
Microbial Community Composition Correlates with Metal Sorption in an Ombrotrophic Boreal Bog: Implications for Radionuclide Retention



Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling Site and Sample Pretreatment
2.2. Nucleic Acid Isolation
2.3. Amplicon Library Preparation
2.4. Sequence Processing and Analysis
2.5. Sorption Batch Experiments Data and Metal Concentrations
2.6. Statistical Analyses
3. Results
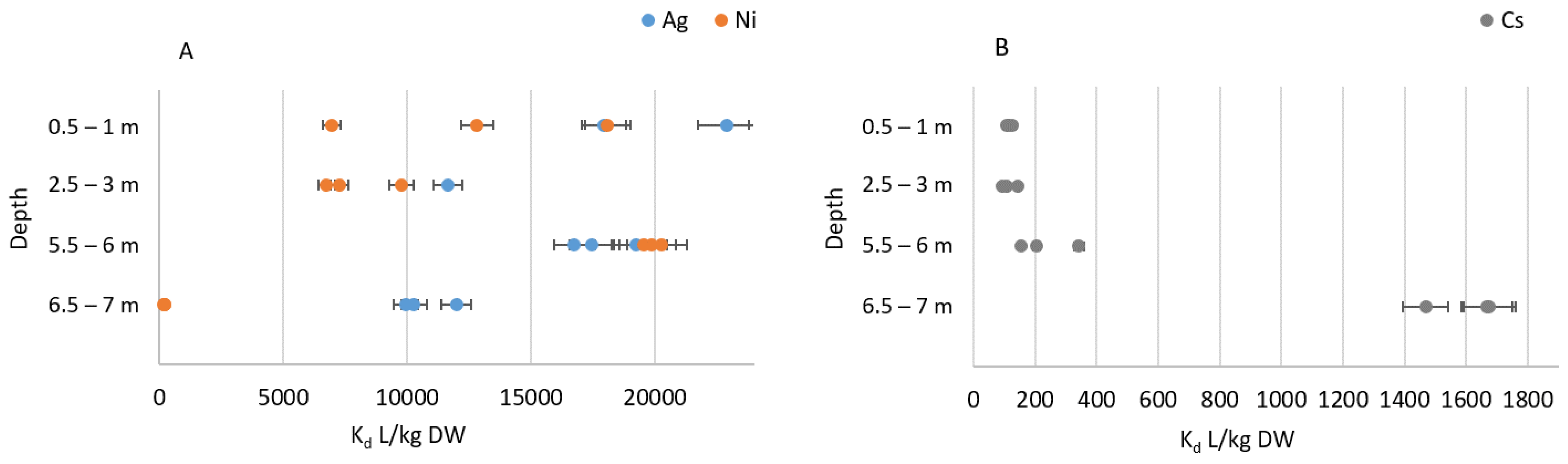
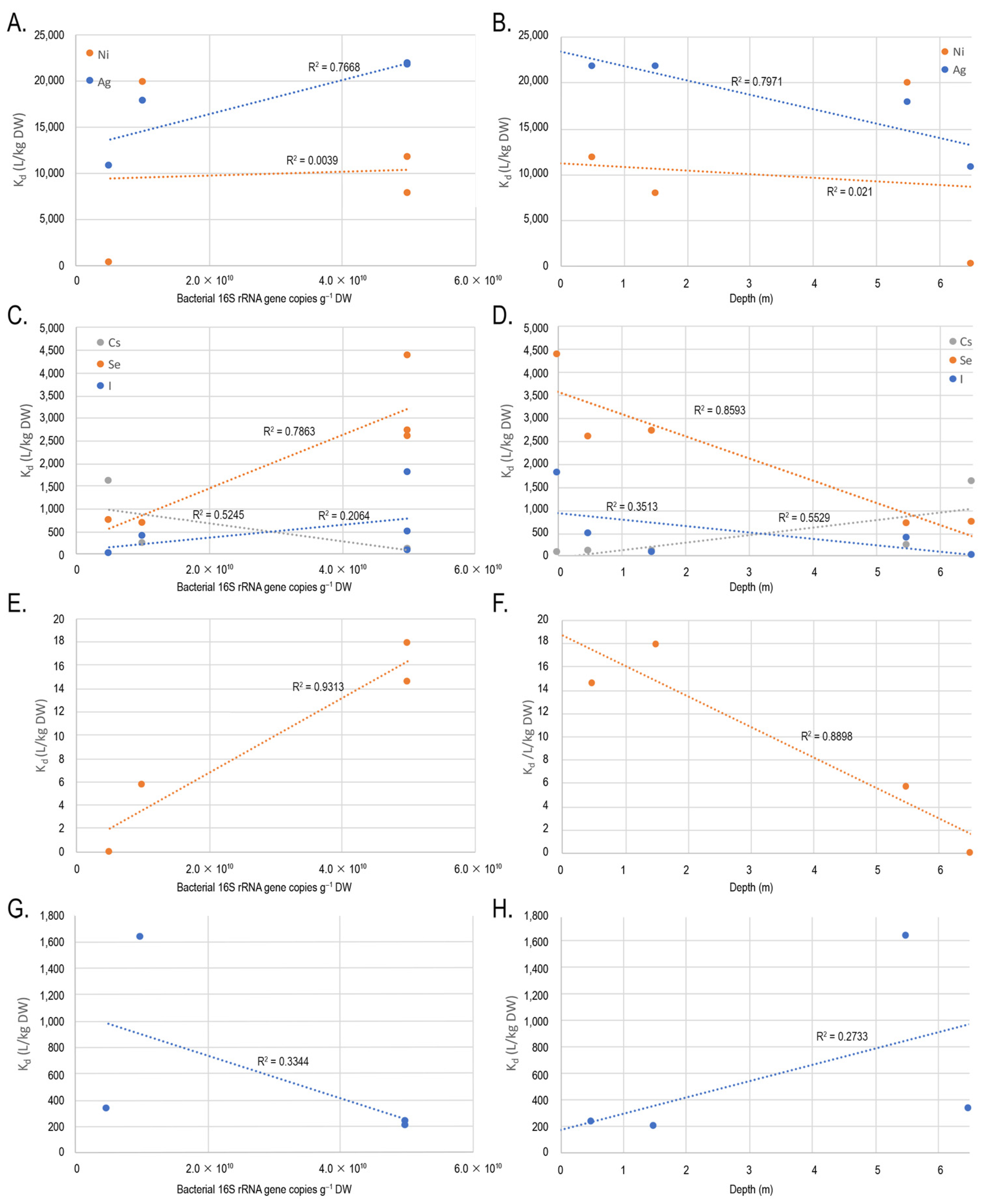
3.1. Sorption Experiments
3.2. Metal Concentrations
3.3. Archaeal, Fungal, and Bacterial Diversity
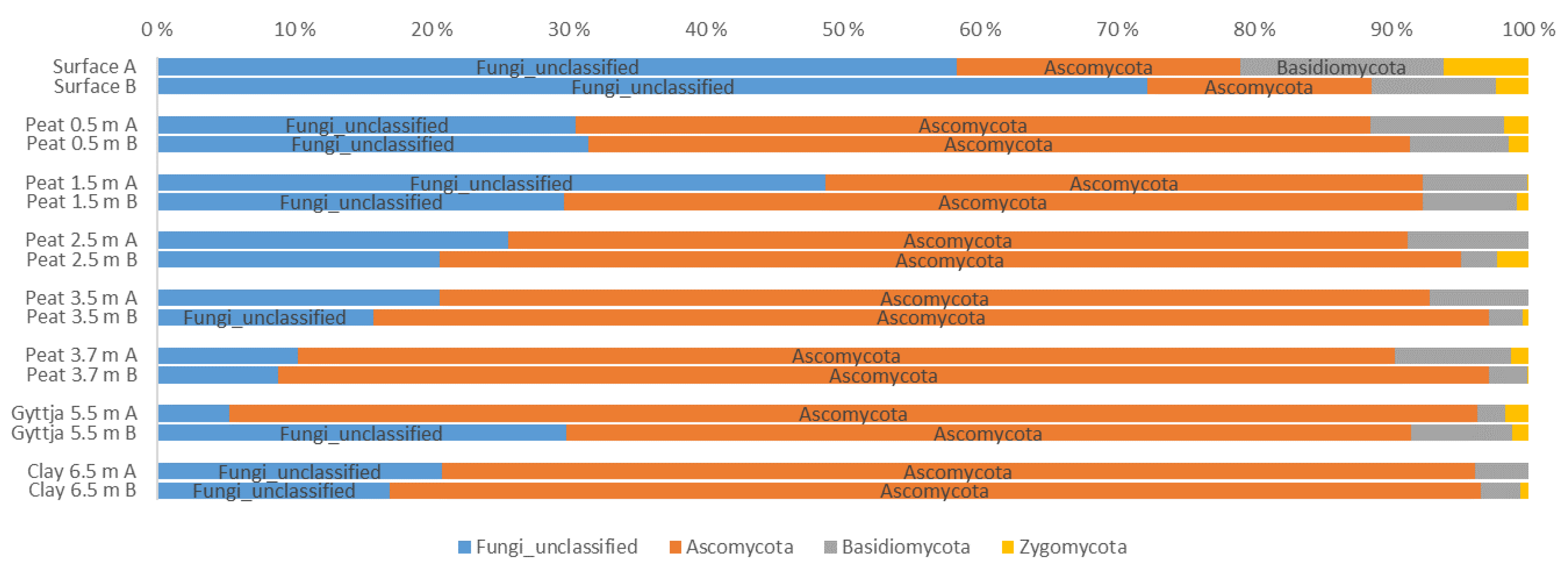
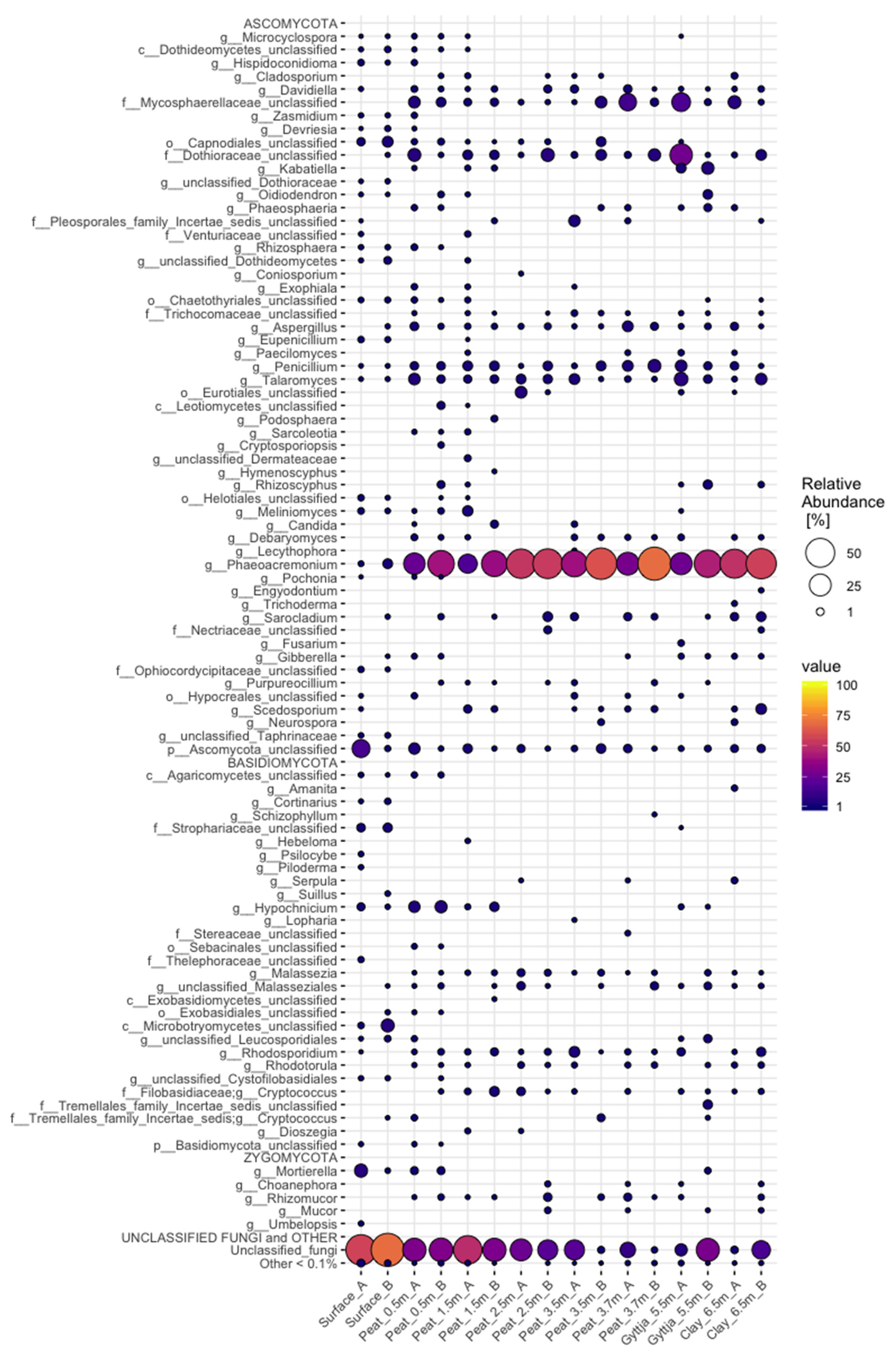
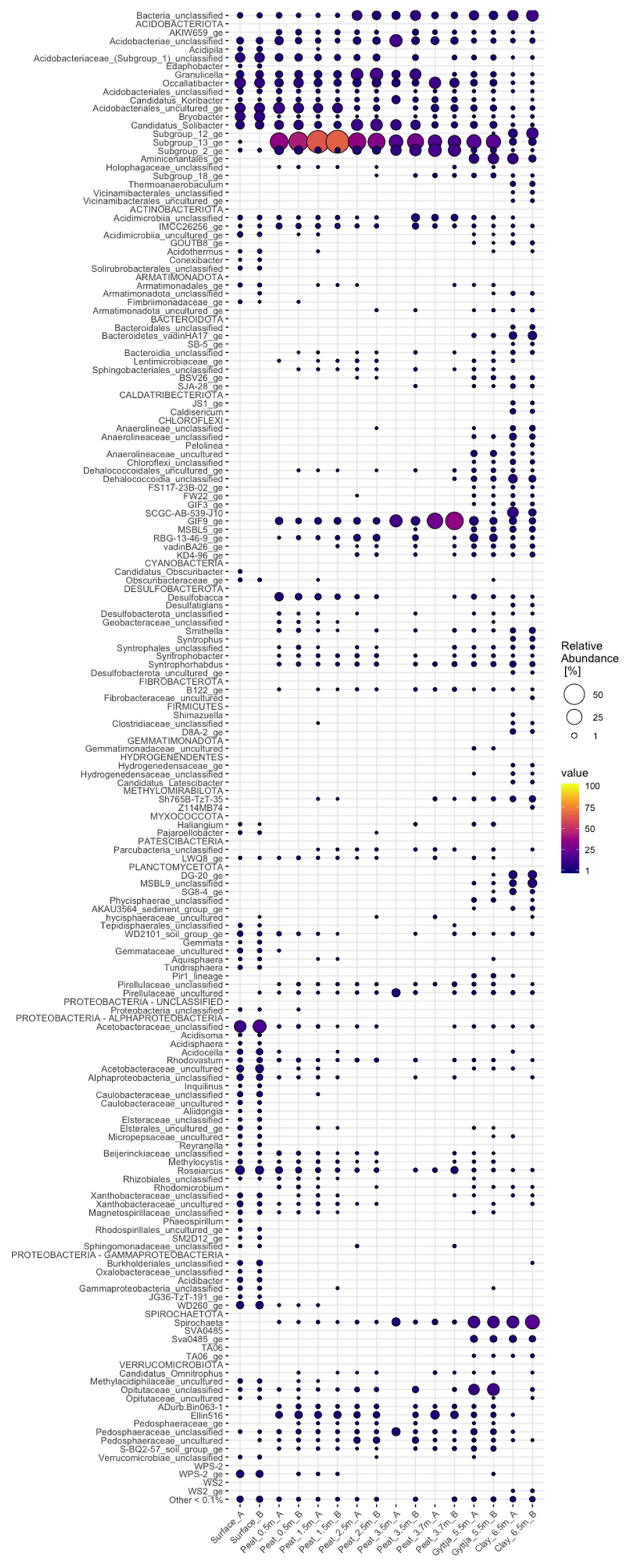
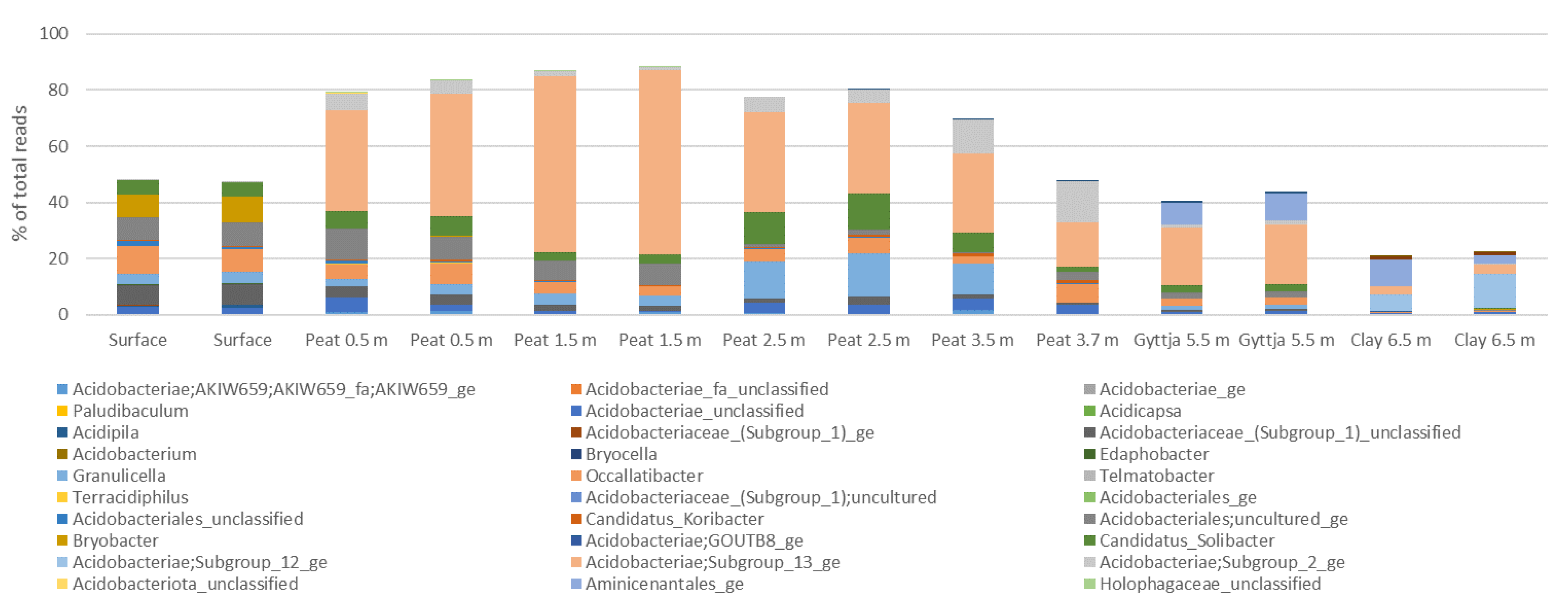
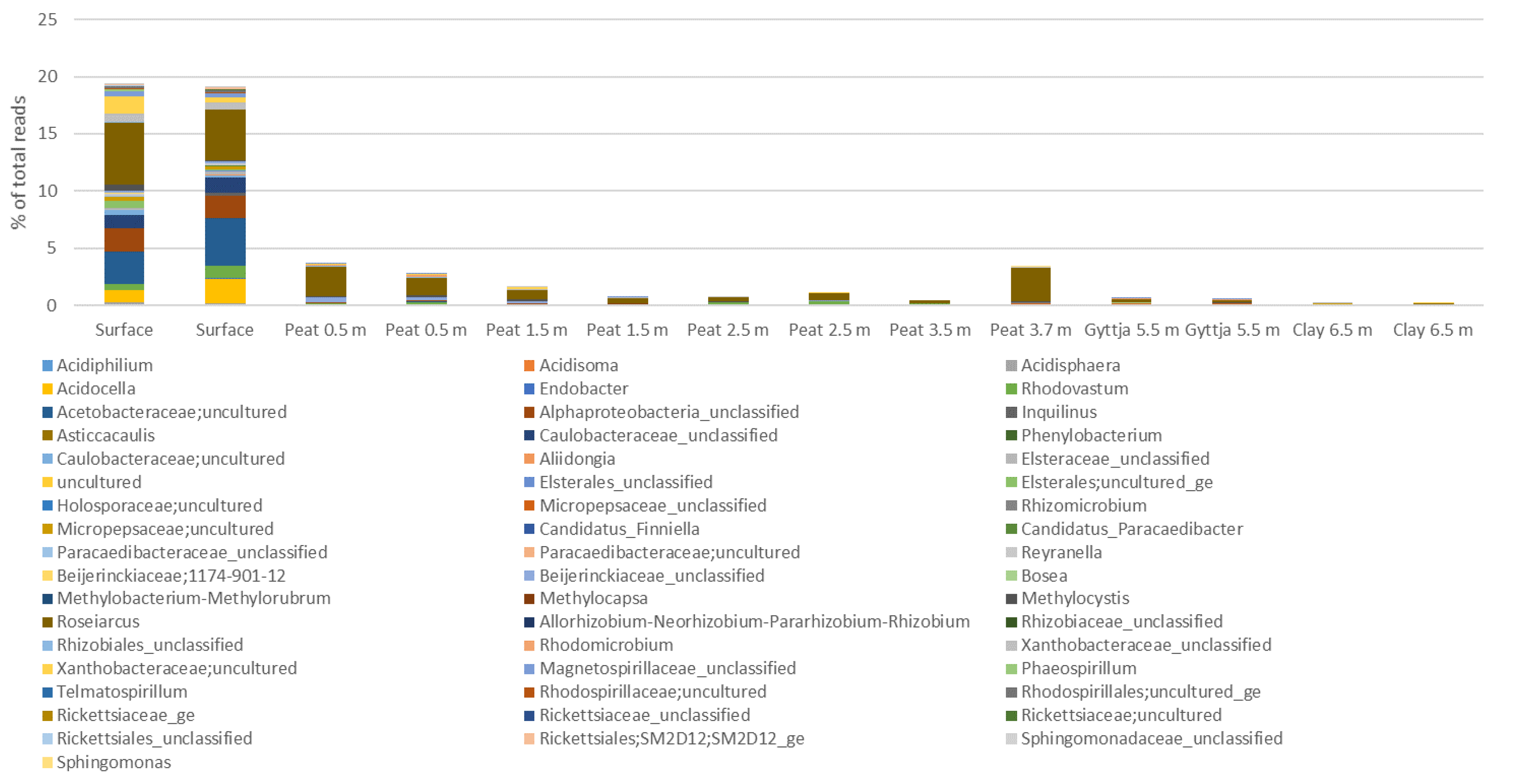
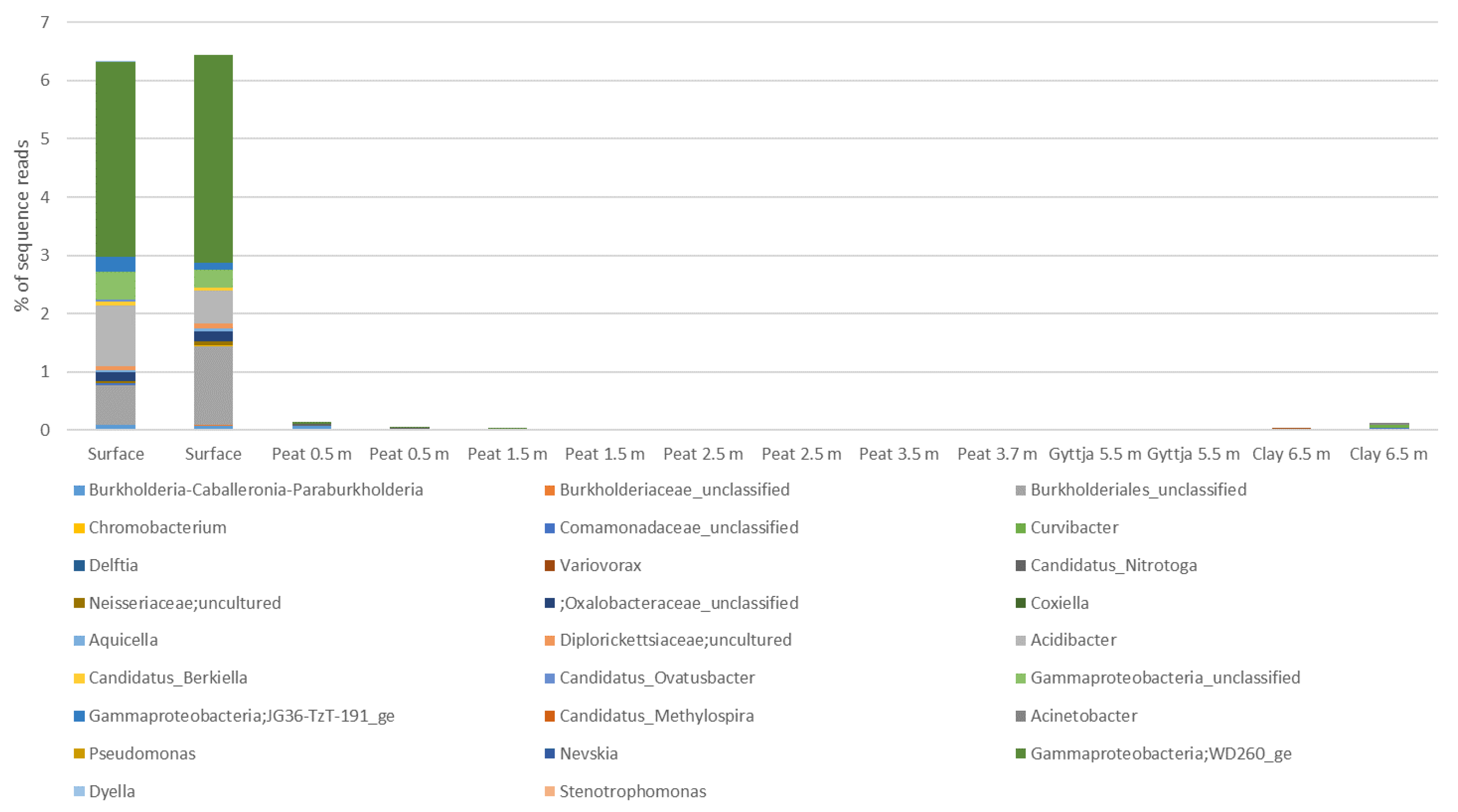
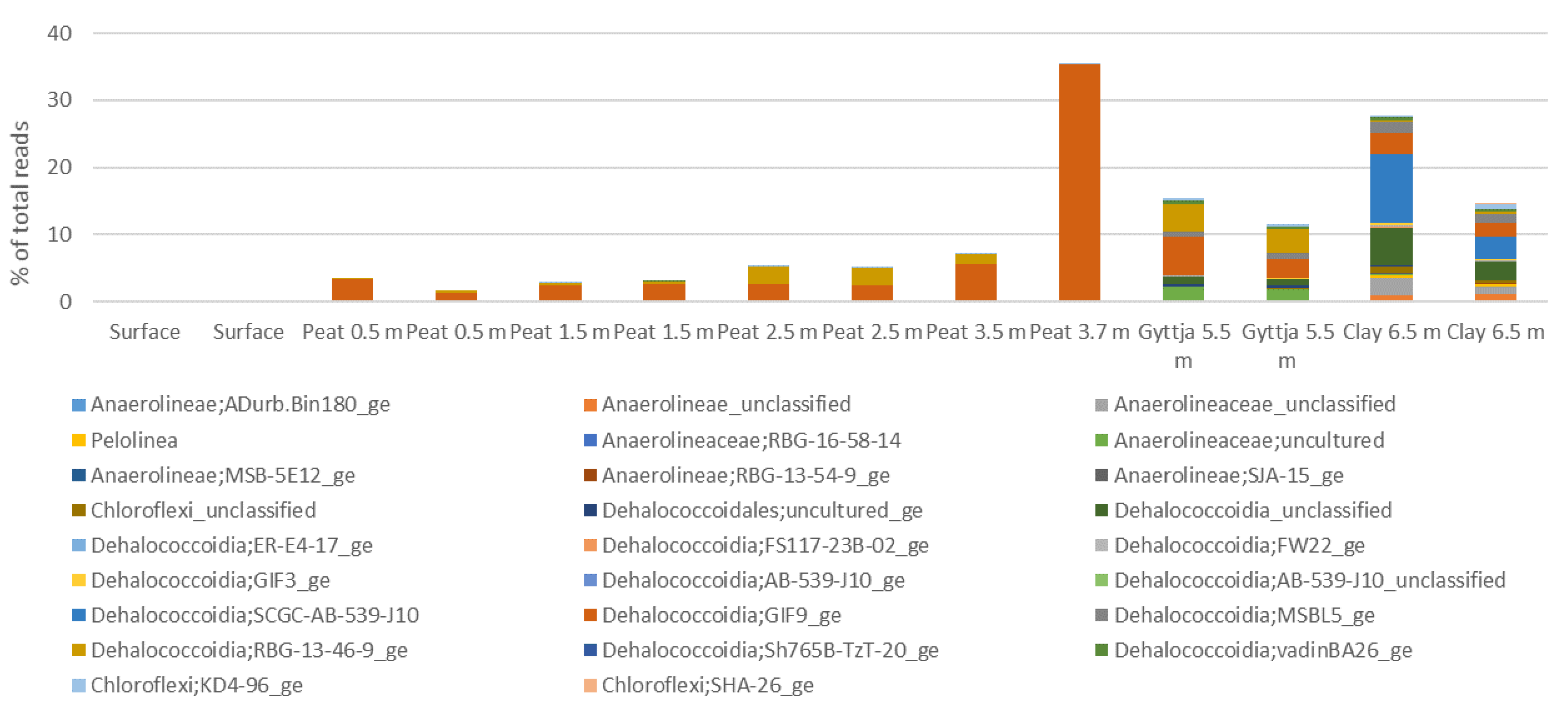
3.4. Archaeal, Fungal, and Bacterial Community Compositions
3.5. Correlation between Microbial Communities, Alphadiversity, and Physicochemical Parameters
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Candidatus Altiarchaeum | Lokiarchaeia Genus | Crenarchaeota Unclassified | HADARCHAEALES GENUS | Methanomicrobiaceae Uncultured | Methanosaeta | Methanomassiliicoccaceae Uncultured | |
|---|---|---|---|---|---|---|---|
| pH | 0.98 *** | 0.92 ** | 0.85 | 0.98 *** | 0.86 | ||
| Eh | −0.98 | ||||||
| Humification degree | 0.88 | ||||||
| Organic matter | −0.98 *** | −0.96 *** | −0.92 ** | −0.90 | |||
| Mg | 0.88 | 0.91 * | 0.89* | 0.85 | 0.93 ** | ||
| Fe | 0.88 | 0.94 ** | 0.98 *** | 0.96 *** | 0.92 * | 0.90 * | |
| Co | 0.88 | 0.96 ** | 0.99 *** | 0.96 *** | 0.93 ** | 0.90 * | |
| Cu | 0.90 * | 0.91 * | |||||
| Zn | 0.88 | 0.88 | |||||
| Se | 0.88 | 0.93 ** | 0.98 *** | 0.96 *** | 0.90 * | 0.89 * | |
| Cs | 0.88 | 0.93 ** | 0.98 *** | 0.96 *** | 0.90 * | 0.89 * | |
| Th | 0.87 | 0.97 *** | 0.99 *** | 0.95 *** | 0.95 *** | 0.90 * | |
| U | 0.86 | 0.98 *** | 0.98 *** | 0.93 ** | 0.97 *** | 0.90 * |
| pH | Organic Matter | Mg | Al | Fe | Co | Cu | Se | Cs | Th | U | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Acetothermiia | 0.95 | −1.0 *** | 0.92 | 1.0 *** | 0.99 *** | 1.0 *** | 1.0 *** | 0.99 *** | 0.97 *** | ||
| GOUTB8 | −1.0 *** | 0.92 | 1.0 *** | 1.0 *** | 0.99 *** | 0.99 *** | 1.0 *** | 0.98 *** | |||
| Acidobacteriae Subgroup 12 | 0.94 | 0.93 | 0.94 | 0.94 | 0.93 | ||||||
| Thermoanaerobaculum | −1.0 *** | 0.92 | 1.0 *** | 0.99 *** | 1.0 *** | 1.0 *** | 0.99 *** | 0.97 *** | |||
| Vicinamibacterales uncultured | 0.92 | 0.92 | 0.93 | 0.92 | 0.92 | ||||||
| Bacteroidales unclassified | 0.94 | 0.93 | 0.94 | 0.94 | 0.93 | ||||||
| Bacteroidetes vadinHA17 | −0.98 *** | 0.98 *** | 0.98 *** | 0.98 *** | 0.98 *** | 0.98 *** | 0.97 *** | ||||
| Prolixibacteraceae BSV13 | −0.98 *** | 0.98 *** | 0.98 *** | 0.98 *** | 0.98 *** | 0.97 *** | 0.95 *** | ||||
| BSV26 | 0.92 | ||||||||||
| Anaerolineae unclassified | −0.98 *** | 0.98 *** | 0.98 *** | 0.98 *** | 0.98 *** | 0.97 *** | 0.95 * | ||||
| Pelolinea | −1.0 *** | 0.93 | 1.0 *** | 1.0 *** | 1.0 *** | 1.0 *** | 0.99 *** | 0.97 *** | |||
| SJA-15 | −0.96 | 0.96 * | 0.95 * | 0.98 * | 0.96 * | 0.95 | 0.93 | ||||
| Chloroflexi unclassified | 0.91 | 0.92 | 0.91 | ||||||||
| Dehalococcoidia unclassified | 0.92 | 0.93 | 0.94 | 0.94 | |||||||
| GIF3 | 0.98 *** | 0.94 | 0.94 | 0.93 | 0.93 | 0.95 | 0.95 * | ||||
| MSBL5 | 0.94 | 0.92 | 0.96 * | ||||||||
| vadinBA26 | 0.94 | ||||||||||
| Desulfatiglans | 0.93 | 0.92 | 0.93 | 0.93 | 0.92 | ||||||
| Smithella | 0.93 | 0.92 | 0.93 | 0.93 | 0.92 | ||||||
| Syntrophus | −0.99 *** | 0.92 | 1.0 *** | 0.99 *** | 0.99 *** | 0.99 *** | 0.98 *** | 0.96 * | |||
| Firmicutes unclassified | −0.96 | 0.96 * | 0.95 * | 0.96 * | 0.96 * | 0.96 | 0.93 | ||||
| TSAC18 | −0.96 | 0.96 * | 0.95 * | 0.96 * | 0.96 * | 0.95 | 0.93 | ||||
| Hydrogenedensaceae | −0.99 *** | 0.91 | 0.99 *** | 0.98 *** | 0.99 *** | 0.99 *** | 0.98 *** | 0.96 * | |||
| Candidatus Latescibacter | −0.98 ** | 0.98 *** | 0.98 *** | 0.98 *** | 0.98 *** | 0.97 *** | 0.95 | ||||
| Latescibacteraceae unclassified | 0.94 | −1.0 *** | 0.92 | 1.0 *** | 0.99 *** | 1.0 *** | 1.0 *** | 0.99 *** | 0.97 *** | ||
| Methylomirabilaceae Sh765B-TzT-35 | −0.97 * | 0.92 | 0.97 *** | 0.97 *** | 0.96 *** | 0.96 * | 0.97 *** | 0.97 *** | |||
| MidBa8 | −1.0 *** | 0.92 | 1.0 *** | 0.99 *** | 1.0 *** | 1.0 *** | 0.99 *** | 0.97 *** | |||
| DG-20 | −1.0 *** | 0.92 | 1.0 *** | 0.99 *** | 1.0 *** | 1.0 *** | 0.99 *** | 0.97 *** | |||
| MSBL9 unclassified | 0.93 | 0.92 | 0.93 | 0.93 | 0.92 | ||||||
| AKAU3564 sediment group | −0.96 | 0.95 * | 0.95 * | 0.95 * | 0.95 * | 0.95 | 0.93 | ||||
| Planctomycetota unclassified | −0.94 | ||||||||||
| Spirochaeta | 0.93 | ||||||||||
| Sva0485 | 0.93 | ||||||||||
| WCHB1-41 | −0.96 | 0.96 * | 0.95 * | 0.96 * | 0.96 * | 0.95 | 0.93 |
References
- Hovmand, M.F.; Kemp, K.; Kystol, J.; Johnsen, I.; Riis-Nielsen, T.; Pacyna, J.M. Atmospheric heavy metal deposition accumulated in rural forest soils of southern Scandinavia. Environ. Pollut. 2008, 155, 537–541. [Google Scholar] [CrossRef] [PubMed]
- Ukonmaanaho, L.; Nieminen, T.M.; Rausch, N.; Shotyk, W. Heavy metal and arsenic profiles in ombrogenous peat cores from four differently loaded areas in Finland. Water Air Soil Pollut. 2004, 158, 277–294. [Google Scholar] [CrossRef]
- Laird, K.R.; Das, B.; Cumming, B.F. Enrichment of uranium, arsenic, molybdenum, and selenium in sediment cores from boreal lakes adjacent to northern Saskatchewan uranium mines. Lake Reserv. Manag. 2014, 30, 344–357. [Google Scholar] [CrossRef]
- Roberts, S.; Kirk, J.L.; Wiklund, J.A.; Muir, D.C.; Keating, J.; Yang, F.; Gleason, A.; Lawson, G.; Wang, X.; Evans, M. Sources of atmospheric metal (loid) pollution recorded in Thompson Manitoba lake sediment cores within the Canadian boreal biome. Sci. Total Environ. 2020, 732, 139043. [Google Scholar] [CrossRef] [PubMed]
- Tchounwou, P.B.; Yedjou, C.G.; Patlolla, A.K.; Sutton, D.J. Heavy metal toxicity and the environment. Mol. Clin. Environ. Toxicol. 2012, 101, 133–164. [Google Scholar]
- Sigee, D.C.; Al-Rabaee, R.H. Nickel toxicity inPseudomonas tabaci: Single cell and bulk sample analysis of bacteria cultured at high cation levels. Protoplasma 1986, 130, 171–185. [Google Scholar] [CrossRef]
- Rajapaksha, R.M.C.P.; Tobor-Kapłon, M.A.; Bååth, E. Metal toxicity affects fungal and bacterial activities in soil differently. Appl. Environ. Microbiol. 2004, 70, 2966–2973. [Google Scholar] [CrossRef]
- Pennanen, T.; Frostegard, A.S.A.; Fritze, H.; Baath, E. Phospholipid fatty acid composition and heavy metal tolerance of soil microbial communities along two heavy metal-polluted gradients in coniferous forests. Appl. Environ. Microbiol. 1996, 62, 420–428. [Google Scholar] [CrossRef]
- Haapanen, R.; Aro, L.; Lahdenperae, A.M.; Helin, J.; Ikonen, A.T.K. Studies on Reference Mires: 1. Lastensuo and Pesaensuo in 2010–2011; No. POSIVA-WR--12-102; Posiva Oy: Eurajoki, Finland, 2013. [Google Scholar]
- Hjerpe, T.; Broed, R.; Ikonen, A.T.K. Biosphere Assessment Report 2009; No. POSIVA--10-03; Posiva Oy: Eurajoki, Finland, 2010. [Google Scholar]
- Dönmez, G.Ç.; Aksu, Z.; Öztürk, A.; Kutsal, T. A comparative study on heavy metal biosorption characteristics of some algae. Process Biochem. 1999, 34, 885–892. [Google Scholar] [CrossRef]
- Wong, J.P.K.; Wong, Y.S.; Tam, N.F.Y. Nickel biosorption by two chlorella species, C. Vulgaris (a commercial species) and C. Miniata (a local isolate). Bioresour. Technol. 2000, 73, 133–137. [Google Scholar] [CrossRef]
- Aksu Ksu, Z. Determination of the equilibrium, kinetic and thermodynamic parameters of the batch biosorption of nickel (II) ions onto Chlorella vulgaris. Process Biochem. 2002, 38, 89–99. [Google Scholar] [CrossRef]
- Grandjean, P. Human Exposure to Nickel; IARC Scientific Publications: Lyon, France, 1984; pp. 469–485. [Google Scholar]
- Scott-Fordsmand, J.J. Toxicity of nickel to soil organisms in Denmark. Rev. Environ. Contam. Toxicol. 1997, 148, 1–34. [Google Scholar]
- Gupta, V.K.; Rastogi, A.; Nayak, A. Biosorption of nickel onto treated alga (Oedogonium hatei): Application of isotherm and kinetic models. J. Colloid Interface Sci. 2010, 342, 533–539. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, N.; Iqbal, J.; Iqbal, M. Removal and recovery of nickel (II) from aqueous solution by loofa sponge-immobilized biomass of Chlorella sorokiniana: Characterization studies. J. Hazard. Mater. 2004, 108, 85–94. [Google Scholar] [CrossRef] [PubMed]
- White, C.E.; Wilkinson, S.C.; Gadd, G.M. The role of microorganisms in biosorption of toxic metals and radionuclides. Intern. Biodeterior. Biodegrad. 1995, 35, 17–40. [Google Scholar] [CrossRef]
- Ledin, M.; Pedersen, K. The environmental impact of mine wastes—Roles of microorganisms and their significance in treatment of mine wastes. Earth-Sci. Rev. 1996, 41, 67–108. [Google Scholar] [CrossRef]
- López, A.; Lázaro, N.; Priego, J.M.; Marqués, A.M. Effect of pH on the biosorption of nickel and other heavy metals by Pseudomonas fluorescens 4F39. J. Ind. Microbiol. Biotechnol. 2000, 24, 146–151. [Google Scholar] [CrossRef]
- Sar, P.; Kazy, S.K.; Singh, S.P. Intracellular nickel accumulation by Pseudomonas aeruginosa and its chemical nature. Lett. App. Microbiol. 2001, 32, 257–261. [Google Scholar] [CrossRef]
- Mulrooney, S.B.; Hausinger, R.P. Nickel uptake and utilization by microorganisms. FEMS Microbiol. Rev. 2003, 27, 239–261. [Google Scholar] [CrossRef]
- Knuutinen, J.; Bomberg, M.; Kemell, M.; Lusa, M. Ni(II) interactions in boreal Peanibacillus sp., Methylobacterium sp., Paraburkholderia sp. and Pseudomonas sp. strains isolated from an acidic, ombrotrophic bog. Front. Microbiol. 2019. [Google Scholar] [CrossRef]
- Eitinger, T.; Mandrand-Berthelot, M.-A. Nickel transport systems in microorganisms. Arch. Microbiol. 2000, 173, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Pedroso, M.S.; Pinho, G.L.; Rodrigues, S.C.; Bianchini, A. Mechanism of acute silver toxicity in the euryhaline copepod Acartia tonsa. Aquat. Toxicol. 2007, 82, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, A.R.; McBride, M.B.; Baveye, P.; Steenhuis, T.S. Environmental factors determining the trace-level sorption of silver and thallium to soils. Sci. Total Environ. 2005, 345, 191–205. [Google Scholar] [CrossRef] [PubMed]
- Smith, I.; Carson, B. Silver; MI7 Ann Arbor Science Publishers: Ann Arbor, MI, USA, 1977. [Google Scholar]
- Hadrup, N.; Sharma, A.K.; Loeschner, K. Toxicity of silver ions, metallic silver, and silver nanoparticle materials after in vivo dermal and mucosal surface exposure: A review. Regul. Toxicol. Pharmacol. 2018, 98, 257–267. [Google Scholar] [CrossRef]
- Rosenman, K.D.; Seixas, N.; Jacobs, I. Potential nephrotoxic effects of exposure to silver. Occup. Environ. Med. 1987, 44, 267–272. [Google Scholar] [CrossRef]
- Das, D.; Das, N.; Mathew, L. Kinetics, equilibrium and thermodynamic studies on biosorption of Ag (I) from aqueous solution by macrofungus Pleurotus platypus. J. Hazard. Mater. 2010, 184, 765–774. [Google Scholar] [CrossRef]
- SKB. Long-Term Safety for the Final Repository for Spent Nuclear Fuel at Forsmark Main Report of the SR-Site Project Volume III; Errata 2011-10, SKB TR-11-01; Svensk Kärnbränslehantering AB: Stockholm, Sweden, 2011. [Google Scholar]
- Williams, M.; Wohlers, D.W.; Citra, M.; Diamond, G.L.; Swarts, S.G. Toxicological profile of Cesium; U.S. Department of Health and Human Services, Public Health Service Agency for Toxic Substances and Disease Registry: Atlanta, GA, USA, 2004.
- Sparks, D.L. Environmental Soil Chemistry; Elsevier: Amsterdam, The Netherlands, 2003. [Google Scholar]
- Helin, J.; Ikonen, A.T.K.; Hjerpe, T. Review of Element-Specific Data for Biosphere Assessment BSA-2009; Working Report 2010-37; Posiva Oy: Eurajoki, Finland, 2010. [Google Scholar]
- Bostick, B.C.; Vairavamurthy, M.A.; Karthikeyan, K.G.; Chorover, J. Cesium adsorption on clay minerals: An EXAFS spectroscopic investigation. Environ. Sci. Technol. 2002, 36, 2670–2676. [Google Scholar] [CrossRef]
- Chang, K.P.; Hsu, C.N.; Tamaki, H. Basic study of 137Cs sorption on soil. J. Nucl. Sci. Technol. 1993, 30, 1243–1247. [Google Scholar] [CrossRef]
- Zhuang, J.; Flury, M.; Jin, Y. Colloid-facilitated Cs transport through water-saturated Hanford sediment and Ottawa sand. Environ. Sci. Technol. 2003, 37, 4905–4911. [Google Scholar] [CrossRef]
- Lusa, M.; Bomberg, M.; Virtanen, S.; Lempinen, J.; Aromaa, H.; Knuutinen, J.; Lehto, J. Factors affecting the sorption of cesium in a nutrient-poor boreal bog. J. Environ. Radioact. 2015, 147, 22–32. [Google Scholar] [CrossRef]
- Sasaki, T.; Kubota, T.; Mito, S.; Kauri, T.; Kudo, A. Radionuclide sorption to a mixture of anaerobic bacteria in the repository environment. J. Nucl. Sci. Technol. 2002, 39 (Suppl. 3), 954–957. [Google Scholar] [CrossRef]
- Higo, M.; Kang, D.J.; Isobe, K. First report of community dynamics of arbuscular mycorrhizal fungi in radiocesium degradation lands after the Fukushima-Daiichi Nuclear disaster in Japan. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Ohnuki, T.; Sakamoto, F.; Kozai, N.; Nanba, K.; Neda, H.; Sasaki, Y.; Niizato, T.; Watanabe, N.; Kozaki, T. Role of filamentous fungi in migration of radioactive cesium in the Fukushima forest soil environment. Environ. Sci. Process. Impacts 2019, 21, 1164–1173. [Google Scholar] [CrossRef] [PubMed]
- Gyuricza, V.; Thiry, Y.; Wannijn, J.; Declerck, S.; Dupré de Boulois, H. Radiocesium transfer between Medicago truncatula plants via a common mycorrhizal network. Environ. Microbiol. 2010, 12, 2180–2189. [Google Scholar] [PubMed]
- Coppin, F.; Chabroullet, C.; Martin-Garin, A. Selenite interactions with some particulate organic and mineral fractions isolated from a natural grassland soil. Eur. J. Soil Sci. 2009, 60, 369–376. [Google Scholar] [CrossRef]
- Sharmasarkar, S.; Vance, G.F. Soil and plant selenium at a reclaimed uranium mine. J. Environ. Qual. 2002, 31, 1516–1521. [Google Scholar] [CrossRef]
- Manceau, A.; Gallup, D.L. Removal of Selenocyanate in Water by Precipitation: Characterization of Copper− Selenium Precipitate by X-ray Diffraction, Infrared, and X-ray Absorption Spectroscopy. Environ. Sci. Technol. 1997, 31, 968–976. [Google Scholar] [CrossRef]
- Yasin, M.; El Mehdawi, A.F.; Jahn, C.E.; Anwar, A.; Turner, M.F.; Faisal, M.; Pilon-Smits, E.A. Seleniferous soils as a source for production of selenium-enriched foods and potential of bacteria to enhance plant selenium uptake. Plant Soil 2015, 386, 385–394. [Google Scholar] [CrossRef]
- De Souza, M.P.; Chu, D.; Zhao, M.; Zayed, A.M.; Ruzin, S.E.; Schichnes, D.; Terry, N. Rhizosphere bacteria enhance selenium accumulation and volatilization by Indian mustard. Plant Physiol. 1999, 119, 565–574. [Google Scholar] [CrossRef]
- Barceloux, D.G. Selenium. J. Toxicol. Clin. Toxicol. 1999, 37, 145–172. [Google Scholar] [CrossRef]
- Xu, C.; Zhang, S.; Ho, Y.-F.; Miller, E.J.; Roberts, K.A.; Li, H.-P.; Schwehr, K.A.; Otosaka, S.; Kaplan, D.I.; Brinkmeyer, R.; et al. Is soil natural organic matter a sink or source for mobile radioiodine (129I) at the Savannah River Site? Geochim. Cosmochim. Acta 2011, 75, 5716–5735. [Google Scholar] [CrossRef]
- World Health Organization. Trace Elements in Human Nutrition and Health; World Health Organization: Geneva, Switzerland, 1996. [Google Scholar]
- Antonyak, H.; Iskra, R.; Panas, N.; Lysiuk, R. Selenium. In Trace Elements and Minerals in Health and Longevity; Springer International Publishing: Berlin/Heidelberg, Germany, 2018. [Google Scholar]
- Oldfield, J.E. Selenium World Atlas; Selenium-Tellurium Development Association (STDA): Grimbergen, Belgium, 2002. [Google Scholar]
- Fleming, G.A.; Walsh, T. Selenium occurrence in certain Irish soils and its toxic effects on animals. In Proceedings of the Royal Irish Academy. Section B: Biological, Geological, and Chemical Science; Hodges, Figgis, Co.: Dublin, Ireland, 1956; pp. 151–166. [Google Scholar]
- Li, H.F.; McGrath, S.P.; Zhao, F.J. Selenium uptake, translocation and speciation in wheat supplied with selenate or selenite. New Phytol. 2008, 178, 92–102. [Google Scholar] [CrossRef]
- Bebien, M.; Chauvin, J.P.; Adriano, J.M.; Grosse, S.; Verméglio, A. Effect of selenite on growth and protein synthesis in the phototrophic bacterium Rhodobacter sphaeroides. Appl. Environ. Microbiol. 2001, 67, 4440–4447. [Google Scholar] [CrossRef]
- Tarze, A.; Dauplais, M.; Grigoras, I.; Lazard, M.; Ha-Duong, N.-T.; Barbier, F.; Blanquet, S.; Plateau, P. Extracellular production of hydrogen selenide accounts for thiol-assisted toxicity of selenite against Saccharomyces cerevisiae. J. Biol. Chem. 2007, 282, 8759–8767. [Google Scholar] [CrossRef]
- Oremland, R.S.; Herbel, M.J.; Blum, J.S.; Langley, S.; Beveridge, T.J.; Ajayan, P.M.; Sutto, T.; Ellis, A.V.; Curran, S. Structural and spectral features of selenium nanospheres produced by Se-respiring bacteria. Appl. Environ. Microbiol. 2004, 70, 52–60. [Google Scholar] [CrossRef]
- Lusa, M.; Knuutinen, J.; Bomberg, M. Uptake and reduction of Se (IV) in two heterotrophic aerobic Pseudomonads strains isolated from boreal bog environment. AIMS Microbiol. 2017, 3, 798. [Google Scholar] [CrossRef]
- Oremland, R.S.; Steinberg, N.A.; Maest, A.S.; Miller, L.G.; Hollibaugh, J.T. Measurement of in situ rates of selenate removal by dissimilatory bacterial reduction in sediments. Environ. Sci. Technol. 1990, 24, 1157–1164. [Google Scholar] [CrossRef]
- Zhang, Y.; Moore, J.N. Selenium fractionation and speciation in a wetland system. Environ. Sci. Technol. 1996, 30, 2613–2619. [Google Scholar] [CrossRef]
- Stolz, J.F.; Oremland, R.S. Bacterial respiration of arsenic and selenium. FEMS Microbiol. Rev. 1999, 23, 615–627. [Google Scholar] [CrossRef]
- Nancharaiah, Y.V.; Lens, P.N.L. Ecology and biotechnology of selenium-respiring bacteria. Microbiol. Mol. Biol. Rev. 2015, 79, 61–80. [Google Scholar] [CrossRef] [PubMed]
- De Souza, M.P.; Huang, C.P.A.; Chee, N.; Terry, N. Rhizosphere bacteria enhance the accumulation of selenium and mercury in wetland plants. Planta 1999, 209, 259–263. [Google Scholar] [CrossRef]
- Lusa, M.; Help, H.; Honkanen, A.P.; Knuutinen, J.; Parkkonen, J.; Kalasová, D.; Bomberg, M. The reduction of selenium (IV) by boreal Pseudomonas sp. strain T5-6-I—Effects on selenium (IV) uptake in Brassica oleracea. Environ. Res. 2019, 177, 108642. [Google Scholar] [CrossRef]
- Rosenfeld, C.E.; Kenyon, J.A.; James, B.R.; Santelli, C.M. Selenium (IV, VI) reduction and tolerance by fungi in an oxic environment. Geobiology 2017, 15, 441–452. [Google Scholar] [CrossRef] [PubMed]
- Charlet, L.; Scheinost, A.; Tournassat, C.; Greneche, J.-M.; Gehin, A.; Fernández-Martınez, A.; Coudert, S.; Tisserand, D.; Brendle, J. Electron transfer at the mineral/water interface: Selenium reduction by ferrous iron sorbed on clay. Geochim. Cosmochim. Acta 2007, 71, 5731–5749. [Google Scholar] [CrossRef]
- Assemi, S.; Erten, H. Sorption of radioiodine on organic rich soil, clay minerals and alumina. J. Radioanal. Nucl. Chem. 1994, 178, 193–204. [Google Scholar] [CrossRef]
- Evans, G.J.; Hammad, K.A. Radioanalytical studies of iodine behaviour in the environment. J. Radioanal. Nucl. Chem. 1995, 192, 239–247. [Google Scholar] [CrossRef]
- Sheppard, M.I.; Hawkins, J.L. Iodine and microbial interactions in an organic soil. J. Environ. Radioact. 1995, 29, 91–109. [Google Scholar] [CrossRef]
- Ashworth, D.J.; Shaw, G.; Butler, A.P.; Ciciani, L. Soil transport and plant uptake of radio-iodine from near-surface groundwater. J. Environ. Radioact. 2003, 70, 99–114. [Google Scholar] [CrossRef]
- Ashworth, D.J.; Shaw, G. A comparison of the soil migration and plant uptake of radioactive chlorine and iodine from contaminated groundwater. J. Environ. Radioact. 2006, 89, 61–80. [Google Scholar] [CrossRef]
- Li, H.-P.; Yeager, C.M.; Brinkmeyer, R.; Zhang, S.; Ho, Y.-F.; Xu, C.; Jones, W.L.; Schwehr, K.A.; Otosaka, S.; Roberts, K.A.; et al. Bacterial production of organic acids enhances H2O2-dependent iodide oxidation. Environ. Sci. Technol. 2012, 46, 4837–4844. [Google Scholar] [CrossRef]
- Bostock, A.C.; Shaw, G.; Bell, J.N.B. The volatilisation and sorption of 129I in coniferous forest, grassland and frozen soils. J. Environ. Radioact. 2003, 70, 29–42. [Google Scholar] [CrossRef]
- Yamaguchi, N.; Nakano, M.; Takamatsu, R.; Tanida, H. Inorganic iodine incorporation into soil organic matter: Evidence from iodine K-edge X-ray absorption near-edge structure. J. Environ. Radioact. 2010, 101, 451–457. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Miller, E.J.; Zhang, S.; Li, H.-P.; Ho, Y.-F.; A Schwehr, K.; Kaplan, D.I.; Otosaka, S.; A Roberts, K.; Brinkmeyer, R.; et al. Sequestration and remobilization of radioiodine (129I) by soil organic matter and possible consequences of the remedial action at Savannah River Site. Environ. Sci. Technol. 2011, 45, 9975–9983. [Google Scholar] [CrossRef] [PubMed]
- Li, H.P.; Yeager, C.M.; Brinkmeyer, R.; Jones, W.L.; Zhang, S.J.; Xu, C.; Schwehr, K.A.; Santschi, P.H.; Kaplan, D.I.; Yeager, C.M. Iodide accumulation by aerobic bacteria isolated from subsurface sediments of an I-129 contaminated aquifer at the Savannah River Site, South Carolina. Appl. Environ. Microbiol. 2011, 77, 2153–2160. [Google Scholar] [CrossRef]
- Xu, C.; Chen, H.; Sugiyama, Y.; Zhang, S.; Li, H.P.; Ho, Y.F.; Chuang, C.Y.; Schwehr, K.A.; Kaplan, D.I.; Yeager, C.; et al. Novel molecular-level evidence of iodine binding to natural organic matter from Fourier transform ion cyclotron resonance mass spectrometry. Sci. Total Environ. 2013, 449, 244–252. [Google Scholar] [CrossRef]
- Amachi, S.; Mishima, Y.; Shinoyama, H.; Muramatsu, Y.; Fujii, T. Active transport and accumulation of iodide by newly isolated marine bacteria. Appl. Environ. Microbiol. 2005, 71, 741–745. [Google Scholar] [CrossRef] [PubMed]
- Ban-Nai, T.; Muramatsu, Y.; Amachi, S. Rate of iodine volatilization and accumulation by filamentous fungi through laboratory cultures. Chemosphere 2006, 65, 2216–2222. [Google Scholar] [CrossRef]
- Lusa, M.; Bomberg, M.; Aromaa, H.; Knuutinen, J.; Lehto, J. Sorption of radioiodide in an acidic, nutrient-poor boreal bog: Insights into the microbial impact. J. Environ. Radioact. 2015, 143, 110–122. [Google Scholar] [CrossRef]
- Dupont, C.L.; Grass, G.; Rensing, C. Copper toxicity and the origin of bacterial resistance—New insights and applications. Metallomics 2011, 3, 1109–1118. [Google Scholar] [CrossRef]
- Vincent, M.; Duval, R.E.; Hartemann, P.; Engels-Deutsch, M. Contact killing and antimicrobial properties of copper. J. Appl. Microbiol. 2018, 124, 1032–1046. [Google Scholar] [CrossRef]
- Antsotegi-Uskola, M.; Markina-Iñarrairaegui, A.; Ugalde, U. New insights into copper homeostasis in filamentous fungi. Int. Microbiol. 2020, 23, 65–73. [Google Scholar] [CrossRef]
- Richard, D.; Ravigné, V.; Rieux, A.; Facon, B.; Boyer, C.; Boyer, K.; Grygiel, P.; Javegny, S.; Terville, M.; Canteros, B.I.; et al. Adaptation of genetically monomorphic bacteria: Evolution of copper resistance through multiple horizontal gene transfers of complex and versatile mobile genetic elements. Mol. Ecol. 2017, 26, 2131–2149. [Google Scholar] [CrossRef]
- Nies, D.H. Bacterial transition metal homeostasis. In Molecular Microbiology of Heavy Metals; Springer: Berlin/Heidelberg, Germany, 2007; pp. 117–142. [Google Scholar]
- Rosen, B.P.; Liu, Z. Transport pathways for arsenic and selenium: A minireview. Environ. Int. 2009, 35, 512–515. [Google Scholar] [CrossRef]
- Turner, R.J.; Weiner, J.H.; Taylor, D.E. Selenium metabolism in Escherichia coli. Biometals 1998, 11, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Amachi, S. Microbial contribution to global iodine cycling: Volatilization, accumulation, reduction, oxidation, and sorption of iodine. Microbes Environ. 2008, 23, 269–276. [Google Scholar] [CrossRef]
- Gozlan, R.S. Isolation of iodine-producing bacteria from aquaria. Antonie Leeuwenhoek 1968, 34, 226. [Google Scholar] [CrossRef] [PubMed]
- Amachi, S.; Muramatsu, Y.; Akiyama, Y.; Miyazaki, K.; Yoshiki, S.; Hanada, S.; Kamagata, Y.; Ban-Nai, T.; Shinoyama, H.; Fujii, T. Isolation of iodide-oxidizing bacteria from iodide-rich natural gas brines and seawaters. Microb. Ecol. 2005, 49, 547–557. [Google Scholar] [CrossRef] [PubMed]
- Li, H.-P.; Daniel, B.; Creeley, D.; Grandbois, R.; Zhang, S.; Xu, C.; Ho, Y.-F.; Schwehr, K.A.; Kaplan, D.I.; Santschi, P.H.; et al. Superoxide production by a manganese-oxidizing bacterium facilitates iodide oxidation. Appl. Environ. Microbiol. 2014, 80, 2693–2699. [Google Scholar] [CrossRef] [PubMed]
- Amachi, S.; Kimura, K.; Muramatsu, Y.; Shinoyama, H.; Fujii, T. Hydrogen peroxide-dependent uptake of iodine by marine Flavobacteriaceae bacterium strain C-21. Appl. Environ. Microbiol. 2007, 73, 7536–7541. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Amachi, S.; Minami, K.; Miyasaka, I.; Fukunaga, S. Ability of anaerobic microorganisms to associate with iodine: 125I tracer experiments using laboratory strains and enriched microbial communities from subsurface formation water. Chemosphere 2010, 79, 349–354. [Google Scholar] [CrossRef]
- Lusa, M.; Lehto, J.; Aromaa, H.; Knuutinen, J.; Bomberg, M. Uptake of radioiodide by Paenibacillus sp., Pseudomonas sp., Burkholderia sp. and Rhodococcus sp. isolated from a boreal nutrient-poor bog. J. Environ. Sci. 2016, 44, 26–37. [Google Scholar] [CrossRef]
- Duborská, E.; Urík, M.; Bujdoš, M. Comparison of iodide and iodate accumulation and volatilization by filamentous fungi during static cultivation. Water Air Soil Pollut. 2017, 228, 1–8. [Google Scholar] [CrossRef]
- Fang, L.; Cai, P.; Chen, W.; Liang, W.; Hong, Z.; Huang, Q. Impact of cell wall structure on the behavior of bacterial cells in the binding of copper and cadmium. Colloids Surf. A Physicochem. Eng. Asp. 2009, 347, 50–55. [Google Scholar] [CrossRef]
- Bhainsa, K.C.; D’Souza, S.F. Thorium biosorption by Aspergillus fumigatus, a filamentous fungal biomass. J. Hazard. Mater. 2009, 165, 670–676. [Google Scholar] [CrossRef]
- Fein, J.B.; Daughney, C.J.; Yee, N.; Davis, T.A. A chemical equilibrium model for metal adsorption onto bacterial surfaces. Geochim. Cosmochim. Acta 1997, 61, 3319–3328. [Google Scholar] [CrossRef]
- Gadd, G.M.; White, C. Microbial treatment of metal pollution—A working biotechnology? Trends Biotechnol. 1993, 11, 353–359. [Google Scholar] [CrossRef]
- Mishra, B.; Boyanov, M.; Bunker, B.A.; Kelly, S.D.; Kemner, K.M.; Fein, J.B. High-and low-affinity binding sites for Cd on the bacterial cell walls of Bacillus subtilis and Shewanella oneidensis. Geochim. Cosmochim. Acta 2010, 74, 4219–4233. [Google Scholar] [CrossRef]
- Antsotegi-Uskola, M.; Markina-Iñarrairaegui, A.; Ugalde, U. Copper resistance in Aspergillus nidulans relies on the PI-type ATPase CrpA, regulated by the transcription factor AceA. Front. Microbiol. 2017, 8, 912. [Google Scholar] [CrossRef] [PubMed]
- Orell, A.; Navarro, C.A.; Arancibia, R.; Mobarec, J.C.; Jerez, C.A. Life in blue: Copper resistance mechanisms of bacteria and archaea used in industrial biomining of minerals. Biotechnol. Adv. 2010, 28, 839–848. [Google Scholar] [CrossRef]
- Nies, D.H. Efflux-mediated heavy metal resistance in prokaryotes. FEMS Microbiol. Rev. 2003, 27, 313–339. [Google Scholar] [CrossRef]
- Aro, L.; Korpela, L.; Mäkinen, V.; Salemaa, M.; Saarinen, M.; Lahdenperä, A.M.; Parviainen, L.; Kuusisto, J. Studies on Reference Mires: 2. Lastensuo, Pesänsuo and Häädetkeidas in 2010–2015; Posiva WR 2017-15; Posiva Oy: Eurajoki, Finland, 2018. [Google Scholar]
- Mäkilä, M.; Grundström, A. Turpeen ikä ja Kerrostumisnopeus Lounais-Suomen Soilla; Working Report 2008–12; Posiva Oy: Eurajoki, Finland, 2008. (In Finnish) [Google Scholar]
- Tsitko, I.; Lusa, M.; Lehto, J.; Parviainen, L.; Ikonen, A.T.; Lahdenperä, A.M.; Bomberg, M. The variation of microbial communities in a depth profile of an acidic, nutrient-poor boreal bog in southwestern Finland. Open J. Ecol. 2014, 4, 832. [Google Scholar] [CrossRef]
- Lusa, M.; Knuutinen, J.; Lindgren, M.; Virkanen, J.; Bomberg, M. Microbial communities in a former pilot-scale uranium mine in Eastern Finland–Association with radium immobilization. Sci. Total Environ. 2019, 686, 619–640. [Google Scholar] [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes-application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.J.W.T.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. Guide Methods Appl. 1990, 18, 315–322. [Google Scholar]
- Edwards, U.; Rogall, T.; Blöcker, H.; Emde, M.; Böttger, E.C. Isolation and direct complete nucleotide determination of entire genes. Characterization of a gene coding for 16S ribosomal RNA. Nucleic Acids Res. 1989, 17, 7843–7853. [Google Scholar] [CrossRef] [PubMed]
- Muyzer, G.; De Waal, E.C.; Uitterlinden, A.G. Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Appl. Environ. Microbiol. 1993, 59, 695–700. [Google Scholar] [CrossRef] [PubMed]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef]
- Pruesse, E.; Quast, C.; Knittel, K.; Fuchs, B.M.; Ludwig, W.; Peplies, J.; Glöckner, F.O. SILVA: A comprehensive online resource for quality checked and aligned ribosomal RNA sequence data compatible with ARB. Nucleic Acids Res. 2007, 35, 7188–7196. [Google Scholar] [CrossRef] [PubMed]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef]
- Kõljalg, U.; Nilsson, R.H.; Abarenkov, K.; Tedersoo, L.; Taylor, A.F.S.; Bahram, M.; Bates, S.T.; Bruns, T.D.; Bengtsson-Palme, J.; Callaghan, T.M.; et al. Towards a unified paradigm for sequence-based identification of fungi. Mol. Ecol. 2013, 22, 5271–5277. [Google Scholar] [CrossRef]
- Nilsson, R.H.; Larsson, K.-H.; Taylor, A.F.S.; Bengtsson-Palme, J.; Jeppesen, T.S.; Schigel, D.; Kennedy, P.; Picard, K.; Glöckner, F.O.; Tedersoo, L.; et al. The UNITE database for molecular identification of fungi: Handling dark taxa and parallel taxonomic classifications. Nucleic Acids Res. 2019, 47, D259–D264. [Google Scholar] [CrossRef]
- Abarenkov, K.; Zirk, A.; Piirmann, T.; Pöhönen, R.; Ivanov, F.; Nilsson, R.H.; Kõljalg, U. UNITE Mothur Release for Fungi. Version 04.02.2020. UNITE Community. 2020. Available online: https://plutof.ut.ee/#/doi/10.15156/BIO/786381 (accessed on 20 April 2020).
- Lusa, M.; Bomberg, M.; Aromaa, H.; Knuutinen, J.; Lehto, J. The microbial impact on the sorption behaviour of selenite in an acidic, nutrient-poor boreal bog. J. Environ. Radioact. 2015, 147, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Hammer, Ø.; Harper, D.A.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- McMurdie, P.J.; Holmes, S. phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014; Available online: http://www.R-project.org/ (accessed on 9 September 2020).
- Scheller, S.; Goenrich, M.; Boecher, R.; Thauer, R.K.; Jaun, B. The key nickel enzyme of methanogenesis catalyses the anaerobic oxidation of methane. Nature 2010, 465, 606–608. [Google Scholar] [CrossRef] [PubMed]
- Madhaiyan, M.; Poonguzhali, S.; Sa, T. Metal tolerating methylotrophic bacteria reduces nickel and cadmium toxicity and promotes plant growth of tomato (Lycopersicon esculentum L.). Chemosphere 2007, 69, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Ho, Y.S.; Wase, D.J.; Forster, C.F. Batch nickel removal from aqueous solution by sphagnum moss peat. Water Res. 1995, 29, 1327–1332. [Google Scholar] [CrossRef]
- Vila Domínguez, A.; Ayerbe Algaba, R.; Miró Canturri, A.; Rodríguez Villodres, Á.; Smani, Y. Antibacterial activity of colloidal silver against gram-negative and gram-positive bacteria. Antibiotics 2020, 9, 36. [Google Scholar] [CrossRef]
- Lusa, M.; Lehto, J.; Bomberg, M. The uptake of Ni2+ and Ag+ by bacterial strains isolated from a boreal nutrient-poor bog. AIMS Microbiol. 2016, 2, 120–137. [Google Scholar] [CrossRef]
- Andreasson, A.; Jönsson, B.; Lindman, B. Surface and colloid chemistry of peat and peat dewatering. Electrostatic effects. Colloid Polym. Sci. 1988, 266, 164–172. [Google Scholar] [CrossRef]
- Strelko, V., Jr.; Malik, D.J. Characterization and metal sorptive properties of oxidized active carbon. J. Colloid Interface Sci. 2002, 250, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Corapcioglu, M.O.; Huang, C.P. The adsorption of heavy metals onto hydrous activated carbon. Water Res. 1987, 21, 1031–1044. [Google Scholar] [CrossRef]
- Tan, K.H. Humic Matter in Soil and the Environment: Principles and Controversies, 2nd ed.; CRC Press, Taylor and Francis Group: Boca Raton, FL, USA, 2014; p. 324. [Google Scholar]
- Martinez, E.R.; Sharma, P.; Kappler, A. Surface binding site analysis of Ca2+-homoionized clay–humic acid complexes. J. Colloid Interface Sci. 2010, 352, 526–534. [Google Scholar] [CrossRef] [PubMed]
- Koch-Steindl, H.; Pröhl, G. Considerations on the behaviour of long-lived radionuclides in the soil. Radiat. Environ. Biophys. 2001, 40, 93–104. [Google Scholar] [CrossRef]
- Huber, R.; Sacher, M.; Vollmann, A.; Huber, H.; Rose, D. Respiration of arsenate and selenate by hyperthermophilic archaea. Syst. Appl. Microbiol. 2000, 23, 305–314. [Google Scholar] [CrossRef]
- Sarret, G.; Avoscan, L.; Carrière, M.; Collins, R.; Geoffroy, N.; Carrot, F.; Covès, J.; Gouget, B. Chemical forms of selenium in the metal-resistant bacterium Ralstonia metallidurans CH34 exposed to selenite and selenate. Appl. Environ. Microbiol. 2005, 71, 2331–2337. [Google Scholar] [CrossRef] [PubMed]
- Oremland, R.S.; Hollibaugh, J.T.; Maest, A.S.; Presser, T.S.; Miller, L.G.; Culbertson, C.W. Selenate reduction to elemental selenium by anaerobic bacteria in sediments and culture: Biogeochemical significance of a novel, sulfate-independent respiration. Appl. Environ. Microbiol. 1989, 55, 2333–2343. [Google Scholar] [CrossRef]
- Handley, K.M.; Verberkmoes, N.C.; I Steefel, C.; Williams, K.H.; Sharon, I.; Miller, C.S.; Frischkorn, K.R.; Chourey, K.; Thomas, B.C.; Shah, M.B.; et al. Biostimulation induces syntrophic interactions that impact C, S and N cycling in a sediment microbial community. ISME J. 2013, 7, 800–816. [Google Scholar] [CrossRef]
- Nakamaru, Y.M.; Altansuvd, J. Speciation and bioavailability of selenium and antimony in non-flooded and wetland soils: A review. Chemosphere 2014, 111, 366–371. [Google Scholar] [CrossRef]
- Pettine, M.; Gennari, F.; Campanella, L.; Casentini, B.; Marani, D. The reduction of selenium (IV) by hydrogen sulfide in aqueous solutions. Geochim. Cosmochim. Acta 2012, 83, 37–47. [Google Scholar] [CrossRef]
- Yeager, C.M.; Amachi, S.; Grandbois, R.; Kaplan, D.I.; Xu, C.; Schwehr, K.A.; Santschi, P.H. Microbial Transformation of Iodine: From Radioisotopes to Iodine Deficiency. Adv. Appl. Microbiol. 2017. [Google Scholar] [CrossRef]
- Fernández-Calviño, D.; Bååth, E. Interaction between pH and Cu toxicity on fungal and bacterial performance in soil. Soil Biol. Biochem. 2016, 96, 20–29. [Google Scholar] [CrossRef]
- Starr, M.; Lindroos, A.J.; Ukonmaanaho, L.; Tarvainen, T.; Tanskanen, H. Weathering release of heavy metals from soil in comparison to deposition, litterfall and leaching fluxes in a remote, boreal coniferous forest. Appl. Geochem. 2003, 18, 607–613. [Google Scholar] [CrossRef]
- Moffett, B.F.; Nicholson, F.A.; Uwakwe, N.C.; Chambers, B.J.; Harris, J.A.; Hill, T.C. Zinc contamination decreases the bacterial diversity of agricultural soil. FEMS Microbiol. Ecol. 2003, 43, 13–19. [Google Scholar] [CrossRef]
- Kour, R.; Jain, D.; Bhojiya, A.A.; Sukhwal, A.; Sanadhya, S.; Saheewala, H.; Jat, G.; Singh, A.; Mohanty, S.R. Zinc biosorption, biochemical and molecular characterization of plant growth-promoting zinc-tolerant bacteria. 3 Biotech 2019, 9, 1–17. [Google Scholar] [CrossRef]
- Myers, B.; Webster, K.L.; Mclaughlin, J.W.; Basiliko, N. Microbial activity across a boreal peatland nutrient gradient: The role of fungi and bacteria. Wetl. Ecol. Manag. 2012, 20, 77–88. [Google Scholar] [CrossRef]
- Song, N.; Xu, H.; Yan, Z.; Yang, T.; Wang, C.; Jiang, H.L. Improved lignin degradation through distinct microbial community in subsurface sediments of one eutrophic lake. Renew. Energy 2019, 138, 861–869. [Google Scholar] [CrossRef]
- Lin, X.; Tfaily, M.M.; Steinweg, J.M.; Chanton, P.; Esson, K.; Yang, Z.K.; Chanton, J.P.; Cooper, W.T.; Schadt, C.W.; Kostka, J.E. Microbial community stratification linked to utilization of carbohydrates and phosphorus limitation in a boreal peatland at Marcell Experimental Forest, Minnesota, USA. Appl. Environ. Microbiol. 2014, 80, 3518–3530. [Google Scholar] [CrossRef] [PubMed]
- Coolen, M.J.; van de Giessen, J.; Zhu, E.Y.; Wuchter, C. Bioavailability of soil organic matter and microbial community dynamics upon permafrost thaw. Environ. Microbiol. 2011, 13, 2299–2314. [Google Scholar] [CrossRef]







| Sample | Bog Water pH | Bog Layer pH | Humification Degree | Organic Matter | Water Content | Number of Bacterial 16S rRNA Genes | S Content g/kg DW |
|---|---|---|---|---|---|---|---|
| Surface moss | 3.8 | 3.1 | H1 | 99.2 | 92.0 | 5.0 × 1010 | 1.7 |
| Peat 0.5 m | 4.7 | 3.1 | H3 | 99.5 | 88.8 | 5.0 × 1010 | 1.3 |
| Peat 1.5 m | ND | 3.1 | H4 | 99.6 | 90.9 | 1.0 × 1010 | 2.7 |
| Peat 2.5 m | 4.9 | 3.2 | H4 | 99.8 | 66.2 | 2.0 × 109 | 2.4 |
| Peat 3.5 m | ND | 3.2 | H4/H5 | 99.8 | 94.1 | 1.0 × 1010 | 2.4 |
| Peat 3.7 m | ND | 3.3 | H5 | 99.8 | 91.0 | 1.0 × 1010 | ND |
| Gyttja 5.5 m | 5.2 | 4.0 | H6 | 95.0 | 83.3 | 1.0 × 1010 | 1.2 |
| Clay 6.5 m | ND | 5.3 | Clay | 15.3 | 73.2 | 5.0 × 109 | 17.3 |
| Na NH4AC | K NH4AC | Ca NH4AC | Mg NH4AC | Al NH4AC | Sum (K+, Ca2+, Mg2+, Na+, Al3+) | ||
| Surface moss | 1.1 | 4.4 | 2.6 | 3.4 | 1.5 | 13 | |
| Peat 0.5 m | 0.2 | 0.3 | 1.0 | 3.0 | 1.1 | 6 | |
| Peat 1.5 m | 0.2 | 0.2 | 1.8 | 3.4 | 0.6 | 6 | |
| Peat 2.5 m | 0.3 | 0.4 | 2.5 | 4.3 | 0.5 | 8 | |
| Peat 3.5 m | 0.4 | 0.2 | 2.8 | 7.0 | 0.4 | 11 | |
| Peat 3.7 m | 0.3 | 0.2 | 5.6 | 9.1 | 0.5 | 16 | |
| Gyttja 5.5 m | 0.2 | 0.3 | 18.7 | 11.8 | 6.2 | 37 | |
| Clay 6.5 m | ND | ND | ND | ND | ND | ND | |
| Minerals in <2 µm fraction | Minerals in >2 µm fraction | ||||||
| Gyttja and Clay | Illite, clinochlore and kaolinite | Quartz, microcline, plagioclase, pyrite, Fe-hornblende | |||||
| Sample | Ni | Ag | Cs | Se | I | Se Anoxic * | I Anoxic * |
|---|---|---|---|---|---|---|---|
| Surface moss | nd | nd | 93 | 4388 | 1797 | nd | nd |
| Peat 0.5 m | 11,727 | 21,743 | 115 | 2602 | 478 | 15 | 226 |
| Peat 2.5 m | 7817 | 21,783 | 113 | 2716 | 80 | 18 | 194 |
| Gyttja 5.5 m | 19,880 | 17,778 | 221 | 696 | 316 | 6 | 1629 |
| Clay 6.5 m | 190 | 10,699 | 1601 | 735 | 24 | 0 | 333 |
| Surface | 0.5 m | 1.5 m | 2.5 m | 3.5 m | 5.5 m | 6.5 m | |
|---|---|---|---|---|---|---|---|
| Na | 433 | 0 | 0 | 0 | 202 | 0 | 430 |
| Mg | 634 | 687 | 581 | 831 | 2410 | 1520 | 6562 |
| Al | 273 | 350 | 221 | 197 | 650 | >1486 | >1486 |
| K | 4673 | 344 | 186 | 319 | 241 | 271 | 6156 |
| Ca | 1619 | 1193 | 955 | 938 | 2566 | 4030 | 4124 |
| Fe | 423 | 522 | 401 | 271 | 558 | 1509 | 25,543 |
| Co | 0.04 | 0.03 | 0.04 | 0.02 | 0.02 | 1.01 | 11 |
| Ni | 0 | 20 | 15 | 0 | 0 | 0 | 34 |
| Cu | 13 | 5.01 | 3.02 | 0 | 1.04 | 25 | 35 |
| Zn | 48 | 37 | 23 | 19 | 21 | 23 | 90 |
| Se | 0 | 0 | 0 | 0 | 0 | 0 | 6.05 |
| Mo | 0.03 | 4.00 | 4.00 | 0 | 0 | 0.09 | 6.02 |
| Ag | 0.02 | 0.02 | 0.01 | 0.02 | 0.01 | 0.01 | 0.02 |
| Cs | 0.05 | 0.01 | 0.01 | 0 | 0.01 | 0.01 | 3.03 |
| Th | 0.01 | 0.01 | 0.01 | 0 | 0.01 | 1.03 | 7.06 |
| U | 0 | 0 | 0 | 0 | 0 | 1.01 | 4.04 |
| Sample | Sequences | OTUs | Chao 1 | Coverage (%) | Shannon H′ |
|---|---|---|---|---|---|
| Surface A | 1228 | 283 | 505 | 88 | 4.7 |
| Surface B | 1215 | 302 | 520 | 87 | 4.8 |
| Peat 0.5 m A | 12,587 | 145 | 311 | 99 | 1.7 |
| Peat 0.5 m B | 13,314 | 148 | 285 | 99 | 1.6 |
| Peat 1.5 m A | 11,034 | 128 | 204 | 99 | 2.2 |
| Peat 1.5 m B | 11,470 | 141 | 252 | 99 | 2.1 |
| Peat 2.5 m A | 2212 | 100 | 159 | 98 | 2.8 |
| Peat 2.5 m B | 5971 | 160 | 273 | 99 | 2.8 |
| Peat 3.5 m A | 9871 | 170 | 411 | 99 | 2.3 |
| Peat 3.5 m B | 11,672 | 129 | 263 | 99 | 2.1 |
| Peat 3.7 m A | 13,120 | 177 | 445 | 99 | 2.0 |
| Peat 3.7 m B | 4354 | 80 | 143 | 99 | 2.0 |
| Gyttja 5.5 m A | 23,698 | 280 | 493 | 99 | 2.4 |
| Gyttja 5.5 m B | 19,757 | 200 | 325 | 100 | 2.3 |
| Clay 6.5 m A | 18,925 | 200 | 314 | 100 | 1.9 |
| Clay 6.5 m B | 14,913 | 195 | 293 | 99 | 2.1 |
| Sample | Sequences | OTUs | Chao 1 | Coverage (%) | Shannon H′ |
|---|---|---|---|---|---|
| Surface A | 7067 | 323 | 418 | 99 | 3.8 |
| Surface B | 4495 | 237 | 343 | 98 | 3.7 |
| Peat 0.5 m A | 20,265 | 308 | 463 | 99 | 3.8 |
| Peat 0.5 m B | 17,543 | 286 | 379 | 99 | 3.1 |
| Peat 1.5 m A | 18,224 | 291 | 406 | 99 | 3.7 |
| Peat 1.5 m B | 17,433 | 209 | 307 | 100 | 3.1 |
| Peat 2.5 m A | 5689 | 120 | 171 | 99 | 2.7 |
| Peat 2.5 m B | 10,079 | 206 | 307 | 99 | 2.9 |
| Peat 3.5 m A | 7023 | 156 | 307 | 99 | 2.9 |
| Peat 3.5 m B | 8419 | 159 | 240 | 99 | 2.6 |
| Peat 3.7 m A | 14,233 | 327 | 465 | 99 | 3.4 |
| Peat 3.7 m B | 17,236 | 218 | 287 | 100 | 2.2 |
| Gyttja 5.5 m A | 13,782 | 259 | 445 | 99 | 2.9 |
| Gyttja 5.5 m B | 18,917 | 246 | 350 | 100 | 3.3 |
| Clay 6.5 m A | 13,924 | 241 | 388 | 99 | 3.2 |
| Clay 6.5 m B | 17,840 | 310 | 433 | 99 | 3.1 |
| Sample | Sequences | OTUs | Chao 1 | Coverage (%) | Shannon H′ |
|---|---|---|---|---|---|
| Surface A | 3089 | 654 | 1348 | 88 | 5.5 |
| Surface B | 7988 | 1063 | 2015 | 93 | 5.5 |
| Peat 0.5 m A | 2961 | 546 | 1365 | 88 | 4.6 |
| Peat 0.5 m B | 9295 | 697 | 1422 | 96 | 4.1 |
| Peat 1.5 m A | 36,102 | 1122 | 2420 | 98 | 3.3 |
| Peat 1.5 m B | 8723 | 321 | 603 | 98 | 2.9 |
| Peat 2.5 m A | 7937 | 382 | 812 | 97 | 3.6 |
| Peat 2.5 m B | 15,327 | 634 | 1376 | 98 | 3.7 |
| Peat 3.5 m * | 3841 | 191 | 385 | 97 | 3.4 |
| Peat 3.7 m * | 5256 | 261 | 410 | 98 | 2.9 |
| Gyttja 5.5 m A | 5992 | 556 | 1196 | 95 | 4.4 |
| Gyttja 5.5 m B | 9743 | 731 | 1393 | 96 | 4.3 |
| Clay 6.5 m A | 2811 | 575 | 1502 | 87 | 5.1 |
| Clay 6.5 m B | 5002 | 633 | 1142 | 93 | 4.7 |
| Ag Sorption | Cs Sorption | Se Sorption | Se Anoxic Sorption | I Anoxic Sorption | |
|---|---|---|---|---|---|
| Lokiarchaeia genus | −0.98 | ||||
| Methanosaeta | −0.98 | ||||
| Hadarchaeales genus | 0.98 | ||||
| Acetothermiia_ge | 1.0 | ||||
| GOUTB8_ge | 1.0 | ||||
| Thermoanaerobaculum | 1.0 | ||||
| Pelolinea | 1.0 | ||||
| Anaerolineaceae;uncultured | 0.99 | ||||
| Latescibacteraceae_unclassified | 1.0 | ||||
| MidBa8_ge | 1.0 | ||||
| DG-20_ge | 1.0 | ||||
| Bacterial number | 0.99 *** | 0.96 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lusa, M.; Bomberg, M. Microbial Community Composition Correlates with Metal Sorption in an Ombrotrophic Boreal Bog: Implications for Radionuclide Retention. Soil Syst. 2021, 5, 19. https://doi.org/10.3390/soilsystems5010019
Lusa M, Bomberg M. Microbial Community Composition Correlates with Metal Sorption in an Ombrotrophic Boreal Bog: Implications for Radionuclide Retention. Soil Systems. 2021; 5(1):19. https://doi.org/10.3390/soilsystems5010019
Chicago/Turabian StyleLusa, Merja, and Malin Bomberg. 2021. "Microbial Community Composition Correlates with Metal Sorption in an Ombrotrophic Boreal Bog: Implications for Radionuclide Retention" Soil Systems 5, no. 1: 19. https://doi.org/10.3390/soilsystems5010019
APA StyleLusa, M., & Bomberg, M. (2021). Microbial Community Composition Correlates with Metal Sorption in an Ombrotrophic Boreal Bog: Implications for Radionuclide Retention. Soil Systems, 5(1), 19. https://doi.org/10.3390/soilsystems5010019