by comparison with the conventional growth in agricultural soil. A lower H. annuus biomass production was observed in the industrial soil, compared to the non-contaminated agricultural soil. This is associated with the high Zn and Cd contamination level [32], which, as a consequence, was partly bioaccumulated by the plant. The contamination of a soil with such levels of metals can impair plant growth not only by disturbing its nutrient uptake—high concentrations of metals in the soil will result in reduced uptake of water and nutrients [6]—but also by damaging severely its metabolic activities, by inhibiting physiologically active enzymes and mineral metabolism and, consequently, decreasing the rates of photosynthesis [43]. In the present study, the impairment was more severe for the plant’s roots compared to the upper parts of the plants, as previously described in the literature for H. annuus. For example, Alaboudi et al. [43] reported decreases in the root and shoot dry weights of H. annuus of 35 and 14%, respectively, when exposed to a 10 mg Cd kg−1 spiked soil. Marques et al. [13] showed that root and shoot dry weights of H. annuus were reduced by up to 83 and 40%, respectively, when exposed to a 500 mg Zn kg−1 spiked soil; twice higher values when compared to those obtained in the present study.
Despite the greater effect in H. annuus root, the overall biomass decrease was lower than 20%. This decrease was accompanied by an increase in both Zn and Cd accumulation in all plant tissues; maximum increases of up to ca. 6.5 and 6.1-fold were registered for the root and stems tissues for Zn, and up to 15 times for the root and stem tissues for Cd. This resulted in Zn and Cd accumulation levels exceeding the phytotoxic concentrations proposed by Kabata Pendias [32]—100 to 400 mg Zn kg−1 and 5 mg Cd kg−1—with values as high as 434 mg Zn kg−1 and 24 mg Cd kg−1. H. annuus plants seemed to have adopted an immobilization strategy of the metals in the roots, which presented a significantly higher level of both Zn and Cd accumulation. This generally resulted in translocation factors (TFs) lower than 1—similarly to what happened in previous studies [13]. The range of Zn and Cd concentrations observed in this study for the tissues of H. annuus grown in the industrial site are within the range of other studies with similar soil contamination profiles; for a metal-contaminated soil (575 mg Zn kg−1 and 6 mg Cd kg−1), Meers et al. [44] described levels of Zn and Cd accumulation in the shoots of H. annuus of up to ca. 250 and 5 mg kg−1, respectively; Nehnevajova et al. [45] reported concentrations of Zn and Cd in the stems of up to 274 and 0.67 mg kg−1, respectively, and in the leaves of up to 401 and 1.58 mg kg−1, respectively, for H. annus plants grown in a soil contaminated with average levels of 759 mg Zn kg−1 and 0.81 mg Cd kg−1. However, the values reported in these studies are converted in bioaccumulation factors (BFs) lower than those observed in the present report for the industrial soil for both Cd and Zn. In this work, BFs and consequently remediation ratios (RRs) were always higher for Cd than for Zn, which is in accordance with the commonly recognized higher Cd transfer coefficient between soil and plant [46], probably affected by the positive relation between metal electro-negativity (according to the Pauling scale, Zn = 1.65 and Cd = 1.69) and metal uptake ability. For Zn, both BF and RR values decreased when sunflower was grown in contaminated soil and the opposite happened for Cd. Nevertheless, values were generally higher for all cases than those registered in the report of Mani et al. [35]: RRs up to ca. 0.6 and. 0.2, and BFs up to ca. 2 and 0.75, for Cd and Zn, respectively, for a soil showing a contamination of 2.2 mg Cd kg−1 and 39 mg Zn kg−1.
Despite the growth reduction associated with increased metal uptake, H. annuus plants were able to grow and produce significant amounts of biomass under the harsh conditions of metal toxicity presented by the industrial soil. Although this could be explained by different mechanisms described in the literature for the plant to endure the presence of such heavy metals, such as enhancement of metal detoxification through metal-sequestering molecule production [47], improvement of antioxidant systems [48], and tolerance mechanisms implicating plant growth regulators [49], the establishment of a symbiotic association between the plant and the mixture of beneficial microorganisms might have played an important role in H. annuus growth and phytoremediation ability. AMF and PGPR can promote a higher resistance to toxicity and ability to grow and produce biomass under metal stress. In the present work, in order to help the plants to cope with the installed metal contamination conditions, the industrial soil was inoculated with the AMF Rhizophagus irregularis and consequently the H. annuus plants growing in this soil presented higher AMF colonization rates in their roots. High arbuscle formation is revealing of active AMF symbiosis, as they are exchange sites in the root cortex for mineral elements [20,21]. This increase is similar to the one reported in the study by Zhang et al. [18] when a soil from a recycling base was inoculated with the AMF Funneliformis caledonium for assisting the growth of garlic chives and H. annuus. Similarly, the industrial soil was inoculated with a heavy-metal-site-derived bacterial strain Cupriavidus sp. strain 1C2. From the follow-up of the microbial community conducted throughout the experiment, it was observed that the inoculated PGPR prevailed in the rhizosphere, possibly keeping its activity until the end of the experiment. These results were somehow expected, since, as aforementioned, this strain was initially isolated from an industrial soil contaminated with heavy metals [26]. Different mechanisms of microbial protection and stabilization of H. annuus can play a part under such metal stress, having an action either on growth promotion of the plant (recovering the potential effect of the metals on plant yields) or on the regulation of metal [14]. Although inoculating AMF or PGPR alone can improve plant performance under different abiotic stressing conditions, by helping plant’s establishment, survival, growth and physiology [50], several studies have shown that AMF and PGPR together can provide a more positive environment for the plant and the rhizosphere microbiota [17,22,35]. The AMF action might not only be associated with healthier plants’ physiological traits, but also due to its positive effect on soil bacteria; AMF exudates can be used as source of nutrients by bacteria, while creating bacterial habitats with their hyphae, and soil microbes produce compounds that can increase the amount of root exudates as well as plant hormones, influencing, in this way, AMF establishment and rate of root colonization [17,22].
Another key strategy to increase the effectiveness of phytoremediation is to increase HM bioavailability [3], and both Cd and Zn became more bioavailable at the end of the experiment. Exudates from plant roots can influence the pH of the soil solution and consequently affect metal availability, as soil acidification on the rhizosphere is a key mechanism accountable for rising metal solubility [51]. Biosurfactants and organic acids, produced by the bacterial community present in the rhizosphere, can also increase metal bioavailability, by helping the desorption of metals from the soil matrix and maximizing metals’ chelation [2]. Also, arbuscular mycorrhizal fungi can alter soil properties through their metabolic processes and consequently promote changes in the availability of the existing metals [52]. Some of these factors may have contributed to the rise of metal availability in both tested situations.
As cadmium and Zn are toxic metals, their presence and increased bioavailability throughout the experiments can cause changes in the composition of soil microbial communities. The functional diversity of the soil microbial community has been used as a pointer for assessing the effect of revegetation on soil quality [11] and a considerable number of studies have focused on the liaison between soil microbial communities and soil contamination by heavy metals [11,13,17]. Despite the different use and contamination status of the tested soils, the initial bacterial phyla dominance was similar. A deeper analysis into the taxonomical classification allowed identifying a higher diversity of bacterial genera present in the agricultural compared to the industrial soil. These results are in accordance with the report of de Quadros et al. [53], which showed a decrease in the diversity of the microbial community of post-mined sites by comparison with undisturbed land, as expected, due to the lower level of abiotic stressors present in the agricultural soil. Acidobacteria, Firmicutes, Actinobacteria and Proteobacteria were found to be among the most dominant bacterial phyla present in soil [54]. The initial higher numbers of Acidobacteria, and Acidobacterium, in the agricultural soil could be explained by the fact that members of the Acidobacteriaceae family are abundant in mature soils with plant cover, in the presence of carbon, nutrients and lower pH [55]. Gaiella, from the Rubrobacteria class (Actinobacteria) was dominant in both soils, regardless of the land use and contamination status. Gaiella might be ubiquitous in soil and tolerant to HMs, since it was previously identified in farmland [56] and in heavy-metal-contaminated soil [57]. Nakamurella (Actinobacteria class and phyla) was also present in both soils. This is a more rare genus, with some species isolated from soil [58]. Sphingomonas sp. (Alphaproteobacteria class) was also identified with high relative abundance in the tested soils. This bacterium is widely distributed in water and soil, associated with plants and can improve plants’ development in stress conditions such as drought, salinity and in the presence of heavy metals [59]. On the other hand, some bacteria were identified with higher relative abundance in the industrial soil, such as Arthrobacter. These bacteria are normally found in the rhizosphere, being also present in HM-contaminated sites [60].
Ammonium-oxidizing bacteria (AOB) and nitrite-oxidizing bacteria (NOB) are sensitive to HMs and their enzyme activities can be used as bioindicators of HM contamination [61]. Representative of AOB and dominant in soil microbial communities [62], both Nitrosomonas and Nitrosospira bacterial genera were identified in the tested soils. The presence of a lower abundance of Nitrosospira (AOB) in the industrial compared to the agricultural soil might be associated with HM contamination sensitivity, while the increased abundance of Nitrospira (NOB) after the phytoremediation process might be linked to a decrease in HM concentration in the rhizosphere. Other bacteria possibly favoured by the phytoremediation process were Acidobacterium and Candidatus Koribacter. Despite being known as HM-tolerant [63], Gemmatimonadetes was not present in the industrial soil in relative abundance during the phytoremediation process.
The background levels of Zn in soils should not be higher than 300 mg Zn kg−1 dry soil and 1 mg Cd kg−1 dry soil (EC Directive 86/278/CEE from 12 June). The industrial soil is above the legislated limits and fell within the range of contamination seen as potential for the application of phytoremediation-based clean-up strategies. Using the data presented on this study, considering equal plant development (4 crops/year) and similar soil contamination characteristics, a minimum of 20 crops would be necessary to achieve Cd soil clean-up and 1211 crops for Zn (which will correspond for 1 ton of soil to a minimum time span of 5 and more than 300 years, respectively). The accumulations of Zn and Cd in sunflowers grown in the industrial site were above the range for plants cultivated on uncontaminated parcels [64], and well above concentrations shown to be damaging to animals [65], and therefore their use as products for human or livestock consumption is not allowed. Alternative applications for this biomass should be thus considered, namely as sources of biomass for energy production [66]. This option has emerged as a possibility in recent reports [11,27] but needs further study, as heavy metals may affect the product generation routes and ultimately contamination levels should always be quantified to understand if the final product is compatible with the anticipated purpose [11].
Here, the assessment of the potential of the H. annuus biomass produced for the generation of biofuels, namely using an integrated strategy encompassing the utilization of all of the plant sections and analysis of the resulting products concerning heavy metal contents, is further analysed in the study by Paulo et al. [30]. This report shows that the production of biofuels (namely oil, bioethanol, biodiesel and biogas) from the biomass derived from such a phytomanagement strategy of this industrial HM-contaminated soil can be feasible and can serve to counterpart the growing need for biomass for biofuels generation.
5. Conclusions
The total H. annuus biomass obtained after the phytoremediation strategy was less than 20% lower compared to growth in an agricultural soil. The highest metal accumulation occurred in the plant’s roots (434 mg Zn kg−1 and 24 mg Cd kg−1) followed by the stems (343 mg Zn kg−1 and 15 mg Cd kg−1), flowers (129 mg Zn kg−1 and 5.3 mg Cd kg−1) and seeds (4 mg Zn kg−1 and 0.5 mg Cd kg−1), associated with low translocation factors (generally < 1). An expected increase in Cd and Zn mobilization in soil was also verified (up to 34 and 19% in agricultural and industrial soils, respectively). The persistence of inoculated PGPR and AMF colonization in the treated industrial soil was observed, indicating the ability of both types of microorganisms to survive.
A phytoremediation strategy encompassing the application of an energetic crop inoculated with known beneficial microbiota can lead to the production of a significant amount of biomass, increasing its potential use for different valorisation purposes.
Author Contributions
A.M.S.P.: Conceptualization, Validation, Investigation, Methodology, Visualization; N.S.C.: Writing; P.M.L.C.: Conceptualization, Writing; A.P.G.C.M.: Conceptualization, Validation, Investigation, Methodology, Writing, Visualization, Supervision, Project Administration, Funding Acquisition. All authors have read and agreed to the published version of the manuscript.
Funding
This work was financially supported by: project PHYTOENERGY-PTDC/BTA-BTA/28761/2017 (POCI-01-0145-FEDER-028761), funded by National Funds from FCT—Fundação para a Ciência e a Tecnologia; and UIDB/50016/2020 (CBQF), LA/P/0045/2020 (ALiCE), UIDB/00511/2020 and UIDP/00511/2020 (LEPABE), and UIDB/04730/2020 (CIETI), funded by national funds through FCT/MCTES (PIDDAC).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available in the article.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Bermudez, G.M.A.; Jasan, R.; Plá, R.; Pignata, M.L. Heavy metals and trace elements in atmospheric fall-out: Their relationship with topsoil and wheat element composition. J. Hazard. Mater. 2012, 213–214, 447–456. [Google Scholar] [CrossRef] [PubMed]
- Shah, V.; Daverey, A. Phytoremediation: A multidisciplinary approach to clean up heavy metal contaminated soil. Environ. Technol. Innov. 2020, 18, 100774. [Google Scholar] [CrossRef]
- Yan, A.; Wang, Y.; Tan, S.N.; Mohd Yusof, M.L.; Ghosh, S.; Chen, Z. Phytoremediation: A Promising Approach for Revegetation of Heavy Metal-Polluted Land. Front. Plant Sci. 2020, 11, 359. [Google Scholar] [CrossRef] [PubMed]
- Qian, Q.; Hu, C.; Lin, J.; Wang, X.; Tang, C.; Dai, Z.; Xu, J. Contamination with multiple heavy metals decreases microbial diversity and favors generalists as the keystones in microbial occurrence networks. Environ. Pollut. 2022, 306, 119406. [Google Scholar] [CrossRef]
- Xu, Y.; Seshadri, B.; Sarkar, B.; Wang, H.; Rumpel, C.; Sparks, D.; Farrell, M.; Hall, T.; Yang, X.; Bolan, N. Biochar modulates heavy metal toxicity and improves microbial carbon use efficiency in soil. Sci. Total Environ. 2018, 621, 148–159. [Google Scholar] [CrossRef]
- Khan, A.; Khan, S.; Khan, M.A.; Qamar, Z.; Waqas, M. The uptake and bioaccumulation of heavy metals by food plants, their effects on plants nutrients, and associated health risk: A review. Environ. Sci. Pollut. Res. 2015, 22, 13772–13799. [Google Scholar] [CrossRef]
- Marques, A.P.G.C.; Rangel, A.O.S.S.; Castro, P.M.L. Remediation of heavy metal contaminated soils: Phytoremediation as a potentially promising clean-Up technology. Crit. Rev. Environ. Sci. Technol. 2009, 39, 622–654. [Google Scholar] [CrossRef]
- Reddy, K.R.; Amaya-Santos, G.; Yargicoglu, E.; Cooper, D.E.; Negri, M.C. Phytoremediation of heavy metals and PAHs at slag fill site: Three-year field-scale investigation. Int. J. Geotech. Eng. 2019, 13, 32–47. [Google Scholar] [CrossRef]
- Willscher, S.; Mirgorodsky, D.; Jablonski, L.; Ollivier, D.; Merten, D.; Büchel, G.; Wittig, J.; Werner, P. Field scale phytoremediation experiments on a heavy metal and uranium contaminated site, and further utilization of the plant residues. Hydrometallurgy 2013, 131–132, 46–53. [Google Scholar] [CrossRef]
- Moreira, H.; Pereira, S.I.A.; Mench, M.; Garrbisu, C.; Kidd, P.; Castro, P.M.L. Phytomanagement of Metal(loid)-Contaminated Soils: Options, Efficiency and Value. Front. Environ. Sci. 2021, 9, 661423. [Google Scholar] [CrossRef]
- Agnello, A.C.; Potysz, A.; Fourdrin, C.; Huguenot, D.; Chauhan, P.S. Impact of pyrometallurgical slags on sunflower growth, metal accumulation and rhizosphere microbial communities. Chemosphere 2018, 208, 626–639. [Google Scholar] [CrossRef]
- Evangelou, M.W.H.; Deram, A. Phytomanagement: A realistic approach to soil remediating phytotechnologies with new challenges for plant science (Mini review). Int. J. Plant Biol. Res. 2014, 2, 1023. [Google Scholar]
- Marques, A.P.G.C.; Moreira, H.; Franco, A.R.; Rangel, A.O.S.S.; Castro, P.M.L. Inoculating Helianthus annuus (sunflower) grown in zinc and cadmium contaminated soils with plant growth promoting bacteria—Effects on phytoremediation strategies. Chemosphere 2013, 92, 74–83. [Google Scholar] [CrossRef]
- Abeed, A.H.A.; Mahdy, R.E.; Alshehri, D.; Hammami, I.; Eissa, M.A.; Abdel Latef, A.A.H.; Mahmoud, G.A. Induction of resilience strategies against biochemical deteriorations prompted by severe cadmium stress in sunflower plant when Trichoderma and bacterial inoculation were used as biofertilizers. Front. Plant Sci. 2022, 13, 1004173. [Google Scholar] [CrossRef]
- Li, J.; Wang, J.; Liu, H.; Macdonald, C.A.; Singh, B.K. Application of microbial inoculants significantly enhances crop productivity: A meta-analysis of studies from 2010 to 2020. J. Sustain. Agric. Environ. 2022, 3, 216–225. [Google Scholar] [CrossRef]
- Ma, Y.; Rajkumar, M.; Zhang, C.; Freitas, H. Beneficial role of bacterial endophytes in heavy metal phytoremediation. J. Environ. Manag. 2016, 174, 14–25. [Google Scholar] [CrossRef]
- Moreira, H.; Pereira, S.I.A.; Marques, A.P.G.C.; Rangel, A.O.S.S.; Castro, P.M.L. Mine land valorization through energy maize production enhanced by the application of plant growth-promoting rhizobacteria and arbuscular mycorrhizal fungi. Environ. Sci. Pollut. Res. 2016, 23, 6940–6950. [Google Scholar] [CrossRef]
- Zhang, Y.; Hu, J.; Bai, J.; Qin, H.; Wang, J.; Wang, J.; Lin, X. Intercropping with sunflower and inoculation with arbuscular mycorrhizal fungi promotes growth of garlic chive in metal-contaminated soil at a WEEE-recycling site. Ecotoxicol. Environ. Saf. 2019, 167, 376–384. [Google Scholar] [CrossRef]
- Sánchez-Castro, I.; Gianinazzi-Pearson, V.; Cleyet-Marel, J.C.; Baudoin, E.; van Tuinen, D. Glomeromycota communities survive extreme levels of metal toxicity in an orphan mining site. Sci. Total Environ. 2017, 598, 121–128. [Google Scholar] [CrossRef]
- Read, S.E.; Smith, D. Mycorrhizal Symbiosis, 2nd ed.; Academic Press: London, UK, 2008. [Google Scholar]
- Riaz, M.; Kamran, M.; Fang, Y.; Wang, Q.; Cao, H.; Yang, G.; Deng, L.; Wang, Y.; Zhou, Y.; Anastopoulos, I. Arbuscular mychorrizal fungi induced mitigation of heavy metal phytotoxicity in metal contaminated soils: A critical review. J. Hazard. Mater. 2021, 402, 123919. [Google Scholar] [CrossRef]
- Pereira, S.I.A.; Moreira, H.; Argyras, K.; Castro, P.M.L.; Marques, A.P.G.C. Promotion of sunflower growth under saline water irrigation by the inoculation of beneficial microorganisms. Appl. Soil Ecol. 2016, 105, 36–47. [Google Scholar] [CrossRef]
- Jin, Z.; Deng, S.; Wen, Y.; Jin, Y.; Pan, L.; Zhang, Y.; Black, T.; Jones, K.C.; Zhang, H.; Zhang, D. Application of Simplicillium chinense for Cd and Pb biosorption and enhancing heavy metal phytoremediation of soils. Sci. Total Environ. 2019, 697, 134148. [Google Scholar] [CrossRef] [PubMed]
- Tabassum, B.; Khan, A.; Tariq, M.; Ramzan, M.; Khan, M.S.I.; Shahid, N.; Aaliya, K. Bottlenecks in commercialisation and future prospects of PGPR. Appl. Soil Ecol. 2017, 121, 102–117. [Google Scholar] [CrossRef]
- Marques, A.P.G.C.; Pires, C.; Moreira, H.; Rangel, A.O.S.S.; Castro, P.M.L. Assessment of the plant growth promotion abilities of six bacterial isolates using Zea mays as indicator plant. Soil Biol. Biochem. 2010, 42, 1229–1235. [Google Scholar] [CrossRef]
- Pires, C. Bacteria in Heavy Metal Contaminated Soil: Diversity, Tolerance and Use in Remediation Systems; Cranfield University: Cranfield, UK, 2010. [Google Scholar]
- Balsamo, R.A.; Kelly, W.J.; Satrio, J.A.; Ruiz-Felix, M.N.; Fetterman, M.; Wynn, R.; Hagel, K. Utilization of Grasses for Potential Biofuel Production and Phytoremediation of Heavy Metal Contaminated Soils. Int. J. Phytoremediation 2015, 17, 448–455. [Google Scholar] [CrossRef]
- Olivares, A.R.; Carrillo-González, R.; González-Chávez, M.C.A.; Hernández, R.M.S. Potential of castor bean (Ricinus communis L.) for phytoremediation of mine tailings and oil production. J. Environ. Manag. 2013, 114, 316–323. [Google Scholar] [CrossRef]
- Chami, Z.A.; Amer, N.; Smets, K.; Yperman, J.; Carleer, R.; Dumontet, S.; Vangronsveld, J. Evaluation of flash and slow pyrolysis applied on heavy metal contaminated Sorghum bicolor shoots resulting from phytoremediation. Biomass Bioenergy 2014, 63, 268–279. [Google Scholar] [CrossRef]
- Paulo, A.M.S.; Caetano, N.S.; Marques, A.P.G.C. Bioenergetic Products from Phytoremediation Derived Biomass: Assessment of the Potential of Sunflower Grown in Metal Contaminated Soils for the Production of Biofuels. 2023. Available online: http://dx.doi.org/10.2139/ssrn.4441114 (accessed on 26 July 2023).
- Marques, A.P.G.C.; Moreira, H.; Rangel, A.O.S.S.; Castro, P.M.L. Arsenic, lead and nickel accumulation in Rubus ulmifolius growing in contaminated soil in Portugal. J. Hazard. Mater. 2009, 165, 174–179. [Google Scholar] [CrossRef]
- Kabata-Pendias, A. Trace Elements in Soils and Plants, 2nd ed.; CRC Press: Boca Raton, FL, USA, 1992. [Google Scholar]
- Pereira, S.I.A.; Barbosa, L.; Castro, P.M.L. Rhizobacteria isolated from a metal-polluted area enhance plant growth in zinc and cadmium-contaminated soil. Int. J. Environ. Sci. Technol. 2015, 12, 2127–2142. [Google Scholar] [CrossRef]
- Wallinga, I.; Vark, W.; Houba, V.J.G.; Lee, J.J. Plant Analysis Procedures. In Syllabus; Department of Soil Science and Plant Nutrition, Wageningen Agricultural Univerdsity: Wageningen, The Netherlands, 1998. [Google Scholar]
- Mani, D.; Kumar, C.; Kumar Patel, N. Integrated micro-biochemical approach for phytoremediation of cadmium and zinc contaminated soils. Ecotoxicol. Environ. Saf. 2015, 111, 86–95. [Google Scholar] [CrossRef]
- Moreira, H.; Marques, A.P.G.C.; Franco, A.R.; Rangel, A.O.S.S.; Castro, P.M.L. Phytomanagement of Cd-contaminated soils using maize (Zea mays L.) assisted by plant growth-promoting rhizobacteria. Environ. Sci. Pollut. Res. 2014, 21, 9742–9753. [Google Scholar] [CrossRef]
- de Koe, T. Agrostis castellana and Agrostis delicatula on heavy metal and arsenic enriched sites in NE Portugal. Sci. Total Environ. 1994, 145, 103–109. [Google Scholar] [CrossRef]
- Turner, S.; Pryer, K.M.; Miao, V.P.W.; Palmer, J.D. Investigating deep phylogenetic relationships among cyanobacteria and plastids by small subunit rRNA sequence analysis. J. Eukaryot. Microbiol. 1999, 46, 327–338. [Google Scholar] [CrossRef]
- Kisand, V.; Cuadros, R.; Wikner, J. Phylogeny of culturable estuarine bacteria catabolizing riverine organic matter in the northern Baltic Sea. Appl. Environ. Microbiol. 2002, 68, 379–388. [Google Scholar] [CrossRef]
- Paulo, A.M.S.; Amorim, C.L.; Costa, J.; Mesquita, D.P.; Ferreira, E.C.; Castro, P.M.L. Long-term stability of a non-adapted aerobic granular sludge process treating fish canning wastewater associated to EPS producers in the core microbiome. Sci. Total Environ. 2021, 756, 144007. [Google Scholar] [CrossRef]
- Vierheilig, H.; Coughlan, A.P.; Wyss, U.; Piché, Y. Ink and vinegar, a simple staining technique for arbuscular-mycorrhizal fungi. Appl. Environ. Microbiol. 1998, 64, 5004–5007. [Google Scholar] [CrossRef]
- Giovannetti, M.; Mosse, B. An Evaluation of Techniques for Measuring Vesicular Arbuscular Mycorrhizal Infection in Roots. New Phytol. 1980, 84, 489–500. [Google Scholar] [CrossRef]
- Alaboudi, K.A.; Ahmed, B.; Brodie, G. Phytoremediation of Pb and Cd contaminated soils by using sunflower (Helianthus annuus) plant. Ann. Agric. Sci. 2018, 63, 123–127. [Google Scholar] [CrossRef]
- Meers, E.; Ruttens, A.; Hopgood, M.; Lesage, E.; Tack, F.M.G. Potential of Brassic rapa, Cannabis sativa, Helianthus annuus and Zea mays for phytoextraction of heavy metals from calcareous dredged sediment derived soils. Chemosphere 2005, 61, 561–572. [Google Scholar] [CrossRef]
- Nehnevajova, E.; Herzig, R.; Bourigault, C.; Bangerter, S.; Schwitzguébel, J.-P. Stability of Enhanced Yield and Metal Uptake By Sunflower Mutants for Improved Phytoremediation. Int. J. Phytoremediation 2009, 11, 329–346. [Google Scholar] [CrossRef]
- Kloke, A.; Sauerbeck, D.R.; Vetter, H. The Contamination of Plants and Soils with Heavy Metals and the Transport of Metals in Terrestrial Food Chains. In Changing Metal Cycles and Human Health: Report of the Dahlem Workshop on Changing Metal Cycles and Human Health; Nriagu, J.O., Ed.; Springer-Verlag: Berlin/Heidelberg, Germany, 1984; pp. 113–141. [Google Scholar] [CrossRef]
- Tomas, M.; Pagani, M.A.; Andreo, C.S.; Capdevila, M.; Atrian, S.; Bofill, R. Sunflower metallothionein family characterisation. Study of the Zn(II)- and Cd(II)-binding abilities of the HaMT1 and HaMT2 isoforms. J. Inorg. Biochem. 2015, 148, 35–48. [Google Scholar] [CrossRef] [PubMed]
- Nehnevajova, E.; Lyubenova, L.; Herzig, R.; Schröder, P.; Schwitzguébel, J.P.; Schmülling, T. Metal accumulation and response of antioxidant enzymes in seedlings and adult sunflower mutants with improved metal removal traits on a metal-contaminated soil. Environ. Exp. Bot. 2012, 76, 39–48. [Google Scholar] [CrossRef]
- Tassi, E.; Pouget, J.; Petruzzelli, G.; Barbafieri, M. The effects of exogenous plant growth regulators in the phytoextraction of heavy metals. Chemosphere 2008, 71, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Moreira, H.; Pereira, S.I.A.; Vega, A.; Castro, P.M.L.; Marques, A.P.G.C. Synergistic effects of arbuscular mycorrhizal fungi and plant growth-promoting bacteria benefit maize growth under increasing soil salinity. J. Environ. Manag. 2020, 257, 109982. [Google Scholar] [CrossRef]
- Luo, Y.M.; Christie, P.; Baker, A.J.M. Soil solution Zn and pH dynamics in non-rhizosphere soil and in the rhizosphere of Thlaspi caerulescens grown in a Zn/Cd-contaminated soil. Chemosphere 2000, 41, 161–164. [Google Scholar] [CrossRef]
- Leung, H.M.; Wang, Z.W.; Ye, Z.H.; Yung, K.L.; Peng, X.L.; Cheung, K.C. Interactions Between Arbuscular Mycorrhizae and Plants in Phytoremediation of Metal-Contaminated Soils: A Review. Pedosphere 2013, 23, 549–563. [Google Scholar] [CrossRef]
- de Quadros, P.D.; Zhalnina, K.; Davis-Richardson, A.G.; Drew, J.C.; Menezes, F.B.; Camargo, F.A.O.; Triplett, E.W. Coal mining practices reduce the microbial biomass, richness and diversity of soil. Appl. Soil Ecol. 2016, 98, 195–203. [Google Scholar] [CrossRef]
- Fierer, N.; Bradford, M.A.; Jackson, R.B. Toward an ecological classification of soil bacteria. Ecology 2007, 88, 1354–1364. [Google Scholar] [CrossRef]
- Ivanova, A.A.; Zhelezova, A.D.; Chernov, T.I.; Dedysh, S.N. Linking ecology and systematics of acidobacteria: Distinct habitat preferences of the Acidobacteriia and Blastocatellia in tundra soils. PLoS ONE 2020, 15, e0230157. [Google Scholar] [CrossRef]
- Liu, P.; Jia, S.; He, X.; Zhang, X.; Ye, L. Different impacts of manure and chemical fertilizers on bacterial community structure and antibiotic resistance genes in arable soils. Chemosphere 2017, 188, 455–464. [Google Scholar] [CrossRef]
- Hu, Q.; Tan, L.; Gu, S.; Xiao, Y.; Xiong, X.; Zeng, W.A.; Feng, K.; Wei, Z.; Deng, Y. Network analysis infers the wilt pathogen invasion associated with non-detrimental bacteria. NPJ Biofilms Microbiomes 2020, 6, 8. [Google Scholar] [CrossRef]
- Nouioui, I.; Göker, M.; Carro, L.; Montero-Calasanz, M.d.C.; Rohde, M.; Woyke, T.; Kyrpides, N.C.; Klenk, H.P. High quality draft genome of Nakamurella lactea type strain, a rock actinobacterium, and emended description of Nakamurella lactea. Stand. Genom. Sci. 2017, 12, 4. [Google Scholar] [CrossRef]
- Asaf, S.; Numan, M.; Khan, A.L.; Al-Harrasi, A. Sphingomonas: From diversity and genomics to functional role in environmental remediation and plant growth. Crit. Rev. Biotechnol. 2020, 40, 138–152. [Google Scholar] [CrossRef]
- Gallo, I.F.L.; Furlan, J.P.R.; Sanchez, D.G.; Stehling, E.G. Heavy metal resistance genes and plasmid-mediated quinolone resistance genes in Arthrobacter sp. isolated from Brazilian soils. Antonie Van Leeuwenhoek 2019, 112, 1553–1558. [Google Scholar] [CrossRef]
- Li, C.; Quan, Q.; Gan, Y.; Dong, J.; Fang, J.; Wang, L.; Liu, J. Effects of heavy metals on microbial communities in sediments and establishment of bioindicators based on microbial taxa and function for environmental monitoring and management. Sci. Total Environ. 2020, 749, 141555. [Google Scholar] [CrossRef]
- Nardi, P.; Laanbroek, H.J.; Nicol, G.W.; Renella, G.; Cardinale, M.; Pietramellara, G.; Weckwerth, W.; Trinchera, A.; Ghatak, A.; Nannipieri, P. Biological nitrification inhibition in the rhizosphere: Determining interactions and impact on microbially mediated processes and potential applications. FEMS Microbiol. Rev. 2020, 037, 874–908. [Google Scholar] [CrossRef]
- Juan, Z.; Shi, Q.; Fan, S.; Zhang, Y.; Zhang, M.; Zhang, J. Distinction between Cr and other heavy–metal–resistant bacteria involved in C/N cycling in contaminated soils of copper producing sites. J. Hazard. Mater. 2021, 402, 123454. [Google Scholar] [CrossRef]
- Chaney, R.L. Toxic Element Accumulation in Soils and Crops: Protecting Soil Fertility and Agricultural Food-Chains. In Inorganic Contaminants in the Vadose Zone; Springer: Berlin/Heidelberg, Germany, 1989; pp. 140–158. [Google Scholar] [CrossRef]
- Underwood, E.J.; Suttle, N.F. The Mineral Nutrition of Livestock, 3rd ed.; CAB International: Wallingford, CT, USA, 1999. [Google Scholar]
- Marques, A.P.G.C.; Caetano, N.S.C.; Castro, P.M.L. Chapter 12: Strategies for Enhancing Soil Phytoremediation and Biomass Valorization. In Handbook of Environmental Remediation: Cloassic and Modern Techniques; Hussein, C.M., Ed.; Royal Society of Chemistry: London, UK, 2020; pp. 331–356. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).