Abstract
The phospholipid fatty acid method was used to determine the shifts in microbial biomass due to irrigation with reverse-osmosis (RO) concentrate (or highly saline reject water) and brackish groundwater (BGW). In this greenhouse study, RO concentrate and BGW were applied to irrigate pecan trees for 8 months for two consecutive seasons. The objectives of the study were to (i) evaluate how irrigation with RO concentrate and BGW impacts soil microbial composition in pecan rhizospheres using microbial phospholipid fatty acid (PLFA) biomarkers as indicators, and (ii) evaluate its implications on soil health. Three treatments of RO concentrate (EC = 8.0 dS/m), BGW (EC = 4.0 dS/m), and the city of Las Cruces’s water (EC = 0.8 dS/m) as a control were used to irrigate pecan trees. EC, pH, and organic matter (OM%) content of the soil samples were measured, and PLFA biomarkers for the microbial community were determined. Na-, Cl-, and K-ion concentrations were 26.16, 32.54, and 5.93 meq/L in 2017 and 25.44, 24.26, and 5.49 meq/L in 2018, respectively, in RO irrigation pots. For two seasons, gram-positive bacteria were dominant, while gram-negative bacteria were not detected in the second season. PLFA biomarkers of fungi were found among all three treatments in the first season; however, they appeared only with BGW in the second season. Actinomycetes were recorded in the first season while they were not seen in the second season. Increases in soil salinity and microbial shifts could have important implications for soil health. Irrigating with RO and BGW shifted the soil microbial composition; therefore, long-term irrigation with BGW and RO concentrate would be deleterious for pecan production and soil health.
1. Introduction
Natural soil microbes are essential for longevity and soil health, which entails a balance between the chemical, physical, and biological characteristics of the soil [1]. Soil microbial communities perform important roles in cycling nutrients from soil organic matter and maintaining plant productivity [2]. Microbes contribute to maintaining the physical and biochemical characteristics necessary for soil fertility by converting organic matter, depending on the amount of it in the soil, into simple, absorbable nutrients. [3]. Nutrient availability to plants is regulated by the rhizosphere microbial activity [4]. Thus, any factor influencing the microbial community and its activities will have an impact on the soil’s quality, the availability of nutrients, as well as the growth of plants. Salinity is even more influential in changing microbial composition than pH or temperature [4]. This is because higher salt concentrations outside of microbial cells have an adverse physiological impact, which causes the microbial cells to become dehydrated [5] due to lower soil osmotic potential and decreased water availability [6].
In saline soil, it has been observed that the ratio of bacteria to fungi rises with decreasing fungal biomass [7]. Saline soil negatively affects microbial enzymatic activities [8], respiration, and soil microbial biomass [9] and decreases soil organic matter [10]. Furthermore, microbial diversity has been reported to linearly decrease with increasing salinity in Gurbantunggut desert soil [11]. According to Zhang et al. [10], the efficiency and abundance of bacteria, arbuscular mycorrhizal fungi, and other microorganisms were all significantly impacted by increasing soil salinity.
Different groups of microbes contain different fatty acids in their structural living fat cells [12]; therefore, the PLFA method was used to monitor the formation of microbial communities in different environments, such as agricultural soils. PLFAs are present in the living cell membranes of the microbes [13]; therefore, a shift in the microbial composition is an indicator of the negative changes in the surrounding conditions [14]. PLFA analysis has been used to assess and identify microbial biomass and structure [15]. PLFA biomarker analysis revealed a broad and diversified spectrum of soil microbial composition in seawater-irrigated soil [6]. PLFA analysis in a study on the effects of salinity above the threshold of 31.7 dS/m on a mangrove forest found no shift in microbial community structure; however, increased microbial biomass–prokaryotes were observed due to flooding [6]. Irrigation with saline water showed that the bacterial/fungal PLFA ratio increased with the addition of soil nitrogen and phosphorus contents [16]. Another study reported that PLFA evaluation was not affected by salinity [13]. Chowdhury et al. [12] applied various levels of salinity (EC 0.7, 9.3, and 17.6 dS/m) in the presence of organic matter and found that high salinity levels decreased fungal PLFAs. On the contrary, increases in the fungal PLFA biomarker (18:2w6,9) were the trend with higher soil salinity levels (22.9 dS/m) [17]. Increases in PLFAs of gram-negative and gram-positive bacteria were observed in a brackish-water-irrigated mangrove peat soil in Florida [6], likely impacting the soil health.
Soil health is inferred from a combination of physicochemical and biological properties of the soil that assist plant productivity and growth. These factors are quantified to assess soil health correctly [18]. Kumar et al. [19] mentioned that the amounts of soil organic matter and nutrients are the most important indicators when considering plant yield and soil health. Salinity stress has been identified as one of the main reasons for declining soil health and plant growth due to ion toxicity, osmotic stress, nutrient deficiency, and microbial diversity imbalance [20,21]. Salinity is reported to reduce the root uptake of nitrogen, phosphorus, and potassium as essential plant nutrients and water due to osmotic pressure [21,22,23]. The interaction between plants and soil microbes, which affects the availability of nutrients, is essential to maintaining soil health, microbe diversity, and crop productivity under duress [19,24]. Soil microbes’ diversity plays a vital role in improving plant growth and soil health, ameliorating biotic and abiotic stresses and enhancing crop productivity [25,26]. According to Kumar et al. [19] and Kumar et al. [21], soil health and crop growth can be improved by certain salinity-resistant microbes such as Achromobacter, Arthrobacter, Bacillus, Chryseobacterium Enterobacter, Ochrobactrum, and Pseudomonas. By fixing atmospheric nitrogen, solubilizing phosphate, and releasing nutrients, cyanobacteria also increase soil fertility and crop yield [27].
All the studies reported above utilized soils that were already saline, or NaCl was added to the soil to create a gradient of salinity. To the best of our knowledge, no study used irrigation with reverse osmosis (RO) concentrate (or highly saline reject water, a byproduct of brackish groundwater desalination) and brackish groundwater (BGW) to investigate the effects on soil health and microbial composition using the phospholipid fatty acid (PLFA) analysis method.
Due to a lack of information on the effects of calcium-dominant saline water on soil microbial composition, this study focused on utilizing calcium-dominant brackish groundwater and RO concentrate. In addition, this study was conducted for two consecutive growth seasons. The present study hypothesizes that irrigation with brackish groundwater and RO concentrate would decrease the microbial composition in the soil and negatively affect soil health. The objective of this study is to evaluate the irrigation-water salinity impacts on soil microbial composition and soil health in pecan rhizospheres using microbial phospholipids fatty acids as indicators.
2. Materials and Methods
2.1. Site Description and Plant Material
This research was carried out in a greenhouse over the course of two seasons, from May of 2017 to December of 2018, at the Fabian Garcia Science Center of New Mexico State University in Las Cruces, New Mexico (32°16’704” N, 106°46’376” W). The roof of the greenhouse is made entirely of plastic, and the building also includes an evaporative air conditioner, an electric heater, and a temperature-regulating system. Four-year-old pecan trees (Carya illinoinensis L., variety Wichita; Archer Farms, Inc., Las Cruces, NM, USA) were grown in cylindrical pots measuring 0.035 m3, 36 cm in height, and 36 cm in diameter in May 2017 [28]. The cultivation media was classified as sandy loam soil by the United States Department of Agriculture classification system (USDA) [29].
2.2. Experimental Design and Treatments
Twelve pots were randomly distributed in a completely randomized design. Randomization was accomplished by generating random numbers in MS Excel (2013). The pecan trees were irrigated with three different levels of saline water treatments in four replicates. The three treatments were Fabian Garcia (city) water (control, EC = 0.8 dS/m), reverse osmosis extracts (RO), and brackish groundwater (BGW; EC = 4.0 dS/m) [28]. The soil absorption ratio SAR for all irrigation water was <5.1; some cation and anion concentrations are shown in Table 1. BGW and RO concentrate were provided by the Brackish Groundwater National Desalination Research Facility in Alamogordo (32°53’04” N, 105°58’37” W). A 7 L plastic bucket was used to irrigate the pecan trees every 14 days for 15 irrigations throughout the growing season.

Table 1.
Magnesium (Mg), calcium (Ca), sodium (Na), chloride (Cl), phosphorus (P), potassium (K), and sulfur (S) ion concentrations (meq/L), SAR, and pH for the irrigation waters.
The same volumes of soil and deionized water were combined and then passed through a piece of clothing to determine the electrical conductivity (dS/m) and pH of the soil using portable probes [30]. The paste extract was then collected in a glass tube, and measurements were made by dipping the probes in the solution. Soil samples were air-dried, ground, and bagged in plastic bags for analysis of soil ion concentrations and organic matter (OM) by AgSource laboratory (Lincoln, NE, USA) using the loss-on-ignition technique [31]. Soil health impacts were assessed using soil chemical and microbial data.
2.3. Soil Samples Preparation
Soil samples were collected from the Fabian Garcia Research Center of New Mexico State University in early January at the end of every season. A soil probe was used to collect soil samples from 0–25 cm depth. Samples were then bagged in plastic bags and frozen at −20 °C until they were analyzed after one week.
2.4. Samples Drying and Lipid Extraction
The method applied was published by Buyer and Sasser [32]. Two grams of wet sieved sandy loam soil was placed in every 13 mm × 100 mm screw-cap tube of known weight. After freezing in liquid nitrogen, samples were dried in a vacuum (Labconco, freeze-dry system. Labconco Corporation, Kansas City, MO, USA) overnight, and dry weight was obtained. Four mL of Bligh–Dyer extractant consisting of 200 mL of 50 mM K2HPO4 in H2O, 500 mL methanol, and 250 mL chloroform was mixed with the internal standard (19:0 phosphatidylcholine, Avanti Polar Lipids, Alabaster, AL, USA) and added to every tube. The tubes were sonicated in an ultrasonic cleaning bath (Bransonic 2210, Branson Ultrasonics Corporation, Danbury, CT, USA) for 10 min then shacked using (Apollo HP50 shaker oven. Amerex Instruments, Inc. Concord, CA, USA) for 2 h. After centrifuging for 10 min using (Beckman model J-6B. Phoenix Equipment Inc, Phoenix, USA), the liquid phase was transferred to another clean 13 mm × 100 mm tube. One mL chloroform and one mL distilled water were added to every tube. The tubes were vortexed for 5 s. The samples were centrifuged for 10 min, and the upper phase was removed and discarded. The lower phase containing the extracted lipids was evaporated using the nitrogen gas flow (The Meyer N-EVAP, Analytical evaporator, Organomation Association, Berlin, MA, USA), and the samples were stored overnight at −20 °C.
2.5. Lipids Separation
Solid-phase extraction (SPE) was used for lipid separation as described by Christie [33] with the following modifications using a silica gel-packed glass Pasteur pipette. The glass Pasteur pipettes were packed with a small amount of untreated glass wool (Ohio Valley specialty chemical, OH, USA) then 50 mg of silica gel (SIELC Technologies, Inc., Prospect Heights, IL, USA) was added to every Pasteur pipette. Each Pasteur pipette was equipped with 3 mL methanol followed by 3 mL chloroform. The stored dry sample was dissolved in 1 mL chloroform and passed through silica. After that, 1 mL of chloroform followed by 1 mL of acetone was added to wash the Pasteur pipette. After eluting with 0.5 mL of 5:5:1 methanol: chloroform: H2O, the phospholipids were collected in 1.5 mL screwcap glass vials (Multi-Tier microplate, E&K Scientific, Santa Clara, CA, USA). The resulting solution was evaporated in a speed vacuum for 30 min at 60 °C until dried.
2.6. Fatty Acids Transesterification
As mentioned by Buyer and Sasser [32], a 0.2 mL transesterification reagent (0.561 g KOH dissolved in 75 mL methanol and 25 mL toluene) was added to each lipid residue. The tubes were closed with PTFE/silicon caps (E&K Scientific, Santa Clara, CA, USA) and incubated at 37 °C for 15 min. After that, acetic acid (0.075 moles) followed by 0.4 mL chloroform was added to each vial, shaken for 10 s, and kept stable in a rack for a while, allowing the phases to separate. Using a pipette, 0.3 mL of the liquid bottom was removed to a new 1 mL vial. This process was repeated, but 0.4 mL was removed and combined with previous extractions. The chloroform was evaporated in a sped vacuum at room temperature. The samples were dissolved in 75 µL hexane and transferred to gas chromatography vials containing conical glass and stored at −20 °C.
2.7. PLFA Analysis
Gas chromatography (Agilent Technologies, Wilmington, DE, USA) was performed on a Varian 3900 equipped with an FID detector as previously described by Buyer and Sasser [32]. An Agilent Ultra 2 column—25 m in length, 0.2 mm internal diameter, and 0.33 µm film thickness—was used to separate the fatty acid methyl esters, or FAMEs. A split ratio of 30:1 was used with hydrogen gas at a 1.2 mL/min flow rate. The injector temperature was 250 °C and the detector temperature was 300 °C. Fatty acid methyl esters (FAMEs) were identified using relative retention time to ISTD [33]. The data was recorded for each season. The recovery was around 83.04%. The limit of detection LOD ranged from 2 to 6 ug/mL, and the limit of quantification LOQ ranged from 6 to 17 ug/mL. The fatty acids i14:0, i15:0, a15:0, i16:0, i17:0, a17:0, i18:0, and 10Me16:0 were used as indicators of gram-positive bacteria PLFAs while 15:03OH, 15:1w6c, 16:12OH, 16:1w57 9c, 17:1w8c, 18:1w7 9c, and cy17:0 PLFAs were indicators of gram-negative bacteria PLFAs [34]. The fatty acids 18:2w6 9c, 20:1w9c, 18:1w9c, and 16:1w5c were used as indicators of fungal PLFAs; 10Me16:0, 10Me17:0, and 10Me18:0 were used as indicators of actinomycete PLFAs; and 16:1w5c was the indicator of AM fungi [34].
2.8. Data and Statistical Analysis
PLFA biomass (nmol/g) was calculated for bacteria, fungi, actinomycetes, and other eukaryotes. Differences due to treatments were determined using a one-way analysis of variance (ANOVA). Soil pH, EC, and OM data were analyzed using a one-way analysis of variance (ANOVA). The statistical analysis was performed using SAS software, v 9.4 (SAS Institute Inc. Cary, NC, USA). Means were separated by the least significant difference (LSD) and were considered significant at α = 0.05.
3. Results and Discussion
Soil electrical conductivity and pH are shown in Table 2. For the 2017season, it can be observed that the highest EC was for the soil irrigated with RO concentrate (6.60 ± 0.03 dS/m), followed by the BGW-irrigated soil (4.80 ± 0.02 dS/m). The soil pH ranged from 8.60 ± 0.10 for control treatment to 7.53 ± 0.14 for RO-concentrate-irrigated soil.
Table 2.
Mean and standard error of soil electrical conductivity EC (dS/m), pH, and organic matter OM (%) for three treatments and two seasons of 2017 and 2018.
The 2018 season showed the highest EC for soil irrigated with RO concentrate (7.30 ± 0.03 dS/m) followed by BGW (4.81 ± 0.01 dS/m) (Table 2). A similar trend was observed in soil pH with the control treatment displaying the highest pH (7.93 ± 0.13) followed by BGW (7.73 ± 0.06), and RO-concentrate-irrigated soil had the lowest pH (7.53 ± 0.14) (Table 2). Although most of the references indicated no direct relation between pH and EC [35], our results showed a slight, non-significant reduction in pH with increasing salinity. In contrast to our results, in reference [10], increases in soil pH were reported because of applying seawater versus non-saline water. This could be due to the difference between our irrigation water, which was Ca-ion dominant, and the seawater.
Decreases in soil pH between the two seasons were more pronounced. In contrast, EC showed increases in the second season in 2018 (Table 2). This suggested that the microbial composition and activities could be affected by the changes in pH and EC. Reduction in microbial composition was observed as a result of the increasing soil EC irrigated with seawater compared with non-saline soil [6].
Soil irrigated with RO concentrate had the lowest OM content (0.86 ± 0.31%) followed by BGW (0.93 ± 0.18%), while the highest OM content was for soil irrigated with the control treatment (1.3 ± 0.05%) in the first season, 2017. In the second season, the highest percentage of OM content was for the soil irrigated with BGW (2.13 ± 0.90%) followed by the control (1.66 ± 0.41%), whereas the lowest percentage OM content was for the soil irrigated with RO concentrate (0.76 ± 0.14%). For the two seasons of the experiment, the soil irrigated with RO concentrate had the lowest percentage of OM. In addition, this clearly showed that increases in soil salinity decreased soil organic matter content, which was also reflected in the microbial biomass and diversity.
These results agree with a previous study where organic matter and microbial biomass were reported to decrease with increasing salinity [10]. According to the USDA, our three treatments were considered as between slightly and moderately saline. Salinity can decrease soil organic matter by enhancing the activity of carbon-degrading extracellular enzymes and affecting soil bacterial communities [10].
In the first season with an EC of 2.60 dS/m and pH of 8.0, the PLFA biomass of gram-positive bacteria of RO-concentrate irrigation presented as 15:0iso, 15:0anteiso, 16:0iso, 16:0 10-methyl, and 17:0anteiso was the highest; followed by BGW irrigation, which presented as 15:0iso, 15:0anteiso, 16:0 10-methyl, and 17:0anteiso; compared with the control treatment, which presented as 15:0iso, 15:0anteiso, 16:0iso, and 16:0 10-methyl, showing the bacterial ability to tolerate the osmotic stress caused by salinity (Table 3) (Figure 1). This was the opposite with PLFAs of gram-negative bacteria, which presented as 16:1w5c, 17:1w8c, and 18:1w7c compared with the control treatment, which presented as 16:1w5c and 18:1w7c, representing the effects of RO concentrate on the microbial composition (Table 3) (Figure 1). This trend partially agreed with another study where both gram-positive and gram-negative bacteria increased with increasing salinity and suggested that gram-negative bacteria has more tolerance for salinity [6]; in contrast in the present study, gram-positive bacteria were more tolerant than gram-negative bacteria and remained for the two seasons. This could be due to the different structures of the cell membrane that provide gram-positive bacteria with an advantage over gram-negative bacteria in salinity tolerance.
Table 3.
Phospholipid fatty acid (PLFA) biomarkers of gram-negative bacteria, gram-positive bacteria, AM fungi, fungi, and actinomycetes for three treatments and two seasons of 2017 and 2018.
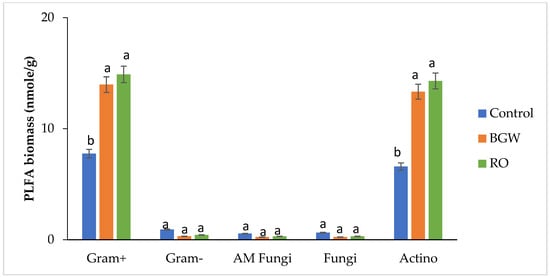
Figure 1.
Phospholipid fatty acid (PLFA) biomass of gram-positive bacteria, gram-negative bacteria, AM fungi, fungi, and actinomycetes (nmol/g); measurements of three treatments with four replications for the first season, 2017. Lowercase letters correspond to a statistically significant difference in measurements between treatments at ∝ = 0.05. Control = 0.8 dS/m, brackish groundwater BGW= 4.0 dS/m, and RO concentrate = 8.0 dS/m. nmol/g = nanomoles per gram.
In the second season, however, with an EC of 2.84 dS/m and pH of 7.53, decreases in gram-positive PLFA biomass presented as 18:0iso with RO-concentrate irrigation, whereas BGW irrigation’s higher PLFA biomass presented as 18:0iso, and the control irrigation presented with the same PLFA biomarker (Table 3) (Figure 2). In this second season, gram-negative bacteria completely disappeared from all three treatments (Table 3) (Figure 2).
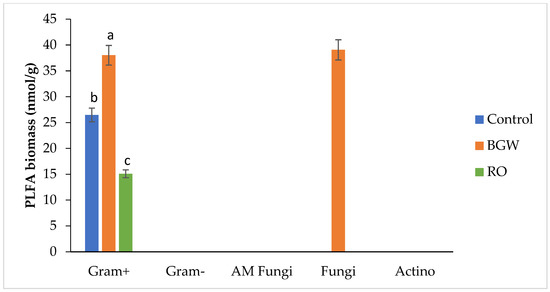
Figure 2.
Phospholipid fatty acid (PLFA) biomass of gram-positive bacteria, gram-negative bacteria, AM fungi, fungi, and actinomycetes (nmol/g); measurements of three treatments with four replications for the second season, 2018. Lowercase letters correspond to a statistically significant difference in measurements between treatments at ∝ = 0.05. Control = 0.8 dS/m, brackish groundwater BGW = 4.0 dS/m, and RO concentrate = 8.0 dS/m. nmol/g = nanomoles per gram.
The PLFA biomass of AM fungi presented as 16:1w5c, slightly decreased with RO-concentrate and BGW irrigation compared to the control treatment in the first season (Table 3) (Figure 1). In the second season, AM fungi did not appear with all three treatments (Table 3) (Figure 2). These results agree with the findings of [10], where the AM fungi biomass decreased in coastal saline soils. As reported by Zhang et al. [10], the decrease in AM fungi negatively affected the host plant’s nutrition and physiology. In addition, this could be one of the reasons—in combination with increases in salinity—of the field observations on pecan tree leaves that showed signs of damage such as edge-burning, small yellowish leaves compared with the control trees because of increasing salinity (Figure 3).

Figure 3.
Pecan leaves showing signs of increasing salinity.
The PLFA biomass of fungi with the control irrigation, presented as 16:1w5c and 18:2w6c, was higher than the PLFA biomass of fungi with RO-concentrate irrigation, which presented as 16:1w5c in the first season (Table 3) (Figure 1). A similar decrease was reported in a fungi PLFA biomass with higher salinity levels [12]. The results of the second season presented the PLFAs of fungi as 18:2 w6c and 20:1 w9c only with BGW irrigation (Table 3) (Figure 2).
Mohamed and Martiny [36] reported that with a soil salinity gradient, the fungal composition has changed on marshland. Zhang et al. [10] reported decreases in the fungal Hydropisphaera species, which is responsible for lignin degradation in coastal soils, affecting the nutrient availabilities in the soil. This is what happened in this study, where fungi and organic matter decreased by increasing the salinity gradient. Another study disagreed with our findings where the fungal biomass was higher with increasing NaCl levels in the soil [17].
Irrigation with RO concentrate showed the highest PLFA biomass of actinomycetes, presenting as 16:0 10-methyl and 18:0 10-methyl, followed by BGW irrigation, compared with the PLFA biomass with the control irrigation, which presented as 16:0 10-methyl in the first season (Table 3) (Figure 1). The PLFA biomarker 18:0 10-methyl has been reported to increase with saline soil [15], and this is similar to the present study. The second season showed a disappearance in actinomycetes among all three treatments (Table 3) (Figure 2). This confirms the observation of [6], who reported that actinomycetes represent a small fraction of the soil microbial composition.
The not-assigned microbial PLFA biomass showed the greatest number of other eukaryotes with RO-concentrate irrigation compared with the PLFA biomass with the BGW and control irrigation treatments in the first season (Figure 4). Decreases in not-assigned PLFA microbial biomass with RO irrigation were more pronounced in the second season compared to the BGW-irrigated soil, while BGW irrigation showed the highest amount of not-assigned PLFA biomass compared with the control treatment (Figure 5). Among the three treatments, the total PLFA microbial biomass was the greatest with RO-concentrate irrigation, followed by the total PLFA biomass with BGW irrigation, while the control treatment showed the lowest total PLFA microbial biomass in the first season (Figure 4). This trend disagreed with another study that reported that total biomass was not affected by increasing salinity [13], while [37] improved the effects of salinity in the total microbial biomass. BGW irrigation showed the highest total PLFA microbial biomass, while RO-concentrate irrigation biomass decreased in the second season (Figure 5). This could be related to the decrease in OM content to 0.76 ± 0.14% with RO-concentrate irrigation, which was less than the OM content 2.13 ± 0.90% for BGW irrigation (Table 2).
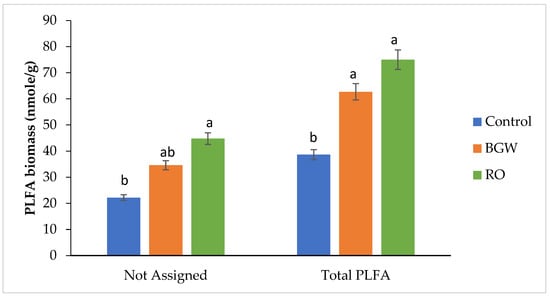
Figure 4.
Phospholipid fatty acid (PLFA) biomass of not-assigned and total PLFA (nmol/g) measurements of three treatments with four replications for the first season, 2017. Lowercase letters correspond to a statistically significant difference in measurements between treatments at ∝ = 0.05. Control = 0.8 dS/m, brackish groundwater BGW= 4.0 dS/m, and RO concentrate = 8.0 dS/m. nmol/g = nanomoles per gram.
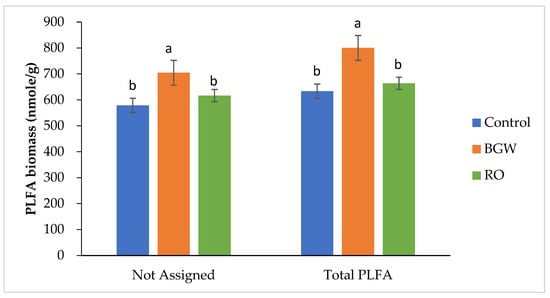
Figure 5.
Phospholipid fatty acid (PLFA) biomass of not-assigned and total PLFA (nmol/g) measurements of three treatments with four replications for the second season, 2018. Lowercase letters correspond to a statistically significant difference in measurements between treatments at ∝ = 0.05. Control = 0.8 dS/m, brackish groundwater BGW= 4.0 dS/m, and RO concentrate = 8.0 dS/m. nmol/g = nanomoles per gram.
Soil quality, productivity, and organic matter are strongly related to microbial community composition. Every component of the microbial community has a role in the soil ecosystem. According to Hoorman [38], gram-positive bacteria, gram-negative bacteria, and actinomycetes concentrate their biomass in the rhizosphere zone. In the rhizosphere, the interaction between plants and bacteria (nitrogen fixation) assists soil and plant health by making a mutualist relationship with plant roots. In general, as stated by Zhang et al. [37], bacteria have more advantages over fungi because of their higher activity. In this study, with RO-concentrate irrigation, the soil microbial community shifted to a bacterial community, as the gram-positive bacteria were the dominant species in the second season, with increases in soil salinity levels. This, consequently, would affect the role of bacteria in decomposing the simple sugars and carbon compounds. Moreover, bacteria can improve soil structure by producing polysaccharides, which are important for cementing clay, silt, and sand [38].
In the present study, as the results showed, the decrease in fungi biomass with RO-concentrate irrigation caused a decrease in lignin decomposition because fungi are responsible for the lignin degradation; however, the BGW showed increases in fungal biomass in the second season, meaning that the degradation of lignin was higher with BGW irrigation than with the RO and control treatments. The decreases in AM fungi in the first season followed by the absence of the AM fungi in the second season could negatively affect plant nutrition by decreasing the potassium and phosphorus uptake and negatively affecting plant survival and growth. This is confirmed by [10,39] where some AM fungal species such as Rhizophagus intraradices, Claroideoglomus etuncatum, and Septoglomus constrictum can assist plant growth under saline conditions.
On the other hand, actinomycetes have the same role as fungi because of their ability to decompose hard organic matter such as lignin [38]. As shown in Figure 1 and Table 3, the high actinomycete PLFA biomass observed suggests that the breakdown of lignin with RO-concentrate and BGW irrigation was higher than with the control irrigation. However, this advantage disappeared in the second season with the disappearance of the actinomycetes with all three treatments (Figure 2).
Our results were in line with a previous study where reductions in microbial biomass, total PLFA, bacterial biomass, and fungal biomass were recorded because of applying 4.61 and 8.04 dS/m saline water [40]. Shifting toward bacterial abundance is a result of applying different concentrations of seawater [6]. Another study concluded the same when it was observed that high salinity (24 dS/m) inhibited the growth of microbial biomass [41]. Xie et al. [42] stated that these conflicting observations might be attributed to soil texture, nutrient availability, microbial acclimation to salt stress, and quantity of soluble salts. Lynch and Whipps [43] stated that microbial growth in the soil is mostly carbon-limited; therefore, the organic matter has a great influence on microbial composition. Moreover, the specific microbiota of many plants suggests that microbial diversity increases with increasing plant diversity and therefore the heterogeneity for the microorganisms, as well. This might explain the disappearances of several microbial species in the second season even with the control treatment since there was no plant diversity and the organic matter was limited and affected by the salinity in the pots.
Table 4 and Table 5 show the concentrations of Na, Mg, Ca, S, Zn, and Cl ions and SAR in the soil in the 2017 and 2018 seasons. The data recorded from both seasons showed increases in sodium, magnesium, calcium, sulfur, zinc, and chloride ions with increasing water salinity; however, potassium significantly decreased by 5.93 ± 0.56 meq/L with the increasing irrigation-water salinity level in the first season (Table 5). Significant increases were observed in Na-ion concentrations in the pots irrigated with RO concentrate by 26.16 ± 3.43–25.44 ± 4.26 meq/L for the 2017 and 2018 seasons, respectively (Table 4). Similarly, the Mg-ion concentration increased in the pots irrigated with RO concentrate to 32.23 ± 13.15–32.11 ± 4.83 meq/L for the 2017 and 2018 seasons, respectively (Table 4). In agreement with our results, a previous study reported increases in Na- and Mg-ion concentrations in a soil irrigated with increasing irrigation-water salinity up to 8.0 dS/m [44]. Another study reported decreases in Ca, P, and Zn upon irrigation with 4 dS/m KCl, NaCl, and CaCl2 saline water [45]. The greatest ion concentration was calcium with RO-concentrate irrigation at 164.15 ± 38.31–32.11 ± 4.83 meq/L for the two seasons, respectively (Table 4). Since the EC of the soil was >4.0 dS/m and the SAR ratio was <13 in BGW- and RO-concentrate-irrigated soil, the soil was considered saline but not sodic for the two seasons (Table 4).
Table 4.
Mean and standard error of soil Na-, Mg-, and Ca-ion concentrations (meq/L), and SAR for three treatments and two seasons of 2017 and 2018.
Table 5.
Mean and standard error of soil S-, K-, Zn-, and Cl-ion concentrations (meq/L) for three treatments and two seasons of 2017 and 2018.
Sulfur-ion concentrations significantly increased to 52.88 ± 36.62–56.62 ± 16.48 meq/L in RO-irrigated pots in the 2017 and 2018 seasons, respectively, while K-ion concentrations decreased to 5.93 ± 0.56–5.49 ± 0.38 meq/L in RO pots in 2017 and 2018 seasons, respectively (Table 5). No differences in Zn-ion concentrations throughout the two seasons were found; however, the Cl-ion concentrations increased up to 32.54 ± 9.70–24.26 ± 7.39 meq/L in RO irrigation pots of the 2017 and 2018 seasons, respectively (Table 5).
Salinity was found to increase the concentration of sodium in soil, although it also reduced the amount of organic matter, soil K exchangeable, and microbial biomass [10,37]. These reported results are in assent with our findings where Na increased while the OM, K, and microbial biomass decreased with RO-concentrate irrigation pots. This, consequently, would negatively affect soil health and productivity. The K/Na ratio must be higher to reduce the negative effects of Na on soil microbial composition, and accordingly, soil health. As mentioned above, every microbe species has a role in the soil ecosystem; thus, the absence of such species would interrupt the biogeochemical processes, which are responsible for organic matter degradation and nutrient availabilities, and accordingly, soil health [38,40].
Previous studies suggested that several parameters should be examined for soil health assessment. Such parameters include texture, bulk density, hydraulic conductivity, salinity, OM, pH, and microbial activity [46,47]. In this study, we found that with increasing irrigation-water salinity, soil salinity increased. This increase in soil salinity would negatively affect the soil structure, decreasing soil porosity, water retention, and nutrient availability. Ikemura and Shukla [48] mentioned that soil with EC from 7 to 10 dS/m has severe limitations for productivity. In this study, the soil EC was higher than 7 dS/m in the pots irrigated with BGW and RO concentrate, and limitations in soil health led to decreases in pecan growth (Figure 3). A range of 0.5–1.0% soil organic matter content caused severe limitations in soil productivity [48]. Soil OM decreased with continuous RO irrigation to 0.76%, which could cause reductions in nutrient availability and microbial diversity that could adversely affect productivity and soil health. Previous studies mentioned reductions in soil nutrient availability, microbes, enzyme activities, and physiochemical properties due to decreases in soil OM content because of increases in soil salinity [49,50,51,52]. Specifically, as the K/Na ratio decreased in RO-irrigated soil, the availability of K for plants and microbes’ activities was inhibited, resulting in declining productivity. According to [19,53,54], increases in Na-ion concentrations in the soil led to changes in soil porosity, aeration, and water conductance, thereby decreasing nutrient mobility, microbial composition, and crop yield.
Bulk density was reported to increase with increasing soil salinity for soils irrigated with RO concentrate (EC around 8.0 dS/m) [29]. Increases in soil bulk density and decreases in infiltration rates were the result of saline-sodic drainage (EC 11.2 dS/m), irrigation with saline water [55], and irrigation with treated wastewater [56]. Greater decreases in soil hydraulic conductivity have been observed in clay than in sand soil irrigated with brackish groundwater (4.1 dS/m) or RO concentrate (8.0 dS/m) used to irrigate halophytic species [57]. According to [58,59], decreases in clay-loam soil irrigated with highly saline-sodic waters, 5.64 and 8.5 dS/m, respectively, were recorded with attendant decreases in soil macro-porosity. Thus, irrigation with RO concentrate has the potential to deteriorate productivity and soil health unless proper irrigation management is applied.
4. Conclusions
RO concentrate and BGW were used to irrigate pecan trees in a greenhouse. This caused shifts in the soil microbial composition considering PLFA biomarkers as indicators. Gram-positive bacteria were found to be dominant even with control, BGW, and RO-concentrate treatments. PLFAs of fungi appeared in both seasons; however, in the second, they appeared in a higher amount only with BGW. Soil organic matter decreased as the irrigation-water salinity increased. Na and Cl concentrations increased in the soils irrigated with BGW and RO concentrate. These changes would affect the pecan tree’s growth through the nutrient availabilities necessary for a healthy soil term.
This study shows how the shift in the PLFA microbial biomass due to salinity could affect the soil environment and be reflected in pecan tree growth. According to this study, the important role of micro-organisms in soil health and productivity becomes clear, as well as how soil microorganisms are affected by salinity, especially reverse-osmosis concentrate. Continuous irrigation with brackish groundwater and RO concentrate will cause a reduction in pecan growth, productivity, and soil health.
Author Contributions
Writing original draft, review, and edit (A.R.B.A.); technical support, review, and editing (F.F.O.H.); supervision, review, and editing (M.K.S.). All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by [Nakayama Professorship] grant number [904144], [Hatch] grant number [135433], and [NIFA] grant number [GR0006853].
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors thank Nakayama Professorship, NMSU Agricultural Experiment Station, NIFA, Hatch, and Brackish Groundwater National Desalination Research Facility in Alamogordo.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Moebius-Clune, B.N.; Moebius-Clune, D.J.; Gugino, B.K.; Idowu, O.J.; Schindelbeck, R.R.; Schindelbeck, A.J.; Ristow, H.M.; van Es, J.E.; Thies, H.A.; Shayler, M.B.; et al. Comprehensive Assessment of Soil Health; The Cornell Framework Manual; Crnell University: Geneva, Switzerland, 2016. [Google Scholar]
- De Souza Silva, A.M.; Fay, E.F. Effect of salinity on soil microorganisms. In Soil Health and Land Use Management; IntechOpen: London, UK, 2012; Volume 10, pp. 177–198. [Google Scholar]
- Tajik, S.; Ayoubi, S.; Lorenz, N. Soil Microbial Communities Affected by Vegetation, Topography and Soil Properties in a Forest Ecosystem. Appl. Soil Ecol. 2020, 149, 103514. [Google Scholar] [CrossRef]
- Rath, K.M.; Rousk, J. Salt Effects on the Soil Microbial Decomposer Community and Their Role in Organic Carbon Cycling: A Review. Soil Biol. Biochem. 2015, 81, 108–123. [Google Scholar] [CrossRef]
- Empadinhas, N.; da Costa, M.S. Osmoadaptation Mechanisms in Prokaryotes: Distribution of Compatible Solutes. Int. Microbiol. 2008, 11, 151–161. [Google Scholar]
- Chaudhary, D.R.; Rathore, A.P.; Jha, B. Effects of Seawater Irrigation on Soil Microbial Community Structure and Physiological Function. Int. J. Environ. Sci. Technol. 2016, 13, 2199–2208. [Google Scholar] [CrossRef]
- Pankhurst, C.E.; Yu, S.; Hawke, B.G.; Harch, B.D. Capacity of Fatty Acid Profiles and Substrate Utilization Patterns to Describe Differences in Soil Microbial Communities Associated with Increased Salinity or Alkalinity at Three Locations in South Australia. Biol. Fertil. Soils 2001, 33, 204–217. [Google Scholar] [CrossRef]
- Pan, C.; Liu, C.; Zhao, H.; Wang, Y. Changes of Soil Physico-Chemical Properties and Enzyme Activities in Relation to Grassland Salinization. Eur. J. Soil Biol. 2013, 55, 13–19. [Google Scholar] [CrossRef]
- Egamberdieva, D.; Davranov, K.; Wirth, S.; Hashem, A.; Abd_Allah, E.F. Impact of Soil Salinity on the Plant-Growth–Promoting and Biological Control Abilities of Root Associated Bacteria. Saudi J. Biol. Sci. 2017, 24, 1601–1608. [Google Scholar] [CrossRef]
- Zhang, W.; Chong, W.; Rui, X.; Wang, L. Effects of Salinity on the Soil Microbial Community and Soil Fertility. J. Integr. Agric. 2019, 18, 1360–1368. [Google Scholar] [CrossRef]
- Zhang, K.; Shi, Y.; Cui, X.; Yue, P.; Li, K.; Liu, X.; Tripathi, B.M.; Chu, H. Salinity Is a Key Determinant for Soil Microbial Communities in a Desert Ecosystem. mSystems 2019, 4, e00225-18. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, N.; Marschner, P.; Burns, R. Response of Microbial Activity and Community Structure to Decreasing Soil Osmotic and Matric Potential. Plant Soil 2011, 344, 241–254. [Google Scholar] [CrossRef]
- Baumann, K.; Marschner, P. Effects of Salinity on Microbial Tolerance to Drying and Rewetting. Biogeochemistry 2013, 112, 71–80. [Google Scholar] [CrossRef]
- Chambers, L.G.; Guevara, R.; Boyer, J.N.; Troxler, T.G.; Davis, S.E. Effects of Salinity and Inundation on Microbial Community Structure and Function in a Mangrove Peat Soil. Wetlands 2016, 36, 361–371. [Google Scholar] [CrossRef]
- Tunç, E.; Gul, O. Analysis of Phospholipid Fatty Acids (PLFA) as a Soil Bioindicator in Karkamis/Gaziantep Pistachio Orchards. Fresenius Environ. Bull. 2014, 23, 385. [Google Scholar]
- Zheng, Q.; Hu, Y.; Zhang, S.; Noll, L.; Böckle, T.; Dietrich, M.; Herbold, C.W.; Eichorst, S.A.; Woebken, D.; Richter, A.; et al. Soil Multifunctionality Is Affected by the Soil Environment and by Microbial Community Composition and Diversity. Soil Biol. Biochem. 2019, 136, 107521. [Google Scholar] [CrossRef]
- Kamble, P.N.; Gaikwad, V.B.; Kuchekar, S.R.; Baath, E. Microbial Growth, Biomass, Community Structure and Nutrient Limitation in High PH and Salinity Soils from Pravaranagar (India). Eur. J. Soil Biol. 2014, 65, 87–95. [Google Scholar] [CrossRef]
- Nunes, M.R.; Veum, K.S.; Parker, P.A.; Holan, S.H.; Karlen, D.L.; Amsili, J.P.; van Es, H.M.; Wills, S.A.; Seybold, C.A.; Moorman, T.B. The Soil Health Assessment Protocol and Evaluation Applied to Soil Organic Carbon. Soil Sci. Soc. Am. J. 2021, 85, 1196–1213. [Google Scholar] [CrossRef]
- Kumar, A.; Singh, S.; Mukherjee, A.; Rastogi, R.P.; Verma, J.P. Salt-Tolerant Plant Growth-Promoting Bacillus Pumilus Strain JPVS11 to Enhance Plant Growth Attributes of Rice and Improve Soil Health under Salinity Stress. Microbiol. Res. 2021, 242, 126616. [Google Scholar] [CrossRef]
- Mbodj, D.; Effa-Effa, B.; Kane, A.; Manneh, B.; Gantet, P.; Laplaze, L.; Diedhiou, A.; Grondin, A. Arbuscular Mycorrhizal Symbiosis in Rice: Establishment, Environmental Control and Impact on Plant Growth and Resistance to Abiotic Stresses. Rhizosphere 2018, 8, 12–26. [Google Scholar] [CrossRef]
- Kumar, A.; Singh, S.; Gaurav, A.K.; Srivastava, S.; Verma, J.P. Plant Growth-Promoting Bacteria: Biological Tools for the Mitigation of Salinity Stress in Plants. Front. Microbiol. 2020, 11, 1216. [Google Scholar] [CrossRef]
- Nautiyal, C.S.; Srivastava, S.; Chauhan, P.S. Rhizosphere Colonization: Molecular Determinants from Plant-Microbe Coexistence Perspective. In Molecular Mechanisms of Plant and Microbe Coexistence; Springer: Berlin/Heidelberg, Germany, 2008; pp. 99–123. [Google Scholar]
- Jaiswal, D.K.; Verma, J.P.; Prakash, S.; Meena, V.S.; Meena, R.S. Potassium as an Important Plant Nutrient in Sustainable Agriculture: A State of the Art. In Potassium Solubilizing Microorganisms for Sustainable Agriculture; Springer: New Delhi, India, 2016; pp. 21–29. [Google Scholar]
- Bhise, K.K.; Bhagwat, P.K.; Dandge, P.B. Synergistic Effect of Chryseobacterium Gleum Sp. SUK with ACC Deaminase Activity in Alleviation of Salt Stress and Plant Growth Promotion in Triticum aestivum L. 3 Biotech 2017, 7, 105. [Google Scholar] [CrossRef] [PubMed]
- Zahid, M.; Abbasi, M.K.; Hameed, S.; Rahim, N. Isolation and Identification of Indigenous Plant Growth Promoting Rhizobacteria from Himalayan Region of Kashmir and Their Effect on Improving Growth and Nutrient Contents of Maize (Zea mays L.). Front. Microbiol. 2015, 6, 207. [Google Scholar] [CrossRef]
- Kumawat, K.C.; Sharma, P.; Nagpal, S.; Gupta, R.K.; Sirari, A.; Nair, R.M.; Bindumadhava, H.; Singh, S. Dual Microbial Inoculation, a Game Changer?—Bacterial Biostimulants with Multifunctional Growth Promoting Traits to Mitigate Salinity Stress in Spring Mungbean. Front. Microbiol. 2021, 11, 3491. [Google Scholar] [CrossRef] [PubMed]
- Gr, S.; Yadav, R.K.; Chatrath, A.; Gerard, M.; Tripathi, K.; Govindsamy, V.; Abraham, G. Perspectives on the Potential Application of Cyanobacteria in the Alleviation of Drought and Salinity Stress in Crop Plants. J. Appl. Phycol. 2021, 33, 3761–3778. [Google Scholar] [CrossRef]
- Ben Ali, A.R.; Shukla, M.K.; Schutte, B.J.; Gard, C.C. Irrigation with RO Concentrate and Brackish Groundwater Impacts Pecan Tree Growth and Physiology. Agric. Water Manag. 2020, 240, 106328. [Google Scholar] [CrossRef]
- Ben Ali, A.R.; Yang, H.; Shukla, M. Brackish Groundwater and Reverse Osmosis Concentrate Influence Soil Physical and Thermal Properties and Pecan Evapotranspiration. Soil Sci. Soc. Am. J. 2021, 85, 1519–1533. [Google Scholar] [CrossRef]
- Gavlak, R.G.; Horneck, D.A.; Miller, R.O. Plant, Soil, and Water Reference Methods for the Western Region; Western Rural Development Center: Logan, UT, USA, 1994. [Google Scholar]
- Nelson, D.W.; Sommers, L.E. Total Carbon, Organic Carbon, and Organic Matter. In Methods of Soil Analysis; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 1996; pp. 961–1010. ISBN 978-0-89118-866-7. [Google Scholar]
- Buyer, J.S.; Sasser, M. High Throughput Phospholipid Fatty Acid. Appl. Soil Ecol. 2012, 61, 127–130. [Google Scholar] [CrossRef]
- Christie, W.W. Lipid Analysis: Isolation, Separation, Identification and Structural Analysis of Lipids; Oily Press: Bridgewater, UK, 2003. [Google Scholar]
- Rios-Velazquez, C.; Malave-Orengo, J.; Borglin, S.; Hazen, T. A Modified Cell Extraction Method to Access Microbial Community Structure in Soil Samples by Phospholipid Fatty Acid Analysis. Curr. Res. Technol. Educ. Top. Appl. Microbiol. Microb. Biotechnol. 2011, 2, 1562–1568. [Google Scholar]
- Provin, T.; Pitt, J.L. Managing Soil Salinity. In Texas Farmer Collection; Texas A&M University: College Station, TX, USA, 2001; pp. 1–7. [Google Scholar]
- Mohamed, D.J.; Martiny, J.B. Patterns of Fungal Diversity and Composition along a Salinity Gradient. ISME J. 2011, 5, 379–388. [Google Scholar] [CrossRef]
- Zhang, S.; Zheng, Q.; Noll, L.; Hu, Y.; Wanek, W. Environmental Effects on Soil Microbial Nitrogen Use Efficiency Are Controlled by Allocation of Organic Nitrogen to Microbial Growth and Regulate Gross N Mineralization. Soil Biol. Biochem. 2019, 135, 304–315. [Google Scholar] [CrossRef]
- Hoorman, J.J. The Role of Soil Bacteria. Fact sheet, Agriculture and Natural Resources; Ohio State University: Columbus, OH, USA, 2011; pp. 1–4. [Google Scholar]
- Estrada, B.; Aroca, R.; Maathuis, F.J.; Barea, J.M.; Ruiz-Lozano, J.M. Arbuscular Mycorrhizal Fungi Native from a Mediterranean Saline Area Enhance Maize Tolerance to Salinity through Improved Ion Homeostasis. Plant Cell Environ. 2013, 36, 1771–1782. [Google Scholar] [CrossRef]
- Min, W.; Guo, H.; Zhang, W.; Zhou, G.; Ma, L.; Ye, J.; Liang, Y.; Hou, Z. Response of Soil Microbial Community and Diversity to Increasing Water Salinity and Nitrogen Fertilization Rate in an Arid Soil. Acta Agric. Scand. Sect. B—Soil Plant Sci. 2016, 66, 117–126. [Google Scholar] [CrossRef]
- Wichern, J.; Wichern, F.; Joergensen, R.G. Impact of Salinity on Soil Microbial Communities and the Decomposition of Maize in Acidic Soils. Geoderma 2006, 137, 100–108. [Google Scholar] [CrossRef]
- Xie, L.-N.; Ge, Z.-M.; Li, Y.-L.; Li, S.-H.; Tan, L.-S.; Li, X.Z. Effects of Waterlogging and Increased Salinity on Microbial Communities and Extracellular Enzyme Activity in Native and Exotic Marsh Vegetation Soils. Soil Sci. Soc. Am. J. 2020, 84, 82–98. [Google Scholar] [CrossRef]
- Lynch, J.M.; Whipps, J.M. Substrate Flow in the Rhizosphere. Plant Soil 1990, 129, 1–10. [Google Scholar] [CrossRef]
- Kankarla, V.; Shukla, M.K.; VanLeeuwen, D.; Schutte, B.J.; Picchioni, G.A. Growth, Evapotranspiration, and Ion Uptake Characteristics of Alfalfa and Triticale Irrigated with Brackish Groundwater and Desalination Concentrate. Agronomy 2019, 9, 789. [Google Scholar] [CrossRef]
- Ahmadi, M.; Souri, M.K. Growth and Mineral Content of Coriander (Coriandrum sativum L.) Plants under Mild Salinity with Different Salts. Acta Physiol. Plant. 2018, 40, 1–8. [Google Scholar] [CrossRef]
- Doran, J.W.; Parkin, T.B. Defining and Assessing Soil Quality. In Defining Soil Quality for a Sustainable Environment; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 1994; pp. 1–21. ISBN 978-0-89118-930-5. [Google Scholar]
- Doran, J.W.; Zeiss, M.R. Soil Health and Sustainability: Managing the Biotic Component of Soil Quality. Appl. Soil Ecol. 2000, 15, 3–11. [Google Scholar] [CrossRef]
- Ikemura, Y.; Shukla, M.K. Soil Quality in Organic and Conventional Farms of New Mexico, USA. J. Org. Syst. 2009, 4, 34–47. [Google Scholar]
- Rekaby, S.A.; Awad, M.Y.; Hegab, S.A.; Eissa, M.A. Effect of Some Organic Amendments on Barley Plants under Saline Condition. J. Plant Nutr. 2020, 43, 1840–1851. [Google Scholar] [CrossRef]
- Mahmoud, E.; Ibrahim, M.; Ali, N.; Ali, H. Effect of Biochar and Compost Amendments on Soil Biochemical Properties and Dry Weight of Canola Plant Grown in Soil Contaminated with Heavy Metals. Commun. Soil Sci. Plant Anal. 2020, 51, 1561–1571. [Google Scholar] [CrossRef]
- Gunarathne, V.; Senadeera, A.; Gunarathne, U.; Biswas, J.K.; Almaroai, Y.A.; Vithanage, M. Potential of Biochar and Organic Amendments for Reclamation of Coastal Acidic-Salt Affected Soil. Biochar 2020, 2, 107–120. [Google Scholar] [CrossRef]
- Liu, D.; Ding, Z.; Ali, E.F.; Kheir, A.M.S.; Eissa, M.A.; Ibrahim, O.H.M. Biochar and Compost Enhance Soil Quality and Growth of Roselle (Hibiscus sabdariffa L.) under Saline Conditions. Sci. Rep. 2021, 11, 8739. [Google Scholar] [CrossRef] [PubMed]
- Porcel, R.; Aroca, R.; Ruiz-Lozano, J.M. Salinity Stress Alleviation Using Arbuscular Mycorrhizal Fungi. A Review. Agron. Sustain. Dev. 2012, 32, 181–200. [Google Scholar] [CrossRef]
- Paul, D.; Lade, H. Plant-Growth-Promoting Rhizobacteria to Improve Crop Growth in Saline Soils: A Review. Agron. Sustain. Dev. 2014, 34, 737–752. [Google Scholar] [CrossRef]
- Al-Nabulsi, Y.A. Saline Drainage Water, Irrigation Frequency and Crop Species Effects on Some Physical Properties of Soils. J. Agron. Crop Sci. 2001, 186, 15–20. [Google Scholar] [CrossRef]
- Aiello, R.; Cirelli, G.L.; Consoli, S. Effects of Reclaimed Wastewater Irrigation on Soil and Tomato Fruits: A Case Study in Sicily (Italy). Agric. Water Manag. 2007, 93, 65–72. [Google Scholar] [CrossRef]
- Flores, A.M.; Shukla, M.K.; Daniel, D.; Ulery, A.L.; Schutte, B.J.; Picchioni, G.A.; Fernald, S. Evapotranspiration Changes with Irrigation Using Saline Groundwater and RO Concentrate. J. Arid. Environ. 2016, 131, 35–45. [Google Scholar] [CrossRef]
- Adhikari, P.; Shukla, M.K.; Mexal, J.G. Spatial Variability of Hydraulic Conductivity and Sodium Content of Desert Soils: Implications for Management of Irrigation Using Treated Wastewater. Trans. ASABE 2012, 55, 1711–1721. [Google Scholar] [CrossRef]
- Duan, R.; Fedler, C.B.; Sheppard, C.D. Field Study of Salt Balance of a Land Application System. Water Air Soil Pollut. 2011, 215, 43–54. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).